

Age and growth of the endangered Maugean skate (Zearaja maugeana) by using microchemical analysis
Claire van Werven A * , David Moreno A , Sean Tracey A and Jeremy Lyle A
A * , David Moreno A , Sean Tracey A and Jeremy Lyle A
A
Abstract
The Maugean skate (Zearaja maugeana) is an endangered species micro-endemic to two small embayments in western Tasmania, Australia. Given the species’ range restriction, status and exposure to large environmental variability, emerging impacts from anthropogenic pressures are likely to cause additional stress to the population. Information on Maugean skate age and growth is limited.
To determine the age and estimate growth parameters of the Maugean skate using vertebral banding patterns supplemented with microchemical analysis.
This study estimated the ages of opportunistically sampled Maugean skate (n = 44) from Macquarie Harbour between 2012 and 2019, by using incremental banding in their vertebrae. A multi-model inference framework using Akaike’s information criterion corrected for small sample size was used to determine the best fit growth function base on length at age data. Annual periodicity of growth bands was indirectly verified using laser ablation–inductively coupled plasma–mass spectrometry (LA-ICP-MS) measuring the uptake of heavy metals in the vertebrae.
Age estimates ranged from 2 to 10 years. A two-parameter von Bertalanffy growth function provided the best fit when applied with a fixed length at birth on the basis of two neonate individuals (L0 fixed at 103.50-mm total length, TL), with the parameters as follows: the theoretical maximum length (L∞) and growth coefficient (k) estimated at 843.53 mm TL and 0.27 for females, 747.47 mm TL and 0.32 for males and 836.34 mm TL and 0.25 for pooled data. Females grew faster and reached a larger size than did males.
The Maugean skate’s reasonably fast growth, early maturity and short lifespan may help it withstand disturbances in the dynamic harbour. However, density-dependent responses to population declines could limit its ability to regulate numbers, especially amid ongoing threats.
The use of archived samples of vertebrae indirectly verified with microchemistry demonstrates a novel technique to describe the age and growth of Maugean skate, and provides critical life-history parameters to contribute to population assessments of this endangered population. The need to carry out age-validation studies for this species is also highlighted.
Keywords: age verification, chondrichthyans, elasmobranchs, laser ablation–inductively coupled plasma–mass spectrometry, Macquarie Harbour, Maugean skate, multi-model inference, schlerochronology, vertebral microchemistry.
Introduction
Skates (superorder: Batoidea; family: Rajidae) are a diverse group of cartilaginous fish comprising over one-quarter of all extant chondrichthyans (sharks, skates, rays and chimeras) (Bizzarro et al. 2014). Skate typically display k-selected life-histories, distinguished by low fecundity, late maturity and slow growth; however, relative to other chondrichthyans such as sharks, they tend to exhibit a high reproductive rate (Simpfendorfer and Kyne 2009). Nevertheless, skate populations have undergone significant worldwide declines over the past few decades (Dulvy et al. 2000). Although the magnitude of population declines is subject to continued discussion, overexploitation, habitat modification and various other anthropogenic pressures have been recognised as primary reasons for these losses, resulting in the classification of some as threatened (Siskey et al. 2019). Despite their high diversity and far-reaching global distribution (Griffiths et al. 2010), the development of fishery or conservation management plans for skates is difficult. This is due, in part, to the lack of detailed biological information and species-specific data (Stevens et al. 2000), given that many skate species are of low economic value or, as is the case for the Maugean skate (Zearaja maugeana) a result of its endangered status, where low abundances and ethical limitations may impede research efforts.
A commonly used technique for estimating age and growth in chondrichthyans involves the analysis of band pairs in vertebral centra (hereafter ‘vertebrae’). As chondrichthyans grow, vertebrae develop ‘bands’, where alternating opaque and translucent rings are deposited during periods of slow (winter) and faster (summer) growth (Goldman 2005). To estimate age, rings within vertebral cross-sections are counted and an age is estimated in years (Cailliet 2015). For many species, a band pair occurs annually at a rate that parallels growth, where one band pair represents 1 year and, hence, growth is assumed to be annual (Cailliet et al. 2006). For some chondrichthyan species such as the Pacific angel shark (Squatina californica) bands are not deposited annually, but instead bi-annually, with some studies demonstrating band pair deposition to cease completely in older fish (Francis et al. 2007).
The verification or validation for these bands across a sample group is therefore important to ensure that the number of ‘growth’ bands reflects the accurate age of the individual. Although verification techniques generally require expansive sample sizes to examine seasonal or environmental trends in band formation, validation techniques require physical calcified structures to be marked and studied either in captivity, or released and recaptured at a later point (e.g. oxytetracycline, OTC, or marginal increment analysis, MIA; Cailliet et al. 2006).
The growth rate and recovery potential of diminishing populations of skates have typically involved demographic analyses based on estimates of age to determine important life-history characteristics (e.g. age at sexual maturity; Cortés et al. 2023). Nevertheless, the paucity of reliable age estimates is often a limitation in such analyses, notably threatened species’ life-history information.
Given that these traditional validation techniques are either logistically unachievable or costly for many species, microchemical fingerprinting has emerged as an alternative approach to age validation in chondrichthyans, given that vertebrae grow concentrically over time (Tillett et al. 2011). Once incorporated, the hard tissue is assumed to remain metabolically inert and it is unlikely that the elemental composition is altered following deposition, therefore giving rise to the potential of these individuals to carry chronological records of environmental history within their tissues (Scharer et al. 2012). Elements that are present in an individual’s environment are known to be absorbed into the vertebral hydroxyapatite matrix during the biomineralisation process (Outridge et al. 1995). Although studies validating elemental uptake pathways into vertebrae are currently limited, concentrations of certain elements are thought to be associated with environmental variables such as salinity (e.g. strontium, Sr; and barium, Ba), temperature (e.g. Ba), dietary intake (e.g. zinc, Zn; and manganese, Mn) and reproductive cues (e.g. Zn) (Tillett et al. 2011; Raoult et al. 2018). As a result, the chemical analysis of vertebrae may provide a retrospective tool to show changes in the environmental conditions experienced throughout an individual’s life (Scharer et al. 2012). Taking advantage of this theory, microchemical analysis of vertebrae using laser ablation–inductively coupled plasma–mass spectrometry (LA-ICP-MS) has been used to verify the periodicity of the growth band formation in vertebrae, as changes in specific elemental concentrations can be related with growth band deposition or seasons (Hale et al. 2006; Mohan et al. 2018). This technique is particularly useful for endangered species, for which samples are often rare (Scharer et al. 2012), or for species with few or difficult to discern growth bands.
Occupying one of the most restricted distributions of any chondrichthyan, the Maugean skate inhabits a unique ecosystem in western Tasmania (Australia) (Moreno et al. 2022). Maugean skate reaches a maximum size of 840 mm in total length (TL) and has been described as a relic species from the late Cretaceous period (80 million–100 million years ago) when Gondwana fragmented (Last and Gledhill 2007). On the basis of its limited geographic range and assumed small population size, Maugean skate has been listed as Endangered under the Threatened Species Protection Act 1995 (Tas.) (Threatened Species Link 2020), the Environmental Protection and Biodiversity Conservation Act 1999 (Cth), as well as the IUCN Red List of Threatened Species (Last et al. 2016). Primarily an estuarine species, Maugean skate is known to inhabit only Macquarie Harbour (western Tasmania) (Treloar et al. 2017), Australia (Fig. 1). The species had been identified in Bathurst Harbour; however, the last recorded sighting of the species in this harbour was in 1992 (Bell et al. 2016). Recent eDNA work has presented clear evidence that the population in this harbour is either extremely small or locally extinct (Moreno et al. 2022). Hence, it is likely that Macquarie Harbour now represents the last remaining habitat for this species (Bell et al. 2016).
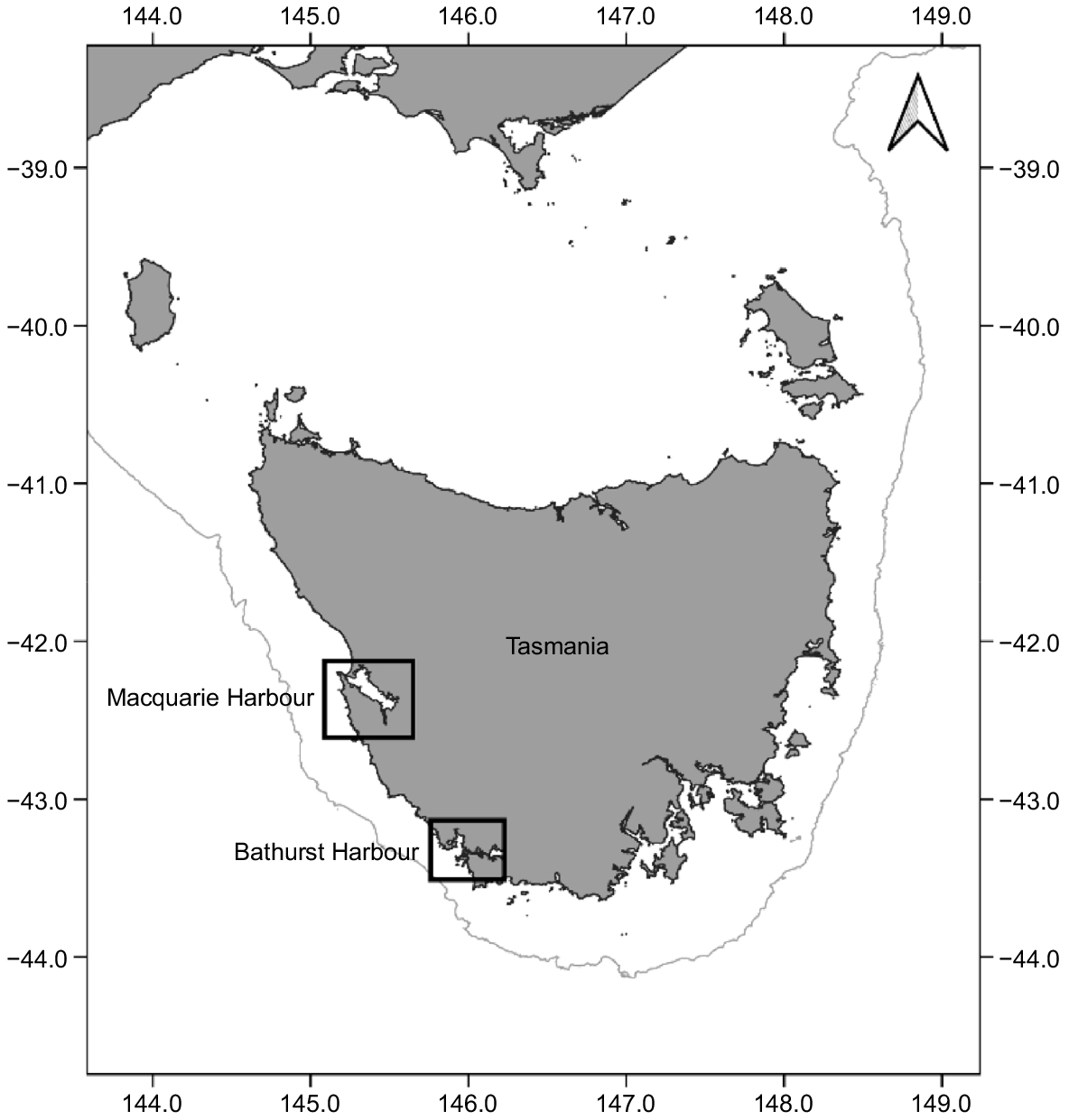
Macquarie Harbour’s ria embayments are highly stratified, with the predominantly freshwater surface layer heavily tannined, significantly hindering light penetration below the surface (Ross and MacLeod 2018). Within Macquarie Harbour, the stratified brackish middle layer exhibits little thermal variation and reduced dissolved oxygen (DO), whereas the bottom layer is nearly completely saline (31 g kg−1), with limited oceanic mixing (Ross and MacLeod 2017). The modification and pollution of rivers entering Macquarie Harbour through damming and historical mining, in addition to the development of salmonid aquaculture and climate-change impacts, may alter the physical and chemical environment of Macquarie Harbour, with potential ramifications for the population (Moreno et al. 2020). A considerable threat to the future wellbeing of Maugean skate in Macquarie Harbour also stems from recent observations of water health, where dramatic declines in deep water dissolved oxygen (DO) concentrations have been recorded and are likely affecting the already narrow depth range (5–15 m) of Maugean skate (Bell et al. 2016). Given the study site (Macquarie Harbour) experiences very little oceanic mixing because of its narrow and shallow entrance, this distinct bio-geochemical environment may be useful as a microchemical fingerprint.
Underlining the need for comprehensive life-history data on Maugean skate, this study aims to (1) estimate the age of Maugean skate through the reading of growth band-pair counts deposited on vertebrae, (2) investigate the utility of LA-ICP-MS as a technique for verifying the periodicity of these growth bands with visually aged vertebrae counts, and (3) compare and obtain best growth models to inform key life-history parameters for this species. Results from this study are intended to augment life-history and demographic knowledge of Maugean skate for improved assessment of the endangered population in Macquarie Harbour (Awruch et al. 2021).
Materials and methods
Sample collection
Vertebrae from Maugean skate were opportunistically sourced from Macquarie Harbour between 2012 and 2019 (Fig. 1), with 11 specimens collected in 2012 (Lyle et al. 2014), 13 collected in 2014 (Bell et al. 2016), and the remaining 20 collected in 2019 (Morash et al. 2020; Moreno et al. 2020). The total number of samples available was 44. Most of the Maugean skates were captured by monofilament gill nets deployed in depths ranging from 1 to 19 m, with the exception of three illegally discarded individuals found deceased at the local boat ramp. Additional details of capture methods, site location and biological sampling methodologies are detailed in Bell et al. (2016), Treloar et al. (2017) and Moreno et al. (2020).
Ethics and permits
All procedures were undertaken with University of Tasmania Animal Ethics Committee approval (permit A13468 and permit A16526) and scientific permits (13125, 14139, 17083, 17197 and 19026) issued under Section 14 of the Living Marine Resources Management Act 1995 and permits (TFA 13982, 14019, 14253, 17107 and 17359) issued under Regulation 4 of the Threatened Species Protection Regulations 2006 and Section 29 of the Nature Conservation Act 2002. These latter permits covered the capture, possession, tagging and biological sampling of an endangered species, the deployment of research fishing gear and the deployment of moorings within Macquarie Harbour, including the World Heritage Area.
Sample processing
Seeing as the location in the vertebral column from which samples are taken for ageing can have a statistically significant effect on increment counts, this study ensured that the vertebral sampling region was standardised to use only the thoracic centra (Officer et al. 1996). Five individual vertebrae from this region in each sample were cleaned of excess tissue, and neural and haemal arches were removed using a scalpel blade, leaving the centra for further preparation. Vertebrae were then immersed in 50% bleach solution (sodium hypochlorite 42 g L−1; sodium hydroxide 9 g L−1) for 5 min, with additional time (between 10 and 20 min) allocated to larger vertebrae to ensure that any remaining soft tissue was removed. Vertebrae were then immediately placed into individual labelled tissue cassettes submerged in a running water bath, left to rinse for 30 min and then air dried under a covered fume hood for 24–48 h. Dry vertebrae were embedded into a two-part casting polyester resin block and, once cured, sectioned along the sagittal plane by using an Isomet low-speed diamond-bladed saw (Isomet; Buehler, Lake Bluff, IL, USA) at ~240 rpm with a 75-g load weight. Sections were ~500 μm thick and were rinsed immediately after cutting in deionised water to remove potential contaminants (Moreno Iturria 2018).
Sections were sonicated for 10 min in distilled water and allowed to air dry in a laminar flow-filtered fume hood. Sections reserved for microchemistry were further sonicated three times. The dry samples used for age determination were mounted using two-part resin in a randomised order onto glass microscope slides and stored in covered containers. The sections used for microchemistry were mounted to microscope slides using double-sided tape and kept in a microscope slide storage box to reduce contamination risk.
Age determination
Vertebrae mounted to slides were photographed with a digital camera-mounted stereomicroscope by using transmitted light (Leica MZ7.5). The software package ‘Image J’ (W. S Rasband, US National Institutes of Health, Bethesda, MD, USA, see https://imagej.nih.gov/ij/; Schindelin et al. 2012) was used for image enhancement and annotation for ageing. Banding was visible along the corpus calcareum reading from the focus to the outer centrum edge (Fig. 2). The change of angle caused by differences in growth rate from intra-uterine to post-natal life-history stages was recorded as the point of birth (or birth mark) and assumed to represent year zero. Two full-term neonate skates (112 and 92 mm) were used to validate the position of the birth increment in vertebrae (~0.5 mm) and size at birth, used for models requiring a fixed L0 parameter (103.50 mm TL). Multiple counts were read on each ‘horn’ of vertebra and they did not vary so the most readable section was analysed.
A vertebral centra section from a 617-mm total length (TL) male Maugean skate (Zearaja maugeana) estimated to be 4 years of age. The white dots indicate translucent bands. Laser ablation–inductively coupled plasma–mass spectrometry (LA-ICP-MS) ablation path is depicted by a yellow line transect.
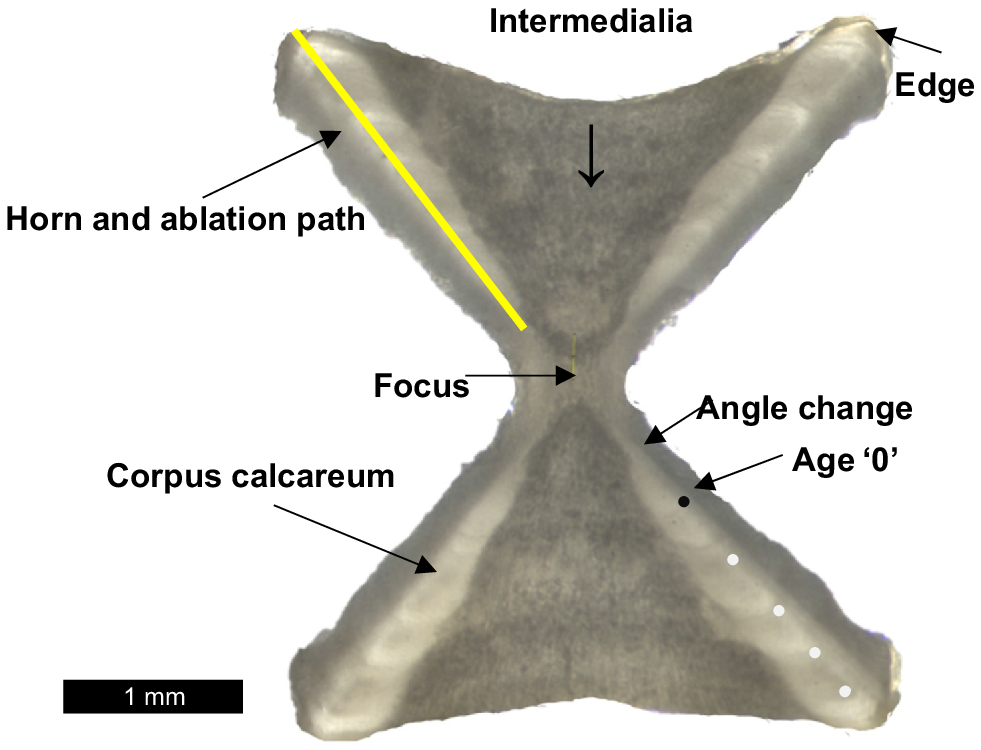
The centrum radius at time of capture (CRc) of each vertebra from each individual (defined as the distance from the focus of centra to the edge along the corpus calcareum) was measured to the nearest micrometre (Fig. 2). The relationship between CRc and TL was explored. Linear and quadratic models were fitted to males and females separately, model fit was compared using ANOVA, and the adjusted coefficient of determination was used to evaluate fit.
Age estimates were recorded without any knowledge of the size and sex of the individual. Each vertebra was read multiple times by two different readers (Reader A, first author; and Reader B, second author). When estimating age, a subjective readability index (RI) (between 1 and 6) was assigned to each vertebra, to allow a confidence factor to provide quantitative comparisons of perceived readability between readers (Officer et al. 1996). Reader A performed a practice read and then two non-consecutive reads on the samples to ensure accuracy, indicating the readability of each vertebra. Reader B performed one read on the samples and indicated readability. The latter read of Reader A was compared with the read of Reader B, and any vertebrae with discrepancies were re-read a third time by Reader A to determine a consensus.
Precision
Precision, defined as the reproducibility of repeated measurements on a given structure, was estimated through average percentage error (APE) (Beamish and Fournier 1981), as follows:
where n is the number of skate aged, r is the number of readings, xij is the ith age estimation of the jth skate at the ith reading, and xj is the mean age calculated for the jth skate.
Both intra- and inter-reader precision, in addition to comparison of visual age count and microchemical peak count, were compared using average percentage error (APE), with uncertainty surrounding the estimate calculated using a bootstrapping procedure (n = 500). Systematic bias was analysed by comparing the bootstrapped mean APE to a bias-corrected mean APE, and through a bias plot (Beamish and Fournier 1981). Internal error was calculated to understand bias between the visual band counts and the different elements.
Growth modelling
A multi-model inference (MMI) (Katsanevakis and Maravelias 2008) approach was applied, with six candidate growth models being fitted to the observed age and length data for males and females separately, and pooled. These models were fitted because they are commonly used to model growth in elasmobranch studies (Liu et al. 2021): (1) the von Bertalanffy growth model (VBGM), (2) the two-parameter reformulation of the VBGM with a fixed Lo (two-parameter VBGM), (3) the Gompertz growth model (GGM), (4) a logistic model (LM), (5) the hyper-k two-phase VBGM (TPVB hyper-k) and (6) the log-L∞ VBGM (TPVB log-L∞). The fixed-value L0 for both sexes was 103.50 mm TL on the basis of mean size at birth of two skates hatched in captivity in a previous study (Moreno et al. 2020). The biphasic models (i.e. hyper-k and log-L∞) were initially fitted using a model-estimated inflection point; however, no convergence was reached using this full parametrisation. As a result, fixed values were applied on the basis of observed size at maturity data from a previous report; females (t50 = 662 mm TL), males (t50 = 632 mm TL) and the average of these to generate the pooled value (t50 = 647 mm TL) (Bell et al. 2016). Confidence intervals (95% CI) for the estimated parameters were calculated by applying a bootstrap procedure (n = 1000).
Model selection
As per Moreno Iturria (2018), models were evaluated using a MMI theoretic approach on the basis of a small sample-corrected Akaike’s information criterion (AICc) (Katsanevakis and Maravelias 2008). AICc modifies the standard AIC (Akaike 1973) by considering the sample size and number of variables and was selected over AIC because of its improved performance when sample sizes are <200 (Zhu et al. 2009). The model with the lowest AICc value (AICmin) was considered the most parsimonious, all candidate models were ranked on the basis of the AICc differences:
AICc was estimated as follows:
where for least squares:
where σ2 = RSS ÷ n, n is the sample size and Ø is the total number of regression parameters estimated including the variance (σ2).
Normal deviations with constant variance were assumed. AIC weights (wi) for each model represent the probability of choosing the correct model from the set of candidates (Harry et al. 2011). The Akaike weight (wi) acts as a Bayesian a posterior model probability for the candidate models representing the true fit for a given dataset. The threshold for selecting single ‘best’ model fit for the data was wi ≥ 0.9 (Katsanevakis and Maravelias 2008). Akaike weights were calculated as follows:
The 95% confidence intervals (CI) and prediction intervals were calculated around the best-fit parameter estimates. All six models were parameterised to include a length-at-birth parameter (L0) and an asymptotic length parameter (L∞) because both can be compared directly between growth functions. Model-averaged estimates of the coefficients L0 and L∞ were estimated as a weighted average, by applying Akaike weights as a scaling factor, noting that the two-parameter VBGM (M2) had a fixed value for L0 (Katsanevakis and Maravelias 2008).
The growth coefficient of the different models (k) was not directly comparable, so the derivative form of each model was used to calculate instantaneous growth rates at birth (dL/dt0), age at 50% maturity (dL/dt50) and age at 95% of the asymptotic length (dL/dtL∞, c), as per Thorson and Simpfendorfer (2009).
A likelihood-ratio test (LRT) was conducted on the best-fitting candidate model by using Microsoft Excel to determine whether there was a significant difference in the growth functions between sexes, using the method described by Haddon (2011).
Longevity
Two estimates of longevity were used for the species, the first being the initial values of life span (tmax = observed) for both sexes obtained from the oldest vertebral age estimate in this study. A theoretical longevity estimate was also calculated according to the equation proposed by Fabens (1965), which is defined by the time it takes to reach 95% of L∞ and is denoted as follows:
where k is a coefficient of the von Bertalanffy growth curve.
Elemental analysis
Analytical measurements were taken using a NewWave NWR213-ESI laser system coupled to a quadruple ICP–MS housed at the Centre for Ore Deposits and Earth Sciences (CODES) analytical laboratory (Moreno Iturria 2018). Each sample was pre-ablated over the complete sampling transect with the laser to remove any residual surface contamination. To correct for background signals, a 28-s blank preceded all analysis. The laser was operated with a pulse frequency of 5 Hz, a scan speed of 10 μm s−1, and an energy output of 0.2–0.3 mJ pulse−1. The spot ablation size was 76 μm. Parameters were kept consistent between runs. The standards NIST612, BCR-2G, GSD-1G and Durango were measured at regular intervals throughout the analytical run to account for instrumental drift. The limit of detection (Morrison et al. 2019) was calculated following Longerich et al. (1996), and elements that did not meet this standard were excluded from the analysis. The suite of elements analysed (see Supplementary Table S1) was selected because they have previously been used as chemical tracers for determining movement and habitat, or, alternatively, if they have provided results for previous attempts at age verification of two morphologically similar sharks (Tillett et al. 2011). Transects were drawn along the sagittal plane of the vertebrae, beginning at the focus through to the centre of the corpus calcareum (Fig. 2). The transect was programmed to reach the edge of the vertebrae into the resin mount to ensure that the most recent material was analysed and therefore the last known point of location before capture of the skate. To compare inter-sample variation, multiple edges of the same vertebrae were ablated. Data reduction was performed using the standalone software package LADR (Norris Scientific). The average background levels in the pre-analysis blank were subtracted from the data, and calcium (43Ca) was used as an internal standard to transform count data into parts per million and subsequently to molar masses (Tillett et al. 2011). All statistical analyses were undertaken in R (ver. 3.2.3, R Foundation for Statistical Computing, Vienna, Austria, see https://www.r-project.org/) at a critical probability level of 0.05.
Elemental age verification
Element concentrations were fitted with a 5-point running average to each transect to smooth the data and reduce instrumental noise. Butterfly transects (mirrored plot profiles) were analysed to ensure that elemental concentrations were consistent across both sides of the vertebrae. This also corroborates the assumption that elements are present proportional to initial incorporation. The degree to which smoothed peaks in elemental profiles corresponded to opaque bands was evaluated and overlaying the position of opaque bands on plots obtained by measuring the distance (mm) from the focus to the mid-point of each opaque band of the vertebra. The first discernible peak was assumed the birth band (Year 0); thus, every consecutive peak following was an additional ‘count’.
Results
Size distribution
Vertebrae from 44 Maugean skate individuals collected in western Tasmania between 2012 and 2019 were assessed in this study. The size range of individuals varied from 450 to 819 mm TL for females (n = 27; mean ± s.e. = 709 ± 17.40 mm TL) and from 520 to 720 mm TL for males (n = 17; mean ± s.e = 644 ± 13.61 mm TL). The length distribution of females was significantly different from that of males (Kolmogorov–Smirnov test; D = 0.556, P < 0.05).
Precision
Across the two independent readers, no samples were excluded from the lack of consensus and there was no systematic bias of age estimates detected. Precisions for intra- and inter-reader variation were 3.17 and 8.56% respectively, as determined by mean APE. Inter-reader differences in estimated visual counts from vertebrae were higher than were intra-reader differences; however, neither was significant (P < 0.05). For both intra- and inter-reader estimates, ages were more difficult to discern in older individuals, with less certainty for estimates between 8 and 10. Both APE were therefore within the range (<10%) of what is commonly considered acceptable in ageing studies. The APE between the observed counts and those from elemental peaks were 27.41% for Sr:Ba, 15.35% for uranium, 10.47% for manganese and 6.9% for zinc.
Age determination
Centrum vertebral radius at capture (CRc) was positively correlated with length for both sexes, with a quadratic function providing a better fit in females, males and pooled data (P < 0.05).
The oldest individual was estimated to be 10 years for both females and males, with the youngest age for females and males being estimated to be 2 and 4 years respectively.
Growth analysis
Female Maugean skate grew to a larger size than males (Fig. 3). For female, male and pooled sexes, all growth models reached convergence, with the two-parameter VBGM (fixed L0) (M2) displaying the lowest AICc (females = 308.33, males = 192.48) and highest wi, (females = 58.16, males = 62.16) regardless of sex (Tables S2–S4). This model (M2) was considered the best for describing Maugean skate growth, with L0 fixed at 103.50 mm TL and L∞ and k estimated at 843.53 mm TL and 0.27, for females, 747.47 mm TL and 0.32 for males and 836.34 mm TL and 0.25 for pooled data. A LRT identified a significant difference between female and male (M2) growth curves (χ2 = 9.11, d.f. = 2, P < 0.05), and hence subsequent results have presented sex-specific life-history parameters for management.
Estimated growth for observed data on the basis of vertebral age estimates of (a) female, (b) male and (c) pooled female and male Maugean skates (Zearaja maugeana). Model shown is the best-ranked for each dataset (solid line) 95% confidence intervals (dashed line) and prediction intervals (dotted line). The best-ranked model regardless of sex was the two-parameter VBGM (fixed L0 = 103.50 mm TL) (M2).
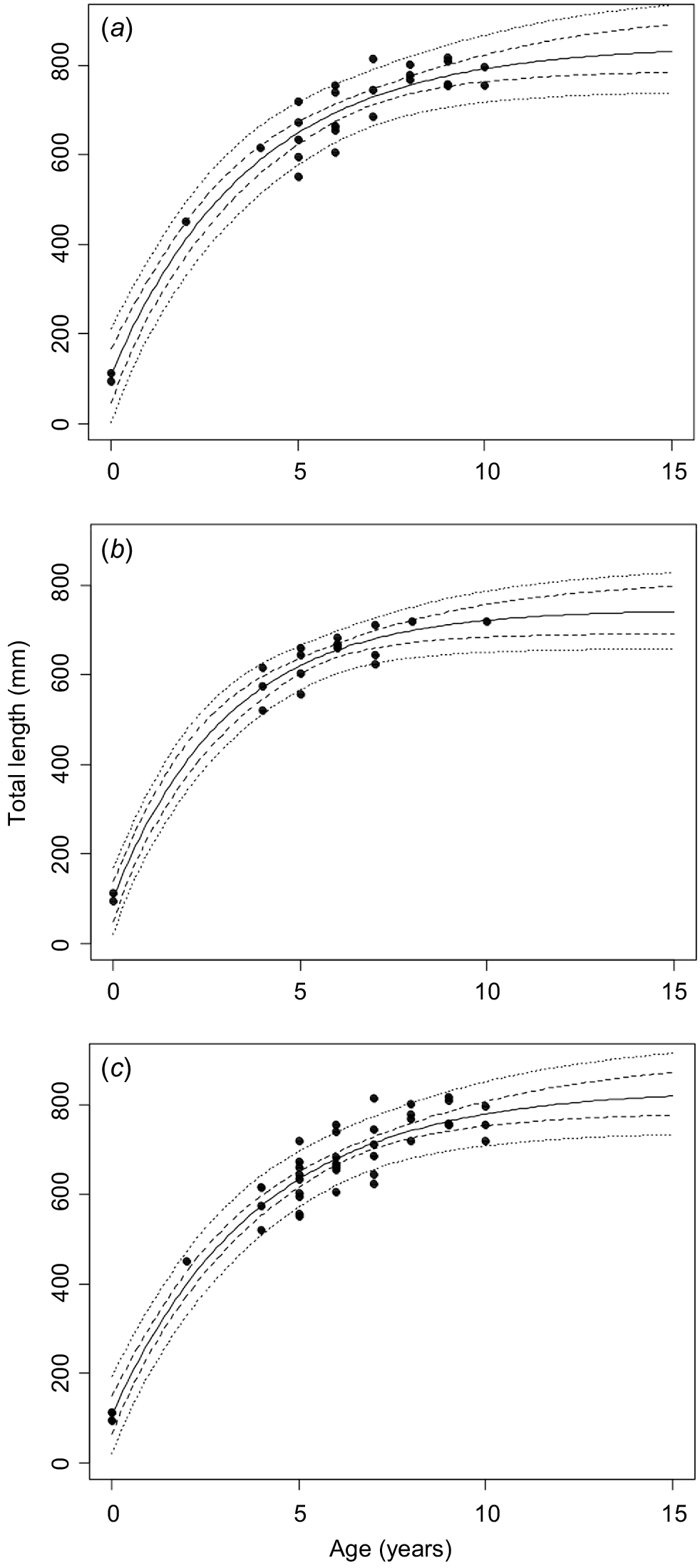
The selection of this candidate model was supported by the robust performance of the model across Akaike weights (wi), with values of >50% (Burnham and Anderson 2002) (Supplementary Tables S2–S4). In the absence of a clear superior candidate model, the model-averaged parameters provided a good compromise by incorporating uncertainty, and in data-poor situations, as was encountered in this study, should be selected over the single-model estimates when informing demographic analysis or management (Moreno Iturria 2018). However, no single model resulted in wi of >90%, so no single best model could be selected. Weighted model-averaged parameters were calculated to help characterise modelling uncertainty. The weakest of all candidate models (wi < 3%) was the Logistic (M4) for females and pooled data, and the two-phase hyper-k (M5) model. The Logistic, two-parameter VBGM (fixed L0) and model-averaged parameters had the lowest errors regardless of sex or dataset. Theoretic estimates of longevity were much greater than was the observed maximum age (Table 1).
| Species | Age and growth | Longevity | Source | |||||
|---|---|---|---|---|---|---|---|---|
| t0 (year−1) | L0 (mm) | L∞ (mm) | k (year−1) | tmax (year−1) | tmaxth (year−1) | |||
| Z. chilensis | −0.51 | 1283 | 0.11 | 27 | 31 | Licandeo et al. (2006) | ||
| Z. nasutus | −1.20 | 913 | 0.16 | 9 | 22 | Francis et al. (2001) | ||
| Z. maugeana | −2.78 | 329.03 | 916 | 0.16 | 10 | 22 | Bell et al. (2016) | |
| Z. maugeana | −0.57 | 107.75 | 835 | 0.26 | 10 | 13 | This study (M1) | |
| Z. maugeana | −3.00 | 355.71 | 878 | 0.17 | 10 | 17 | This study based on Zn ages (M1) | |
Derived longevities, tmaxth (Fabens 1965), were solved using known parameters based on this study’s two definitions of longevity.
Elemental analysis
Each element assayed was consistently above detection limits for 43 Maugean skate microchemistry samples. One sample (MS43) was removed from analysis because of evidence of surface contamination. Of the elements analysed, zinc (66Zn), uranium (238U) and manganese (55Mn) were used for the analysis, given they provided the clearest patterns (Fig. 4). Oscillations in the relative signal intensity of these elements were observed along the transect that closely correlated with the identified growth bands (Fig. 4).
Photograph of the (a) corpus calcareum of a vertebral section (size of section is 3.13 mm) from a mature Maugean skate (Zearaja maugeana) (MS34) aged 9 years, together with the corresponding 5-point moving average smoothed profiles of (b) 66Zn:43Ca, (c) 55Mn:43Ca and (d) 238U:43Ca along vertebra transect distance (mm). All elemental concentrations are relative to 43Ca and are presented per millimole or per micromole. For scale, section is from the top left-hand horn of the vertebrae as depicted in Fig. 2 of this document, encompassing the area from the focus to the vertebral edge. Grey shading aligned on plots depicts visual opaque bands from section (a). Birth bump is indicated by yellow shading.
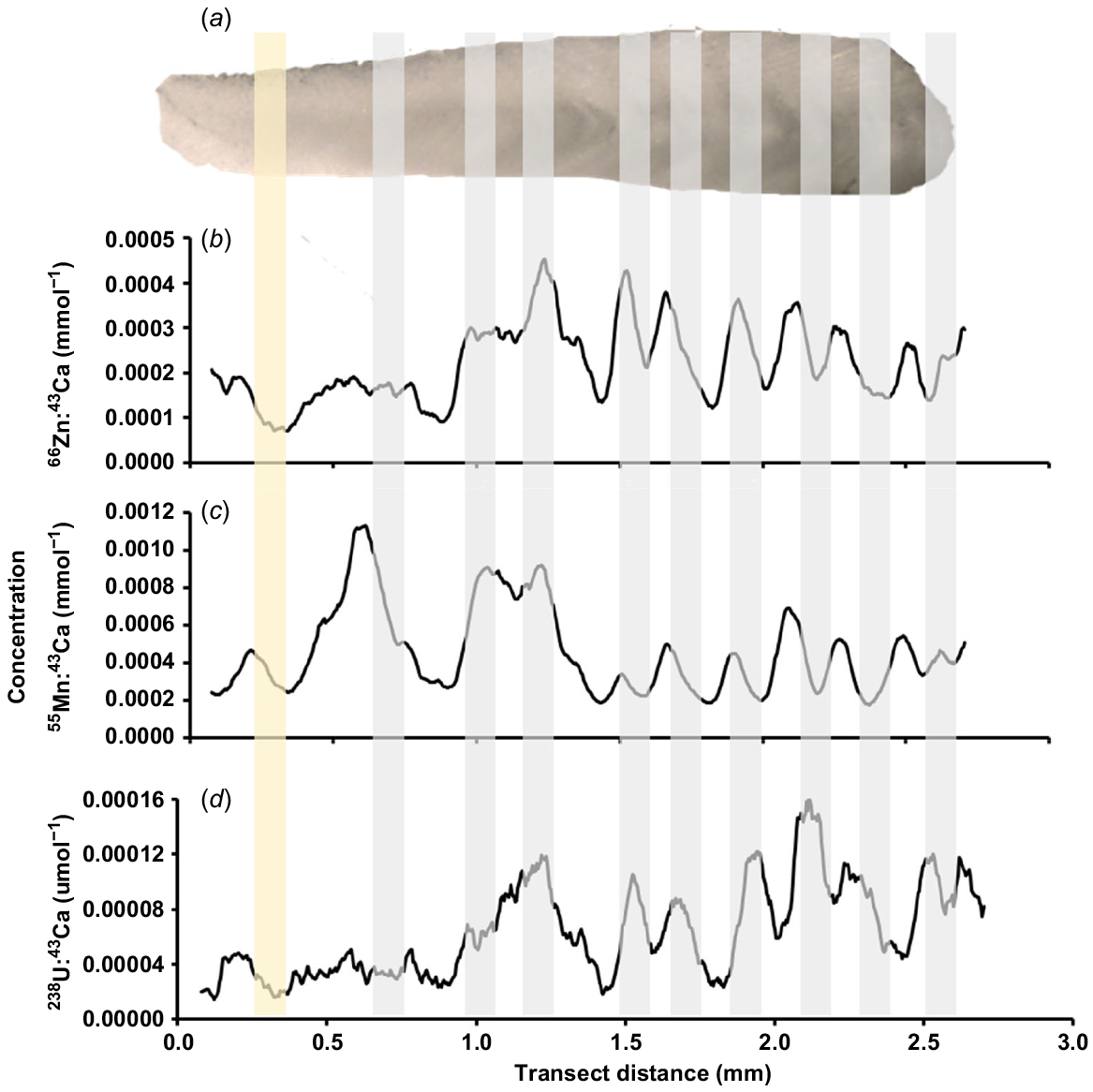
The phenomenon of double-banding was apparent in various signals, characterised by a slight dip in the peak curve evident in some vertebra and was counted only once to remain consistent with ageing rules used in visual estimation. This was also evident in many visual age estimates analysed using light microscopy. Estimates obtained using 66Zn:43Ca had the highest positive correlation to observed ages (n = 43, r2 = 0.744, P < 0.05). Evidence of systematic bias was apparent in observed age estimates of elemental peak counts, with agreement between peak counts favouring skates aged 5–7 years old.
Despite this, precision among these 66Zn:43Ca age estimates and visual counts provided the best correlation, and an APE < 10% (n = 43; APE = 6.52%). Analyses of other elements showed strong discernible signals for 55Mn, 238U and 208Pb that closely tracked the signal for 66Zn:43Ca. Although the 43Ca profiles exhibited some correlation between visual opaque counts, this pattern was not consistent between all samples and hence counts were not included in age verification. Trends in 88Sr:137Ba profile plots did show evidence of potential seasonal fluctuations among vertebrae. Variation in elemental signals was reported for 88Sr:43Ca and 137Ba:43Ca between samples, however these trends did not provide correlations with ageing and therefore were not used for verification.
Discussion
This study has provided the first sex-specific estimate of growth for Maugean skate in Macquarie Harbour. Microchemistry-corroborated age estimates provided indirect verification of seasonally deposited growth rings within the vertebrae. It is likely that Maugean skate is a relatively short-lived and fast-growing skate species, consistent with population census data and the limited growth data of recaptured individuals (sex-pooled data only) (Bell et al. 2016; Awruch et al. 2021).
Age estimates ranged from 2 to 10 years; however, females aged less than 4 years and males older than 2 years were poorly represented in the sample, which was likely to be a result of size selectivity bias in capture methods (i.e. the lack of small individuals) (Moreno et al. 2022). As skates grew older, outer bands observed in vertebrae appeared more tightly spaced, but were still discernible, which is a frequent occurrence, often resulting in age underestimation (Sulikowski et al. 2005). In view of this, precision estimate values (APEs) for both visual counts (between readers), and elemental counts (between consensus counts and elemental counts) were <30%, where agreement was most variable for larger size classes. The comparisons of these counts indicated that no appreciable bias existed between readers.
Small sample sizes often can cause poor growth parameter estimates, especially when not all length or age classes have a good representation in the sample. When biased samples compromise a robust estimation, the MMI approach is an effective tool. The six candidate growth models fitted the observed length-at-age data well regardless of sex. Nevertheless, some variability in model fit was observed; however, the inclusion of the neonatal individuals improved model fit for the under-represented younger size classes and improved length-at-birth (L0) estimates. In this study, none of the six candidate models provided a single best fit (wi > 0.90), which is suggestive of elevated model performance uncertainty and is likely to be caused by the gaps in the dataset (i.e. an absence of the younger age classes). However, the two-parameter VBGM (fixed L0) had the lowest AICc values across both sexes and sexes combined. Although the evaluation of these models was inconclusive, future work on the Maugean skate, and other endangered skate species, should include additional modelling such as cost of reproduction models, because these have the possibility of capturing additional features of growth related to life-history events (Moreno Iturria 2018).
Growth parameters suggested that Maugean skates are sexually dimorphic, where females attain a slightly larger total length than do males and grow faster, confirmed by a LRT. Both males and females have been estimated to mature at approximately the same age (5 years) (Awruch et al. 2021). This pattern is likely to be a result of differing reproductive biology, with females attaining a larger body size to accommodate egg production and storage (Awruch et al. 2021).
Although models with fixed L0 are highly applicable where small individuals are inadequately sampled and their inclusion increases statistical fit, they are limited by a failure to account for variable length-at-birth or rapid early growth, reducing accuracy. This may ensue an overestimation of L∞ and k, causing an impractical interpretation of age-at length despite improved model convergence between sexes. Additional work to update previous estimates of age and size at maturity on the basis of the results of this study should be considered in future work, and may assist in informing demographic analyses of the Maugean skate population. These results should be interpreted with caution when examining the effects of small sample size and the number of larger-sized specimens collected on the estimates of sex-specific growth parameters. Despite this, the established potential for uncertainty cannot be considered a barrier when the focus of the study is an endangered species.
There is only one estimate of age for Maugean skate in published literature, for which this study contained a subset of samples (n = 13) (Bell et al. 2016; Awruch et al. 2021). On the basis of the VBGM (M1), observed longevity is consistent with the results of this study (10 years); however, applying the theoretical calculation, results from this study differ substantially, with a theoretical maximum age of 13 years, which appears closer to a biologically reasonable estimate, given the observed age of the species in Macquarie Harbour. To contrast this, the estimated observed maximum age for larger relatives of Z. maugeana, Z. chilensis, is 27 years (Licandeo et al. 2006), and for Z. nasutus 9 years (Francis et al. 2001), elucidating the variability of skate life-histories (Table 1).
On the basis of the size and age at 50% maturity (647 mm TL for pooled sexes) determined by Bell et al. (2016), Maugean skates mature at ~5 years old, suggesting that they are potentially reproductively active for up to ~7 years. This is important because research catch data suggest that there has been a shift in their size distribution, which may indicate a drop in recruitment success, because the egg stages are significantly more vulnerable to environmental degradation owing to their immobility (Moreno et al. 2020). If there has been a decline in recruitment, or if a period of reduced recruitment persists for a protracted period, the number of active breeders in the remaining reproductive population will have decreased significantly. If this reduced recruitment has been occurring for several years, many of the skate individuals that are still reproductively active may have only a few years of effective reproduction left, exposing a critical window of opportunity for conservation action. This further highlights the population vulnerability to long-term impacts on recruitment as a whole.
Seasonal changes in temperature, light, food availability and movement behaviour can contribute to the deposition of periodic growth bands in the vertebrae of chondrichthyans, although underlying causes have been found to vary depending on the species (Tillett et al. 2011). In the case of Maugean skate, the unique biogeochemical profile of Macquarie Harbour is subject to seasonal fluctuations in environmental conditions, which may facilitate the seasonal deposition of minerals within the vertebrae (Ross et al. 2020). Although annual banding has not been validated directly for Maugean skate, results from studies of other skate species (Smith et al. 2003; McPhie and Campana 2009) and the microchemical analysis conducted here, indirectly support assumptions of an annual single band-pair deposition.
Previous work on elemental age verification has focussed on the detection of signal peaks in single elements, particularly 43Ca, which are counted to estimate age (Scharer et al. 2012). More recently, as a result of the increased measurement accuracy of light spectroscopy, non-Ca elements such as 66Mn, 88Sr and 137Ba have been proposed as an alternative to determine age, particularly in species that move across a salinity gradient such as that in Macquarie Harbour. Despite this, calculations of internal error indicated that traditional predictors of seasonality, such as strontium and barium, were less likely to contribute informative data for age verification in this study, which could be linked to the complex factors driving freshwater inflows and thermal variation in the harbour (Mohan et al. 2018).
Controlled experiments need to be conducted with other, non-endangered elasmobranchs to test the effect of various factors on the incorporation of trace metals in vertebrae. However, the evidence presented herein suggests that the concentration of Sr:Ba in Maugean skate tissue is consistent with expected seasonal fluctuations in salinity and temperature in the system and may serve as a suitable indicator to verify the annual formation of opaque bands in skate vertebrae.
Trends observed in 66Zn:43Ca, 55Mn:43Ca and 238U:43Ca near mirrored that of visual opaque band counts, displaying distinct signals that could be counted. It is understood that Maugean skate spends its entire life within Macquarie Harbour, exhibiting no seasonal migrations (Bell et al. 2016). On the basis of that understanding, if an element is incorporated environmentally and the animal does not leave the harbour system, the likely explanation for changes in microchemical signatures in the vertebrae is that the concentration of the element in the environment is changing. Past work has identified a seasonal change in river flow and oxygen concentrations within Macquarie Harbour, providing a good indication that peaks in the elements and banding in vertebrae may occur seasonally (Ross and MacLeod 2017). Field studies have also suggested that Macquarie Harbour consists of a system controlled by river run-off and seasonal heating and cooling, with surface water warmer in summer and cooler in winter, maintained by the strong salinity-driven density gradient (Wild-Allen et al. 2020). Although the relationship among temperature, salinity and DO requires further investigation, it is known that these factors influence heavy metal resuspension within aquatic environments and may link the variability in available 66Zn, 55Mn and 238U. The fluctuations evident in the signatures may relate to the historical fluctuations in river flow in Macquarie Harbour. This is further reinforced by the flux in manganese, an element potentially dietarily driven, which corresponds with band counts offering a potential source of verification (Mohan et al. 2018). The seasonal fluctuations in the ambient concentrations of these elements reinforce the value of the microchemistry in this study. Where traditional ageing cannot be achieved, counting and verifying the elemental peaks through the application of microchemistry serves as a reliable option to remove some of the uncertainty and bias often associated with ageing, by using visual band counts, a phenomenon made even more difficult in chondrichthyans such as the endangered Maugean skate.
Manganese is a useful indicator of fish age, because it is a known dietarily incorporated marker; however, it is not clear whether this also applies to elasmobranchs, and additional verification is required. (Mohan et al. 2018). Nevertheless, the seasonal nature of the mineralisation process produced a periodicity in the 55Mn:43Ca enriched bands that closely correlates with the visually assessed age of Maugean skate, with seasonal fluctuation being apparent. This relationship was particularly evident in younger fish (between 3 and 5 years) where the bands appeared discrete and were sufficiently spatially resolved to enable the ablation of a discernible signal. Suspended molecules such as 66Zn, 55Mn, 238U and lead (206Pb) determined by the resuspension of the trace metal-rich sediments in Macquarie Harbour, may, as a result, be concentrated in residents of the harbour, such as Maugean skate. Additional work may show evidence of seasonal patterns affecting heavy metal resuspension and subsequent incorporation observed in the vertebrae of Maugean skate as a result of possible resuspension in the water column.
The fast-growing, relatively early maturing and short-lived characteristics of Maugean skate may provide an evolutionary advantage by making the species more robust to acute disturbances likely to be faced in the dynamic harbour system. However, Maugean skate may also exhibit a density-dependent response to reductions in population size, potentially making the species less adept to regulating the population size, especially in response to ongoing threats (Matich et al. 2021).
Conclusions
This study has supplemented knowledge of Maugean skate life-history and ecology, allowing the generation of sex-specific growth modelling of the species by using MMI, a biologically robust approach for demographic analyses. The indirect verification of band periodicity in vertebrae through microchemical analysis suggests that, unlike in previous studies, it is heavy metals that display the most prominent chemical fingerprint for this species. The establishment of a library of existing hard parts of ageing structures (vertebrae) obtained through this study has provided a medium for future life-history studies of Maugean skate. Such results may have not been possible otherwise, where small sample sizes or endangered populations typically would impede the use of traditional procedures for age validation. Although this study emphasises the use of multi-model approaches to address uncertainties in data-poor contexts and does show promise, further analytical techniques are needed to be developed to consider and evaluate the effectiveness of the ageing technique used in this study. Future studies and long-term monitoring will help ensure that this skate population can be managed properly; in particular, age validation should be performed to verify annual vertebral band-pair periodicity for Maugean skate in Macquarie Harbour. The importance of this information is emphasised considering the uncertain status of the skate and dramatically declining environmental condition of Macquarie Harbour – the last known habitat for this endangered species.
Data availability
The datasets generated during or analysed during the current study are available from the corresponding author on reasonable request.
Declaration of funding
This study utilised samples collected by past projects funded by the Fisheries Research and the Development Corporation (FRDC).
Acknowledgements
This study would not have been possible without the work of previous sampling efforts to make the skate vertebrae available for ageing and analysis, including by various IMAS staff, in particular Justin Bell, Graeme Ewing, Ed Forbes and Jayson Semmens. We also thank Maxwell Morissette and Leonid Danyushevsky for their technical assistance with LA-ICP-MS analysis.
References
Akaike H (1973) Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika 60(2), 255-265.
| Crossref | Google Scholar |
Awruch CA, Bell JD, Semmens JM, Lyle JM (2021) Life history traits and conservation actions for the Maugean skate (Zearaja maugeana), an endangered species occupying an anthropogenically impacted estuary. Aquatic Conservation: Marine and Freshwater Ecosystems 31(8), 2178-2192.
| Crossref | Google Scholar |
Beamish RJ, Fournier DA (1981) A method for comparing the precision of a set of age determinations. Canadian Journal of Fisheries and Aquatic Sciences 38(8), 982-983.
| Crossref | Google Scholar |
Bell J, Lyle J, Semmens J, Awruch C, Moreno D, Currie S, Morash A, Ross J, Barrett N (2016) Movement, habitat utilisation and population status of the endangered Maugean skate and implications for fishing and aquaculture operations in Macquarie Harbour. Fisheries Research and Development Corporation project number 2013/008, final report. (Institute for Marine and Antarctic Studies, University of Tasmania: Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/2013-008-DLD.pdf
Bizzarro JJ, Broms KM, Logsdon MG, Ebert DA, Yoklavich MM, Kuhnz LA, Summers AP (2014) Spatial segregation in Eastern North Pacific skate assemblages. PLoS ONE 9(10), e109907.
| Crossref | Google Scholar |
Cailliet GM (2015) Perspectives on elasmobranch life-history studies: a focus on age validation and relevance to fishery management. Journal of Fish Biology 87(6), 1271-1292.
| Crossref | Google Scholar | PubMed |
Cailliet GM, Smith WD, Mollet HF, Goldman KJ (2006) Age and growth studies of chondrichthyan fishes: the need for consistency in terminology, verification, validation, and growth function fitting. Environmental Biology of Fishes 77(3), 211-228.
| Crossref | Google Scholar |
Cortés F, Colonello JH, Sammarone M, Zavatteri A, Hozbor NM (2023) Demographic analyses reveal differential biological vulnerability in four southwestern Atlantic skates. Canadian Journal of Fisheries and Aquatic Sciences 80(5), 851-865.
| Crossref | Google Scholar |
Dulvy NK, Metcalfe JD, Glanville J, Pawson MG, Reynolds JD (2000) Fishery stability, local extinctions, and shifts in community structure in skates. Conservation Biology 14(1), 283-293.
| Crossref | Google Scholar |
Fabens AJ (1965) Properties and fitting of the von Bertalanffy growth curve. Growth 29, 265-289.
| Google Scholar | PubMed |
Francis MP, Maolagáin CÓ, Stevens D (2001) Age, growth, and sexual maturity of two New Zealand endemic skates, Dipturus nasutus and D. innominatus. New Zealand Journal of Marine and Freshwater Research 35(4), 831-842.
| Crossref | Google Scholar |
Francis MP, Campana SE, Jones CM (2007) Age under-estimation in New Zealand porbeagle sharks (Lamna nasus): is there an upper limit to ages that can be determined from shark vertebrae? Marine and Freshwater Research 58(1), 10-23.
| Crossref | Google Scholar |
Goldman KJ (2005) Age and growth of elasmobranch fishes. FAO Fisheries Technical Paper 474. (Food and Agriculture Organization of the United Nations) Available at https://www.fao.org/4/a0212e/A0212E10.htm
Griffiths AM, Sims DW, Cotterell SP, El Nagar A, Ellis JR, Lynghammar A, McHugh M, Neat FC, Pade NG, Queiroz N, Serra-Pereira B (2010) Molecular markers reveal spatially segregated cryptic species in a critically endangered fish, the common skate (Dipturus batis). Proceedings of the Royal Society B: Biological Sciences 277(1687), 1497-1503.
| Crossref | Google Scholar |
Hale LF, Dudgeon JV, Mason AZ, Lowe CG (2006) Elemental signatures in the vertebral cartilage of the round stingray, Urobatis halleri, from Seal Beach, California. In ‘Age and growth of chondrichthyan fishes: new methods, techniques and analysis. Developments in Environmental Biology of Fishes, Vol. 25’. (Eds JK Carlson, KJ Goldman) pp. 317–325. (Springer: Dordrecht, Netherlands) 10.1007/978-1-4020-5570-6_12
Harry AV, Macbeth WG, Gutteridge AN, Simpfendorfer CA (2011) The life histories of endangered hammerhead sharks (Carcharhiniformes, Sphyrnidae) from the east coast of Australia. Journal of Fish Biology 78(7), 2026-2051.
| Crossref | Google Scholar | PubMed |
Katsanevakis S, Maravelias CD (2008) Modelling fish growth: multi-model inference as a better alternative to a priori using von Bertalanffy equation. Fish and Fisheries 9(2), 178-187.
| Crossref | Google Scholar |
Last PR, Gledhill DC (2007) The Maugean skate, Zearaja maugeana sp. nov. (Rajiformes: Rajidae) – a micro-endemic, Gondwanan relict from Tasmanian estuaries. Zootaxa 1494(65), 45-65.
| Crossref | Google Scholar |
Last PR, Gledhill DC, Sherman CS (2016) Maugean skate Dipturus maugeanus (formerly as: Zearaja maugeana). In ‘The IUCN Red List of Threatened Species 2016’. e.T64442A68650404. (International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/species/64442/68650404
Licandeo RR, Lamilla JG, Rubilar PG, Vega RM (2006) Age, growth, and sexual maturity of the yellownose skate Dipturus chilensis in the south-eastern Pacific. Journal of Fish Biology 68(2), 488-506.
| Crossref | Google Scholar |
Liu K-M, Wu C-B, Joung S-J, Tsai W-P, Su K-Y (2021) Multi-model approach on growth estimation and association with life history trait for elasmobranchs. Frontiers in Marine Science 8, 591692.
| Crossref | Google Scholar |
Longerich HP, Günther D, Jackson SE (1996) Elemental fractionation in laser ablation inductively coupled plasma mass spectrometry. Fresenius’ Journal of Analytical Chemistry 355(5), 538-542.
| Crossref | Google Scholar |
Lyle JM, Bell JD, Chuwen BM, Barrett N, Tracey SR, Buxton CD (2014) Assessing the impacts of gillnetting in Tasmania: implications for by-catch and biodiversity. Fisheries Research and Development Corporation project number 2010/016, final report. (Institute for Marine and Antarctic Studies, University of Tasmania: Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/2010-016-DLD.pdf
McPhie RP, Campana SE (2009) Bomb dating and age determination of skates (family Rajidae) off the eastern coast of Canada. ICES Journal of Marine Science 66(3), 546-560.
| Crossref | Google Scholar |
Matich P, Plumlee JD, Fisher M (2021) Grow fast, die young: does compensatory growth reduce survival of juvenile blacktip sharks (Carcharhinus limbatus) in the western Gulf of Mexico? Ecology and Evolution 11(22), 16280-16295.
| Crossref | Google Scholar | PubMed |
Mohan JA, Miller NR, Herzka SZ, Sosa-Nishizaki O, Kohin S, Dewar H, Kinney M, Snodgrass O, Wells RJD (2018) Elements of time and place: manganese and barium in shark vertebrae reflect age and upwelling histories. Proceedings of the Royal Society of London – B. Biological Sciences 285, 20181760.
| Crossref | Google Scholar |
Morash AJ, Lyle JM, Currie S, Bell JD, Stehfest KM, Semmens JM (2020) The endemic and endangered Maugean skate (Zearaja maugeana) exhibits short-term severe hypoxia tolerance. Conservation Physiology 8(1), coz105.
| Crossref | Google Scholar |
Moreno Iturria DA (2018) An unusual habitat for a common shark: life history, ecology and demographics of the spiny dogfish (Squalus acanthias) in Macquarie Harbour, Tasmania. PhD thesis, Institute for Marine and Antarctic Studies, University of Tasmania, Hobart, Tas., Australia. 10.25959/100.00030224
Moreno D, Lyle J, Semmens J, Morash A, Stehfest K, McAllister J, Bowen B, Barrett N (2020) Vulnerability of the endangered Maugean Skate population to degraded environmental conditions in Macquarie Harbour. Fisheries Research and Development Corporation project number 2016-068, final report. (Institute for Marine and Antarctic Studies, University of Tasmania: Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/2016-068-DLD.pdf
Moreno D, Patil J, Deagle B, Semmens JM (2022) Application of environmental DNA to survey Bathurst Harbour (Tasmania) for the Endangered Maugean Skate (Zearaja maugeana). Report to the National Environmental Science Program. Institute for Marine and Antarctic Studies, University of Tasmania, Hobart, Tas., Australia.
Morrison CM, Kunegel-Lion M, Gallagher CP, Wastle RJ, Lea EV, Loewen TN, Reist JD, Howland KL, Tierney KB (2019) Decoupling of otolith and somatic growth during anadromous migration in a northern salmonid. Canadian Journal of Fisheries and Aquatic Sciences 76(11), 1940-1953.
| Crossref | Google Scholar |
Officer RA, Gason AS, Walker TI, Clement JG (1996) Sources of variation in counts of growth increments in vertebrae from gummy shark (Mustelus antarcticus, and school shark, Galeorhinus galeus): implications for age determination. Canadian Journal of Fisheries and Aquatic Sciences 53(8), 1765-1777.
| Crossref | Google Scholar |
Outridge PM, Veinott G,, Evans RD (1995) Laser ablation ICP-MS analysis of incremental biological structures: archives of trace-element accumulation. Environmental Reviews 3(2), 160-170.
| Crossref | Google Scholar |
Raoult V, Howell N, Zahra D, Peddemors VM, Howard DL, de Jonge MD, Buchan BL, Williamson JE (2018) Localized zinc distribution in shark vertebrae suggests differential deposition during ontogeny and across vertebral structures. PLoS ONE 13(1), e0190927.
| Crossref | Google Scholar |
Ross J, MacLeod C (2017) Environmental research in Macquarie Harbour: interim synopsis of benthic and water column conditions. Report prepared for the Environmental Protection Authority and Department of Primary Industries, Parks, Water and Environment Tasmania. (Institute for Marine and Antarctic Studies, University of Tasmania: Hobart, Tas., Australia) Available at https://www.imas.utas.edu.au/__data/assets/pdf_file/0019/940303/IMAS-Technical-Report-on-Macquarie-Harbour-Condition.pdf
Ross J, MacLeod C (2018) Environmental Research in Macquarie Harbour. FRDC 2016/067: understanding oxygen dynamics and the importance for benthic recovery in Macquarie Harbour. Fisheries Research and Development Corporation project number 2016-067, 2016-067 IMAS Progress Report on Macquarie Harbour February 2018. (Institute for Marine and Antarctic Studies, University of Tasmania: Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/2016-067%20IMAS%20Progress%20Report%20on%20Macquarie%20Harbour%20February%202018.pdf
Ross J, Wild-Allen K, Andrewartha J, Beard J, Moreno D (2020) Environmental research in Macquarie Harbour. Understanding oxygen dynamics and the importance for benthic recovery in Macquarie Harbour. Fisheries Research and Development Corporation project number 2016-067, 2016-067 IMAS Progress Report on Macquarie Harbour February 2020. (Institute for Marine and Antarctic Studies, University of Tasmania, Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/2016-067%20IMAS%20Progress%20Report%20on%20Macquarie%20Harbour%20February%202020.pdf
Scharer RM, Patterson WF, III, Carlson JK, Poulakis GR (2012) Age and growth of endangered smalltooth sawfish (Pristis pectinata) verified with LA-ICP-MS analysis of vertebrae. PLoS ONE 7(10), e47850.
| Crossref | Google Scholar |
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY (2012) Fiji: an open-source platform for biological-image analysis. Nature Methods 9(7), 676-682.
| Crossref | Google Scholar |
Simpfendorfer C, Kyne PM (2009) Limited potential to recover from overfishing raises concerns for deep-sea sharks, rays and chimaeras. Environmental Conservation 36, 97-103.
| Crossref | Google Scholar |
Siskey MR, Shipley ON, Frisk MG (2019) Skating on thin ice: identifying the need for species-specific data and defined migration ecology of Rajidae spp. Fish and Fisheries 20(2), 286-302.
| Crossref | Google Scholar |
Smith SE, Mitchell RA, Fuller D (2003) Age-validation of a leopard shark (Triakis semifasciata) recaptured after 20 years. Fishery Bulletin 101(1), 194-198.
| Google Scholar |
Stevens JD, Bonfil R, Dulvy NK, Walker PA (2000) The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES Journal of Marine Science 57(3), 476-494.
| Crossref | Google Scholar |
Sulikowski JA, Tsang PCW, Howell WH (2005) Age and size at sexual maturity for the winter skate, Leucoraja ocellata, in the western Gulf of Maine based on morphological, histological and steroid hormone analyses. Environmental Biology of Fishes 72(4), 429-441.
| Crossref | Google Scholar |
Thorson JT, Simpfendorfer CA (2009) Gear selectivity and sample size effects on growth curve selection in shark age and growth studies. Fisheries Research 98(1-3), 75-84.
| Crossref | Google Scholar |
Threatened Species Link (2020) Species management profile Zearaja maugeana Maugean skate. (Department of Natural Resources and Environment Tasmania, Government of Tasmania) Available at https://www.threatenedspecieslink.tas.gov.au/Pages/Maugean-Skate.aspx
Tillett BJ, Meekan MG, Parry D, Munksgaard N, Field IC, Thorburn D, Bradshaw CJA (2011) Decoding fingerprints: elemental composition of vertebrae correlates to age-related habitat use in two morphologically similar sharks. Marine Ecology Progress Series 434, 133-142.
| Crossref | Google Scholar |
Treloar MA, Barrett NS, Edgar GJ (2017) Biology and ecology of Zearaja maugeana, an Endangered skate restricted to two south-western Tasmanian estuaries. Marine and Freshwater Research 68(5), 821-830.
| Crossref | Google Scholar |
Wild-Allen K, Andrewartha J, Baird M, Bodrossy L, Brewer E, Eriksen R, Skerratt J, Revill A, Sherrin K, Wild D (2020) Macquarie Harbour Oxygen Process model l (FRDC 2016-067): CSIRO Final Report. Fisheries Research and Development Corporation project number 2016-067, FRDC_MH_Final_Rep_June_2020. (CSIRO Oceans & Atmosphere: Hobart, Tas., Australia) Available at https://www.frdc.com.au/sites/default/files/products/FRDC_MH_Final_Rep_June_2020.pdf
Zhu L, Li L, Liang Z (2009) Comparison of six statistical approaches in the selection of appropriate fish growth models. Chinese Journal of Oceanology and Limnology 27(3), 457-467.
| Crossref | Google Scholar |