

Effects of growth path, sire type, calving time and sex on growth and carcass characteristics of beef cattle in the agricultural area of Western Australia
B. L. McIntyre A B G , G. D. Tudor A C D , D. Read A C E , W. Smart A C * , T. J. Della Bosca A C , E. J. Speijers A B and B. Orchard A FA Cooperative Research Centre for Cattle and Beef Quality, University of New England, Armidale, NSW 2351, Australia.
B Department of Agriculture and Food, Western Australia, Baron Hay Court, South Perth, WA 6151, Australia.
C Department of Agriculture and Food, Western Australia, PO Box 1231, Bunbury, WA 6230, Australia.
D 6153 Bolsena Circuit, Hope Island, Qld 4212, Australia.
E Rabobank, 2/5 Wollaston Street, Bunbury, WA 6230, Australia.
F New South Wales Department of Primary Industries Agricultural Institute, Private Bag, Wagga Wagga, NSW 2650, Australia.
G Corresponding author. Email: bmcintyre@agric.wa.gov.au
Animal Production Science 49(6) 504-514 https://doi.org/10.1071/EA08180
Submitted: 6 June 2008 Accepted: 12 November 2008 Published: 13 May 2009
Abstract
Growth, carcass characteristics and meat quality of the steer and heifer progeny of autumn (AC: March–April) and winter (WC: June–July) calving cows following weaning in January in each of 3 years (2003–05) were measured. The cows were mated to sires with a high estimated breeding value for either retail beef yield (RBY), intramuscular fat (IMF) or both RBY and IMF. After weaning, the progeny entered one of three growth paths until slaughter at an average steer liveweight of 500 kg: (i) fast – fast growth from weaning on a high concentrate feedlot diet; (ii) slow – slow growth from weaning (~0.6 kg/day) to 400 kg liveweight followed by growth at over 1 kg/day on high quality pasture; or (iii) comp. – 10% weaning weight loss, immediately after weaning followed by compensatory or rapid growth of over 1 kg/day on high quality pasture. Steers on the fast growth path had higher (P < 0.001) P8 fat thickness than those on the slow or comp. growth paths whereas heifers on the fast growth path only had higher (P < 0.001) P8 fat thickness than those on the slow growth path. Animals on the fast growth treatment had higher (P < 0.001) levels of IMF% than the slow animals which were higher (P < 0.001) than the comp. growth treatment. AUS-MEAT and US marbling scores were not different among growth paths. Animals finished on the fast growth path had a lower (P < 0.001) RBY% than those on either the slow or comp. growth paths. The RBY-sired progeny had higher (P < 0.001) finishing liveweight and hot standard carcass weight than either RBY and IMF or IMF-sired animals. IMF-sired progeny had higher (P < 0.01) rib fat thickness than either RBY or RBY- and IMF-sired animals. There was also a similar trend for P8 fat thickness but the effects were not significant. The RBY-sired animals had lower AUS-MEAT marbling scores (P < 0.01), US marbling scores (P < 0.001) and levels of IMF% (P < 0.01) than either of the other two sire treatments. RBY-sired animals also had higher (P < 0.001) estimated RBY% than those from the IMF sires while those by RBY and IMF sires were intermediate and not significantly different from either. Calving time had little influence on most carcass characteristics. However, WC animals tended to be fatter and have higher marbling scores than AC animals. The IMF% was higher (P < 0.01) in WC animals from RBY and IMF sires than in the corresponding AC animals. Heifers had lighter slaughter liveweight, carcass weight, were fatter and had higher marbling scores than steers. Heifers also had lower (P < 0.001) RBY% than the steers. Ossification scores for heifers were higher (P < 0.001) than for steers by ~30 units in AC calves and by 20 units in WC calves. The results of this experiment confirm the effectiveness of using sires with high estimated breeding value for the required characteristics in producing the desired improvements in the progeny. The absence of any interactions of sire type with growth path indicates that differences between sire types will be similar regardless of environmental conditions. Animals raised on a faster growth path after weaning produce carcasses with more fat and more IMF% than those grown on slower growth paths.
Introduction
The agricultural area of the south-west of Western Australia, which extends from Geraldton to Esperance, is responsible for the production of almost all of the high quality beef for the domestic market in Western Australia (Quin and Manners 1999). Typically, this beef comes from cattle that are less than 2 years old, with carcass weight of 220–280 kg and P8 fat thickness between 4 and 20 mm.
In the Mediterranean environment of this region, several different production systems are utilised for the supply of animals that meet this description. One source of supply is the production and slaughter of weaner calves that have been born in the late summer–autumn and slaughtered directly off the dam from early spring to mid summer at less than 12 months of age. The supply of animals suitable for slaughter in this production system is quickly reduced once the pasture dries off in summer. For the weaners that do not meet weight and fat specifications for slaughter at that time there are several possible growth paths to slaughter. First, they may enter a feedlot to be grown rapidly and slaughtered once they reach specifications. Second, they may continue to grow at a relatively slow rate in areas where summer–autumn pasture quantity and quality are maintained at a reasonable level, where animals are fed supplements or where they are grazed on irrigated pasture. Third, they may experience a period of weight stasis or loss where pasture dries off in late summer–autumn. However, animals that have been grown on pasture over summer–autumn will normally gain sufficient weight and fatness on the high quality pasture that grows over the winter–spring period to enable them to be slaughtered at ~20 months of age (Tudor et al. 1996).
The effects of different growth paths on carcass composition and meat quality characteristics have been the subject of several studies. In one study where animals were grown from weaning to slaughter at specific market weights, lot-fed animals were fatter, had more marbling (Reverter et al. 2003) and more desirable meat quality characteristics (Johnston et al. 2003) than those grown on pasture at a slower rate. In contrast, the application of divergent growth paths either before weaning (Hennessy and Morris 2003) or immediately after weaning (Tomkins et al. 2006) followed by an extended period of growth to slaughter resulted in no effect on carcass composition or meat quality characteristics.
In Western Australia there is an increasing argument for calving later in the year, after the commencement of the rainy season in autumn, onto nutritious pasture and weaning younger and lighter animals in December and January when pasture senesces. Delaying calving ensures that high quality pasture is available for the cow and calf and reduces the demand for and cost of feeding large quantities of hay (Della Bosca et al. 2004).
There is a diverse range of breeds, each with their own range of production and carcass characteristics, available for beef production. The estimated breeding values (EBV) system has been developed to quantify the genetic potential of breeding animals (Nicol et al. 1985). It is based on genetic analysis of the carcass characteristics of progeny and their relatives as well as traits measured on the live animal. With EBV of various live and carcass traits now available to producers, a clearer understanding of the likely carcass outcome from different combinations of the EBV in different management systems within a range of environments is required (Gardiner and Rutley 1995). Intramuscular fat (IMF) and retail beef yield (RBY) are two traits that can significantly influence the value of a carcass. IMF is associated with meat eating quality and can command a premium price in some export and high quality domestic markets (Hart 2001), whereas RBY is important through its effect on the proportion of the carcass that is saleable.
The aim of this project was to investigate the hypotheses that postweaning growth path will affect carcass composition and meat quality characteristics of cattle, that growth path effects will not interact with sire type and that sire types (selected for RBY and/or IMF) will perform consistent with their EBV.
Materials and methods
The details of the progeny and the production systems from both the autumn (AC, March–April) or winter (WC, June–July) calving cows has been described by Read et al. (2004) and McKiernan et al. (2005). Approximately equal numbers of weaned steers and heifers from each of the AC and WC progeny groups were selected for raising to a liveweight suitable for the Western Australian domestic market at Vasse Research Station (115.24E, 33.43S) over 3 years (2003–05). These animals were produced from a cow herd with predominantly either Angus or Murray Grey breeding. The cows were artificially inseminated using Angus sires with high accuracy (70% and above) EBV in the one percentile band for either RBY, IMF or both RBY and IMF. Two days after insemination, Angus bulls with unknown EBV status were run with the cows for a period of 6 weeks in order to maximise the reproductive rate. A total of 23 sires were used, 10 with high EBV for RBY, 11 with high EBV for IMF and 2 with high EBV for both RBY and IMF. All calves that were known to be sired by the AI bulls, either because of their gestation period, or as a result of confirmation by a DNA test were included in the study. The remainder were selected at random from progeny sired by the backup bulls. The calves were transported to the Vasse Research Station within 2–3 days of weaning in early January.
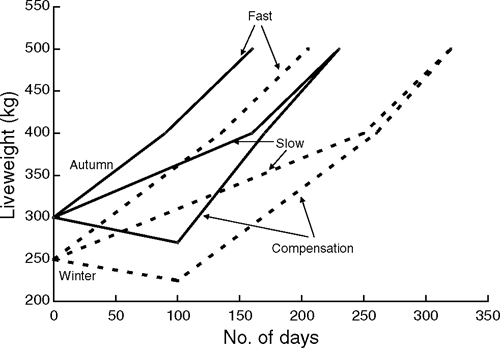
On arrival at Vasse the animals were held in small paddocks where they were fed hay and handled in the yards several times over the next 10–14 days. They were stratified into groups based on sex, sire type and liveweight and then randomly allocated to one of three replicates in each growth path treatment. The proposed growth paths are shown in Fig. 1 and further details are described below in the following sections.
|
In year 1, following almost 7 weeks on the different finishing growth path treatments the animals were reallocated to the treatments. This was due to problems associated with the initial allocation methods.
Growth paths
AC and WC – fast growth
This treatment comprised fast growth from weaning to slaughter at a final average steer liveweight of 500 kg in a feedlot on predominantly high grain diets. The fast growth path groups were fed a total mixed ration consisting of grains such as wheat, oats, barley, lupins and roughage such as pasture and oaten hay. The diets were designed to promote liveweight gains of over 1.0 kg/day and the proportions of grain and roughage contained in the mix varied over the feeding period and from year to year depending on cost and availability of ingredients.
AC and WC – slow growth
This treatment comprised slow growth from weaning (~0.6 kg/day) to 400 kg liveweight then over 1 kg/day growth rate on pasture to slaughter at a final average steer liveweight of 500 kg. In the first year of the experiment, the slow growth groups were allocated to paddocks where they grazed the available dry pasture and were fed supplementary hay at the rate of ~4 kg/head.day. It soon became apparent that the animals were not growing at the required rate so ad libitum silage feeding was commenced in early April. Two weeks later, following continued poor growth, the cattle were allowed ad libitum hay; they continued on the supplementary feed until early June but their growth was lower than expected. At this point supplementary feeding stopped and animals were moved into larger paddocks with sufficient green pasture to ensure growth.
In years 2 and 3, the slow growth groups were confined to a paddock where they were fed a restricted amount of a mixed ration of barley, lupins and hay plus ad libitum hay. This feeding system was adopted to ensure that these animals grew at their target rate of ~0.6 kg/head.day (GrazFeed 1990; AFRC 1992). This feeding method continued until the end of May when there was sufficient pasture growth to allow commencement of grazing. Supplementary feeding was discontinued and the animals were rotationally grazed at the rate of 0.35 ha/head until slaughter.
AC and WC – compensatory growth
This treatment comprised weight loss of ~10% from weaning, over a 4–5-month period, followed by compensatory (comp.) growth and finishing on pasture for slaughter at a final average steer liveweight of 500 kg. Slow and comp. growth animals were slaughtered at the same time. In the first year the comp. growth groups grazed plots previously grazed by cows to reduce the pasture to a level expected to be appropriate for the weight loss regime. They received supplementary hay at 2 kg/head.day and performed above expectation so feed had to be severely restricted to achieve the desired liveweight loss. From the end of May the cattle were stocked under conditions similar to the slow growth treatments.
In years 2 and 3 all animals in the comp. treatments were confined to a paddock where they received supplementary hay. This feeding method was adopted to better control the rate at which animals lost weight; they reached their minimum weight (loss of around 10% of weaning weight) by about the end of May. At this time they were placed onto pasture and rotationally grazed at ~0.70 ha/head.
Measurements
Animals were weighed at ~2-week intervals, to monitor progress against growth targets, and fat thickness at the P8 site was measured approximately monthly using an ultrasound scanner (PIE 200, PIE Medical, Maastricht, the Netherlands). All animals (including heifers) were slaughtered when the average liveweight of steers in the various treatment groups reached the target of ~500 kg. They were transported to the abattoir on the day before slaughter. Following slaughter, standard AUS-MEAT carcass measurements of hot standard carcass weight (HSCW), P8 fat thickness, sex and dentition (AUS-MEAT 2009) were recorded. All carcasses were graded by a Meat Standards Australia (MSA) grader who measured eye muscle area (EMA), rib fat, ossification, AUS-MEAT marbling score, US marbling score, ultimate pH, meat colour and fat colour. Carcasses were also assessed by VIAscan (Cedar Creek Company, Qld, Australia; Ferguson et al. 1995) the day following slaughter for RBY. The IMF% was measured on samples of striploin that were removed from each carcass after boning.
Statistical analysis
Carcass data were analysed using a residual maximum likelihood procedure in Genstat version 9 (Lawes Agricultural Trust: Rothamsted) to fit linear mixed models. The initial model included fixed effects of growth path treatment, season of birth, ‘type’ of sire (RBY, RBY and IMF or IMF) and sex and all interactions. Random effects comprised sire (within type), dam, slaughter group, replicate (within growth treatments), year of birth and all interactions. The association between individuals through common sire and/or dam was also included within the random effects structure. The model was refined over progressive runs by eliminating random terms showing negative or non-significant variance components and by removing non-significant interactions. The final model was then used to evaluate treatment effects and generate predicted means. Age at start of treatment and age at slaughter were included as covariates in all analyses, but were often not significant. HSCW was included as a covariate for all carcass traits (except HSCW) and was significant in most cases. Weaning liveweight was included as a covariate in analyses of slaughter liveweight and HSCW, whereas P8 fat scan was included as a covariate for the analysis of P8 fat thickness at slaughter. Log-transformations of the data (base e) were required for rib fat, AUS-MEAT marbling score and IMF%. Because probability values using the Wald test tend to give significant results too frequently (Payne 2008), P < 0.01 was chosen for tests of significance.
Predicted means were adjusted for significant covariates. Covariate adjustments were made within treatment groups where differences in covariate measurements between the groups were known to exist.
Results
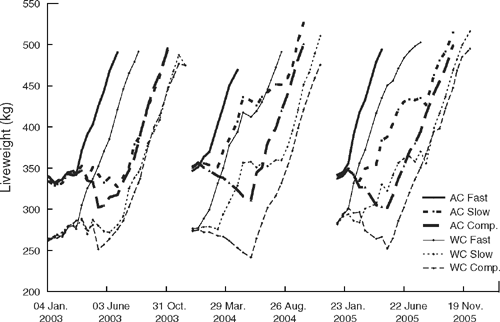
The growth of AC and WC animals from weaning in the fast, slow and comp. growth treatments in each of the 3 years of the experiment is shown in Fig. 2. In the first year, the delay to mid March in allocation of animals to their final treatments meant that there was no divergence in liveweight among the treatments for the first 3 months. By early April, the cattle in the fast growth treatment were growing rapidly once they were on their final diet. There was one period in the second year when the WC fast growth animals suffered a short period of weight loss. This was due to particularly muddy conditions in the feedlot which hampered their performance. However, rapid growth continued as soon as the animals were removed from the muddy pens. In the first year, AC and WC slow growth animals grew more slowly than anticipated on the pasture and supplementary feed and the maximum difference in liveweight between the slow and comp. growth animals for AC and WC was 40 and 28 kg, respectively. In the second and third years of the experiment, the difference in liveweight between the slow and comp. treatments varied from 77 kg for WC animals in year 3 to 121 kg for AC animals in year 2. Because of the greater divergence in weights in the second and third years, the comp. animals were lighter at slaughter than their slow growth counterparts.
|
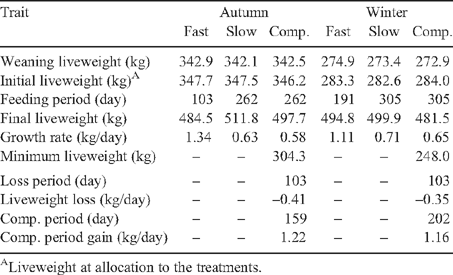
Table 1 shows the liveweights, liveweight gains and feeding periods for AC and WC calves for the different growth path treatments. All AC groups were very similar in liveweight at weaning and at the time of allocation to the treatments as were the WC groups. The AC progeny were on average 90 days older and ~75 kg heavier at weaning than the WC progeny. Liveweight gained between weaning and commencement of the treatments was 4.6 and 9.9 kg for the AC and WC animals, respectively. The average feedlot period for AC animals was 103 days compared with 191 days for the WC animals. Growth rates over the feedlot period were 1.34 and 1.11 kg/day for the AC and WC animals, respectively. While the AC animals in the slow and comp. treatments were slaughtered after 262 days with average growth rates of 0.63 and 0.58 kg/day, respectively, the WC animals took 305 days to reach the target slaughter weight and grew at 0.71 and 0.65 kg/day, respectively. The target loss of 10% liveweight in the comp. treatments was achieved with the AC animals losing 41.9 kg (12.1%) over 103 days at –0.41 kg/day and the WC animals losing 36.0 kg (12.7%) at –0.35 kg/day. Following the restriction period, the AC comp. growth animals remained on pasture for 159 days gaining at 1.22 kg/day while the WC animals grazed for 202 days and gained at an average of 1.16 kg/day.
|
Following the commencement of the rainy season in April–May in each year the available feed supply was sufficient to allow grazing by the end of May or early June; levels of pasture availability remained relatively static until the end of September when they began to increase. As a result of the lower stocking rate on the comp. growth treatments, the available pasture was maintained at a slightly higher level than in the slow growth path treatments.
Effects of growth path
The effects of growth path on live and carcass characteristics are shown in Table 2. There was no difference in slaughter liveweight among growth path treatments. However, the difference of 17 kg between the slow growth treatment compared with the fast and comp. treatments approached significance (P = 0.014). Animals from the fast growth treatment had significantly (P < 0.01) higher HSCW than those from both the slow and comp. treatments with the slow treatment being significantly (P < 0.01) heavier than the comp. treatment. For dressing%, a significant (P < 0.001) interaction was found between growth rate and calving time. The fast WC treatment had the highest (P < 0.001) dressing% followed by fast AC. These were both higher (P < 0.001) than all other growth path and calving time treatments. Comp. AC and WC treatments also had lower (P < 0.001) dressing% than the slow AC treatment. There was also a significant (P < 0.001) interaction for dressing% between growth path and sex. In both steers and heifers the fast growth treatment had a higher (P < 0.001) dressing% than the other growth path treatments. Steers had significantly (P < 0.001) higher dressing% than heifers in the fast and comp. treatments but there was no difference in the slow treatments. There was a significant growth path treatment × sex interaction for P8 fat thickness. Heifers from the fast treatment had higher (P < 0.001) P8 fat thickness than all steer treatments and heifers on the slow growth path. Steers from the slow and comp. treatments had lower P8 fat than steers on the fast treatment and heifers on the slow and comp. treatments. A significant (P < 0.001) growth path × sex interaction was also present for log rib fat thickness. It showed a similar pattern to that for P8 fat with heifers from the fast growth path having significantly (P < 0.001) higher rib fat than steers on the slow and comp. treatments and heifers on the slow growth paths. Steers from the slow and comp. treatments had lower (P < 0.001) rib fat than steers on the fast treatment and heifers on the comp. treatment. No differences in EMA, log AUS-MEAT marbling score or US marbling score were found among growth path treatments. Growth path significantly (P < 0.001) affected log IMF%, with animals from the fast treatment having a higher level then both the slow and comp. treatments. There was a significant (P < 0.01) growth path × calving time × sex interaction for ossification score. The fast treatment had lower ossification scores than the slow and comp. treatments in both steers and heifers from AC calves. In addition, the comp. growth treatment had a lower ossification score than the slow growth treatment in AC heifer calves. There were no differences in ossification score among growth treatments in either steer or heifer WC calves. RBY as estimated from VIAscan was significantly (P < 0.001) affected by growth path and was highest for the slow growth treatment and lowest for the fast treatment. Growth path had no effect on sensory meat quality score estimated from the MSA model.

|
Effects of sire type
The effects of sire type on live and carcass characteristics are shown in Table 3. The progeny of animals with high EBV for RBY were heavier at slaughter (P < 0.001) and had ~10 kg heavier carcasses (P < 0.001) than either of the other two types, which were not different. Neither dressing% nor P8 fat thickness was significantly (P > 0.01) different among sire type treatments. The log rib fat thickness of the progeny of IMF sires was higher (P < 0.01) than progeny of either RBY or RBY and IMF sires, which were not different. There were no differences in EMA among sire types. Log AUS-MEAT marbling scores were significantly (P < 0.01) lower in the RBY-sired individuals than in both their RBY and IMF and IMF-sired contemporaries. US marbling score showed a very similar pattern with the RBY animals having lower scores (P < 0.001) than the other two types. For log IMF% there was a significant (P < 0.01) sire carcass type × calving time interaction. RBY-sired progeny had lower (P < 0.01) log IMF% than both the other two types in the AC group whereas in the WC groups the differences were only significant (P < 0.01) between RBY and IMF. Ossification scores did not differ between the three sire types. RBY for the RBY-sired animals was significantly (P < 0.001) higher than for the IMF-sired progeny with RBY and IMF intermediate and not significantly different from the other sire types. Meat quality scores estimated from the MSA model were significantly (P < 0.01) higher for the IMF-sired animals than for the RBY-sired animals but were similar to those for the RBY- and IMF-sired animals.

|
Effects of calving time
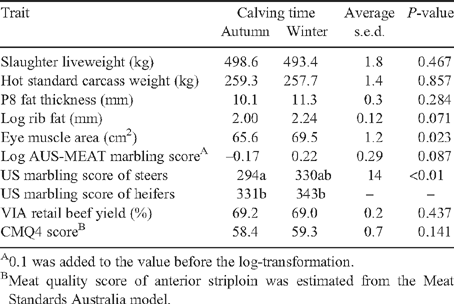
The effect of calving time on the performance and carcass characteristics of the AC- and WC-born calves shows there were no significant differences in slaughter liveweight, carcass weight, P8, log rib fat, EMA and MSA marbling (Table 4). There was a significant (P < 0.001) interaction between growth path and calving time (Table 2) for dressing%, with differences between calving times significant only within the fast growth path treatments, within which the value for WC was greater than that for AC. A significant (P < 0.01) calving time × sex interaction was evident for US marbling score with WC heifers having a higher score than AC steers but no different from WC steers or AC heifers (Table 4). There was a significant calving time × sire type interaction for log IMF% (Table 3). In all sire types, WC had higher log IMF% and the differences were significant (P < 0.01) for both RBY- and IMF-sired animals. Ossification scores showed significant (P < 0.01) interactions among calving time, growth path and sex (Table 2). The AC and WC heifers from the slow and comp. growth path treatments had higher ossification scores than their WC counterparts. In steers, AC from the fast growth path had lower ossification than WC from the slow and comp. treatments. RBY and CMQ4 score were not different between AC and WC.
|
Effects of sex
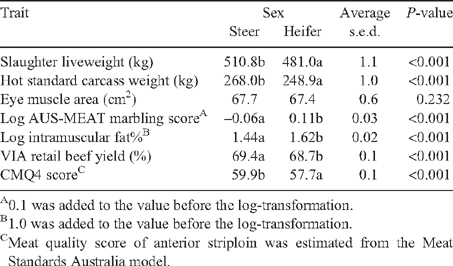
Predicted means for postweaning growth and carcass characteristics of steers and heifers are shown in Table 5. Slaughter liveweights for steers were higher (P < 0.001) than heifers and steers had ~20 kg higher HSCW (P < 0.001) than heifers. A significant (P < 0.01) growth path × sex interaction was present for dressing% (Table 2) with steers having higher dressing% than heifers in both the fast and slow growth paths but not in the comp. growth path. There were significant (P < 0.001) growth path × sex interaction for both P8 fat thickness and log rib fat (Table 2). For P8 fat thickness, steers had lower fat thickness within each growth path treatment. Similarly, in the log rib fat thickness, steers were leaner in both the slow and comp. treatments but not in the fast growth treatment. The EMA did not differ between heifers and steers. Heifers had higher AUS-MEAT marbling score (P < 0.01) than steers. For the US marbling score there was a significant (P < 0.01) calving time × sex interaction (Table 4) where AC steers had a lower score than AC heifers but there was no difference in score between WC steers and heifers. For log AUS-MEAT marbling score, heifers had higher (P < 0.001) scores than steers. Log IMF% of the steers was significantly (P < 0.001) lower than that of the heifers. As shown in Table 2, there was a significant (P < 0.01) sex × growth path × calving time interaction for ossification score. In all combinations of calving time and growth path, heifers had higher (P < 0.01) ossification scores than steers. While ossification score for steers ranged between 120 and 130, the range for heifers was between 150 and 170. The estimated RBY for steers was significantly (P < 0.001) greater than that for heifers by 0.7%. Steers were assessed as being significantly (P < 0.001) more tender than heifers with over a 2-point difference in CMQ4 score as estimated from the MSA model.
|
Discussion
Finishing growth path had several important effects on carcass characteristics, most of which related to the composition of the animal, particularly fatness. Animals on the fast growth path generally had higher P8 and rib fat thickness and IMF% than the animals in the pasture-finished treatments. These differences in fat content are likely to be a combination of diet and growth rate. McIntyre and Ryan (1984) reported significantly higher fat thickness at the 12th rib in cattle grown at a similar rate on a ration containing 70% grain compared with those grown on pasture. Tudor and Utting (1978), Tudor (1992) and Reverter et al. (2003) found higher fat content in the carcasses of grain compared with pasture-fed animals. Robinson et al. (2001) grew cattle on three different growth pathways to a liveweight of 400 kg before finishing on either a feedlot (75% grain) or pasture for two different market destinations. For each prefinishing growth pathway and market destination, after adjusting for differences in carcass weight between finishing diet, P8 fat, rib fat and IMF% were higher for the feedlot-finished animals. Pethick et al. (2004) in reviewing results of the Beef Cooperative Research Centre studies concluded that at the same carcass weight IMF content in pasture-finished animals was reduced by 40% for every 10-kg increase in carcass weight compared with feedlot-finished animals.
The absence of any differences in composition, yield or meat quality measurements between the slow and comp. growth path treatments is consistent with results reported by Hennessy and Morris (2003) and Tomkins et al. (2006). In both of these studies restricted growth followed by an extended period of realimentation on pasture before slaughter did not affect carcass composition or meat quality characteristics.
There was also a significant effect of growth path on RBY% with the fast growth treatment having the lowest yield, whereas the comp. growth treatment had the highest yield. These differences in yield were inversely associated with the patterns in both P8 and rib fat, which were lowest in the slow growth treatment and highest in the fast growth treatment. The effects of fatness on yield were highlighted earlier in the paper.
As a result of their higher dressing%, animals on the fast growth treatment had the highest HSCW despite having the lightest final liveweight. This is consistent with previous studies which showed slaughtered cattle finished on high grain diets had less gut fill and a higher dressing% than cattle finished on pasture (Tudor et al. 1980; McIntyre and Ryan 1982, 1984).
Ossification of the cartilaginous tissue over the bone in the sacral, lumbar and thoracic vertebrae has long been used along with other assessments to estimate age or ‘maturity’ in the US as part of their quality grading system (USDA 1965, 1989). Ossification has been adopted by the MSA grading system in Australia and Anon. (2001) indicated that as ossification scores range between 100–170 age ranges from 9 to 24 months. Hence the ossification scores of carcasses in this experiment were well within the expected range. The lower ossification scores that were found on the AC fast growth path rather than on the slow and comp. treatments in both steers and heifers were consistent with differences in the age at slaughter, since animals in the fast growth treatment were over 150 days younger than those in the slow and comp. treatments. However, the same trend was not apparent in the WC group where there was a similar difference in age between the fast and slow and comp. treatments. The inconsistency of these results may be explained by the difficulty in maintaining a consistent assessment of ossification by MSA graders. Over the 3 years of the experiment there were 12 different slaughters as a result of separate slaughter times for animals from the AC and WC treatments as well as the fast growth path treatment having a different slaughter time than the slow and comp. growth path treatments. In addition, the use of six different graders over the course of the experiment further adds to the possibility of inconsistency in the assessment of ossification.
The results of this experiment provide unequivocal evidence for the use of EBV in cattle breeding programs designed to enhance the performance in some important carcass characteristics. The use of bulls differing in EBV for both IMF and RBY resulted in significant differences in those characteristics in the progeny. The average EBV (for IMF%) of the sires selected to represent high IMF potential was 1.5% compared with –1.0% for those sires selected for RBY. Based on these values a difference of 1.25 would be expected in the progeny from the two sire carcass types. The actual difference of ~1.0% for AC and 0.7% for WC was, therefore, slightly below the expected value. Part of the explanation of these differences (0.25 and 0.55%) may be due to the fact that these cattle were slaughtered at relatively low carcass weights and low levels of fatness and marbling and they may not have had a chance to fully express their marbling potential. Such differences from expectation may also occur because the accuracy of the EBV is not 100%. Pethick et al. (2004) noted that IMF is relatively late developing and is deposited at a greater rate than muscle later in life so that its concentration will increase as the animal gets heavier and fatter. When compared with the time on feed that animals are normally fed for high marbling markets and carcass weights, those in this experiment were at a relatively early stage in their development of IMF. Also, Sundstrom (2002) pointed out that the EBV are expressed as equivalent to a carcass weight of 300 kg, considerably heavier than the carcasses in our experiment. In studies with sires selected on a similar basis as those in our experiment, Graham et al. (2009), Wilkins et al. (2009) and Deland et al. (pers. comm.) found differences in IMF content of 0.8, 0.7 and 0.5%, respectively, between sires selected for IMF and RBY. The commercially important measures of IMF, namely the AUS-MEAT, marbling score and the US marbling score were consistent with the measured IMF content.
Important differences between sire types were evident by the time their progeny had reached weaning where the IMF progeny were lighter and had more P8 fat cover (as measured by ultrasound scan) than RBY progeny (Read et al. 2005). These differences were carried through to slaughter where IMF-sired animals had higher rib fat thickness than those with RBY sires. P8 fat thickness also showed a similar trend although the difference was not significant. These fat thickness measurements were also consistent with the EBV for those traits as IMF sires invariably had EBV for both rib and P8 fat thickness greater than 0.0 whereas those for RBY sires were always less than 0.0. This association between IMF/marbling and fat thickness raises a question about what is being represented by the EBV. The EBV is an estimate of genetic merit for a defined trait relative to some specified contemporaries. The higher fat thickness and presumably greater overall carcass fat content may mean that the animals with higher EBV for IMF are simply further along the growth path as described by Pethick et al. (2004) and are therefore earlier maturing animals than those with a lower EBV for IMF. A more desirable situation would be that the higher EBV represents a difference in fat distribution whereby a higher proportion of the total fat is located in the intramuscular depot.
The results from the RBY assessments were also consistent with expectations from the EBV with differences between the RBY- and IMF-sired animals. While RBY-sired progeny would be expected to have an RBY of around 2% higher than IMF-sired animals, the actual value was only 0.7%. According to Sundstrom (2002) the EBV for RBY is calculated mainly from measurements of weight, fat thickness and EMA. Measures of fatness have a close relationship with carcass composition and in particular the percentage of fat in the carcass (Butterfield 1965; Charles 1977; Johnson and Vidyadaran 1981). Numerous workers have derived relationships between yield of meat and measurements of fat thickness. McIntyre and Frapple (1988) found that the regression coefficient of the relationship between yield of saleable meat and either P8 or 12th rib fat thickness was ~–0.7 indicating a decline in yield of 0.7% for each 1-mm increase in thickness. Ball (1984) found the decline in yield with increasing fat thickness varied between –0.14 and –0.39% when different relationships for P8 and 12th rib fat thickness were derived for steers and heifers whereas Johnson (1987) found similarly low regression coefficients of between –0.15 and –0.27 for P8 and 12th rib measurements. In the present study, the yield of RBY-sired animals was ~0.7% higher than that of the IMF-sired animals whereas the P8 fat thickness was ~1 mm lower. This indicates that at least some of the advantage in yield for the RBY types was due to their lower fat thickness. However, some of the difference was also due to EMA (Sundstrom 2002) as EMA often contributes to the accuracy of predicting yield% (Henderson et al. 1966; Martin et al. 1970; Perry et al. 1993a, 1993b). Although there were no significant differences, RBY-sired animals tended to have higher EMA than IMF-sired animals and this could account for the remaining difference in yield estimated using the VIAscan. These results were consistent with those of Graham et al. (2009) and Wilkins et al. (2009) who also found differences in VIAscan estimates of RBY- and IMF-sired animals. As in the present study, RBY-sired animals tended to have lower fat thickness and larger EMA. The higher finishing liveweight of the RBY-sired animals compared with the RBY and IMF and IMF-sired animals (Table 3) was also consistent with the sire EBV for liveweight. HSCW followed the same pattern as liveweight since dressing% for the three types was very similar.
The palatability of the striploin as predicted by the MSA model showed that the IMF-sired animals scored 1.5 units higher than the RBY-sired animals (Table 3). Although this seems to be a small difference on the sensory scoring scale from 0 to 100 it was significant (P < 0.001) and would be mostly accounted for by the 35-point difference in US marbling score between IMF- and RBY-sired progeny. Thompson (2002), in describing the operation of the MSA model, indicated that an increase in US marbling score from 250 to 550 in the striploin resulted in an increase in 8 units in the estimated palatability score. Assuming a linear relationship between US marbling score and palatability this equates to the equivalent of 1 unit for every 37.5 US marbling score units. Other factors, such as HSCW, rib fat, muscle pH, aging time and ossification that can influence the MSA score differed between types by such small amounts that they would have minor effects on the estimated score. Wilkins et al. (pers. comm.) and Graham et al. (2009) found similar differences in the palatability score estimated from the MSA model. The striploin of animals in the study of Wilkins et al. (2009) were also subjected to the MSA sensory taste panel. These results confirmed the pattern estimated from the MSA predictive model but the differences between the RBY and IMF types were much larger and amounted to 5.5 units for the sensory panel compared with only 0.9 units estimated from the model (J. Wilkins, pers. comm.).
One of the important findings arising from this experiment was the absence of any interactions between sire type and growth path. This means that the sire carcass types can be used with confidence provided sires used have EBV at least as accurate as those of the bulls used here. The performance expected in the progeny will be predictable from the EBV, at least over the range of growth scenarios imposed in this experiment. Selection for one trait will be associated with changes in other traits. In our experiment, the use of sires selected for high EBV for IMF was associated with increases in EBV for fatness and reduced EBV for yield and growth rate. However, the use of EBV provides the opportunity to select sires that allow simultaneous improvement in more than one trait, such as those selected for both RBY and IMF in this experiment. Therefore, it is most important to assess the total package so that improvements in one characteristic are not at the expense of others.
Calving time had very little influence on most carcass characteristics (Table 4). However, there was a consistent trend for WC animals to have higher marbling and IMF% than the AC animals although the differences were only significant for IMF%. These animals were slightly younger and fatter (but not significantly so) than the AC animals and this may account for their greater tendency to marble (Pethick et al. (2004). There was also a slight tendency for AC calves to have higher ossification scores than WC calves. This is possibly due to the chronological age difference of ~2 months at slaughter. However, this trend was only present in heifers from the slow and comp. treatments. In all other cases no effects were apparent. The performance of the WC animals demonstrated that they have the potential to fulfil an important role in the production systems in the south-west of Western Australia. Weaning these animals younger and at lighter weights than their AC counterparts presents the industry with animals that may be more suited to turnoff at different times than the current majority practice. This, in turn, will produce a more consistent supply of high quality beef and reduce the peak in turnoff that still characterises the Western Australian industry. WC had no deleterious effect on subsequent growth rate or carcass characteristics. In fact their slightly superior performance to that of the AC calves in both IMF% and ossification indicates that their MSA grade will be slightly better.
The effects of sex on growth and carcass characteristics showed the expected trends with heifers growing slower and having lighter carcasses than steers. Heifers were also fatter and tended to have slightly higher marbling scores than the steers. As a result of their greater fatness, heifers had lower yield of beef than the steers. The ossification scores confirmed previous findings that heifers have higher scores than steers at the same age (Thompson et al. 2008). In this experiment, the steers had scores ~30 units lower than heifers in AC calves and ~20 units lower in WC calves.
Meeting the target growth paths (Fig. 1), particularly for the slow and comp. pasture treatments, proved relatively difficult to achieve (Fig. 2). As would be expected, the growth of animals on the fast treatment with the regular intake of a high energy, balanced diet most closely matched the target growth path. There was one brief period in year 2 when the WC animals lost weight. This coincided with a period of wet weather that also resulted in weight losses in all other treatments. The animals resumed growth when they were moved from ‘muddy’ to ‘clean’ feedlot pens. Regulating the slow growth path was the most difficult. In the first year, the delay in allocating the weaners to their final growth paths and subsequent poorer than expected growth on hay and silage supplements resulted in these animals being well below their target liveweight by the time sufficient pasture was available to promote rapid liveweight gain. In the subsequent 2 years when animals were fed a mixed ration calculated to achieve a growth rate of 0.6 kg/day, their growth more closely matched the target. Apart from the first year, which was affected by the delay in commencement of the treatments, the animals on the comp. treatment showed a steady decline in liveweight from weaning and lost the target of 10% of their liveweight at weaning by the start of the growing season. Thereafter, all animals gained liveweight through spring and early summer and it was apparent that the imposition of the different growth paths had not influenced their ability to grow rapidly once adequate quantities of high quality green feed were available. The losses in weight of the WC animals just before slaughter in the first year and slightly reduced gains in the third year were undoubtedly due to their later finishing time and the deterioration in pasture quality as the pasture dried off at the end of the growing season.
While the aim of the experiment was to slaughter the slow and comp. growth treatments from the same calving time simultaneously and at the same average final liveweight (500 kg for the steers), this was only achieved in the first year of the experiment. As noted earlier, the maximum divergence in liveweight between the slow and comp. treatments was only 30–40 kg and this difference rapidly disappeared once high quality green pasture was available. However, in the second and third years, the divergence between the slow and comp. treatments commenced much earlier and was much greater (over 75 kg). In addition, the strategy of grazing the animals on the slow treatments at a heavier stocking rate was also successful in restricting their growth rate early in the growing season and ensuring that the divergence in liveweight lasted for much longer. As shown in Fig. 2 the animals in the comp. treatments remained lighter through to slaughter. The failure of the animals on the comp. treatment to fully compensate following the weight lost after weaning is consistent with the results of studies by Tudor and O’Rourke (1980), Ryan et al. (1993) and Tomkins et al. (2006) who also reported that cattle did not completely compensate when finished on pasture.
Slow growth treatments were slaughtered at significantly (P < 0.01) higher liveweights than either fast or comp. growth due to several factors. First, as discussed above, the comp. growth treatments failed to fully compensate in years 2 and 3 by the time the animals reached the target slaughter liveweight. Second, it was not possible to slaughter animals exactly at their target liveweight as the animals had to be scheduled for slaughter some time in advance. Third, the growth during the period between scheduling and actual slaughter could not be accurately predicted.
Conclusion
Finishing growth path from weaning to slaughter had a significant effect on carcass characteristics, including fatness, ossification, IMF% and yield. While fast growth in a feedlot favours greater fatness and IMF%, slow and comp. growth treatments produced greater yield at similar carcass weight. The choice of finishing regime will be determined primarily by economic factors and market requirements.
The use of sires selected for specific EBV produced progeny with carcasses with higher RBY or higher IMF, depending on the sire EBV (and its associated accuracy). This confirmation of the value of using accurate EBV will encourage industry to actively target specific markets, which should increase economic value of each carcass particularly when the results showed a high degree of consistency over a range in growth paths.
The experiment has also demonstrated that there are no disadvantages in terms of growth rate and carcass characteristics from using a later calving strategy.
Acknowledgements
We gratefully acknowledge the support of our commercial cooperator, Alcoa Farmlands and Tony Hiscock in particular. This work was supported by the Department of Agriculture and Food, Western Australia, the Cooperative Research Centre for Cattle and Beef Quality and Meat and Livestock Australia. We also gratefully acknowledge the input of Mr Jim Walkley and his helpful advice in the drafting and revision of this paper.
Butterfield RM
(1965) The relationship of carcase measurements and dissection data to beef carcase composition. Research in Veterinary Science 6, 24–32.
| PubMed |
[Verified 21 January 2009]
Henderson DW,
Goll DE, Kline EA
(1966) Measures of carcass yield and tenderness of two muscles in four groups of beef carcasses. Journal of Animal Science 25, 329–333.

Hennessy DW, Morris SG
(2003) Effect of a preweaning growth restriction on the subsequent growth and meat quality of yearling steers and heifers. Australian Journal of Experimental Agriculture 43, 335–341.
| Crossref | GoogleScholarGoogle Scholar |
Johnson ER
(1987) Comparison of twelfth rib and rump fat thickness measurements for predicting commercial beef yield in local market carcasses. Australian Journal of Experimental Agriculture 27, 613–617.
| Crossref | GoogleScholarGoogle Scholar |
Johnson ER, Vidyadaran MK
(1981) An evaluation of different sites for measuring fat thickness in the beef carcass to determine carcass fatness. Australian Journal of Agricultural Research 32, 999–1007.
| Crossref | GoogleScholarGoogle Scholar |
Johnston DJ,
Reverter A,
Ferguson DM,
Thompson JM, Burrow HM
(2003) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 3. Meat quality traits. Australian Journal of Agricultural Research 54, 135–147.
| Crossref | GoogleScholarGoogle Scholar |
Martin AH,
Fredeen HT,
Weiss GM, Newman JA
(1970) Prediction of lean yield of beef carcasses. Canadian Journal of Animal Science 50, 31–41.
McIntyre BL, Frapple PG
(1988) Estimation of the yield of beef carcases from measurements of fat thickness over the rib and rump. Animal Production in Australia 17, 242–245.
McIntyre BL, Ryan WJ
(1982) Estimating carcase weight in live cattle. Animal Production in Australia 14, 353–356.
McIntyre BL, Ryan WJ
(1984) The influence of type of diet and electrical stimulation on the eating quality of beef. Animal Production in Australia 15, 468–471.
McKiernan WA,
Wilkins JF,
Barwick SA,
Tudor GD,
McIntyre BL,
Graham JF,
Deland MPB, Davies L
(2005) CRC ‘Regional Combinations’ project – effects of genetics and growth paths on beef production and meat quality: experimental design, methods and measurements. Australian Journal of Experimental Agriculture 45, 959–969.
| Crossref | GoogleScholarGoogle Scholar |
Nicol DC,
Graser H-U,
Tier B, Hammond K
(1985) BREEDPLAN – a new within-herd evaluation system for the National Beef Recording Scheme. Proceedings of the Australian Association of Animal Breeding and Genetics 5, 151–153.
Perry D,
McKiernan WA, Yeates AP
(1993a) Muscle score: its usefulness in describing the potential yield of saleable meat from live steers and their carcasses. Australian Journal of Experimental Agriculture 33, 275–281.
| Crossref | GoogleScholarGoogle Scholar |
Perry D,
McKiernan WA, Yeates AP
(1993b) Meat yield and subjective muscle scores in medium weight steers. Australian Journal of Experimental Agriculture 33, 825–831.
| Crossref | GoogleScholarGoogle Scholar |
Pethick DW,
Harper GS, Oddy VH
(2004) Growth, development and nutritional manipulation of marbling in cattle: a review. Australian Journal of Experimental Agriculture 44, 705–715.
| Crossref | GoogleScholarGoogle Scholar |
Read D,
Della Bosca TJ,
Hiscock AJF,
McIntyre BL,
Smart WL,
Taylor EG, Tudor GD
(2004) Optimising feed supply, reproductive efficiency and progeny growth to meet market specifications. 2. Reproduction in autumn and winter calving herds. Animal Production in Australia 25, 304.
Read DM,
McIntyre BL,
Taylor EG, Tudor GD
(2005) Effect of time of calving on growth of progeny to weaning. Proceedings of the Australian Association of Animal Breeding and Genetics 16, 342–345.
Reverter A,
Johnston DJ,
Perry D,
Goddard ME, Burrow HM
(2003) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 2. Abattoir carcass traits. Australian Journal of Agricultural Research 54, 119–134.
| Crossref | GoogleScholarGoogle Scholar |
Robinson DL,
Oddy VH,
Dicker RW, McPhee M
(2001) Post-weaning growth in northern New South Wales. 3. Carry-over effects on finishing, carcass characteristics and intramuscular fat. Australian Journal of Experimental Agriculture 41, 1041–1049.
| Crossref | GoogleScholarGoogle Scholar |
Ryan WJ,
Williams IH, Moir RJ
(1993) Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights. Australian Journal of Agricultural Research 44, 1623–1633.
| Crossref | GoogleScholarGoogle Scholar |
Thompson JM
(2002) Managing meat tenderness. Meat Science 62, 295–308.
| Crossref | GoogleScholarGoogle Scholar |
Thompson JM,
McIntyre BL,
Tudor GD,
Pethick DW,
Polkinghorne R, Watson R
(2008) Effects of Hormonal Growth Promotants (HGP) on growth, carcass characteristics, the palatability of different muscles in the beef carcass and their interaction with aging. Australian Journal of Experimental Agriculture 48, 1405–1414.
| Crossref | GoogleScholarGoogle Scholar |
Tomkins NW,
Harper GS,
Bruce HL, Hunter RA
(2006) Effect of different post-weaning growth paths on long-term weight gain, carcass characteristics and eating quality of beef cattle. Australian Journal of Experimental Agriculture 46, 1571–1578.
| Crossref | GoogleScholarGoogle Scholar |
Tudor GD
(1992) Effect of diet on fat deposition in cattle. Animal Production in Australia 19, 89.
Tudor GD, O’Rourke PK
(1980) The effect of pre- and post-natal nutrition on the growth of beef cattle. II. The effect of severe nutrition in early post-natal life on growth and feed efficiency during recovery. Australian Journal of Agricultural Research 31, 179–189.
| Crossref | GoogleScholarGoogle Scholar |
Tudor GD, Utting DW
(1978) The influence of diet and growth rate on the composition of the 9–10–11th rib. Animal Production in Australia 12, 235.
Tudor GD,
Utting DW, O’Rourke PK
(1980) The effect of pre- and post-natal nutrition on the growth of beef cattle. III. The effect of severe nutrition in early post-natal life on the development of the body components and chemical composition. Australian Journal of Agricultural Research 31, 191–204.
| Crossref | GoogleScholarGoogle Scholar |
Tudor GD,
McMullen GR,
Snowdon JM, Paszkudzka-Baizert L
(1996) Yearling beef production off pasture in the south-west of Western Australia. Animal Production in Australia 21, 81.
Wilkins JF,
McKiernan WA,
Irwin J,
Orchard B, Barwick SA
(2009) Performance of steer progeny of sires differing in genetic potential for fatness and meat yield following post-weaning growth at different rates. 1. Growth and live-animal composition. Animal Production Science 49, 515–524.
| Crossref | GoogleScholarGoogle Scholar |
* Deceased.