

Performance of steer progeny of sires differing in genetic potential for fatness and meat yield following post-weaning growth at different rates. 1. Growth and live-animal composition
J. F. Wilkins A B F , W. A. McKiernan A C , J. Irwin A D , B. Orchard A B and S. A. Barwick A EA Cooperative Research Centre for Beef Genetic Technologies, University of New England, Armidale, NSW 2351, Australia.
B NSW Department of Primary Industries, Wagga Wagga Agricultural Institute, Private Mail Bag, Wagga Wagga, NSW 2650, Australia.
C NSW Department of Primary Industries, Locked Bag 21, Orange, NSW 2800, Australia.
D NSW Department of Primary Industries, Private Mail Bag, Yanco, NSW 2703, Australia.
E Animal Genetics and Breeding Unit, University of New England, Armidale, NSW 2351, Australia.
F Corresponding author. Email: john.wilkins@dpi.nsw.gov.au
Animal Production Science 49(6) 515-524 https://doi.org/10.1071/EA08268
Submitted: 1 November 2008 Accepted: 11 February 2009 Published: 13 May 2009
Abstract
The present experiment, ‘Regional Combinations’, examined growth, and carcass- and meat-quality traits in the progeny of sires genetically diverse for fatness and meat yield when grown at different rates from weaning to feedlot entry. The present paper is the first of several papers describing results from the New South Wales site, one of four in the project. It reports the effects on growth and body composition of steers during backgrounding and feedlot finishing phases. A total of 43 sires within three carcass-class categories, defined as high potential for meat yield, for marbling or for both traits, was used, based on estimated breeding values for retail beef yield and intramuscular fat. Sires were drawn from Angus, Charolais, Limousin, Black Wagyu and Red Wagyu breeds, providing a range of carcass sire types across the three carcass classes. Matings were by artificial insemination to Hereford dams from a single herd. Steer progeny were grown at conventional (slow: ~0.5 kg/day) or accelerated (fast: ~0.7 kg/day) rates from weaning to feedlot entry weight, targeting group means of 400 kg. Accelerated and conventionally grown groups from successive calvings entered the feedlot at similar entry liveweights at the same time, then having identical management during the 100-day finishing phase before slaughter. Within finishing cohorts, fast backgrounding growth resulted in increased subcutaneous fatness at feedlot entry in steers of all carcass types. Slow growth during backgrounding resulted in faster (compensatory) growth in the feedlot in all classes and sire types. This increased the deposition of fat in slow-backgrounded steers compared with that in fast-backgrounded steers during feedlotting, and thus reduced the difference between the groups in P8 and rib fat at feedlot exit. However, there did appear to be an advantage in the level of compensation in the feedlot in favour of those sire types with a genetic propensity for faster growth. Backgrounding growth rate affected body composition and the rate of weight gain during finishing. Faster growth produced more subcutaneous fat during both backgrounding and finishing. Steer progeny groups clearly showed the expected responses in growth and body composition, on the basis of the genetic potential of their sires.
Additional keywords: compensatory growth, growth path, intramuscular fat, estimated breeding values, retail beef yield.
Introduction
Studies into genetic and nutritional issues affecting beef production, such as the importance of sire carcass types and effects of nutritional restrictions during growth on subsequent performance, were initiated within the first phase of Beef Cooperative Research Centres (Beef CRC) research from 1993 to 2000 (Bindon 2001). This was a significant contributor to the establishment of the Meat Standards Australia (MSA) grading scheme (Polkinghorne et al. 2008) and the development of heritability estimates and estimated breeding values (EBV) for carcass traits that formed part of the overall aim of providing industry relevant recommendations and tools to assist management decisions. The scope of genetics research covered by past and ongoing studies within Beef CRC programs was discussed in detail by Burrow and Bindon (2005). Results of the initial work were reported by Robinson et al. (2001) and Upton et al. (2001) and covered a limited range of breed and carcass types (Angus, Hereford, Murray Grey and Shorthorn). Subsequently, the project known as ‘Regional Combinations’, on which we report here, was conducted to extend the previous findings by examining sire carcass types with potential for more extreme production and carcass characteristics while imposing a variety of regional-specific growth and finishing regimens in beef production systems in different environments across southern Australia. The design for this multisite experiment was described by McKiernan et al. (2005). Preliminary results from the various sites have been previously reported by Wilkins et al. (2002, 2004), Tudor et al. (2004), Graham et al. (2005), McKiernan et al. (2006) and Irwin et al. (2006).
Accurate prediction of responses to genetic and nutritional treatments is vital to industry uptake of management strategies so that targeted endpoints can be reliably achieved. Furthermore, the supply of slaughter cattle with consistently high compliance to target specifications for the domestic and export beef trade remains a major industry challenge. There is wide variation for growth and carcass attributes between breeds. Similarly, there is a high degree of variation within breeds, although management decisions are aided by comparing EBV for specific traits. The need also exists to determine growth pathways that best achieve target specifications when sires with differing potential for carcass and/or growth traits are used. Although the choice of suitable genotypes is relatively clear if the production target is total liveweight or carcass weight, fatness at given carcass weights is also a major consideration in meeting current market specifications. Furthermore, meat yield and other traits are assuming greater importance as the industry moves into value-based trading, and age at market weight is also receiving greater attention, flagging the need to grow animals as rapidly as possible to meet age-limited specifications. In this regard, supplying nutrition of sufficient quality and quantity to achieve desired growth rates requires optimal management of existing grazing systems or provision of supplements and/or improved pastures or forage crops.
Beef CRC studies have examined the effects of varying growth rates or restrictions early in life on subsequent performance. Cafe et al. (2006, 2009), Greenwood et al. (2006, 2009) and Greenwood and Cafe (2007) studied the consequences of differing growth during periods before birth and up to weaning. They concluded that, within pasture-based production systems for beef cattle (such as in temperate Australia), animals that were growth-retarded early in life showed little subsequent effects on carcass- and beef-quality characteristics during later stages of growth, although animals grown more rapidly from birth to weaning were slightly fatter than those grown slowly during the same period. Robinson et al. (2001) provided evidence that faster growth following weaning resulted in greater fat deposition at finish. Their results also indicated a small effect on finishing growth rate in favour of those animals with slower growth during backgrounding and a tendency for those with higher backgrounding growth rates to have higher intramuscular fat levels at slaughter. The experiments reported here have been designed to add to those cited above, by further examining effects following weaning and finishing. In the earlier Beef CRC studies that examined effects of steer backgrounding growth paths on subsequent growth and carcass, and eating-quality characteristics (Robinson et al. 2001; Perry and Thompson 2005), a major impact on carcass traits was the separate time of slaughter of the comparative growth treatments. Hence, the present experiment was designed to remove this potential source of confounding by having identical management of treatment groups during finishing, and slaughtering of animals on the same day.
The experiment reported here, from the NSW site, examined the effects of genetic and growth treatments, and their interaction, on liveweight and carcass traits. Groups of steers differing in genetic potential for retail beef yield (RBY%) and intramuscular fat (IMF%) (seven sire progeny groups across three carcass types) were managed by altering postweaning growth (two divergent growth rates) to reach a targeted feedlot entry-weight specification at the same time, having approximately equal mean liveweights although differing in mean age, followed by 100-day grain finish. Responses in production, carcass traits and meat quality were examined. In the present paper, we report the effects on live-animal performance during backgrounding and finishing for the NSW site. Results from the other sites (Victoria and Western Australia) are reported in associated papers (Graham et al. 2009; McIntyre et al. 2009).
Materials and methods
The overall design and methodology covering all sites in the Regional Combinations project were described by McKiernan et al. (2005). Further details specific to our experiment (NSW site) now follow. The experiment reported here differed from those at the other sites in that all animals were finished in a commercial feedlot for ~100 days following postweaning (backgrounding) growth treatments, whereas most groups at the other sites were finished at pasture or with grain feeding as part of a finishing treatment.
Location and timing
The experiment was conducted on a commercial property at Darlington Point, near Griffith (southern NSW; 34.18S, 146.02E). This region was described by Moore et al. (1970) as semiarid shrub woodlands. All matings and treatments were carried out at the home site except for two groups – one that was mated elsewhere and brought to this site in mid-pregnancy, and a second that had to be moved to another site (in the northern tablelands of NSW) with the calves before weaning because of prolonged drought conditions. There were five calvings in total during 3 years: May–June 2001, October–November 2001, May–June 2002, October–November 2002 and the last in May–June 2003. The last groups of steers completed their feedlot finishing and were processed in February 2005.
Generation of progeny
Mating programs were designed to provide groups of calves with 5–7-month difference in average birth dates. Progeny were generated by synchronised artificial insemination (AI), with either one or two cycles of mating. Synchrony programs used CIDR-based systems (EAZI-BREED CIDR, Pfizer Australia West Ryde, NSW, Australia), with device insertion for 8 days, prostaglandin injection on removal and insemination following detection of oestrus. Additional treatment with pregnant mare serum gonadotrophin was given at device removal in some programs when ovarian activity was assessed as low by ultrasound imaging. The cows used were all previously calved pure-bred Herefords of a long established self-replacing herd. Semen from the 43 different sires was allocated to cows at random on presentation for AI. The five AI programs involved 500–800 cows per mating and were conducted in August 2000, 2001 and 2002 and in January 2001 and 2002. Sire and dam were recorded at AI. Ultrasound imaging (Aloka SSD500, Medtel, Brisbane, Australia) was used for pregnancy diagnoses and to determine cycle of conception to confirm sire identity.
Calving and weaning
All progeny were identified to their dams within 24 h of birth (most within 12 h), tagged, weighed and details recorded.
Calves were weaned at averages of 226, 225, 230, 215 and 280 days (7–9 months) across the five calvings. At weaning, calves were weighed and ultrasound imaging was used for measurements of fat depth and eye muscle area (EMA) at the 12/13th rib site and fat depth at the P8 site, by using a B-mode scanner (Upton et al. 1999) Aloka SSD500 (Medtel, Brisbane, Australia) or Pye 200 (Esaote Pie Medical, Maastricht, The Netherlands).
Treatments
Sire genotypes
The 43 sires used were chosen to provide wide diversity in genetic potential in their progeny for the carcass traits of IMF% and RBY%. Sires were selected on trait EBV where available, or on performance expected as a characteristic (carcass class) of their breed. The carcass classes represented were:
-
high meat-yield class – drawn from Charolais (five sires) and Limousin (four sires) breeds, and from Angus on the basis of high EBV for RBY% (nine sires),
-
high marbling/IMF% class – drawn from the Black Wagyu breed (four sires) and from Angus on the basis of high EBV for IMF% (eight sires), and
-
high for both meat yield and marbling/IMF% class – drawn from Angus on the basis of high EBV for both traits (eight sires) and from the Red Wagyu breed (five sires).
These ‘class’ categories will subsequently be referred to as RBY, IMF and RBY&IMF. Reference to different sire ‘types’ within each carcass class relate to their being drawn from different breeds (‘Char’ for Charolais, ‘Lim’ for Limousin, Red Wagyu or Black Wagyu), or in the case of the Angus sires, different types are categorised by their high EBV for RBY% or IMF%, and subsequently referred to as ‘Angus RBY’, ‘Angus IMF’ or ‘Angus RBY&IMF’. Thus, there were seven different ‘types’ (three Angus, two Euro and two Wagyu) among the three carcass ‘classes’, and the results are presented for either or both classifications. The number of sires used for any type within a class restricted the ‘representativeness’ of the type. Hence, caution is needed in considering responses as applying to the entire breed from which the sample of sires was drawn (especially for the non-Angus types), where there were no EBV to indicate their ranking within their breed.
The choice of Angus sires was on the basis of EBV for IMF% and RBY% in BREEDPLAN (Graser et al. 2005) available at the time of sire selection (July 2000). They were generally ranked in the top 1% of the breed for the nominated trait and in the lowest 1–5% of the breed for the other trait. Sires chosen for both traits were on average in the top 5–10% for each of the traits. Where possible the sires within each group were also chosen to represent a range of growth EBV (400-day weight). A complete list of the 43 individual sires used here, as well as those at the other Regional Combinations sites, was given by McKiernan et al. (2005). Many of the sires were used across sites to provide the genetic links for combining sites in an analysis to be reported in a later paper. In total, 60% of the Angus sires, and 56% of the other sire types, were used at two or more sites.
Growth paths
All cows and calves were treated identically until weaning. Steer progeny were then randomly allocated to growth-treatment groups while maintaining a balance for sires and liveweight. There were two replicates per treatment and the steers were grown at either 0.5 or 0.7 kg/day (approximately) from weaning to feedlot entry. These will subsequently be referred to as ‘slow’ or ‘fast’ growth treatment groups. Fast- and slow-growth groups from successive calvings were managed to enter the feedlot at the same time (matched finishing cohorts), with approximately equivalent group mean liveweight, as illustrated in Fig. 1. This alignment of groups for feedlot finishing and processing was specifically designed to prevent confounding of effects on the outcomes (live and carcass data) caused by differing feedlot conditions, day of slaughter and conditions of carcass measurement.
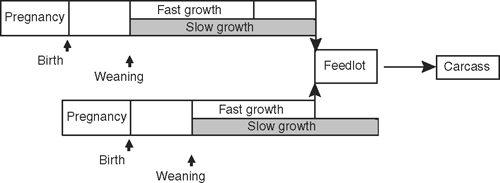
|
The slow-growth groups were grown at conventional rates while grazing native pastures typical of the district, consisting of remnant native grasses, with some introduced temperate annual legumes and grasses. However, the steers had to be supplemented often throughout the experiment to maintain desired growth rates, because of the prolonged drought conditions experienced in this region, as for much of south-eastern Australia. Rainfall was below the long-term average in 3 of the 4 years of the study. The fast-growth groups were provided with improved nutrition from periods of access to irrigated pastures of clover (Trifolium subterraneum spp.) and ryegrass (Lolium perene spp.) for most cohorts, and a dedicated area of irrigated chickory (Cichorium intybus) for one group. Because of drought, irrigation water was much less available than originally planned, which limited the faster growth rates achieved. Steers were weighed regularly (i.e. every 4–6 weeks) to monitor growth and determine adjustments to pasture availability required to achieve targeted group mean liveweights for feedlot entry. The final fast- and slow-growth groups completed their backgrounding growth treatments away from the main site, before returning for finishing at the same feedlot as other cohorts.
There were differences in the mean age of 5.1–6.8 months at feedlot induction between the slow- and fast-growth groups for separate finishing cohorts.
Feedlot finishing and processing
Following their backgrounding period, all steers in matched fast- and slow-growth finishing cohorts were despatched to a commercial feedlot as a single group when the mean overall liveweight was close to 400 kg in the paddock. This resulted in a mean of ~380-kg feedlot-induction weight (empty bodyweight), as recorded by the feedlot operators. It should be noted that calculations of liveweight gain in the feedlot were somewhat inflated, because this empty weight was used at the starting point, whereas the exit weight was taken ‘full’. However, although the apparent absolute rates of gain were affected, comparison between groups remains valid.
Prior to despatch to the feedlot, the measurements taken at weaning (as above) were repeated with the addition of an estimate of IMF% with the Pye 200 ultrasound scanner.
All steers were weighed and given health treatments at induction, according to the standard procedures for the feedlot. These included growth-promotant implants (Revalor, Intervet Australia, Bendigo East, Vic., Australia), internal and external parasite treatments and vaccinations for prevention of clostridial and respiratory infections. In most cases, the entire group was finished in a single feedlot pen; however, where two pens were used, animals were randomly allocated although ensuring equal representation of treatment groups. All animals had identical treatment (feeding and management) during the ~100-day feedlot finishing phase before slaughter (range: 99–116 days for different cohorts). Steers were weighed (full) at exit from the feedlot before despatch to the abattoir. Data were collected at the abattoir by routine procedures as described by McKiernan et al. (2005). Further detail of abattoir processing and subsequent measurements will be given in the papers to follow which report results on carcass traits and meat quality from this experiment (McKiernan et al. 2009; J. F. Wilkins, W. A. McKiernan, J. Irwin, B. Orchard and S. A. Barwick, unpubl. data).
Target market
The production system examined was a cross-breeding enterprise providing a ‘feeder steer’ for feedlot finish, and subsequently producing a 100-day grain-fed product suitable for high-quality domestic and export markets. The feedlot entry specification stated a preferred weight range of 380–440 kg and no permanent teeth, so a group mean liveweight of 400 kg was targeted. All steers were accepted by the feedlot operator to accommodate the project requirements, although a few were outside the weight specification.
Statistical analyses
Analyses of effects on production and carcass traits were performed with the PC Genstat software package (Release 9.1 for Windows XP; VSN International, Hemel Hempstead, UK), using a linear mixed model REML procedure. The procedure involved iterative runs to refine the initial model, eliminating random terms showing negative (or aliased) variance components, and by removing non-significant (P > 0.05) fixed effects. The final model was then used to evaluate treatment effects and generate predicted means.
The initial model included the fixed effects of growth treatment, season of birth, (carcass) ‘class’ of sire (as above), ‘type’ of sire (within class) and all interactions. The random effects included sire (individual sires within type), dam, kill (different slaughter events), replicate (within growth treatments), year of birth and all interactions. ‘Pen’ was also included as a random effect on feedlot-performance measurements, as applicable. An ‘animal pedigree’ matrix to account for the association between individuals through common sire and/or dam was also included within the random-effects structure. ‘Class’ as a main effect was deleted when generating sire ‘type’ means. Age at the start of the treatment was included as a covariate in all analyses, and was often not significant. Carcass traits such as P8 and rib fat depths and EMA can be affected by the size of the carcass, so the model needed to account for differences in HSCW to detect effects independent of response in HSCW (growth-rate response). Therefore, HSCW was included as a covariate for all carcass traits and was significant in most cases. Performance in the feedlot was affected by liveweight at entry. Because the mean entry weights differed between the fast- and slow-growth groups, this was included as a covariate for feedlot growth rate and exit weight. Any effects owing to changes in location (mentioned above) were included in the random variation within the analysis model.
Rates of calving difficulty were compared by chi-square analyses. Where correlations were examined, they were calculated with the Genstat software (Release 9.1 for Windows XP).
Significance of effects in results and discussion assumes P < 0.05 unless otherwise stated.
Results
Performance before and during growth treatment
As background information on the progeny, the variation across genotypes in gestation length and birthweight and the subsequent weaning weights is shown in Table 1. There were significant differences owing to sire type for gestation length, birth and weaning weights, with the European types having larger values for all parameters. There were also differences owing to sex (not tabulated), with males significantly heavier than females at both birth (35.6 v. 33.6 kg; 6% heavier) and weaning (215 v. 205 kg; 5% heavier). Calving difficulties were few, with a total of only 11 cases of assistance in 1307 births (<1%). However, there were more assisted births with Charolais-sired calves (6/108 births) than any other breed type (ranging from 0/235 to 2/232 births), and this was associated with a single sire with a high EBV for birthweight. The rate of calving difficulty for the Charolais-sired progeny was significantly (P < 0.05) higher than that for the Angus types and Red Wagyu, whereas it showed no difference (P > 0.05) from the Black Wagyu- and Limousin-sired calves.

|
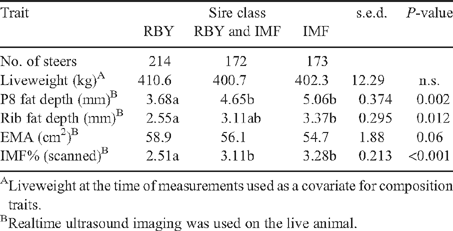
Differences in predicted means for measurements of liveweight and body composition at the end of the growth treatment owing to backgrounding growth rate and sire type are shown in Table 2 and the effects of sire class are shown in Table 3. The average age at the time of live measurement shown in Tables 2 and 3 across all fast-growth groups was 478 days and 636 for the slow-growth groups; however, there were considerable ranges between kills, namely 404–523 days for the fast-growth and 568–687 for the slow-growth groups.

|
|
Sire-type effects
There were significant effects of sire type on all body-composition measurements at the start and the finish of the growth treatment, before feedlot entry (Tables 1 and 2). For the ‘fatness’ traits of fat depth at P8 and rib sites and IMF%, the Limousin and Charolais groups ranked lowest in every case for both fast- and slow-growth treatments, and the Angus RBY groups were the next lowest in most cases. The Angus IMF and Black Wagyu groups were the most consistently highly ranked in the slow-growth treatment. The Angus IMF group was again the most consistently highest in the fast-growth treatment.
Differences in scanned eye muscle area were as expected, with the potentially higher-yielding European types having significantly greater EMA than all other types within both growth-treatment groups.
There were significant differences among classes in P8 and rib fat depths and IMF%, all in accordance with their basis for selection. The trend in the ranking for EMA was as expected although the differences owing to class were not quite statistically significant for this trait (P = 0.06, Table 3).
Growth-treatment effects
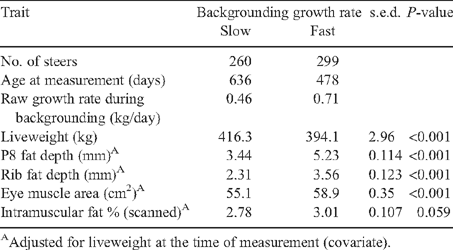
Fast-growth treatments had higher values in all traits for every sire type, except for scanned IMF% in the Black Wagyu, which was not significantly different between the growth treatments (Table 2).
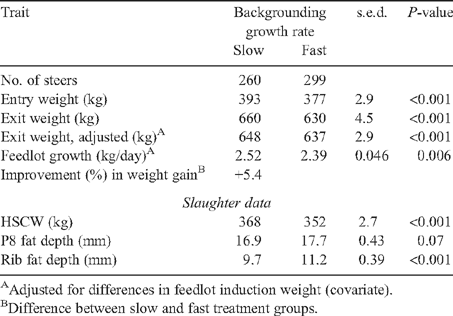
Management was aimed at achieving near-equivalent group means at feedlot entry; however, there was a significant difference of 22 kg in the paddock before entry (16 kg at feedlot induction) in favour of the slow-growth groups. This was taken into account in comparing the live-carcass measurements before feedlot entry (Tables 2–4), with the liveweight at the time as a covariate. Liveweight was also taken into account when considering the performance in the feedlot, with the feedlot entry weight as a covariate (Tables 5–7). Fast-growth groups had significantly (P < 0.001) higher predicted means than did slow-growth groups (Table 4) for P8 and rib fat depths and EMA, with a strong trend in the same direction for the scanned IMF% (P = 0.06).
|

|
|
|
Treatment interactions
There were some inconsistencies in rankings of sire types across growth-treatment groups for P8 and rib fat depths in the Red and Black Wagyu and Angus (RBY&IMF) groups. This resulted in a significant interaction for P8 fat (Table 1).
Performance during feedlot finishing
The predicted means for feedlot performance, as affected by sire type and growth treatments, are shown in Tables 5 and 7, with the effects of sire class shown in Table 6. The results for the carcass P8 and rib fat depths collected at slaughter are also presented here to show the effect of growth during the feedlot phase on fatness. However, the main effects of treatments on carcass traits are fully discussed in a companion paper (McKiernan et al. 2009).
Sire-type effects
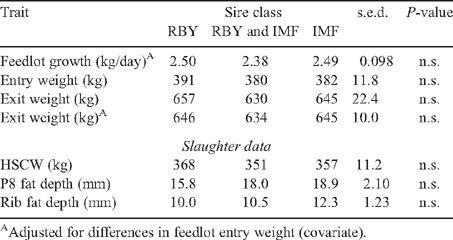
There were significant differences between sire-type progeny groups for feedlot entry (induction) weight. The Charolais progeny were significantly heavier than others whereas both the Black and Red Wagyu were significantly lighter.
Differences among sire types in feedlot performance – daily gain and exit liveweight – were affected by induction (entry) liveweight, and the differences were reduced when this was included as a covariate. The major difference among groups was the significantly (P < 0.001) poorer performance of the Red Wagyu group than that of all others.
Differences in entry weight and feedlot performance among class of sire groups were not significant (Table 6).
Growth-treatment effects
The groups having slower backgrounding growth grew significantly faster in the feedlot than those with rapid backgrounding growth (Table 7), thus exhibiting compensatory growth (5.4% faster than for the fast-backgrounded group). This effect was evident in progeny groups of all sire types, and ranged from a 1.7% to an 8.2% increase in growth (Table 5). Within sire-progeny groups, there was good association between the size of the differences between the slow- and fast-growth groups (compensatory advantage) and the mean growth rates for the type (raw correlation ~0.7). Thus, steers of types with a greater propensity for growth showed a greater compensatory advantage.
Treatment interactions
There were no significant interactions between growth rate and genetic treatments for performance in the feedlot (Table 5).
Discussion
Results discussed here focus on effects during the period of growth for the animals in the present experiment. Effects on traits in the carcass as the endpoint are examined in a companion paper to follow (McKiernan et al. 2009).
Effects of growth treatments
Faster liveweight gain in the feedlot, following slower backgrounding growth, was a significant and very important practical outcome of this experiment. Although the growth rates of the animals in the slow-growth treatments here would not be considered particularly restricted in comparison with experiments designed specifically to examine compensatory growth, the effect was clear. This field of research was well reviewed by Ryan (1990), who concluded that a reduction in the maintenance requirement, increased efficiency of growth and fattening, a reduction in the energy of tissue deposited and an increase in feed intake all contributed to compensatory growth. This was supported by experiments showing that the initial stages of compensatory growth involved greater deposition of protein during recovery of depleted gut and other tissues, which was then followed by the reverse situation of a greater fat to protein ratio during the period of increased intake and faster growth (Ryan et al. 1993a, 1993b). It is the latter phase that is analogous to our situation because growth would not have been restricted to the point of tissue depletion in our experiments. We were not able to measure feed intake in the present experiment although it is likely that steers growing more rapidly were also more efficient, owing to a greater ratio of liveweight gain to the ‘overhead cost’ of maintenance, a significant factor in the economics of feedlot finishing.
An important effect of growth rate seen here was the increase in fatness associated with faster growth, supporting previously reported results from Beef CRC studies (Robinson et al. 2001; Cafe et al. 2006; Greenwood et al. 2006). Although the final fatness in the carcass was greater in the fast-backgrounded groups, the amount of fat deposited during the feedlot phase was greater in the slow-backgrounded groups in association with their compensatory growth, an increase of ~8% for P8 and 3% for rib fat (Tables 4 and 7). Effects on fatness are of obvious practical importance to meeting carcass specifications when targeting specific market end points. The findings here of greater fatness associated with faster growth are consistent with the results reported by Graham et al. (2009) from the Victorian site of the Regional Combination experiment, and by McIntyre et al. (2009) from the Western Australian site. Graham et al. (2009) also cited further supporting evidence from previous studies, showing higher planes of nutrition resulting in increased proportions of fat in the carcass (Berg and Butterfield 1968; Coleman et al. 1993).
Robinson et al. (2001) demonstrated a small effect on the finishing growth rate in favour of animals having slower growth during backgrounding and a tendency for those with higher backgrounding growth rates to have higher intramuscular fat levels. This supports the current findings, where both of these effects were clearly demonstrated. The results here add to the previous studies of effects during pregnancy and lactation (i.e. conception to weaning) reported by Greenwood et al. (2006), and during backgrounding reported by Robinson et al. (2001), in understanding the effects of growth rate at various stages of development on the final outcome measured as carcass traits. This knowledge is vital for providing recommendations to producers that predict consequences of growth restrictions, as well as how best to reach targeted end points. Greenwood and Cafe (2007) concluded that within pasture-based production systems for beef cattle, animals that were growth-retarded early in life showed little residual effect in later stages of growth. They found that cattle grown slowly to weaning had similar composition at slaughter to those grown more rapidly when backgrounded on pasture to the same feedlot entry weight and achieving similar final carcass weights, although at an older age. Similarly, marbling in the carcass was not significantly affected by the preweaning growth rate.
Effects of differing genetic potential
The slow-backgrounded groups grew on average 5.4% faster overall in the feedlot than the fast-backgrounded groups. However, when the prefeedlot growth treatments were evaluated within sire types, those with a propensity for faster growth displayed more compensatory growth in the feedlot. The fastest growing types, Charolais and Limousin, displayed levels of compensation of 8.2% and 7.2%, respectively, whereas the slower growing Red and Black Wagyu progeny displayed growth compensation of only 3.2% and 1.7%, respectively. This suggests that the degree of compensation was related to growth potential when restrictions were removed. The difference in the mean growth rate between the fast- and slow-growth treatment groups for each type (the compensatory effect) was well related to the mean growth rate for the type (raw correlation of ~0.7).
There is a large body of information in the literature from the Meat Animal Research Center, USA (e.g. Gregory et al. 1978, 1994; Cundiff 2006) showing the variation among breeds and various types of cattle (‘biological types’) for growth and carcass attributes. Variation among breeds and genetic parameters for growth and body development for a diverse range of genotypes studied in Australia were reported by Afolayan et al. (2007). Mean differences in the performance of progeny groups from designed matings can be predicted by the use of trait EBV as generated by BREEDPLAN (Graser et al. 2005). Thus, variation in genetic potential for growth and carcass traits is well documented; however, the prediction of the responses of different genotypes to varying growth scenarios remains an important objective of future research.
Although there were only few sires from some breeds used here, the data on traits at birth, weaning and growth rates in their progeny are consistent with performance previously reported for those breeds in both Australian and overseas studies (e.g. Gregory et al. 1978, 1994; Afolayan et al. 2007). Thus, although some caution must be maintained, we can be reasonably confident that the samples of progeny we examined from the various carcass types are not atypical of the breeds from which their sires were drawn, a similar conclusion to that of Graham et al. (2009) who reported the results of the Victorian site of this project. There is little information on the Red Wagyu breed, although there is some anecdotal support for its potential as a dual-purpose type for yield and marbling. Although there were few sires only from this breed, the progeny had the lowest growth rates, which would be a major concern. The carcass traits in relation to the other sire types are discussed in the companion paper (McKiernan et al. 2009), and we suggest more sires need to be sampled to provide a fair assessment of this breed type.
The grouping of sires by ‘class’ produced effects in reasonable accordance with expectations. However, such grouping may also mask important differences among types that need to be identified. Thus, results of type- and class-based analyses need to be assessed in combination when making conclusions. A case in point was the large differences in weight and weight gain between the Charolais and the other types (consistent with that reported by Gregory et al. 1978), and further examples are evident for the carcass traits (McKiernan et al. 2009).
The overall effects of type on fatness were in accordance with expectations on the basis of genetic potential for fat deposition.
Genetic × growth treatment interactions
The only significant interaction in the analyses was caused by a slight reranking of types within growth treatments for P8 fat before feedlot entry. However, the effect on both P8 and rib fat depths favouring faster growth was entirely consistent across types, and the interaction for P8 fat depth did not persist in the carcass data (McKiernan et al. 2009).
Conclusions
The effect of faster growth resulting in greater fatness was a clear result from the present experiment, evident in both the backgrounding phase in the paddock and in the subsequent feedlot finishing. Although the slower-growth treatment after weaning would not be considered a severe restriction, compensatory growth was substantial, and evident across all sire types, indicating that feedlot operators who wish to take advantage of superior growth rates and efficiency should consider selecting animals of the same weight but lower fatness at feedlot entry.
If reaching a minimum specification for fatness at the end of feedlot finishing is critical, types with a low genetic propensity to fatten will need to be treated differently from those of opposite potential. Although slow growth during backgrounding may be well compensated during finishing, it may be detrimental to eventual fatness and cause a failure to meet the specification. Thus, faster growth before feedlot and longer periods on feed may be more suitable for these types, or alternatively they may best be targeted for markets where fatness levels are less critical. Effects on subcutaneous fat were evident in the results reported here, and these are further discussed, along with effects of potentially greater importance on intramuscular fat in the companion paper (McKiernan et al. 2009).
Accurate prediction of the end product will be facilitated by the application of data generated here and elsewhere to growth models currently being refined to provide management-aid tools for use by the industry.
Acknowledgements
We are pleased to acknowledge the large contribution to the project by the support of our commercial cooperator, AgReserves Australia– sincere thanks go to all staff and management at ‘Bringagee’ and ‘Kooba’ for their enthusiastic involvement (Tony Abel and Angus Paterson, in particular). We are also grateful for the cooperation and assistance of Cargill Beef Australia (Harry Waddington and Grant Garey, in particular) at the feedlot finishing (‘Jindalee’, near Temora, NSW) and processing stages (Cargill works, Wagga Wagga, NSW). Special thanks go to Matt Wolcott for his expert assistance with ultrasound measurements, to Diana Perry and staff at the Meat Science Laboratory (Armidale), and to Jeffrey House and Greg Meaker for their valuable inputs in field data collection. The authors were supported by the NSW Department of Primary Industries, the CRC for Cattle and Beef Quality and by Meat and Livestock Australia. Many thanks go to all the field- and administrative-support staff of NSW DPI who assisted with this large experiment and to all our colleagues within the Beef CRC and NSW DPI who provided advice throughout. Finally, we gratefully acknowledge the inputs of Dr Jim Walkley for his helpful advice in the drafting and revision of this paper.
Afolayan RA,
Pitchford WS,
Deland MPB, McKiernan WA
(2007) Breed variation and genetic parameters for growth and body development in diverse beef cattle genotypes. Animal 1, 13–20.
| Crossref | GoogleScholarGoogle Scholar |

Berg RT, Butterfield RM
(1968) Growth patterns of bovine muscle, fat and bone. Journal of Animal Science 27, 611–619.
Bindon BM
(2001) Genesis of the Cooperative Research Centre for the Cattle and Beef Industry: integration of resources for beef quality research (1993–2000). Australian Journal of Experimental Agriculture 41, 843–853.
| Crossref | GoogleScholarGoogle Scholar |
Burrow HM, Bindon BM
(2005) Genetics research in the Cooperative Research Centre for Cattle and Beef Quality. Australian Journal of Experimental Agriculture 45, 941–957.
| Crossref | GoogleScholarGoogle Scholar |
Cafe LM,
Hearnshaw H,
Hennessy DW, Greenwood PL
(2006) Growth and carcass characteristics of Wagyu-sired steers at heavy market weights following slow or rapid growth to weaning. Australian Journal of Experimental Agriculture 46, 951–956.
| Crossref | GoogleScholarGoogle Scholar |
Cafe LM,
Hennessy DW,
Hearnshaw H,
Morris SG, Greenwood PL
(2009) Consequences of prenatal and preweaning growth for feedlot growth, intake and efficiency of Piedmontese- and Wagyu-sired cattle. Animal Production Science 49, 461–467.
| Crossref | GoogleScholarGoogle Scholar |
Coleman SW,
Evans BC, Guenthert JJ
(1993) Body and carcase composition of Angus and Charolais Steers as affected by age and nutrition. Journal of Animal Science 71, 86–95.
|
CAS |
PubMed |
Graham JF,
Byron J, Deland MPB
(2005) Effect on liveweight and carcass traits of progeny sired by bulls selected for extremes in intramuscular fat and retail beef yield and the effect of post weaning growth. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 16, 338–341.
Graham JF,
Byron J,
Clark AJ,
Kearney G, Orchard B
(2009) Effect of postweaning growth and bulls selected for extremes in retail beef yield and intramuscular fat on progeny liveweight and carcass traits. Animal Production Science 49, 493–503.
| Crossref | GoogleScholarGoogle Scholar |
Graser H-U,
Tier B,
Johnston DJ, Barwick SA
(2005) Genetic evaluation for the beef industry in Australia. Australian Journal of Experimental Agriculture 45, 913–921.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL, Cafe LM
(2007) Prenatal and pre-weaning growth and nutrition of cattle: long-term consequences for beef production. Animal 1, 1283–1296.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL,
Cafe LM,
Hearnshaw H,
Hennessy DW,
Thompson JM, Morris SG
(2006) Long-term consequences of birth weight and growth to weaning on carcass, yield and beef quality characteristics of Piedmontese- and Wagyu-sired cattle. Australian Journal of Experimental Agriculture 46, 257–269.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL,
Cafe LM,
Hearnshaw H,
Hennessy DW, Morris SG
(2009) Consequences of prenatal and preweaning growth for yield of beef primal cuts from 30-month-old Piedmontese- and Wagyu-sired cattle. Animal Production Science 49, 468–478.
| Crossref | GoogleScholarGoogle Scholar |
Gregory KE,
Cundiff LV,
Smith GM,
Laster DB, Fitzhugh HA
(1978) Characterization of biological types of cattle–Cycle II: I. Birth and weaning traits. Journal of Animal Science 47, 1022–1030.
Gregory KE,
Cundiff LV,
Koch RM,
Dikeman ME, Koohmaraie M
(1994) Breed effects, retained heterosis, and estimates of genetic and phenotypic parameters for carcass and meat traits of beef cattle. Journal of Animal Science 72, 1174–1183.
|
CAS |
PubMed |
Irwin J,
Wilkins JF,
McKiernan WA, Barwick SA
(2006) Relationships between carcass measures and estimates of meat quality traits in steers with diverse genotypes and having different pre-feedlot growth treatments. Animal Production in Australia Short Communication Number 19. 26,
|
CAS |
McIntyre BL,
Tudor GD,
Read D,
Smart W,
Della Bosca TJ,
Speijers EJ, Orchard B
(2009) Effects of growth path, sire type, calving time and sex on growth and carcass characteristics of beef cattle in the agricultural area of Western Australia. Animal Production Science 49, 504–514.
| Crossref | GoogleScholarGoogle Scholar |
McKiernan WA,
Wilkins JF,
Barwick SA,
Tudor GD,
McIntyre BL,
Graham JG,
Deland MPD, Davies L
(2005) CRC ‘Regional Combinations’ Project – effects of genetics and growth paths on beef production and meat quality: experimental design, methods and measurements. Australian Journal of Experimental Agriculture 45, 959–969.
| Crossref | GoogleScholarGoogle Scholar |
McKiernan WA,
Wilkins JF,
Irwin J,
Orchard B, Barwick SA
(2009) Performance of steer progeny of sires differing in genetic potential for fatness and meat yield following postweaning growth at different rates. 2. Carcass traits. Animal Production Science 49, 525–534.
| Crossref | GoogleScholarGoogle Scholar |
Perry D, Thompson JM
(2005) The effect of growth rate during backgrounding and finishing on meat quality traits in beef cattle. Meat Science 69, 691–702.
| Crossref | GoogleScholarGoogle Scholar |
Polkinghorne R,
Thompson JM,
Watson R,
Gee A, Porter M
(2008) Evolution of the Meat Standards Australia (MSA) beef grading system. Australian Journal of Experimental Agriculture 48, 1351–1359.
| Crossref | GoogleScholarGoogle Scholar |
Robinson DL,
Oddy VH,
Dicker RW, McPhee M
(2001) Post-weaning growth in northern New South Wales. 3. Carry-over effects on finishing, carcass characteristics and intramuscular fat. Australian Journal of Experimental Agriculture 41, 1041–1049.
| Crossref | GoogleScholarGoogle Scholar |
Ryan WJ
(1990) Compensatory growth in sheep and cattle. Nutrition Abstracts and Reviews 60, 653–664.
Ryan WJ,
Williams IH, Moir RJ
(1993a) Compensatory growth in sheep and cattle. 1. Growth pattern and feed intake. Australian Journal of Agricultural Research 44, 1609–1621.
| Crossref | GoogleScholarGoogle Scholar |
Ryan WJ,
Williams IH, Moir RJ
(1993b) Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights. Australian Journal of Agricultural Research 44, 1623–1633.
| Crossref | GoogleScholarGoogle Scholar |
Tudor GD,
Della Bosca TJ,
McIntyre BL,
Read D,
Smart WL,
Taylor EG, Hiskock AJF
(2004) Optimising feed supply, reproductive efficiency and progeny growth to meet market specifications. 1. Background and experimental design. Animal Production in Australia 25, 334.
Upton WH,
Donoghue KA,
Graser H-U, Johnston DJ
(1999) Ultrasound proficiency testing. In Proceedings of the Association for the Advancement of Animal Breeding and Genetics 13, 341–344.
Upton W,
Burrow HM,
Dundon A,
Robinson DL, Farrell EB
(2001) CRC breeding program design, measurements and database: methods that underpin CRC research results. Australian Journal of Experimental Agriculture 41, 943–952.
| Crossref | GoogleScholarGoogle Scholar |
Wilkins JF,
Irwin J,
McKiernan WA, Barwick SA
(2002) Combining genetics and growth rate to produce high quality beef in south eastern Australia. Animal Production in Australia 24, 370.
Wilkins JF,
Irwin J,
McKiernan WA, Barwick SA
(2004) Effects of altering growth rate on carcass traits in a range of beef genotypes that vary in potential for yield and fat deposition. Animal Production in Australia 25, 337.