

Factors influencing the variability in performance of cattle grazing tropical pasture
E. Charmley A * , G. J. Bishop-Hurley B , C. S. McSweeney B , R. Takeuchi B , G. Martinez-Fernandez B and S. Denman B
A * , G. J. Bishop-Hurley B , C. S. McSweeney B , R. Takeuchi B , G. Martinez-Fernandez B and S. Denman B
A
B
Abstract
Whereas it is known that there is variation in liveweight gain (described as performance in this paper) within a group of grazing cattle, the causes of this variation are not well understood.
A 2-year grazing study in northern Queensland was conducted to understand the relative influence of diet selection, grazing behaviour and rumen fermentation on variation in performance of growing cattle.
Eighty-nine Bos taurus × B. indicus (~6 months old) growing steers were grazed as a group on mixed tropical pasture for 23 months. Pasture was characterised for biomass, species and nutrient composition. Cattle were ranked according to overall liveweight (LW) gain and the top (high performance, HP) 20 and bottom (low performance, LP) 20 head were compared for diet composition, rumen fermentation and grazing behaviours.
Pasture biomass, and nutritive value of pasture and diet varied between seasons and years of study. The HP cattle achieved overall LW gains 20% greater than those of the LP cattle mainly due to higher rates of gain in the wet season. However, the nutritive value and rumen fermentation characteristics were similar for both LP and HP cattle, although there was evidence that crude protein and digestibility were modestly higher in diets of HP cattle, especially in the first wet season. Activity, measured as distance travelled, declined as pasture biomass declined, and cattle appeared to favour pasture with higher legume content.
It is concluded that divergence in performance of cattle on tropical pasture could not be fully explained by measurements taken in this study. Diet selection for plant components with improved nutritive value probably played a small role. Differences in initial LW between the LP and HP groups may have been a contributory factor.
Despite comprehensive measurements of factors that influence efficiency and performance on pasture, we were unable to identify definitive causes. Genetic variation in feed intake or efficiency of digestion needs to be investigated.
Keywords: animal performance, beef cattle, feed efficiency, grazing behaviour, intake, nutritive value, pasture, tropical.
Introduction
The northern Australian beef industry is pasture-based, with more than 95% of feed requirements being derived from this resource (Bell and Sangster 2023). Typically, pasture productivity is highly seasonal, with growth occurring in the wet season from approximately December to May, depending on the year. Linked to this seasonality are large changes in nutritive value and performance of grazing cattle (Tothill and Gillies 1992). Being dominated with C4 grasses, diet nutritive value is never high compared with temperate regions and declines from a peak of approximately 65% dry-matter (DM) digestibility in the wet season to a nadir of 40% at the end of the dry season (Poppi and McLennan 1995; Coates 1996). Likewise, crude protein (CP) values follow a similar pattern, ranging between 12% and 4% DM. Nutritive value, particularly CP can be improved when legumes such as Stylosanthes (Cooksley 2003), Desmanthus (Mwangi et al. 2021) or leucaena (Petty and Poppi 2012) are included in the predominantly grass-based swards. Whereas the above generalities define the north, there are also regional variations roughly aligned to rainfall and latitude. Animal performance follows this seasonal cycle with fluctuations in liveweight (LW) change and body condition (Chilcott et al. 2020).
Within this seasonal pattern of performance, there are also differences in performance among individuals within a grazing cohort that can readily be observed in terms of LW change (Silva et al. 2022) or reproductive efficiency (McCosker et al. 2023). However, it is difficult to accurately measure DM intake of grazing cattle, hindering the ability to ascribe causes to these differences in performance.
Understanding feed efficiency is essential for selection of more efficient individuals and for accurate feed budgeting to achieve targeted performance. Residual feed intake (RFI), defined as the difference between the observed and expected amount of feed consumed to achieve a given level of performance, has become a popular metric for assessing feed efficiency (Arthur et al. 2004). Differences in RFI within a cohort of animals receiving the same feed have been ascribed to feeding behaviour, digestion, the rumen microbiota, energy metabolism, protein turnover, and variation in body composition (Cantalapiedra-Hijar et al. 2018). Of these, some, such as feeding behaviour, digestion and the rumen microbiome, are influenced by DM intake (DMI) rather than the genotype per se, whereas energy metabolism and protein turnover appear to be independent of DMI and directly influence RFI (Cantalapiedra-Hijar et al. 2018).
For the grazing animal, the situation is further complicated because grazing behaviour can influence DMI and the nutritive value of ingested DM through diet selection. This strong genetics by environment (G × E) interaction is poorly understood, owing to the difficulties in accurate measurement of grazing behaviour, diet selection and voluntary intake. Animals ranked for RFI under confined feeding may not rank the same when grazing (Lawrence et al. 2013; Kenny et al. 2018). The purpose of this trial was to rank animals for measured LW gain post hoc and relate performance to various measures of grazing behaviour, diet selection and digestion, so as to ascertain key drivers for efficiency under northern Queensland grazing conditions.
Materials and methods
Site
A 276-ha area, comprising three adjoining and interconnected paddocks, was selected for the trial at the Lansdown Research Station (19°39′S, 146°50′E). The site comprised areas of cleared and open woodland with riparian areas. The naturalised pasture was dominated by the grasses Indian couch (Bothriochloa pertusa), black speargrass (Heteropogon contortus), and Queensland bluegrass (Dicanthium sericium), and by the legumes seca stylo (Stylosanthes scabra) and verano stylo (S. hamata). Soils were predominantly loam and sandy loam and the annual rainfall for Lansdown is 902 mm.
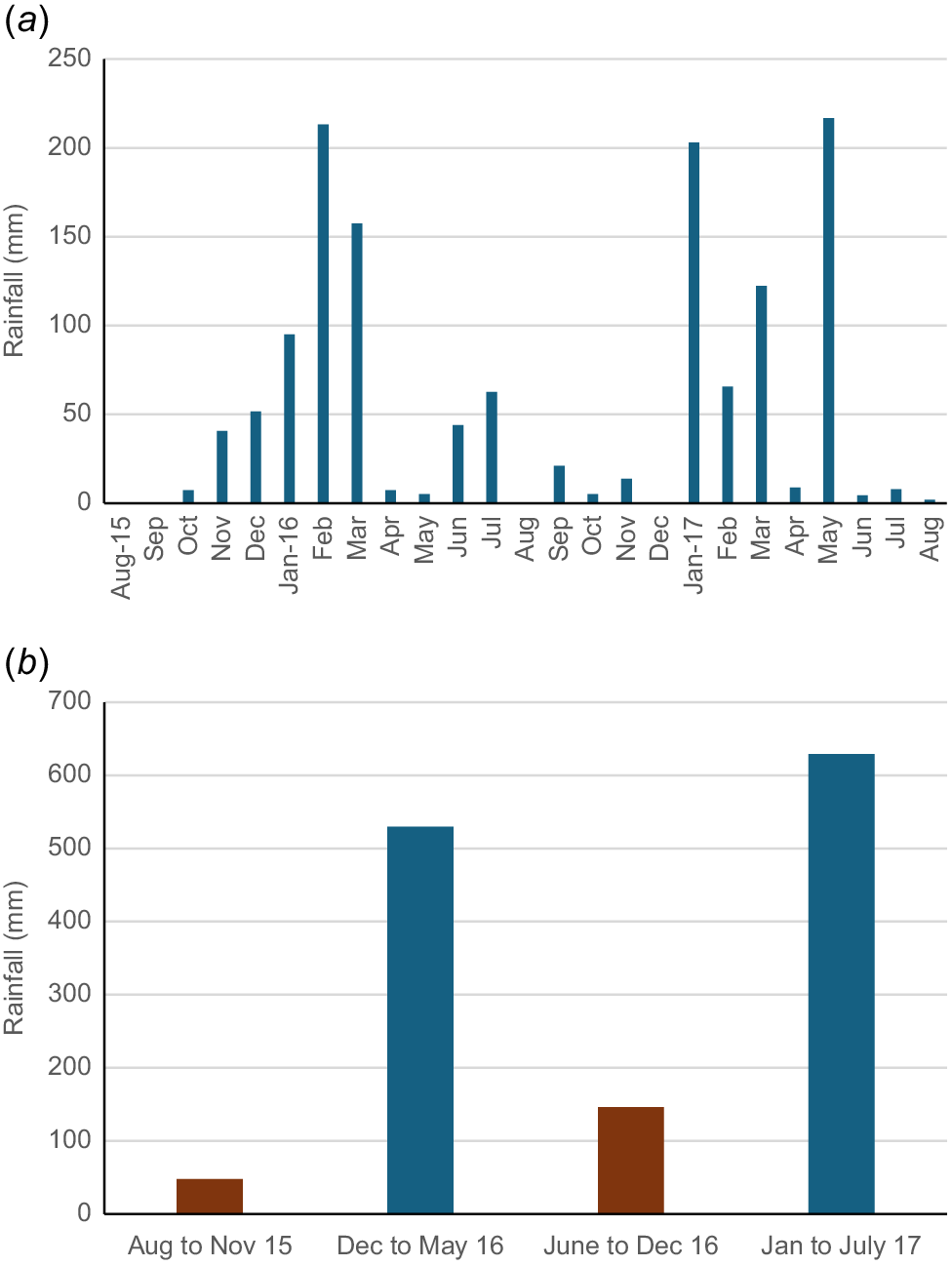
The trial site was set stocked for 23 months, encompassing two dry seasons and two wet seasons. Rainfall patterns corresponding to dry and wet seasons are given in Fig. 1. Three water points were provided
Animals and treatments
Ninety Droughtmaster steers (Bos indicus × B. taurus) with an average age of ~6 months were purchased for the trial locally, and quarantined for 10 days before allocating to the trial on 28 August 2015 with a LW (mean ± s.d.) of 188 ± 30 kg. Cattle were set-stocked on the area for 698 days, at which time they were sent for slaughter on 26 July 2017 at an abattoir 50 km from the trial site and carcase data from the abattoir were obtained for all animals. Cattle were mustered at pre-defined periods (Table 1) for measurement of LW and sampling of rumen fluid, blood and faeces by using industry-standard cattle yards and crush. One animal died during the trial of unknown causes. The LW (mean ± s.d.) at the end of the trial was 554 ± 47 kg. The trial was approved for animal ethics under the Brisbane Animal Ethics Committee Application A 12 2015.
| Season | Botanal pasture assessment | Pasture sampling | Rumen and faecal sampling | GPS collar deployments | |
|---|---|---|---|---|---|
| Dry season 2015 | 1/12/2015 | 1/12/2015 | 1/12/2015 | ||
| Wet season 2016 | 1/03/2016 | 29/02/2016 | 29/02/2016 | 7/03–7/04/2016 | |
| Wet season 2016 | 3/05/2016 | 2/05/2016 | 2/05/2016 | ||
| Transition 2016 | 3/06–5/07/2016 | ||||
| Dry season 2016 | 10/08/2016 | 9/08/2016 | 9/08/2016 | 11/08–8/09/2016 | |
| Dry season 2016 | 23/11/2016 | 6/12/2016 | 6/12/2016 | 8/12–21/2/2017 | |
| Wet season 2017 | 22/02/2017 | 21/02/2017 | 21/02/2017 | ||
| Wet season 2017 | 1/06/2017 | 8/06/2017 | 1/06/2017 |
Dates are shown as DD/MM/YY.
Animals were assigned to the following two groups post hoc according to their overall LW gain: low performance (LP, 20 animals with the lowest LW gain), and high performance (HP, 20 animals with the highest LW gain). The remaining 48 animals were excluded from the statistical analysis of animal variables. Animal and pasture variables were grouped according to season, resulting in a statistical comparison including high versus low performance and wet versus dry season and their interaction. The years of study were defined by rainfall patterns, with Year 1 being from August 2015 to June 2016 and Year 2 from June 2016 to July 2017. Data from each year were treated independently and analysed separately.
Sampling
Cattle were weighed at the beginning, end, and every 2–3 months during the trial at a cattle-handling facility ~1 km from the grazing area. All weights were non-curfewed and recorded within 4 h of mustering. Concurrently with the weight sessions, all animals were sampled for blood via jugular venipuncture, rumen fluid by using an oral stomach tube and faeces by rectal grab sampling. The animals were restrained in a commercial cattle crush with the animal’s head secured in a head bail. For rumen sampling a reinforced plastic suction tube was inserted into the mouth through a protective black polypropylene pipe that was passed over the epiglottis and into the oesophagus before entering the rumen. Approximately a volume of 100–200 mL of rumen fluid was removed via the tube by using negative pressure from a hand-operated pump. The sample was immediately subdivided for pH determination (at the time of sampling by using a portable pH meter), and preservation and storage at −20°C in 20% metaphosphoric acid for volatile fatty acid (VFA) and ammonia-N analysis. A faecal sample for faecal near-infrared (NIR) analysis (100–200 g) was collected by rectal palpation and stored at −20° C within 2 h. A 10 mL blood sample was obtained from the jugular vein and held at room temperature for 30 min before storing on ice for 1.5 h. Samples were then centrifuged at 5°C for 20 min at 4000g. Plasma was analysed for ammonia-N.
Animal behaviour
Up to 12 animals were randomly selected to be fitted with Commonwealth Scientific and Industrial Research Organisation (CSIRO) designed collars (Arablouei et al. 2023) for determination of distance travelled and spatial distribution throughout the grazing area. These collars capture and process sensor data. The spatial distribution of cattle was collected using global positioning system (GPS) technology at 1 s intervals. GPS units were successfully fitted to these animals on three occasions, namely 3/6/2016 to 5/7/2016 (early-dry), 11/8/2016 to 8/9/2016 (mid-dry) and 8/12/2016 to 23/12/2016 (late-dry). The collar included an artificial intelligence of things (AIoT) device (CSIRO, Pullenvale, Australia), battery pack and six photovoltaic modules for harvesting solar energy. The AIoT device featured a Texas Instruments CC2650F128 system-on-chip with an Arm Cortex-M3 CPU at 48 MHz stored at random access memory (RAM) and read-only memory (ROM). It also included a MPU9250 9-axis micro-electro-mechanical (MEMS) inertial measurement unit containing a tri-axial accelerometer sensor to measure acceleration in three axes. The power was supplied by lithium-ion battery pack charged by solar panels. The collars were fitted around the animal’s neck with a strap. The GPS unit was held on top of the neck by means of a counter-balance underneath the neck. As a result of collar failure, data were successfully retrieved from six, seven and five animals in the early-, mid-, and late-dry periods respectively. Collar failure was attributed to moisture entering the electronics housing, collars rotating through 180°, leading to signal failure and loss of collars. Data for analysis were used only on days when all selected collars were generating continuous 1 s fix data.
After each study period, data were downloaded from micro-SD cards on collars and entered and stored in CSV files. GPS data were sorted on Rstudio (Version 2023.06.2). Data were filtered to eliminate points geolocated with less than three satellites and where distance travelled >3.3 m/s. Daily mean distances were included in the analysis when >99% of potential GPS fixes (86,400) were successfully achieved. Daily distances travelled were calculated from fixes every 10 and 30 s to reduce overestimation owing to the frequency of measurement error (McGavin et al. 2018). After filtering, six, seven and five animals were selected from early-dry, mid-dry, late-dry periods respectively. GPS points were imported to QGIS (3.34.3) for visualising preferential use of the grazing area. Preference ratios were calculated as the proportion of GPS points in an area divided by proportion of that area in the total area (Hobbs and Bowden 1982; Tomkins and O’Reagain 2007). In this case, areas were within 200 m of a waterpoint or one of three interconnected paddocks.
Pasture evaluation
Pasture biomass, legume proportion and bare ground (Year 2 only) were assessed using the Botanal technique (Tothill et al. 1992). Three trained individuals each collected data from ~100 0.25-m2 quadrats taken along three parallel trajectories in each of the three interconnected paddocks representing the grazing area. For statistical analysis, 10–15 individual observations were averaged to produce 10 mean values per person within each paddock. Sampling sites were not geo-referenced. Within 1 week of Botanal assessment, 10 random sites were selected within each of the three paddocks and the pasture was sampled by harvesting a 0.25-m2 quadrat to within 50 mm of the ground for NIR analysis of nitrogen, DM digestibility, neutral detergent fibre and acid detergent fibre.
Analyses
The nutritive value of pasture and the diet were scanned by NIR by using a monochromator (Model 6500 NIRSystems, Silver Spring, MD, USA) at the CSIRO Floreat laboratory, according to the method of Coates and Dixon (2011). Pasture samples were dried at 65°C to a constant weight in a forced-air oven and ground though a cyclonic mill (Retsch Twister, Retsch BMGH, Dusseldorf, Germany) with a 1-mm sieve and analysed for nitrogen (N), neutral detergent fibre (NDF), acid detergent fibre (ADF) and DM digestibility (DMD). CP was estimated as N × 6.25 and hemicellulose as the difference between NDF and ADF. Diet nutritive value was determined by NIR spectroscopy of faeces by using the full reference dataset of over 1000 samples from 264 diet–faecal pairs (Coates and Dixon 2011). Faecal samples prepared as described for pasture were analysed for dietary N, faecal N, NDF, ADF, DM digestibility, and organic-matter (OM) digestibility. Random samples of pasture and faeces were analysed for N by using a Leco CN628 N analyser (Leco, St Joseph, MI USA), with the following relationships:
The method of Coates and Dixon (2011) was used to estimate the percentage of non-grass (equivalent to C3 plants) in the diet from faecal NIR predictions. VFA analysis of rumen fluid was performed using the method of Gagen et al. (2014) and ammonia N by using the colorimetric method of Chaney and Marbach (1962).
Statistical analyses
Statistical analyses were performed using SAS general linear models (SAS Institute, Cary, NC, USA). All data for the two successive years (August 2015 to June 2016 and June 2016 to July 2017) were analysed separately. Significance was declared at the 5% level, with trends being declared at the 10% level. The experimental design for pasture characteristics used a general linear model with the fixed effect of season (dry versus wet), paddock and, where appropriate, operator. Operator and paddock effects were not significant and statistical comparisons were made at the season × paddock interaction. For all animal data, the experimental unit was the animal (n = 20 for the LP and HP groups). The fixed effects were season (dry versus wet) and performance group, and the means were compared at the interaction level of season × performance group (low versus high).
Results
Pasture characteristics
Biomass varied between 1.4 and 4.4 t DM/ha (Table 2). In Year 1, biomass was higher in the wet than the dry season as expected (P < 0.001); however, in Year 2, the opposite was observed, with biomass being lower in the wet season (P < 0.001). The legume content averaged 34%, and even though there was no difference between seasons in Year 2 (P > 0.05), the legume content was higher in the dry than the wet season in Year 1 (P = 0.018). The percentage of bare ground was measured only in Year 2 and averaged 34% across both dry and wet seasons (P > 0.05). The N content of pasture was higher in the wet than the dry season in both years (Year 1, P = 0.01; Year 2k P = 0.002). In terms of CP, dry-season values averaged just under 7% DM, whereas wet-season CP was over 9% DM. DMD was very low in both dry seasons, but significantly (P < 0.001) higher in the wet seasons of both years. NDF and ADF followed predictable patterns in Year 1, being lower in the wet than the dry season (NDF, P = 0.07; ADF, P < 0.001). In Year 2, NDF was not significantly (P > 0.05) different between seasons, but ADF content was lower in the wet season (P = 0.05). Because of the seasonal differences in NDF and ADF, hemicellulose was consistently higher in the wet seasons (Year 1, P = 0.01; Year 2, P < 0.001).
| Item | Dry | Wet | s.e. | P-value | |
|---|---|---|---|---|---|
| Year 1 | |||||
| Biomass (t/ha) | 1.38 | 4.39 | 0.25 | <0.001 | |
| Legume (%) | 32.3 | 25.1 | 2.48 | 0.018 | |
| Nitrogen (% DM) | 0.88 | 1.28 | 0.134 | 0.010 | |
| Crude protein (% DM) | 5.55 | 7.97 | 0.835 | 0.010 | |
| Dry-matter digestibility (%) | 21.1 | 43.6 | 2.2 | <0.001 | |
| Neutral detergent fibre (% DM) | 75.1 | 67.5 | 2.48 | 0.007 | |
| Acid detergent fibre (% DM) | 56.1 | 43.9 | 1.66 | <0.001 | |
| Hemicellulose (% DM) | 19.0 | 23.6 | 1.56 | 0.010 | |
| Year 2 | |||||
| Biomass (t/ha) | 2.09 | 1.56 | 0.09 | <0.001 | |
| Legume (%) | 37.3 | 41.9 | 1.78 | 0.074 | |
| Bare ground (%) | 34.5 | 32.7 | 1.86 | 0.420 | |
| Nitrogen (% DM) | 1.34 | 1.65 | 0.07 | 0.002 | |
| Crude protein (% DM) | 8.3 | 10.3 | 0.44 | 0.002 | |
| Dry-matter digestibility (%) | 33.2 | 40.1 | 1.0 | <0.001 | |
| Neutral detergent fibre (% DM) | 67.9 | 69.6 | 1.28 | 0.345 | |
| Acid detergent fibre (% DM) | 44.9 | 41.1 | 0.935 | 0.005 | |
| Hemicellulose (% DM) | 23.0 | 28.5 | 0.725 | <0.001 | |
Diet characteristics
Diet characteristics were estimated from faecal NIR samples (Table 3). The responses to season, performance group and their interactions were inconsistent between the 2 years.
| Item | Low | High | s.e. | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Low vs High | Dry vs Wet | Interaction | |||
| Year 1 | |||||||||
| Dietary N (% DM) | 1.43 | 1.60 | 1.39 | 1.69 | 0.037 | 0.452 | <0.001 | 0.062 | |
| Dietary CP (% DM) | 8.94 | 9.99 | 8.63 | 10.5 | 0.233 | 0.438 | <0.001 | 0.063 | |
| Faecal N (% DM) | 1.64 | 1.86 | 1.53 | 1.93 | 0.035 | 0.455 | <0.001 | 0.009 | |
| DM digestibility (%) | 56.2 | 59.0 | 56.0 | 59.7 | 0.343 | 0.396 | <0.001 | 0.174 | |
| OM digestibility (%) | 57.8 | 60.1 | 57.6 | 60.8 | 0.318 | 0.669 | <0.001 | 0.012 | |
| Non grass (%) | 41.2 | 43.3 | 41.9 | 43.2 | 1.36 | 0.995 | 0.146 | 0.667 | |
| Year 2 | |||||||||
| Dietary N (% DM) | 1.50 | 2.03 | 1.58 | 1.91 | 0.039 | 0.595 | <0.001 | 0.010 | |
| Dietary CP (% DM) | 9.40 | 12.7 | 9.90 | 12.0 | 0.246 | 0.599 | <0.001 | 0.010 | |
| Faecal N (% DM) | 1.68 | 1.82 | 1.74 | 1.71 | 0.034 | 0.514 | 0.085 | 0.009 | |
| DM digestibility (%) | 56.9 | 53.4 | 57.4 | 52.8 | 0.387 | 0.925 | <0.001 | 0.135 | |
| OM digestibility (%) | 58.9 | 55.4 | 59.3 | 54.9 | 0.348 | 0.848 | <0.001 | 0.131 | |
| Non-grass (%) | 45.3 | 43.7 | 47.5 | 42.4 | 1.61 | 0.772 | 0.043 | 0.279 | |
Seasonal effects in Year 1 were highly significant (P < 0.001) for dietary N (and CP), faecal N, DMD, and OM digestibility, with values being greater in the wet than the dry season. In Year 2, dietary N (and CP) were again higher in the wet than the dry season (P < 0.001) and tended to be higher for faecal N (P = 0.085). However, DM and OM digestibility were lower in the wet season than the dry season (P < 0.001). The proportion of C3 species in the diet, mainly represented by legumes, was not affected by season in Year 1 (P > 0.05), but was lower in the wet season in Year 2 (P = 0.043).
The nutritive value of diets selected by LP and HP cattle were not significantly (P > 0.05) different. However, for several diet-quality characteristics, there were significant interactions with season. In Year 1, significant (P < 0.05) or trend (P < 0.10) interactions were seen for dietary N (and CP; P = 0.063), faecal N (P = 0.009) and OM digestibility (P = 0.012). These interactions implied that the HP group consumed a higher-quality diet in the wet season, than in the dry season, but this response was less pronounced for the LP group. By contrast, in Year 2, significant interactions were attributed to the LP group selecting a higher-protein diet than did the HP group, as evidenced by interactions for N (and CP) and faecal N (P = 0.01). However, there were no interactions with performance group for DM and OM digestibility (P > 0.05).
Grazing behaviour
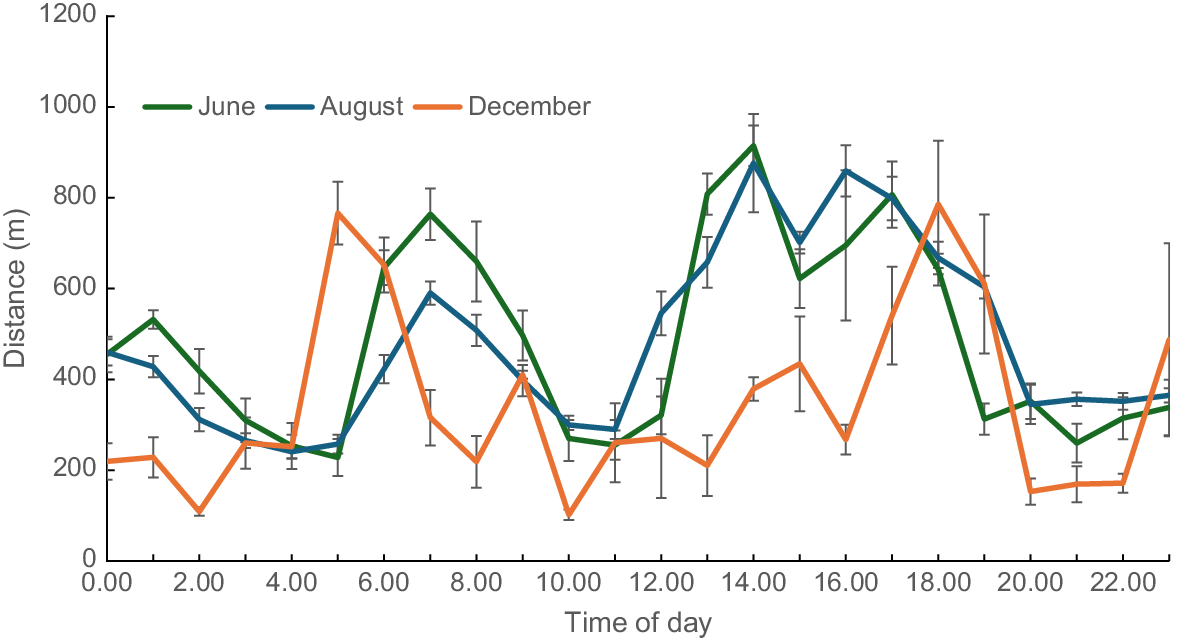
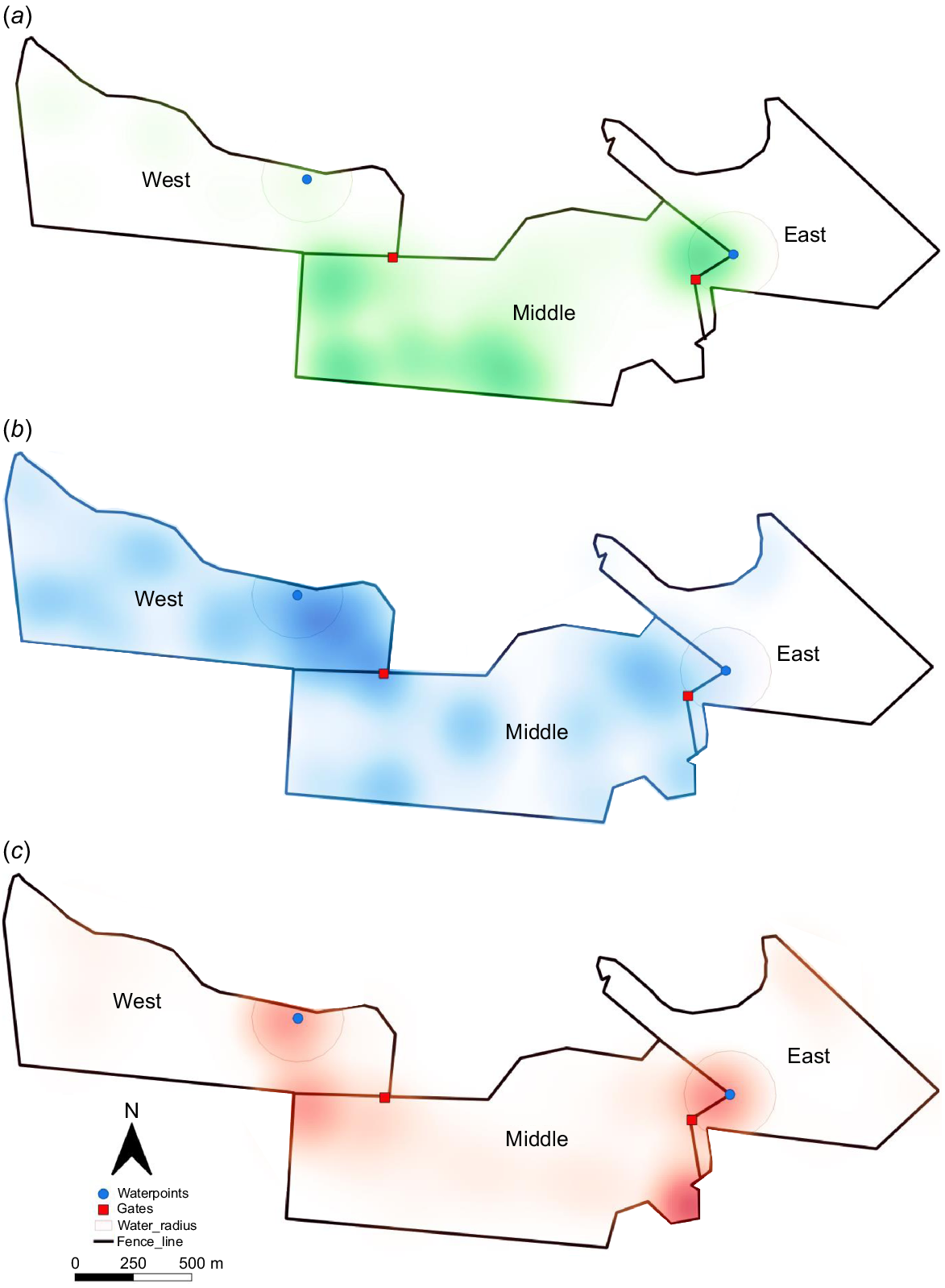
Pasture samples collected close to the periods (Table 1) when collars were on cattle showed a significant time-of-year effect for biomass (P < 0.01) and, in West paddock, legume % (P = 0.037; Table 4). Daily distance travelled by cattle was significantly (P < 0.001) affected by time-of-year, with the distance travelled in the late-dry season (December) being shorter than that in the earlier months of June and August. Travel activity was greatest between 06:00 hours and 10:00 hours and again between 12:00 hours and 18:00 hours in June and August (Fig. 2). In December, activity was much reduced and peak activity was earlier in the morning (04:00 hours to 07:00 hours) and later in the evening (17:00 hours and 20:00 hours) than in earlier periods. Fig. 3 shows the frequency with which cattle spent time in different areas of the three interconnected paddocks (West, Middle, East). In June, cattle were found predominantly within the Middle paddock, whereas in August they were equally dispersed between the West and Middle paddocks. In December, cattle were predominantly found closer to water points. Preference ratios supported the visual evidence (Table 4). In the early-dry season, cattle showed a marked preference (PR = 1.69) for Middle paddock, but 2 months later, preference had shifted to West paddock (PR = 1.64). Throughout the three measurement periods, cattle showed a marked avoidance of the East paddock. Preference ratios showed that cattle spent a disproportionate amount of time (PR on average > 2.0) within 200 m of water points, but the preference for an individual waterpoint changed over time.
| Item | Early-dry | Mid-dry | Late-dry | s.e. | P-value | |
|---|---|---|---|---|---|---|
| Deployment dates | 3/06/2016–5/07/2016 | 11/08/2016–8/09/2016 | 8/12/2016–21/12/2016 | |||
| Collars deployed (n) | 11 | 12 | 9 | – | – | |
| Days per collar (d) | 7 | 7 | 3 | – | – | |
| Functioning collars (n) | 6 | 7 | 5 | – | – | |
| Collars used to calculate distance (n) | 5 | 5 | 5 | – | – | |
| Distance travelled (km/day) | ||||||
| 10 s fix rate | 17.8c | 15.5b | 9.1a | 0.89 | <0.001 | |
| 30 s fix rate | 11.9b | 11.8b | 7.9a | 0.42 | <0.001 | |
| Preference ratios | ||||||
| West paddock | 0.34 | 1.64 | 0.72 | – | – | |
| Middle paddock | 1.96 | 1.01 | 1.52 | – | – | |
| East paddock | 0.05 | 0.17 | 0.39 | – | – | |
| Waterpoint east | 0.06 | 2.56 | 3.90 | – | – | |
| Waterpoint west | 1.95 | 0.47 | 2.91 | – | – | |
| Biomass (t/ha) | ||||||
| West paddock | 6.61 | 2.73 | 1.89 | 0.86 | 0.017 | |
| Middle paddock | 4.40 | 1.89 | 1.42 | 0.48 | 0.010 | |
| East paddock | 6.38 | 2.90 | 1.76 | 0.64 | 0.004 | |
| Legume (%) | ||||||
| West paddock | 24.2 | 52.2 | 26.7 | 6.35 | 0.037 | |
| Middle paddock | 41.4 | 26.7 | 40.0 | 9.80 | 0.540 | |
| East paddock | 32.2 | 44.0 | 30.5 | 7.51 | 0.404 |
Dates are shown as DD/MM/YY.
Means within rows with different lower case letters were significantly different (P < 0.05).
Animal performance
The average initial LW of the HP group was 22 kg heavier (P = 0.028) than that of the LP group, but final LW was 99 kg heavier for the HP group (P < 0.001), resulting in higher LWG of these cattle than of the LP group (P < 0.001; Table 5). These differences in performance resulted in increased carcase weight and value at slaughter (P < 0001). There was no effect of performance group on backfat, muscle score or killing out percentage (P > 0.05). Cattle gained more LW in the wet than dry seasons (P < 0.001; Table 6) and in Year 1, the HP group outgained the LP group (P = 0.041). In Year 2, there was an interaction trend (P = 0.059) between season and performance group, with the HP group having double the increase in LWG (+0.44 kg/day) in the wet compared with the dry season, whereas this response was only +0.22 kg/day in the LP group.
| Item | Low | High | s.e. | P-value | |
|---|---|---|---|---|---|
| Initial LW (kg) | 178 | 200 | 6.91 | 0.028 | |
| Final LW (kg) | 504 | 605 | 8.16 | <0.001 | |
| Final LW (kg) A | 516 | 593 | 3.53 | < 0.001 | |
| LWG (overall) (kg/day) | 0.468 | 0.580 | 0.005 | <0.001 | |
| Carcase weight (kg) | 270 | 320 | 5.01 | <0.001 | |
| Rib fat at quartering site (mm) | 2.90 | 3.00 | 0.290 | 0.089 | |
| Dressing % | 53.4 | 52.8 | 0.350 | 0.225 | |
| Carcass value (AU$) | 1,356 | 1,634 | 27.3 | <0.001 |
| Item | Low | High | s.e. | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Low vs high | Dry vs wet | Interaction | |||
| Overall LWG (kg/day) | |||||||||
| Year 1 | 0.06 | 0.83 | 0.16 | 0.94 | 0.041 | 0.005 | <0.001 | 0.508 | |
| Year 2 | 0.31 | 0.54 | 0.29 | 0.67 | 0.042 | 0.206 | <0.001 | 0.047 | |
| DM intake (g/kg LW) | |||||||||
| Year 1 | 23.0 | 32.5 | 23.2 | 34.4 | 0.596 | 0.075 | <0.001 | 0.157 | |
| Year 2 | 23.7 | 22.1 | 23.8 | 23.7 | 0.395 | 0.210 | 0.006 | 0.299 | |
Initial liveweight used as a covariate.
QuikIntake ver. 6, 2019 (McLennan et al. 2020).
Rumen fermentation and blood urea-N
Rumen fermentation was highly influenced by season, but performance group effects were small or not apparent (Table 7).
| Item | Low | High | s.e. | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Low vs High | Dry vs wet | Interaction | |||
| Year 1 | |||||||||
| Blood urea-N (mg/100 mL) | 9.71 | 10.63 | 9.03 | 10.63 | 0.436 | 0.434 | 0.005 | 0.430 | |
| Rumen NH3 N (mg/100 mL) | 6.15 | 9.06 | 6.11 | 7.71 | 0.651 | 0.297 | 0.001 | 0.323 | |
| Total VFA (mM) | 65.4 | 54.2 | 61.4 | 45.0 | 3.04 | 0.033 | <0.001 | 0.401 | |
| VFA (molar %) | |||||||||
| Acetate | 69.6 | 69.7 | 70.1 | 70.9 | 0.315 | 0.116 | 0.118 | 0.047 | |
| Propionate | 17.5 | 14.4 | 17.3 | 14.5 | 0.282 | 0.975 | <0.001 | 0.571 | |
| Iso-butyrate | 1.02 | 1.39 | 1.01 | 1.54 | 0.086 | 0.369 | <0.001 | 0.348 | |
| Butyrate | 8.86 | 10.9 | 8.45 | 10.6 | 0.202 | 0.076 | <0.001 | 0.840 | |
| Iso-valerate | 1.36 | 1.90 | 1.23 | 2.08 | 0.089 | 0.804 | <0.001 | 0.090 | |
| Valerate | 1.10 | 1.05 | 0.841 | 1.10 | 0.058 | 0.071 | 0.078 | 0.011 | |
| Caproate | 0.535 | 0.629 | 0.375 | 0.554 | 0.061 | 0.056 | 0.027 | 0.484 | |
| Acetate/propionate | 4.00 | 4.93 | 4.12 | 4.83 | 0.090 | 0.877 | <0.001 | 0.219 | |
| pH | 6.93 | 7.02 | 7.08 | 7.25 | 0.061 | 0.003 | 0.030 | 0.550 | |
| Year 2 | |||||||||
| Blood urea-N (mg/100 mL) | 10.9 | 11.2 | 10.1 | 11.4 | 0.447 | 0.464 | <0.001 | 0.310 | |
| Rumen NH3 N (mg/100 mL) | 5.37 | 6.86 | 5.24 | 7.07 | 0.465 | 0.935 | 0.010 | 0.710 | |
| Total VFA (mM) | 60.5 | 63.0 | 59.4 | 58.6 | 3.52 | 0.439 | 0.807 | 0.627 | |
| VFA (molar %) | |||||||||
| Acetate | 73.8 | 73.3 | 73.7 | 73.3 | 0.201 | 0.863 | <0.026 | 0.667 | |
| Propionate | 15.0 | 13.3 | 15.1 | 13.2 | 0.161 | 0.844 | <0.001 | 0.602 | |
| Iso-butyrate | 0.921 | 1.18 | 0.921 | 1.23 | 0.043 | 0.563 | <0.001 | 0.544 | |
| Butyrate | 8.08 | 9.37 | 8.11 | 9.33 | 0.123 | 0.840 | <0.001 | 0.949 | |
| Iso-valerate | 1.15 | 1.53 | 1.17 | 1.65 | 0.059 | 0.223 | <0.001 | 0.473 | |
| Valerate | 1.12 | 1.58 | 1.18 | 1.66 | 0.063 | 0.282 | <0.001 | 0.887 | |
| Caproate | 0.349 | 0.343 | 0.314 | 0.259 | 0.020 | 0.004 | 0.130 | 0.231 | |
| Acetate:propionate | 4.96 | 5.56 | 4.92 | 5.57 | 0.075 | 0.838 | <0.001 | 0.786 | |
| pH | 7.12 | 7.18 | 7.31 | 7.34 | 0.052 | 0.001 | 0.357 | 0.820 | |
Total VFA were lower in the wet than the dry season (P < 0.001), as was the percentage of propionate (P < 0.001). Acetate percentage was not influenced by season (P > 0.05), but all other VFA (except valerate) showed an increase in molar percentage (P < 0.05). Consequently, the acetate to propionate ratio (P < 0.001) and rumen pH (P = 0.030 0.05) were higher in the wet than the dry season. Blood urea-N (P < 0.01) and rumen ammonia N (P < 0.001) concentrations were higher in the wet than the dry season. Total VFA were lower in HP than LP cattle (P < 0.05). While there were no differences in molar proportions of individual VFA between performance groups, there was a significant interaction (P < 0.05) for acetate, with HP cattle having a higher acetate percentage in the wet season, whereas in LP cattle there was no effect. The only other significant performance effect was for rumen pH, which was higher in HP cattle (P < 0.01).
Results in Year 2 were broadly similar to those in Year 1, although in contrast to Year 1, total VFA concentration was not influenced by season or performance group (P > 0.05). Acetate to propionate ratio was not affected by season (P > 0.05) but was higher in the HP cattle (P < 0.001) as was rumen pH (P = 0.001). Blood urea-N (P < 0.001) and rumen ammonia (P < 0.01) were higher in the wet than the dry season, but were not affected by performance group.
Discussion
Pasture and diet characteristics
Pasture biomass in northern Australia is highly responsive to rainfall and stocking rates. Typically, biomass accumulates rapidly in the wet season, to reach a peak of between 4 and 8 t/ha depending on the location, rainfall, and previous and current stocking rates (Bortolussi et al. 2005). This pattern was observed in the first year of the study, but not in the second year, where biomass was lower in the wet season that the dry season. The study was conducted during a period of lower-than-average rainfall. The long-term mean for Lansdown is 901 mm/year, but over the 3 years in which the study occurred, rainfall was 460, 624 and 842 mm, with just 1344 mm falling during the 23-month study period. The pasture area in this almost 2-year study was stocked with 89 growing cattle, with average initial and final LW of 188 and 554 kg respectively. Thus, stocking rate increased over the duration of the study (as animals increased in size) from ~5 ha per adult equivalent (AE) to 3 ha/AE in the following year. We postulate that this led to overutilisation of the pastures in the latter months of the trial, during this lower-rainfall period. Despite this, LWG remained higher in the wet than the dry season in Year 2, although the response to season was smaller than in Year 1.
Throughout the study, legume content in the pasture remained above 25%, being as high as 41% in the second dry season when total pasture biomass was low. Undoubtedly, this contributed to the acceptable rates of gain (overall 0.52 kg/day) observed because tropical legumes are generally of higher nutritive value than are C4 grasses (Archimède et al. 2011) and increase voluntary intake (Kennedy and Charmley 2012). Faecal NIR showed that intake of C3 non-grass (corresponding to legume in the diet) was higher than the proportion of legume measured in pasture by using Botanal techniques. This suggested that cattle were preferentially selecting legume over grass in the diet, as noted by Dixon and Coates (2008). The preference for legume was reflected in the CP content of the diet that was two percentage units higher than in the pasture. It was also apparent that the legume and CP contents of pasture and diets were greater in Year 2 than Year 1, reflective of the lower total biomass in Year 2 having a concentrating effect on legume proportion.
Pasture DMD in the dry season of Year 1 was extremely low (21%). However, only a single sampling was taken for this measurement at the very end of the dry season (December), so the data may not accurately reflect the digestibility of the pasture over the preceding grazing period (August to December). In the following wet season, the expected increase in digestibility was apparent as new vegetative growth replaced the mature and dead material of the dry season (Tothill and Gillies 1992). The seasonal response in pasture digestibility was broadly similar from Year 2 to Year 1.
Diet digestibility from faecal NIR did not follow the same pattern as that observed for digestibility in pasture samples. In Year 1, diet digestibility from NIR was predictably higher than that of the pasture and was higher in the wet than the dry season. These data reflected the selection by cattle for legumes and less stemmy, higher-digestibility grasses (Benvenutti et al. 2015). In contrast, DM digestibility in Year 2 was lower in the wet than dry seasons. It is hypothesised that the low biomass availability restricted the animal’s ability to select the more digestible leaf and green stems of grasses and digestibility was compromised (Chacon and Stobbs 1976).
Grazing behaviour
The relationship between the animal and pasture is critical to both the sustainability of the pasture and the performance of the animals grazing that pasture. We deployed GPS collars periodically throughout the dry season in 2016 to follow the patterns of grazing behaviour as pasture biomass and quality declined. It was not possible to successfully deploy collars in the wet season because many collars failed owing to the wet conditions. Given the small proportion of collard cattle (6–8%), the sampled cattle may not have represented the behaviour of the herd. However, research with virtual fencing technology has shown that the herding instinct in cattle is strong and successful animal control can be achieved with a small proportion of collared cattle (Campbell et al. 2021). Other research has suggested that even a single animal may be sufficient (Moritz et al. 2012). Cattle walked about 14 km/day (at 10 s fix rate) when biomass was above 2 t/ha, but activity dropped off markedly in December when biomass dropped below this value and temperatures increased. Foraging theory would predict that as the feed resource declines, then so too does activity (walking and grazing), to conserve energy for maintenance and production (Owen-Smith et al. 2010). Studies of grazing behaviour (Chacon and Stobbs 1976; da Silva et al. 2013) suggest that initially as biomass declines, cattle will graze longer to maintain intake but ultimately grazing time (and associated activity) will decrease in response to biomass availability. Although it was not possible to differentiate between grazing and walking, it is likely that much of the observed activity was to maximise feed intake. It was speculated that activity (as measured by distance travelled) may be related to growth performance. However, the limited activity data gathered did not allow this theory to be tested.
The determination of accurate travel distance by cattle is difficult. Very high fix rates magnify the inherent GPS error and include the micro-movements of the animal. Very low fix rates increase interpolation error (McGavin et al. 2018); that is the path tortuosity is not captured to the same degree as the length of time between points increases. McGavin et al. (2018) compared distance travelled with fix rates varying from 1 to 600 s and concluded that for cattle in northern Australia values of 5 to 10 s were most appropriate to use.
Cattle do not graze uniformly and in large paddocks will select certain areas over others. Reasons for how animals choose to utilise an area are difficult to ascertain, but factors such as water points and supplement locations, shade, pasture biomass, species and quality, soil type and steepness of terrain are all important factors (Tomkins and O’Reagain 2007; Charmley et al. 2023). In the current study, cattle avoided the East paddock for inexplicable reasons, as biomass and legume content appeared adequate. The highest legume content was seen in Middle paddock (47%) and this paddock showed the highest preference ratio in two of the three measurement periods. In the other period, cattle dispersed into West paddock, again when legume content was high (52%). Although it is ill-advised to make too much of the link between legume content and preference, the data do support the known preference for legumes over grasses in mixed swards (Rutter 2006).
As the season advanced and pasture biomass and quality declined, cattle spent an increasing proportion of their time near waterpoints. When the data from both waterpoints are combined the preference ratio was 1.0, 1.5 and 3.4 for the early-, mid- and late-season measurement periods. The likelihood of overgrazing near these waterpoints is high (Hunt et al. 2007), and overgrazing may explain why cattle switched preference between waterpoints; as biomass became depleted at one point, cattle switched to a point with higher biomass. Cattle are known to have two major grazing periods throughout the day, one from dawn to mid-morning and a second in the afternoon, typically after 16:00 hours (Tomkins and O’Reagain 2007; Tomkins et al. 2009). The current data partially support this observation, although in the early- and mid-dry seasons, peaks were broader than those observed by Tomkins and O’Reagain (2007) and Tomkins et al. (2009). In the late-dry season, peak activity was more clearly defined and linked to dawn and dusk, suggesting that as biomass became depleted, cattle spent less time grazing and more time resting, a supposition reinforced by the higher preference to be near waterpoints in December.
Animal performance
Cattle were sourced from a local herd without data on birth date, breeding values or sires. Thus, it was not possible to take these into account when selecting the LP and HP groups at the completion of the grazing trial. The 22-kg higher initial LW of the HP group could have affected subsequent performance. However, it is unlikely that any pre-trial management, such as age at weaning or compensatory gain, would have persisted throughout the 23 months of study. Such differences usually disappear over time (Berge 1991). Genetic variation in production phenotypes such as feed efficiency or feed intake could have influenced performance both before and during the trial. In a review by Cantalapiedra-Hijar et al. (2018) it was suggested that differences in the rumen microbiome could influence rumen volume and absorptive capacity and, in so doing, affect feed intake. Differences in lean to fat ratio in the carcase and protein turnover rates have also been implicated in affecting feed efficiency (Kenny et al. 2018).
We hypothesised that higher performance by cattle could be attributed to differences in either diet selection, digestion, or animal grazing behaviour, acknowledging that influence on one factor could elicit a response in the others. Consequently, statistical comparisons were drawn between the best- and worst-performing animals as measured by LW gain. Over the 23 months of the trial, annual LW gain was 190 kg/ha, somewhat above the norm for this area of 111–190 kg/ha (Bortolussi et al. 2005). This can be attributed to the high proportion of legume in the diet (Charmley et al. 2023). LW gains of the LP and HP groups were significantly different, as was expected, allowing for a meaningful comparison between the two groups. As a consequence of higher carcase weight, the value of beef sold from the highest-performing cattle was 20% greater than the value of the carcases from the lowest-performing cattle, indicating the importance of selecting for better-performing animals and understanding and managing the reasons for the difference (Silva et al. 2022).
When the diet nutritive value was compared between LP and HP cattle, overall there was no effect. However, when the interaction with season is considered, it is apparent that in the Year 1 wet season, there was some evidence that HP cattle selected diets with higher nutritive value (higher OM digestibility and a trend for higher N content). This was not apparent when cattle were older in Year 2, where the reverse was observed. The VFA concentration of HP cattle was lower than that of LP cattle in Year 1 when forage availability was non-limiting. Although there were no significant effects on acetate and propionate proportions, pH was consistently higher in HP cattle.
The wet-season response in rumen fermentation in Year 1 was contrary to expectations. The improved nutritive value was expected to increase rumen fermentation, resulting in higher total VFA and a trend away from acetate towards propionate fermentation as the fibre content of the diet was lower in the wet season. However, in the first year of our study, total VFA concentration was reduced and the acetate:propionate ratio increased in the wet season. Bowen et al. (2017) also observed a reduction in VFA concentration in the wet season compared with the dry season in steers grazing black speargrass (Heteropogon contortus)-dominated pasture. Similarly, Martinez-Fernandez et al. (2020) observed lower VFA concentration in pregnant heifers grazing Mitchell grass (Bothriochloa erwartiana) in the wet season than in the dry season. These data (Bowen et al. 2017; Martinez-Fernandez et al. 2020), together with data presented in this paper, suggest that lower VFA concentration in the wet season may be due to higher rumen outflow rates (Bowen et al. 2017) which could also imply higher intake (Hendrickson et al. 1981). In support of this theory, Perry et al. (2017) compared tropical grasses of similar nutritive value (~9% CP and ~62% DM digestibility) either as conserved baled hay or cut and carry pasture. They observed lower (nearly halved) VFA concentration and significantly higher rumen outflow rates in the fresh cut and carry forage than in hay. Increased rumen turnover would reduce the instantaneous balance between VFA production and absorption. Other factors such as grazing events and relative sampling times (Playne and Kennedy 1976), and water intake (Rodríguez-Prado et al. 2012) from high-moisture pasture plants may also influence rumen VFA concentration. If we contend that low VFA concentration is linked to higher rumen turnover and intake, then it is possible that HP cattle were achieving higher intake through increased rate of passage from the rumen (Forbes 2003). HP cattle exhibited a marginally greater response to diet nutritive value in the wet season than did LP cattle, especially in Year 1. This suggests that increased LW gain of HP cattle may have been influenced by selection for more nutrient-rich pasture components contributing to higher intake of energy and protein. Additionally, the markedly lower LW gain of the high-intake group in Year 2 (0.68 kg/day) than in Year 1 (0.98 kg/day) demonstrates the added constraint of pasture availability on performance.
In low-protein diets, Silva et al. (2022) contended that variation in N-use efficiency is sufficiently large to affect dry-season performance. The dietary CP contents in the current trial were relatively high for tropical pastures (Hunter and Siebert 1985; Panjaitan et al. 2010), presumably owing to the high proportion of legume in the diet. These concentrations were not likely to limit microbial protein synthesis (Bowen et al. 2017) and rumen ammonia concentrations were above 5 mg/dL, considered to be the minimum to maintain optimum microbial protein synthesis in the rumen (Satter and Slyter 1974). The proportion of branch-chain fatty acids were higher in the wet seasons, in agreement with trials reported by Playne and Kennedy (1976) and Martinez-Fernandez et al. (2020). As branch-chain fatty acids are derived from the deamination of dietary amino acids, higher concentrations in the wet season demonstrate improved N availability in the rumen, even though concentrations were considered adequate in the dry season.
The marked seasonal pattern in animal growth rate was typical of northern Queensland, with low rates of gain in the dry season contrasted with high gains in the wet season. Although this was particularly apparent in Year 1, with gains increasing from ~0.1 in the dry to ~0.9 kg/day in the wet season, it was less so in the second year. Year 2 was characterised by a poor response in pasture biomass production to wet-season rainfall, potentially owing to overstocking in Year 1 during a period of lower-than-average rainfall. Although it was not possible to measure intake in this study, it can be predicted from growth rate, animal weight and activity (McLennan et al. 2020). This approach sums the metabolisable-energy (ME) requirements for maintenance, growth and activity and relates this to the ME content of the diet (from faecal NIR). Dry-season DM intakes for HP and LP cattle were estimated to be 23 g/kg LW in both years. However, wet-season intake was estimated at 34 g/kg LW in Year 1, but only 23 g/kg LW in Year 2. These values suggest that intake was compromised in the second wet season because of low digestibility (~40%) and low biomass availability (1.6 t/ha). Coleman (2005) noted that voluntary intake was constrained when biomass was less than ~1.7 t/ha. Also, Forbes and Coleman (1993) concluded that intake per bite was restricted when green leaf biomass was less than 1.1 t/ha. It was concluded that availability of green biomass was restricting voluntary intake and, hence, LW gain in the second wet season. These observations are supported by da Silva et al. (2013) who manipulated stocking rate to influence biomass, digestibility and intake and showed that biomass had a greater influence on animal performance than did digestibility.
Conclusions
The objective of this study was to determine the relative importance of diet quality and selection, rumen function, and grazing behaviour on performance of grazing cattle. Useful data were obtained on the first two factors; however, for technical reasons we retrieved successful behavioural data only in the dry season where no relationship between activity and performance was found. Also, there were not sufficient data collected to enable a comparison of grazing behaviour between the HP and LP groups.
It is well understood that availability and nutritive value of pasture have a profound effect on growth of grazing cattle. The current study reaffirmed this fact. However, this work also showed that there were differences in how the lowest- and highest-preforming cattle were able to exploit the grazing environment. Although HP cattle appeared to select biomass of marginally better N content and digestibility in the wet season, such small differences did not affect rumen ammonia concentration or the balance of VFA in the rumen. Thus, diet quality differences could only have accounted for a small proportion of the improvement in LW gain observed between LP and HP cattle. The interesting reduction in VFA concentration in HP cattle suggested that perhaps rumen turnover was increased under conditions of ample biomass and high diet quality, as seen in the first wet season. The balance of evidence suggests that the two groups of cattle diverged in performance in the wet season but not the dry season, and this was most pronounced when pasture availability was non-limiting. It remains to be determined whether grazing behaviour could be a factor contributing to animal performance. Evidence for variation among animals in rumen function, diet selection or animal activity was scant, and we assume that in the absence of data, differences in performance could be attributed to differences in initial LW, increased intake in the wet season or genetic differences in efficiency of energy utilisation or protein turnover.
Data availability
Data used to generate the results in the paper will be shared upon reasonable request to the corresponding author.
Conflicts of interest
Ed Charmley is an associate editor of Animal Production Science. To mitigate this potential conflict of interest they had no editor-level access to this manuscript during peer review. The authors declare that they have no other conflicts of interest.
Declaration of funding
Funding for this project was provided by a CSIRO internal competitive funding process.
Acknowledgements
Animal husbandry and technical support was provided by Mel Matthews, Wayne Flintham, Steve Austin, Jag Padmanabha, Pete Giacomantonio, and Elizabeth Hulm.
References
Arablouei R, Wang L, Currie L, Yates J, Alvarenga FAP, Bishop-Hurley GJ (2023) Animal behavior classification via deep learning on embedded systems. Computers and Electronics in Agriculture 207, 107707.
| Crossref | Google Scholar |
Archimède H, Eugène M, Marie Magdeleine C, Boval M, Martin C, Morgavi DP, Lecomte P, Doreau M (2011) Comparison of methane production between C3 and C4 grasses and legumes. Animal Feed Science and Technology 166–167, 59-64.
| Crossref | Google Scholar |
Arthur PF, Archer JA, Herd RM (2004) Feed intake and efficiency in beef cattle: overview of recent Australian research and challenges for the future. Australian Journal of Experimental Agriculture 44, 361-369.
| Crossref | Google Scholar |
Bell A, Sangster N (2023) Research, development and adoption for the north Australian beef cattle breeding industry: an analysis of needs and gaps. Animal Production Science 63, 1-40.
| Crossref | Google Scholar |
Benvenutti MA, Pavetti DR, Poppi DP, Gordon IJ, Cangiano CA (2015) Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality of cattle. Grass and Forage Science 71, 424-436.
| Crossref | Google Scholar |
Berge P (1991) Long-term effects of feeding during calfhood on subsequent performance in beef cattle (a review). Livestock Production Science 28, 179-201.
| Crossref | Google Scholar |
Bortolussi G, McIvor JG, Hodgkinson JJ, Coffey SG, Holmes CR (2005) The northern Australian beef industry, a snapshot. 3. Annual liveweight gains from pasture based systems. Australian Journal of Experimental Agriculture 45, 1093-1108.
| Crossref | Google Scholar |
Bowen MK, Poppi DP, McLennan SR (2017) Efficiency of rumen microbial protein synthesis in cattle grazing tropical pastures as estimated by a novel technique. Animal Production Science 57, 1702-1712.
| Crossref | Google Scholar |
Campbell DLM, Marini D, Lea JM, Keshavarzi H, Dyall TR, Lee C (2021) The application of virtual fencing technology effectively herds cattle and sheep. Animal Production Science 61, 1393-1402.
| Crossref | Google Scholar |
Cantalapiedra-Hijar G, Abo-Ismail M, Carstens GE, Guan LL, Hegarty R, Kenny DA, McGee M, Plastow G, Relling A, Ortigues-Marty I (2018) Review: biological efficiency determinants of between-animal variation in feed efficiency of growing beef cattle. Animal 12(S2), 321-335.
| Crossref | Google Scholar |
Chaney AL, Marbach EP (1962) Modified reagents for determination of urea and ammonia. Clinical Chemistry 8, 130-132.
| Crossref | Google Scholar | PubMed |
Chacon E, Stobbs TH (1976) Influence of progressive defoliation of a grass sward on the eating behaviour of cattle. Australian Journal of Agricultural Research 27, 709-727.
| Crossref | Google Scholar |
Charmley E, Thomas D, Bishop-Hurley GJ (2023) Revisiting tropical pasture intake: what has changed in 50 years? Animal Production Science 63, 1851-1865.
| Crossref | Google Scholar |
Coates DB (1996) Diet selection by cattle grazing Stylosanthes-grass pastures in the seasonally dry tropics: effect of year, season, stylo species and botanical composition. Australian Journal of Experimental Agriculture 36, 781-789.
| Crossref | Google Scholar |
Coates DB, Dixon RM (2011) Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages. Journal of Near Infrared Spectroscopy 19, 507-519.
| Crossref | Google Scholar |
Da Silva SC, Gimenes FMA, Sarmento DOL, Sbrissia AF, Oliveira DE, Hernandez-Garay A, Pires AV (2013) Grazing behaviour, herbage intake and animal performance of beef cattle heifers on marandu palisade grass subjected to intensities of continuous stocking management. Journal of Agricultural Science 151, 727-739.
| Crossref | Google Scholar |
Dixon RM, Coates DB (2008) Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with faecal near infrared reflectance spectroscopy (F.NIRS). Australian Journal of Experimental Agriculture 48, 835-842.
| Crossref | Google Scholar |
Forbes JM (2003) The multifactorial nature of food intake control. Journal of Animal Science 81, E139-E144.
| Crossref | Google Scholar |
Forbes TDA, Coleman SW (1993) Forage intake and ingestive behavior of cattle grazing old world bluestems. Agronomy Journal 85, 808-816.
| Crossref | Google Scholar |
Gagen EJ, Wang J, Padmanabha J, Liu J, de Carvalho IPC, Liu J, Webb RI, Al Jassam R, Morrison M, Denman SE, McSweeney CS (2014) Investigation of a new acetogen isolated from an enrichment of the tammar wallaby forestomach. BMC Microbiology 7, 1122.
| Crossref | Google Scholar |
Hendrickson RE, Poppi DP, Minson DJ (1981) The voluntary intake, digestibility and retention time by cattle and sheep of stem and leaf fractions of a tropical legume (Lablab purpureus). Australian Journal of Agricultural Research 32, 389-398.
| Crossref | Google Scholar |
Hobbs NT, Bowden DC (1982) Confidence intervals on food preference indices. The Journal of Wildlife Management 46, 505-507.
| Crossref | Google Scholar |
Hunt LP, Petty S, Cowley R, Fisher A, Ash AJ, MacDonald N (2007) Factors affecting the management of cattle grazing distribution in northern Australia: preliminary observations on the effect of paddock size and water points. The Rangeland Journal 29, 169-179.
| Crossref | Google Scholar |
Hunter RA, Siebert BD (1985) Utilization of low-quality roughage by Bos taurus and Bos indicus cattle 2. The effect of rumen-degradable nitrogen and sulphur on voluntary food intake and rumen characteristics. British Journal of Nutrition 53, 649-656.
| Crossref | Google Scholar | PubMed |
Kennedy PM, Charmley E (2012) Methane yields from Brahman cattle fed tropical grasses and legumes. Animal Production Science 52, 225-239.
| Crossref | Google Scholar |
Kenny DA, Fitzsimmons C, Waters SM, McGee M (2018) Invited review: Improving feed efficiency of beef cattle – the current state of the art and future challenges. Animal 12, 1815-1826.
| Crossref | Google Scholar | PubMed |
Lawrence P, Kenny DA, Earley B, McGee M (2013) Intake of conserved and grazed grass and performance traits in beef suckler cows differing in phenotypic residual feed intake. Livestock Science 152, 154-166.
| Crossref | Google Scholar |
Martinez-Fernandez G, Jiao J, Padnamabha J, Denman SE, McSweeney CS (2020) Seasonal and nutrient supplement response in rumen microbiota structure and metabolites of tropical rangeland cattle. Microorganisms 8, 1550.
| Crossref | Google Scholar | PubMed |
McCosker KD, Perkins NR, Fordyce G, O’Rourke PK, McGowan MR (2023) Reproductive performance of northern Australia beef herds. 5. Factors influencing risk of non-pregnancy. Animal Production Science 63, 350-364.
| Crossref | Google Scholar |
McGavin SL, Bishop-Hurley GJ, Charmley E, Greenwood PL, Callaghan MJ (2018) Effect of GPS sample interval and paddock size on estimates of distance travelled by grazing cattle in rangeland, Australia. The Rangeland Journal 40, 55-64.
| Crossref | Google Scholar |
McLennan S, McLean I, Paton C (2020) Re-defining the animal unit equivalence (AE) for grazing ruminants and its application for determining forage intake, with particular relevance to the northern Australian grazing industries. Project B.GBP.0036 Final Report. Meat and Livestock Australia, Sydney, NSW, Australia.
Moritz M, Galehouse Z, Hao Q, Garabed RB (2012) Can one animal represent an entire herd? Modelling pastoral mobility using GPS/GIS technology. Human Ecology 40, 623-630.
| Crossref | Google Scholar |
Mwangi FW, Gardiner CP, Walker G, Hall TJ, Malau-Aduli BS, Kinobe RT, Malau-Aduli AEO (2021) Growth performance and plasma metabolites of grazing beef cattle backgrounded on buffel or buffel – Desmanthus mixed pastures. Animals 11, 2355.
| Crossref | Google Scholar |
Owen-Smith N, Fryxell JM, Merrill EH (2010) Foraging theory upscaled: the behavioural ecology of herbivore movement. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2267-2278.
| Crossref | Google Scholar |
Panjaitan T, Quigley SP, McLennan SR, Swain T, Poppi DP (2010) Intake, retention time in the rumen and microbial protein production of Bos indicus steers consuming grasses varying in crude protein content. Animal Production Science 50, 444-448.
| Crossref | Google Scholar |
Perry LA, Al Jassim R, Gaughan JB, Tomkins NW (2017) Effect of feeding forage characteristic of wet-or dry-season tropical C4 grass in northern Australia, on methane production, intake and rumen outflow rates in Bos indicus steers. Animal Production Science 57, 2033-2041.
| Crossref | Google Scholar |
Petty SR, Poppi DP (2012) The liveweight gain response of heifers to supplements of molasses or maize while grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia. Animal Production Science 52, 619-623.
| Crossref | Google Scholar |
Playne HJ, Kennedy PM (1976) Ruminal volatile fatty acids and ammonia in cattle grazing dry tropical pastures. Journal of Agricultural Science 86, 367-372.
| Google Scholar |
Poppi DP, McLennan SR (1995) Protein and energy utilization by ruminants at pasture. Journal of Animal Science 73, 278-290.
| Crossref | Google Scholar | PubMed |
Rodríguez-Prado M, Ferret A, Zwieten J, Gonzalez L, Bravo D, Calsamiglia S (2012) Effects of dietary addition of capsicum extract on intake, water consumption, and rumen fermentation of fattening heifers fed a high-concentrate diet. Journal of Animal Science 90, 1879-1884.
| Crossref | Google Scholar | PubMed |
Rutter SM (2006) Diet preference for grass and legumes in free-ranging domestic sheep and cattle: current theory and future application. Applied Animal Behaviour Science 97, 17-35.
| Crossref | Google Scholar |
Satter LD, Slyter LL (1974) Effect of ammonia concentration on ruminal microbial protein production in vitro. British Journal of Nutrition 32, 199-208.
| Crossref | Google Scholar | PubMed |
Silva LFP, Hegarty RS, Meale SJ, Costa DAF, Fletcher MT (2022) Using the natural abundance of nitrogen isotopes to identify cattle with greater efficiency in protein-limiting diets. Animal 16, 100551.
| Crossref | Google Scholar | PubMed |
Tomkins NW, O’Reagain PJ (2007) Global positioning systems indicate landscape preference s of cattle in the subtropical savannas. The Rangeland Journal 29, 217-222.
| Crossref | Google Scholar |
Tomkins NW, O’Reagain PJ, Swain D, Bishop-Hurley G, Charmley E (2009) Determining the effect of stocking rate on the spatial distribution of cattle for the subtropical savannas. The Rangeland Journal 31, 267-276.
| Crossref | Google Scholar |