

Shoot flammability patterns among plant species of the wildland–urban interface in the fire-prone Greater Blue Mountains World Heritage Area
Brad R. Murray A * , Thomas Hawthorne A , Timothy J. Curran B , Daniel W. Krix A , Molly I. Wallace A , Kieran Young A , Megan L. Murray A , Elisabeth Morley A , Nicola Huber-Smith A and Jonathan K. Webb AA School of Life Sciences, University of Technology Sydney, PO Box 123, Ultimo, NSW 2007, Australia.
B Department of Pest-Management and Conservation, Lincoln University, PO Box 85084, Lincoln, Canterbury 7647, New Zealand.
International Journal of Wildland Fire 32(7) 1119-1134 https://doi.org/10.1071/WF22192
Submitted: 2 September 2022 Accepted: 3 April 2023 Published: 26 April 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Background: Mitigation of wildfires at the wildland–urban interface (WUI) will be enhanced by understanding the flammability of plants growing in this zone.
Aims: We aimed to: (1) compare shoot flammability among wildland native, and both urban native and urban exotic ornamental plants; (2) quantify relationships between shoot traits and flammability; and (3) establish flammability scores to distinguish low- from high-flammability species.
Methods: Flammability and traits of field-collected shoots were measured and relationships quantified in 44 species from the Blue Mountains World Heritage Area, Australia.
Key results: In our study area, urban exotic plants were less flammable than wildland and urban native plants. Slow-igniting shoots had high fuel moisture and bulk density; short-burning shoots had low bulk density and volume; shoots recording low maximum temperatures had high fuel moisture, low bulk density and volume; and shoots with low biomass consumed in flames had high fuel moisture and low volume. Our novel flammability scores distinguished low-flammability (e.g. Lophostemon confertus) from high-flammability native species (e.g. Callistemon citrinus).
Conclusions and implications: Low-flammability plantings at the WUI should preferably use native species given potential ecological impacts of exotics. We suggest that future work should seek to identify broader suites of low-flammability native species.
Keywords: combustibility, consumability, fuel, ignitibility, sustainability, trait, wildfire, wildland–urban interface.
Introduction
Wildfires are an important ecological disturbance in many environments around the world, creating a range of habitats and maintaining native biodiversity (He et al. 2019). However, wildfires that occur close to urban regions at the wildland–urban interface (WUI) pose serious risks to human communities (McWethy et al. 2019). The WUI is the frontline for human–wildfire interactions and typically consists of low-level urban development nested within regions of natural or modified wildland vegetation (Radeloff et al. 2005). Increasing wildfire activity in the coming decades (Bowman et al. 2017a) could lead to enormous social, economic and ecological costs concentrated at the WUI (Nolan et al. 2021; Wang et al. 2021). Given that the WUI is continually being augmented and expanding (Moritz et al. 2022), there is a pressing need to understand wildfire dynamics at the WUI and to develop effective ways to mitigate wildfire impacts (Gill and Stephens 2009).
Plants provide fuel for wildfires, and there is considerable variation among species in the flammability of leaves (Murray et al. 2013), canopy shoots (Kraaij et al. 2022) and whole plants (Gao and Schwilk 2022). This variation plays an important role in dictating wildfire dynamics (Weise et al. 2005; Plucinski et al. 2010; Schwilk 2015; Fares et al. 2017), and species flammability is now a fundamental component of models predicting wildfire behaviour (Zylstra et al. 2016; Prince et al. 2017; Tumino et al. 2019; Burton et al. 2021; Popović et al. 2021; Zylstra and Liow 2021). In this context, determining the relative flammability of three main plant groups – wildland native, urban native and urban exotic plants – at the WUI will enhance our understanding of wildfire dynamics in this zone. Wildland native species are those plants found in natural vegetation immediately adjacent to urban areas, whereas urban native and urban exotic species are ornamental plants growing in residential gardens.
At present, there is a paucity of information about the flammability of plants at the WUI. Previous studies have focused predominantly on the flammability of fine fuels such as leaves (Ganteaume et al. 2013; Murray et al. 2013; Molina et al. 2017; Blackhall and Raffaele 2019; Krix et al. 2019; Romero et al. 2019). This presents an intriguing challenge for understanding wildfire dynamics across WUI landscapes; recent work seems to indicate that leaf flammability may not adequately represent whole plant flammability (Alam et al. 2020). For many plant assemblages, assessing whole plant flammability is simply not feasible (Jaureguiberry et al. 2011). For instance, experimentally burning replicates of tall, isolated trees such as eucalypts is not a realistic option. Assessments of canopy shoot flammability, however, which encompass leaves and branches with intact plant architecture, provide an efficient and robust assessment of whole plant flammability (Wyse et al. 2016; Alam et al. 2020). Although shoot flammability patterns among species in non-WUI areas have provided important insights into wildfire dynamics in Argentina (Santacruz-García et al. 2019), South Africa (Burger and Bond 2015; Calitz et al. 2015; Kraaij et al. 2022) and New Zealand (Wyse et al. 2016; Padullés Cubino et al. 2018; Cui et al. 2020a), interspecific variation in shoot flammability at the WUI has yet to be investigated and there is only one published study of shoot flammability for Australian plant assemblages, from tropical rainforests in Queensland (Potts et al. 2022).
We examined shoot flammability patterns among 44 widespread and abundant plant species of the WUI in the Greater Blue Mountains World Heritage Area in New South Wales, Australia. The first aim of our study was to compare shoot flammability among wildland native, and urban native and exotic ornamental plants of residential gardens. Our second aim was to quantify relationships between shoot traits and flammability. Here, we used phylogenetically informed analyses to relate four shoot traits (fuel moisture, bulk density, volume, branching) to four shoot flammability attributes (ignitibility, sustainability, combustibility, consumability). In addressing these first two aims, our work provides important new plant flammability information that enhances our understanding of wildfire dynamics at the WUI. The third aim of our study was to establish a flammability scoring system to distinguish low- from high-flammability species. Our objective was to provide a scientifically robust method for the selection of low-flammability plant species for use in residential gardens designed to act as opportunistic green firebreaks. Garden firebreaks consisting of low-flammability plantings could help to slow or stop the spread of wildfires in urban regions (Mell et al. 2010; Curran et al. 2018; Gibbons et al. 2018; Murray et al. 2018, 2020; Cui et al. 2019; Corbett 2021). At the same time, our method can be used to inform the selective removal of high-flammability species in close proximity to houses in order to reduce WUI wildfire risks.
Methods
Study region
The Greater Blue Mountains World Heritage Area is located approximately 54 km west of Sydney (New South Wales) in eastern Australia. The area possesses globally unique natural values and is heritage listed due to its ecologically significant flora and fauna, natural habitats for the conservation of biodiversity and threatened species of outstanding value (Hammill and Tasker 2010). The Blue Mountains is a fire-prone environment on the urban fringe and at the wildland edge of Sydney, and as such, provides an ideal case study for examining patterns of plant flammability at the WUI (Supplementary Fig. S1). The region has experienced recent, major wildfire disasters in 1994, 2013 and during the Black Summer fires in 2019–2020. There are nearly 80 000 people currently living in the Blue Mountains in 27 towns and villages, with the region covering large parts of the traditional lands of the Dharug and Gundungurra Indigenous people. In 2016, there were an estimated 32 827 private dwellings in the Blue Mountains (BMEE 2018). The area is characterised by a temperate climate with warm summers (mean maximum temperature 29°C) and cool winters (mean minimum temperature 16°C), and a mean annual rainfall of 1399.6 mm (BOM 2020).
The study region spanned the lower-eastern section of the Blue Mountains from the town of Lapstone (33°46′26.4″N, 150°38′13.2″E) to the mid-mountain town of Faulconbridge (33°41′10.8″S, 150°33′0.5″E). Wildland vegetation predominantly found in the immediate vicinity of urban areas at the WUI comprises plant species of dry sclerophyll forests on Hawkesbury sandstone soils of the Greater Sydney region (Fairley and Moore 2010). This fire-prone vegetation, which experiences inter-fire intervals between 10 and 30 years (Hammill and Tasker 2010), is dominated by sclerophyllous species in families including Myrtaceae (e.g. Eucalyptus spp.), Proteaceae (e.g. Banksia spp.) and Fabaceae (e.g. Acacia spp. in the subfamily Mimosoideae, Pultenaea spp. in the subfamily Faboideae).
Study species and shoot collection
We assessed shoot flammability in 44 woody shrub and tree species occurring in the Blue Mountains WUI, including 25 wildland native species, and 19 urban native and exotic ornamental plants of residential gardens (Supplementary Table S1). Among the urban garden species, nine were ornamental species native to Australia and 10 were ornamental species introduced to Australia since permanent European settlement. These study species were selected based on their floristic dominance with respect to their typical high local abundance as identified in a recent census spanning Lapstone to Faulconbridge (>25% projected canopy cover in 100 m2 survey plots; Hawthorne 2021). In total, we sampled wildland native species from across 56 locations in dry sclerophyll vegetation adjacent to roads and in close proximity to urban housing so as to capture those wildland species at the immediate interface between wildland and urban areas, and urban garden species were sampled from across 54 residential gardens in the region (Hawthorne 2021). Typical planted gardens of homes in the Greater Sydney region, which includes the Blue Mountains, consist of a mix of native and exotic ornamental plant species interspersed with lawns (French et al. 2005). All sampling locations were separated by a minimum distance of 100 m.
When sampling, we collected 70 cm long, sun-exposed terminal shoots from each of nine healthy, mature individuals per species following standardised procedures for collection and shoot replication described in Pérez-Harguindeguy et al. (2013), Wyse et al. (2016) and Alam et al. (2020). Sampling of plant shoots took place in autumn (March to May, 2020) in the Southern Hemisphere, and all shoots were collected at least 48 h after rain. To transport samples back to the laboratory, shoots were placed in large heavy-duty garbage bags, which were sealed shut to prevent sample moisture loss. The cut end of the shoot was wrapped in a small piece of cloth dampened with fresh water and fastened with an elastic band. On return to the laboratory, which was set to a temperature of 20°C and relative humidity of 25–30%, all shoot samples were laid out without overlap on a bench to air dry for approximately 24 h. This dry-down procedure was used to match the ignition source to the moisture content of these samples (White and Zipperer 2010; Wyse et al. 2016, 2018).
Measurement of shoot flammability attributes
To measure shoot flammability, we constructed a portable device meeting the specifications described in Jaureguiberry et al. (2011) and modified by Wyse et al. (2016). We burned 70 cm‐long plant shoots using this device, with shoots containing multiple leaves, twigs and small branches to ensure the inclusion of key factors that influence the flammability of vegetation, including fuel arrangement, continuity and quantity (Martin et al. 1994; Alam et al. 2020). We measured four flammability attributes following the conceptual frameworks of Anderson (1970), Martin et al. (1994) and Wyse et al. 2016. For each individual shoot, we measured: (1) ignitibility as time taken for the shoot to begin flaming combustion (time to flame, TTF); (2) sustainability as length of time the shoot spent in flaming combustion (flame duration, FD); (3) combustibility as maximum heat of combustion of the shoot (flame temperature, FT); and (4) consumability as the proportion of original biomass of the shoot that was burnt during the experiment (biomass burnt, BB). Although flammability testing of complete plants is helpful and can provide direct assessment of whole-plant flammability (Etlinger and Beall 2004; Madrigal et al. 2012; Almeida et al. 2022), it is not always feasible to burn replicates of whole plants, such as tall (>10 m) trees like eucalypts, in a safe environment. Assessments of shoot flammability provide an alternative approach and work well as surrogates of whole-plant flammability. For example, Wyse et al. (2016) and Alam et al. (2020) compared their rankings of species’ shoot flammability with rankings of those species derived from the expert opinion of fire managers (Fogarty 2001), and found a good correlation between the two rankings. The expert rankings were based on field observations by 59 fire managers of the burning characteristics of species during wildfires or prescribed burns across New Zealand (Alam et al. 2020).
The LPG-powered flammability device provided heat when lit to radiate from below and through the shoot sample. Each shoot sample was placed horizontally in the device on a mesh grill set 20 cm above the flames, ensuring that overall shoot architecture was kept intact, with the grill temperature maintained at approximately 185°C. During a pre-heating phase of 2 min (Jaureguiberry et al. 2011), the shoot sample’s length, width and height were measured with a tape measure, while the shoot was on the mesh grill, to allow for calculation of sample volume. A blowtorch positioned on the side of the device was then used as an ignition source to apply direct flame to the shoot (Jaureguiberry et al. 2011; Wyse et al. 2016). The blowtorch was switched on for 10 s and during this time, observers determined at which point the shoot ignited (TTF). When the blowtorch was switched off at the end of the 10 s, the duration of time the shoot then spent flaming was recorded (FD). A handheld infrared laser thermometer (Digitech QM 7226) was used at a distance of 50 cm from the sample to measure the maximum temperature (FT) reached by the external flame plume (Jir-Ming and Jun-Hsien 1996; Calitz et al. 2015; Pérez-Harguindeguy et al. 2013; Wyse et al. 2016). The proportion of biomass consumed (BB) by combustion post ignition was visually estimated by at least two observers, following Burger and Bond (2015).
Measurement of shoot traits
We measured four shoot morphological traits, including fuel moisture content, bulk density, volume and degree of ramification, to determine their relative roles in driving variation in shoot flammability. When field-collected shoots were brought to the laboratory, a 10 cm sub-sample was taken from each shoot for fuel moisture assessment. After a 24 h dry-down period matching that of the whole shoot, sub-samples were weighed to determine air-dried mass just prior to measurement of the flammability of whole shoots. Sub-samples were then oven-dried at 60°C for 48 h, after which time they were reweighed. Fuel moisture was expressed as a percentage and calculated as the ratio of the difference between air-dried and oven-dried mass to air-dried dry mass ([air-dried mass − oven-dried mass]/air-dried mass). The volume of each shoot was estimated as an elliptical cylinder and calculated as (π × shoot length × shoot width × 0.5 × shoot height × 0.5), using measurements of length, width and height taken during the pre-heating phase of shoot flammability measurements. Bulk density was calculated as the shoot air-dried mass (measured directly before burning) divided by the shoot volume. The degree of ramification of each shoot was calculated as the number of branches (or nodes) along the main stem line per metre of branch (Santacruz-García et al. 2019). To confirm that the dry-down procedure did not unduly affect flammability measurements, we determined shoot fuel moisture at collection (just prior to the 24 h dry-down period) to compare with shoot fuel moisture just prior to burning in the flammability assessment. We tested the linear relationship between shoot fuel moisture at collection vs burning. If the coefficient of the relationship deviated from 1.0, that would provide evidence that one or more species were skewing the relationship as a function of dry-down differences between shoots with different morphologies. The relationship on testing had a coefficient of 1.02 (linear model), which did not differ significantly from a value of 1 (F1,43 = 2.71, P = 0.12).
Data analysis
We fitted a series of phylogenetic generalised least squares (PGLS) models using species’ mean values as replicates. To account for the unbalanced design among the plant status groups (i.e. different number of species in each group), marginal effects ANOVA was used throughout the analyses. First, to determine whether there were significant differences in each of the shoot flammability attributes among the three plant status groups (wildland native, urban native and urban exotic plants), we built separate PGLS models for each flammability attribute (as a response variable), with plant status as the sole explanatory variable. In these models, TTF was ln transformed, FD and FT square root transformed, and BB logit transformed. Second, we built separate PGLS models for each of the four shoot traits (fuel moisture, shoot volume, bulk density, degree of ramification) as response variables, with plant status as the sole explanatory variable to determine differences in each of the shoot traits among the plant status groups. Third, we built separate PGLS models for each flammability attribute (as a response variable), with the four shoot traits as continuous explanatory variables (bulk density was ln transformed and degree of ramification was ln transformed), to test relationships between the flammability attributes and the shoot traits. In these flammability–trait relationship models, plant status was added last as a categorical explanatory variable. Here, the emergence of a significant effect of plant status indicated that some unmeasured trait or traits that differed among wildland native, urban native and urban exotic plants contributed to variation in shoot flammability in addition to any effects of differences among the groups in the measured plant traits. In contrast, the emergence of a non-significant effect of plant status meant that after the PGLS model accounted for any variation in the flammability attribute due to the four measured shoot traits, none of the residual variation in the models was explained by differences in plant status. Importantly, a non-significant effect of plant status did not diminish any significant differences in the flammability attributes detected in the preceding PGLS analyses with just plant status as the sole explanatory variable. Rather, what these findings reveal is that any effect of plant status on flammability differences among plant groups (with plant status as the sole explanatory variable) can be accounted for by differences between natives and exotics in one or more of the four measured shoot traits.
The phylogeny used in PGLS models (see Fig. S2) was pruned from the tree published in Smith and Brown (2018). To select the appropriate branch length transformation of the phylogenetic tree used in each model, four initial models were fitted including Brownian motion (no transformation) and kappa, delta and lambda transformations estimated by maximum likelihood. Model fit was then assessed using Akaike information criterion (AIC), and the model with lowest AIC was selected (Table S2). In all graphical presentation of results, model parameter estimates were back transformed to the original response units. In any model where a significant effect emerged for plant status (i.e. a significant difference was detected among wildland native, urban native and urban exotic species), post hoc tests (with Tukey’s correction for multiple comparisons) were used to determine where significant pairwise differences lay between status groups (e.g. between wildland native and urban exotic plants). All post hoc tests are presented in Table S3.
To establish a flammability scoring system to distinguish low- from high-flammability species, a principal component analysis (PCA) with oblique rotation (oblimin; Bernaards and Jennrich 2005) was conducted on the flammability attributes TTF, FD, FT (continuous; ln transformed) and BB (proportion; logit transformed), all scaled (by standard deviation) and centred (minus the mean). For the purposes of this analysis, values of TTF were made negative, so that faster TTF values (indicating quicker ignition) had a larger value relative to slower TTF values. Species’ scores for PC1 and PC2 were then plotted on a bivariate graph of PC1 against PC2. The circular mean angle from zero of the PCA loadings on the first two PCA axes was calculated for the four flammability attributes. This mean loading angle for the flammability attributes (scaled between 0 and 1) was overlaid on the plot of the species’ scores. The coordinates of the intersection of a perpendicular line drawn from of each of the species’ scores to the mean loading angle were then calculated. Each set of intersection coordinates for a species was then transformed to a length along the line of the mean loading angle to give a relative flammability score for each species. Species were then assigned to either low-flammability, high-flammability or intermediate-flammability classes by dividing the flammability score distribution into terciles. We built a PGLS model to test flammability scores against plant status and used a G-test to assess the patterns of distribution of the plant groups among the flammability classes.
All analyses were performed in R 4.2.1 (R Core Team 2022) with ape (Paradis and Schliep 2019), caper (Orme et al. 2018), V.PhyloMaker (Jin and Qian 2019), emmeans (Lenth 2022), psych (Revelle 2022) and DescTools (Signorell et al. 2021) packages.
Results
Each of the four flammability attributes – TTF, FD, FT and BB – varied considerably among species (Fig. 1). We found that urban exotic plants had slower TTF than wildland native plants (F2,41 = 4.474, P = 0.017), whereas urban native plants did not differ in TTF from either urban exotic or wildland native plants (Fig. 2a). Urban exotic plants had shorter FD (F2,41 = 7.859, P = 0.001; Fig. 2b), lower FT (F2,41 = 6.471, P = 0.004; Fig. 2c) and lower BB (F2,41 = 7.882, P = 0.001; Fig. 2d) than both wildland and urban native plants. It is interesting to note that for each of the flammability attributes, although many species showed similar levels of variation about the mean, there was evidence for overlap among species, and some species demonstrated more variation (e.g. Citrus limon for TTF and Isopogon anemonifolius for FD), and some species showed less variation (e.g. Bossiaea obcordata for TTF and Pittosporum undulatum for FD) (Fig. 1). In terms of shoot traits, we found that urban exotic plants had higher fuel moisture than both wildland and urban native plants (F2,41 = 8.239, P = 0.001; Fig. 3a). Urban exotic plants had higher bulk density than wildland native plants (F2,41 = 4.177, P = 0.022), whereas urban native plants did not differ in bulk density from either urban exotic or wildland native plants (Fig. 3b). There were no significant differences among the three plant groups in shoot volume (F2,41 = 2.457, P = 0.10; Fig. 3c) or degree of ramification (F2,41 = 1.154, P = 0.33; Fig. 3d).

|

|

|
Our quantification of relationships between shoot flammability attributes and shoot traits revealed several significant relationships (Fig. 4, Table 1). Slower TTF was related to increases in fuel moisture (F1,37 = 24.603, P < 0.001; Fig. 4a) and bulk density (F1,37 = 5.361, P = 0.026; Fig. 4b), but not to shoot volume (F1,37 = 2.419, P = 0.13; Fig. 4c) or degree of ramification (F1,37 = 0.183, P = 0.67; Fig. 4d). Shorter FD was related to decreases in bulk density (F1,37 = 11.587 P = 0.002; Fig. 4f) and shoot volume (F1,37 = 21.736, P < 0.001; Fig. 4g), but not to degree of ramification (F1,37 = 1.265, P = 0.27; Fig. 4h). There was a marginally non-significant relationship between FD and fuel moisture (F1,37 = 3.899, P = 0.06), with a trend for shorter FD associated with increases in fuel moisture (Fig. 4e). Although lower FT was marginally related to increases in fuel moisture (F1,37 = 4.307, P = 0.045; Fig. 4i), it was more strongly related to decreases in bulk density (F1,37 = 6.354, P = 0.016; Fig. 4j) and shoot volume (F1,37 = 15.568, P < 0.001; Fig. 4k); FT was not related to degree of ramification (F1,37 = 0.320, P = 0.57; Fig. 4l). Lower BB was related to increases in fuel moisture (F1,37 = 8.894, P = 0.005; Fig. 4m) and marginally to decreases in shoot volume (F1,37 = 4.240, P = 0.047; Fig. 4o), but not to bulk density (F1,37 = 1.829, P = 0.18; Fig. 4n) or degree of ramification (F1,37 = 0.952, P = 0.34; Fig. 4p).

|
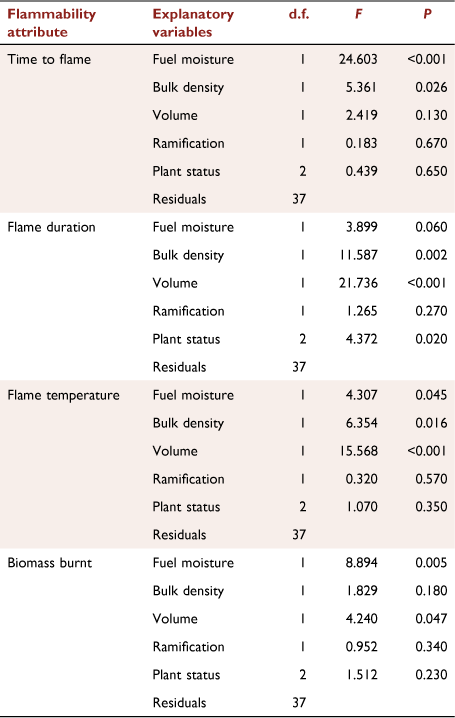
|
Plant status was a non-significant term in the flammability–trait PGLS models for TTF (F2,37 = 0.439, P = 0.65), FT (F2,37 = 1.07, P = 0.35) and BB (F2,37 = 1.512, P = 0.23). This demonstrated that the observed significant effects of plant status in the preceding models of flammability attributes as a function of plant status only were due to differences among plant groups in the measured shoot traits. For TTF, FT and BB, this is mostly attributable to significantly higher fuel moisture in exotic plants (Fig. 3a). In contrast, for FD, the status term was significant (F2,37 = 4.372, P = 0.02), with post hoc tests showing that urban exotic plants had lower FD than wildland native plants (t-ratio = 2.405, P = 0.021), whereas FD was similar between wildland and urban native plants (t-ratio = 1.553, P = 0.13) and urban native plants and urban exotic plants (t-ratio = 1.500, P = 0.14).
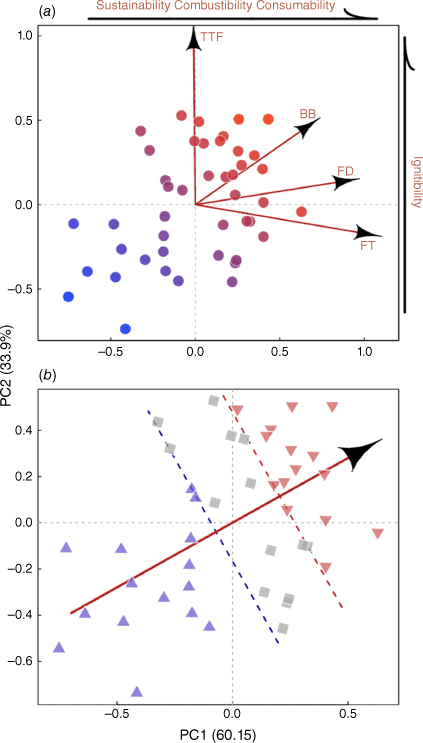
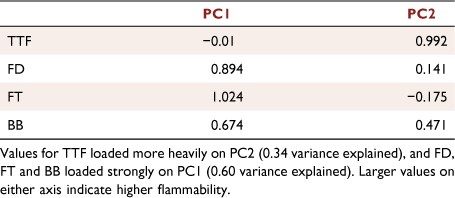
The PCA showed that the three flammability attributes FD, FT and BB loaded strongly on PC1 (0.60 variance explained), whereas TTF loaded more heavily on PC2 (0.34 variance explained) (Fig. 5a, Table 2). The mean angle calculated from the PCA loadings of the four flammability attributes was 61°, with the high-flammability class of species falling in the top right of the PCA and the low-flammability class of species in the bottom left (Fig. 5b). All urban exotic species fell in the low-flammability class, with the exception of Murraya paniculata and Jacaranda mimosifolia, which fell in the high-flammability class. Both wildland and urban native species were relatively evenly distributed across the intermediate-flammability and high-flammability classes (Fig. 6a). Across all species (Fig. 6a), the least flammable was Magnolia grandiflora and the most flammable was Callistemon citrinus. For wildland native plants, the least flammable species was Pittosporum undulatum and the most flammable species was Corymbia gummifera. For urban native plants, the least flammable species was Melaleuca quinquenervia and the most flammable species was C. citrinus. For urban exotic plants, the least flammable species was M. grandiflora and the most flammable was J. mimosifolia. Relative flammability scores were lower for urban exotic plants (F2,41 = 11.534, P < 0.001), relative to wildland and urban native plants, which did not differ significantly from each other (Fig. 6b), with a disproportionately higher number of urban exotic species falling in the low-flammability class (χ2 = 14.780, d.f. = 4, P = 0.005; Fig. 6c).
|
|

|
Discussion
Patterns of flammability among plants of the WUI
Our results reveal important shoot flammability differences between native and exotic plant species of the WUI in the Greater Blue Mountains World Heritage Area. For all four of the shoot flammability attributes that we measured (TTF, FD, FT, BB), we found that urban exotic plants of residential gardens were less flammable than native plants in surrounding wildland vegetation. Our finding for lower shoot flammability in urban exotic plants is likely explained by the comparatively higher fuel moisture content we identified in shoots of exotic species (Fig. 3). Indeed, inspection of the interspecific relationships between shoot fuel moisture and each of the four flammability attributes shows that exotic species occupy the higher moisture–lower flammability areas of the graphs (Fig. 4). Few other studies have compared shoot flammability between native and exotic plants (Wyse et al. 2016, 2018; Msweli et al. 2020; Potts et al. 2022). Although not explicitly setting out to compare native vs exotic flammability, Wyse et al. (2016) found no consistent native–exotic differences in their New Zealand study species; however, the most flammable species of the 60 they tested was the invasive exotic Gorse (Ulex europaeus). Subsequent testing of 194 species in New Zealand found native grasses and exotic species (Pinus ponderosa) had among the highest shoot flammability (Cui et al. 2020b). Potts et al. (2022) found invasive shrubs had relatively low flammability compared with other groups combining growth form (trees, shrubs, vines) and successional status (canopy, understorey, pioneer) in Australian tropical rainforests. In contrast, Msweli et al. (2020) found that shoot flammability was generally highest in invasive exotic plants. Our findings, considered together with this previous work, suggest that it is highly likely that differences in shoot flammability between native and exotic plant species will be regionally context dependent and reliant on underlying native–exotic differences in traits that determine the flammability of species. For our study system of dry sclerophyll vegetation in eastern Australia, the fundamentally higher fuel moisture content in shoots of exotic species leads to their overall lower flammability than native plants. In other ecosystems, where these sorts of shoot traits do not differ between native and exotic species, we would not expect to see native–exotic differences in shoot flammability.
Plants in urban gardens in the Blue Mountains region are typically well watered by residents, particularly when compared with wildland plants in surrounding dry sclerophyll vegetation. Might this difference in watering conditions have had an effect on observed native–exotic differences in plant flammability? We suggest that this is unlikely to play a major role for three of the flammability attributes (FD, FT, BB), because if this had been in effect, we would have expected to see urban native garden plants demonstrate lower flammability than wildland native species, which was not the case. However, our findings suggest that we might have observed a small effect of well watered gardens for TTF. Here, urban native species did not differ significantly from either urban exotic plants or wildland native plants. In general, however, it appears more likely that the intrinsic properties of the different plant groups, most notably shoot water content in exotics, play a predominant role in generating observed native/exotic differences. Our results might be seen as lending support to the notion that plant species of fire-prone communities have evolved characteristics that increase their propensity to burn (Mutch 1970; Bond and Midgley 1995), particularly considering the comparatively high flammability demonstrated by wildland native species. Any support for this hypothesis on the basis of our findings must be viewed cautiously, however, because traits conferring high flammability might simply be exaptations (Snyder 1984; Bowman et al. 2014; Krix and Murray 2018).
Trait relationships with shoot flammability
The quantitative modelling we performed in an explicitly phylogenetic context identified some important relationships between shoot traits and shoot flammability. We found that higher fuel moisture content in shoots was linked to low flammability exhibited primarily by increases in TTF and decreases in BB. Higher moisture content was also somewhat related to decreases in FD and FT, with the former marginally non-significant (P = 0.06) and the latter marginally significant (P = 0.045). In general, our findings are consistent with previous studies of shoot flammability (Wyse et al. 2018; Cui et al. 2020a; Msweli et al. 2020). Our results support the notion of a functional role for water content in lengthening TTF because more time is required for sufficient energy input to remove more water before flaming can commence. High shoot fuel moisture continues to provide a dampening effect on flame initiation and spread, which ultimately limits BB and to some extent FD and FT, with fuel moisture relationships with FD and FT being viewed cautiously.
We also found that lower shoot bulk density was related to decreases in TTF, FD and FT. Previous research on shoot flammability has found no significant relationship between bulk density and shoot flammability (Padullés Cubino et al. 2018; Wyse et al. 2018). The results of our study suggest that increases in bulk density lead to decreasing shoot flammability with respect to TTF, yet at the same time lead to an increase in shoot flammability with respect to FD and FT. A larger shoot mass within a given area provides for less aeration for flames, delaying ignition (Dickinson and Kirkpatrick 1985). However, once the shoot starts to burn, flames can spread more easily to other close parts of the shoot, with the greater mass available to burn leading to longer and hotter fires. It is unclear at this stage why the effect of higher shoot bulk density did not extend to increasing the amount of shoot consumed in flames.
To our knowledge, no previous studies have analysed the relationship between shoot volume and flammability, but our analyses showed that higher shoot volume was significantly related to increases in FD, FT and BB (albeit marginally, P = 0.047), but not to TTF. All else being equal, small- or large-volume shoots have the same likelihood of ignition, which suggests aeration required to reach flaming combustion is independent of shoot size (but not bulk density; see above). Once alight, however, the overall higher volume of shoot that can burn leads to longer and hotter fires with more of the shoot consumed in flames. In contrast to previous work (Santacruz-García et al. 2019), we found no significant correlations between degree of ramification and any of the flammability attributes. It is difficult to capture the architectural complexity of plant canopies using one such simple metric, and in future it may be worth including measures that includes not only the number of branches, but also biomass in different size classes (Burger and Bond 2015) or the density of leaves on a shoot (Calitz et al. 2015), which also contribute to the complex structure of a canopy.
Intriguingly, the PGLS analyses of trait–flammability relationships revealed that plant status was a significant explanatory variable only in the model for FD, with wildland native plants exhibiting longer FD than urban exotic plants. This would suggest that some unmeasured trait that differs between wildland native and urban exotic plants contributed to variation in shoot flammability – in addition to any effects of the plant traits that we included in our study. We suggest that a candidate trait is the presence of volatile organic compounds (VOCs) such as terpenoids, particularly in species within the family Myrtaceae, a family well known for its VOCs (Whiffin and Ladiges 1992; Keszei et al. 2008). Previous work has identified links between flammability and VOCs (Pausas et al. 2016; Romero and Ganteaume 2021). Indeed, Della Rocca et al. (2017) suggested that terpenoids should be included in physical models of the prediction and propagation of wildfire in mediterranean vegetation given their strong influence on plant flammability. To further unpack patterns of flammability among species at the WUI, particularly in fire-prone regions like eastern Australia where the evolution of the flora has been heavily influenced by wildfires, future studies exploring the contribution of VOCs to shoot flammability will be valuable.
Identifying low- and high-flammability plant species
Wildfire impacts on lives and homes is a serious concern for residents living at the WUI in fire-prone areas is (Gill and Stephens 2009). Robust, evidence-based information on the relative flammability of plant species at the WUI (White and Zipperer 2010) is required to guide the selection of low-flammability plant species for opportunistic firebreaks in residential gardens (Mell et al. 2010; Curran et al. 2018; Murray et al. 2018). Green firebreaks have been used extensively in China to help slow or stop the spread of wildfires (Cui et al. 2019). There is a growing body of scientific research from different regions of the world exploring and discussing the use of green firebreaks for wildfire management (Santamarta-Cerezal et al. 2012; Bowman et al. 2017b; Souza and Vale 2019; Batista et al. 2021; Chifa 2021; Khan and Moinuddin 2021). A key premise of the use of green firebreaks is the understanding that all plants are likely to burn under extreme wildfire conditions, but under less intense wildfire conditions the presence of low-flammability plants, which do not ignite quickly and only burn for short durations at low temperatures, can provide a level of protection for residents, their homes and vital infrastructure. Although many organisations provide lists recommending apparent low-flammability species for home gardens, such lists can be problematic because: their method of collation is often unclear; they can be contradictory; most times they lack reference to scientific research; and they can provide a false sense of security by implying that species-specific flammability is known (White and Zipperer 2010; Corbett 2021). Our third aim in this study was to overcome the problems associated with recommendation lists by establishing a robust flammability scoring system to distinguish low- from high-flammability species to guide the selection of low-flammability plant species for use in residential green firebreaks at the WUI. At the same time, our method was devised to help guide the selective removal of high-flammability species in close proximity to houses in order to reduce wildfire risks at the WUI.
In this context, although the overall lower intrinsic flammability of urban exotic plants suggests that these species would be suitable candidates for low-flammability residential gardens, we prefer to support the use of native plant species in urban plantings, over a generalisation that exotic plant species are most suitable, for three reasons. First, two of the exotic species in this study, jacaranda (J. mimosifolia) and orange jasmine (M. paniculata), displayed properties of high-flammability species (e.g. jacaranda ignited within 2 s, and both species burnt on average for more than 20 s at a peak of 500°C and had much higher shoot biomass consumed in flames). Our results would suggest that jacaranda, especially, would not be a good species for reducing fire risks in urban areas. Second, although the moist foliage of exotic species might display low-flammability properties, their dried leaves can be highly flammable when compared with coexisting native species (Murray et al. 2013). Third, exotic plant species that are invasive or have the potential to become invasive (e.g. ‘sleeper weeds’) should be ruled out of consideration for low-flammability gardens given their enormous ecological and economic impacts (IPBES 2019). Indeed, exotic plant invasion can potentially increase flammability causing successional changes of vegetation from less flammable forest to more flammable shrubland or forest–shrub landscape mosaics (Perry et al. 2015). Furthermore, there is a wealth of evidence demonstrating a range of detrimental ecological impacts that exotic species can have on native biodiversity (Davies 2011; Vilà et al. 2011; Jauni and Ramula 2015; Nguyen et al. 2016; Fletcher et al. 2019). All of these issues considered together warn against a reliance on exotic plant species in the design and implementation of green firebreaks.
We found that two of the urban native species fell in the low-flammability class, suggesting their possible use in green firebreaks. Broad-leaved paperbark (Melaleuca quinquenervia) and brush box (Lophostemon confertus) both had relatively short flame durations of less than 10 s, with their short sustainability matched with flame temperatures generally below 500°C (Fig. 1). These two species also took moderately long times to ignite, averaging around 4–5 s before commencing flaming combustion. Brush box might be a candidate low-flammability native species for residential garden firebreaks, but broad-leaved paperbark should not be considered for such a purpose. This species is generally considered highly flammable at the whole plant level, due to its dry, shaggy outer layers of papery bark, which readily provide a ladder fuel that can quickly carry fire into the canopy, destroying leaves and branches (Munger 2005). In addition, wildfire embers can easily get caught in the roughly organised bark (Webster 2000). This provides a good example of the need to look at other plant characteristics, not just shoot-level flammability, when designing low-flammability gardens (Gill and Zylstra 2005; Schwilk 2015). Indeed, previous work has shown that that the accumulation of dead branches in the canopy of plants and lots of leaf litter under the plant further increases their flammable properties (Schwilk 2003). Several of the wildland native species also fell in the low-flammability class, including Pittosporum undulatum, Banksia ericifolia, Petrophile pulchella, Grevillea phylicoides and Acacia linifolia. These are all attractive native plants, worthy of further investigation as potential species for low-flammability urban plantings. In terms of high-flammability urban native species that should be avoided, crimson bottlebrush (C. citrinus) ignited within 2 s, sustained flaming combustion for 30 s on average and burned at temperatures around 600°C (Fig. 1). Other urban native species with comparably high flammability included Banks’ grevillea (Grevillea banksii) and blueberry ash (Elaeocarpus reticulatus) (Fig. 6a). These native species should not be planted near houses if the desired outcome is to create low-flammability residential gardens.
Conclusion
Overall, our findings suggest that native plants currently found at the Blue Mountains WUI, whether they are in wildland vegetation or ornamentals in home gardens, could exacerbate the risks of ignition to houses, which has important ramifications for the establishment of low-flammability residential gardens as opportunistic firebreaks within the WUI in the region. Native plant selection for opportunistic garden firebreaks needs to be performed carefully to avoid the high-flammability native plants. Our flammability scoring has demonstrated its utility for this purpose by distinguishing low-flammability native species such as brush box (L. confertus) from high-flammability species such as crimson bottlebrush (C. citrinus). Our study represents an initial screening of the flammability of a wide range of species in the WUI of the Greater Sydney region. We recommend that subsequent studies should focus on seasonal changes in the flammability of WUI species, especially those identified as being low in flammability in this study.
Building on our findings, future work might focus its attention on identifying a broader suite of low-flammability native plant species for urban gardens, to remove any reliance on exotic plants (given their potential for detrimental ecological effects) as the most suitable species with fire-retardant properties. To this end, our functional trait analysis could be used to direct searches to species with shoots with high fuel moisture content because this is a trait found relatively consistently in our low-flammability species, particularly with respect to increasing time to ignition. If the desired outcome is to have garden plants that do not maintain flaming combustion for long periods, and at comparatively low temperatures, selection targets should be those species with low bulk density and volume. The issue of how long a plant burns is an important one, because although all plants will ignite given hot enough fire fronts, recent work has shown that species maintain idiosyncratic burn durations of their leaves as radiant heat increases (Krix et al. 2022). That is, species with leaves that burn for a long time will do so at both low and high values of radiant heat. In addition, future work might seek to determine from a broader suite of shoot and plant traits the most important traits underpinning flammability to build predictive models (e.g. see Krix and Murray 2022 for leaf-level models). Such predictive models will mean that specialised flammability assessment equipment need not be used to measure flammability directly, but rather, simple-to-measure shoot traits can be used to determine the flammability of a large number of species for many regions of the world.
Finally, urban garden design involves more than just selecting plants that are low in flammability. It is important to recognise that features such as the biodiversity-promoting value of native plant species is an issue that needs to be considered, along with the ability of plant species to cope with climate change and be aesthetically pleasing in home gardens. To this end, broad selection frameworks (Alam 2020, Murray 2018) can be used to take into account the range of selective factors that come into play for people and their gardens.
Supplementary material
Supplementary material is available online.
Data availability
Species flammability and trait data are available in Supplementary Fig. S1.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This research was funded by the Faculty of Science, University of Technology Sydney, NSW.
Acknowledgements
We thank the technical staff in the Faculty of Science at UTS for their assistance with the experimental shoot burns and Greg Dalsanto at the UTS workshop for help with building the portable flammability device. We are grateful to Mel Havelock for helpful comments on a draft of the manuscript.
References
Alam MA, Wyse SV, Buckley HL, Perry GLW, Sullivan JJ, Mason NWH, Buxton R, Richardson SJ, Curran TJ (2020) Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits. Journal of Ecology 108, 641–653.| Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits.Crossref | GoogleScholarGoogle Scholar |
Almeida M, Reza Modarres M, Muñoz JA, Ribeiro LM (2022) Flammability characteristics of typical garden species. In ‘Advances in Forest Fire Research 2022. Chapter 2 – Fire at the wildland urban interface’. (Eds DX Viegas, LM Ribeiro) pp. 602–609. (University of Coimbra, Portugal)
| Crossref |
Anderson HE (1970) Forest fuel ignitibility. Fire Technology 6, 312–319.
| Forest fuel ignitibility.Crossref | GoogleScholarGoogle Scholar |
Batista AC, Biondi D, Martini A (2021) Flammability of ornamental species for fire management in wildland-urban interface in Paraná State. Floresta 51, 192–200.
| Flammability of ornamental species for fire management in wildland-urban interface in Paraná State.Crossref | GoogleScholarGoogle Scholar |
Bernaards CA, Jennrich RI (2005) Gradient projection algorithms and software for arbitrary rotation criteria in factor analysis. Educational and Psychological Measurement 65, 676–696.
| Gradient projection algorithms and software for arbitrary rotation criteria in factor analysis.Crossref | GoogleScholarGoogle Scholar |
Blackhall M, Raffaele E (2019) Flammability of Patagonian invaders and natives: when exotic plant species affect live fine fuel ignitability in wildland-urban interfaces. Landscape and Urban Planning 189, 1–10.
| Flammability of Patagonian invaders and natives: when exotic plant species affect live fine fuel ignitability in wildland-urban interfaces.Crossref | GoogleScholarGoogle Scholar |
BMEE (2018) ‘Blue Mountains demographic and economic profile.’ (Blue Mountains Economic Enterprise: Katoomba, NSW, Australia)
BOM (2020) Australian Bureau of Meteorology Annual Climate Statement 2019. Available at http://www.bom.gov.au/climate/current/annual/aus/[Accessed 17 August 2020]
Bond WJ, Midgley JJ (1995) Kill Thy Neighbour: an individualistic argument for the evolution of flammability. Oikos 73, 79–85.
| Kill Thy Neighbour: an individualistic argument for the evolution of flammability.Crossref | GoogleScholarGoogle Scholar |
Bowman DMJS, French BJ, Prior LD (2014) Have plants evolved to self-immolate? Frontiers in Plant Science 5, 590
| Have plants evolved to self-immolate?Crossref | GoogleScholarGoogle Scholar |
Bowman DMJS, Williamson GJ, Abatzoglou JT, Kolden CA, Cochrane MA, Smith AM (2017a) Human exposure and sensitivity to globally extreme wildfire events. Nature Ecology and Evolution 1, 0058
| Human exposure and sensitivity to globally extreme wildfire events.Crossref | GoogleScholarGoogle Scholar |
Bowman DMJS, Garnett ST, Barlow S, Bekessy SA, Bellairs SM, Bishop MJ, Bradstock RA, Jones DN, Maxwell SL, Pittock J, Toral-Granda MV, Watson JEM, Wilson T, Zander KK, Hughes L (2017b) Renewal ecology: conservation for the Anthropocene. Restoration Ecology 25, 674–680.
| Renewal ecology: conservation for the Anthropocene.Crossref | GoogleScholarGoogle Scholar |
Burger N, Bond WJ (2015) Flammability traits of cape shrubland species with different post-fire recruitment strategies. South African Journal of Botany 101, 40–48.
| Flammability traits of cape shrubland species with different post-fire recruitment strategies.Crossref | GoogleScholarGoogle Scholar |
Burton JE, Cawson JG, Filkov AI, Penman TD (2021) Leaf traits predict global patterns in the structure and flammability of forest litter beds. Journal of Ecology 109, 1344–1355.
| Leaf traits predict global patterns in the structure and flammability of forest litter beds.Crossref | GoogleScholarGoogle Scholar |
Calitz W, Potts AJ, Cowling RM (2015) Investigating species-level flammability across five biomes in the Eastern Cape, South Africa. South African Journal of Botany 101, 32–39.
| Investigating species-level flammability across five biomes in the Eastern Cape, South Africa.Crossref | GoogleScholarGoogle Scholar |
Chifa D (2021) The design of green firebreaks in Portuguese forest: a case study of Alferce, Monchique. MSc Thesis, Estonian University of Life Science, Tartu, Estonia.
Corbett L (2021) ‘Safer gardens: plant flammability and planning for fire.’ (Australian Scholarly Publishing: Vic., Australia)
Cui X, Alam MA, Perry GLW, Paterson AM, Wyse SV, Curran TJ (2019) Green firebreaks as a management tool for wildfires: lessons from China. Journal of Environmental Management 233, 329–336.
| Green firebreaks as a management tool for wildfires: lessons from China.Crossref | GoogleScholarGoogle Scholar |
Cui X, Paterson AM, Alam MA, Wyse SV, Marshall K, Perry GLW, Curran TJ (2020a) Shoot-level flammability across the Dracophyllum (Ericaceae) phylogeny: evidence for flammability being an emergent property in a land with little fire. New Phytologist 228, 95–105.
| Shoot-level flammability across the Dracophyllum (Ericaceae) phylogeny: evidence for flammability being an emergent property in a land with little fire.Crossref | GoogleScholarGoogle Scholar |
Cui X, Paterson AP, Wyse SV, Alam MA, Maurin K, Pieper R, Padullés Cubino J, O’Connell D, Donkers D, Bréda J, Buckley HL, Perry GLW, Curran TJ (2020b) Shoot flammability of vascular plants is phylogenetically conserved and related to habitat fire-proneness and growth form. Nature Plants 6, 355–359.
| Shoot flammability of vascular plants is phylogenetically conserved and related to habitat fire-proneness and growth form.Crossref | GoogleScholarGoogle Scholar |
Curran TJ, Perry GLW, Wyse SV, Alam MA (2018) Managing fire and biodiversity in the wildland-urban interface: a role for green firebreaks. Fire 1, 3
| Managing fire and biodiversity in the wildland-urban interface: a role for green firebreaks.Crossref | GoogleScholarGoogle Scholar |
Davies KW (2011) Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 167, 481–491.
| Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass.Crossref | GoogleScholarGoogle Scholar |
Della Rocca G, Madrigal J, Marchi E, Michelozzi M, Moya B, Danti R (2017) Relevance of terpenoids on flammability of Mediterranean species: an experimental approach at a Low radiant heat flux. iForest 10, 766–775.
| Relevance of terpenoids on flammability of Mediterranean species: an experimental approach at a Low radiant heat flux.Crossref | GoogleScholarGoogle Scholar |
Dickinson KJM, Kirkpatrick JB (1985) The flammability and energy content of some important plant species and fuel components in the forests of southeastern Tasmania. Journal of Biogeography 12, 121–134.
| The flammability and energy content of some important plant species and fuel components in the forests of southeastern Tasmania.Crossref | GoogleScholarGoogle Scholar |
Etlinger MG, Beall FC (2004) Development of a laboratory protocol for fire performance of landscape plants. International Journal of Wildland Fire 13, 479–488.
| Development of a laboratory protocol for fire performance of landscape plants.Crossref | GoogleScholarGoogle Scholar |
Fairley A, Moore PM (2010) ‘Native plants of the Sydney region.’ (Jacana Books: Crows Nest, NSW, Australia)
Fares S, Bajocco S, Salvati L, Camarretta N, Dupuy J-L, Xanthopoulos G, Guijarro M, Madrigal J, Hernando C, Corona P (2017) Characterizing potential wildland fire fuel in live vegetation in the Mediterranean region. Annals of Forest Science 74, 1
| Characterizing potential wildland fire fuel in live vegetation in the Mediterranean region.Crossref | GoogleScholarGoogle Scholar |
Fletcher RA, Brooks RK, Lakoba VT, Sharma G, Heminger AR, Dickinson CC, Barney JN (2019) Invasive plants negatively impact native, but not exotic, animals. Global Change Biology 25, 3694–3705.
| Invasive plants negatively impact native, but not exotic, animals.Crossref | GoogleScholarGoogle Scholar |
Fogarty LG (2001) ‘A flammability guide for some common New Zealand native tree and shrub species’. Forest Research Bulletin 197, Forest and Rural Fire Scientific and Technical Series Report 6. pp. 1–18. (Forest Research Institute in Association with the New Zealand Fire Service Commission and National Rural Fire Authority: Rotorua, Wellington, New Zealand)
French K, Major R, Hely K (2005) Use of native and exotic garden plants by suburban nectarivorous birds. Biological Conservation 121, 545–559.
| Use of native and exotic garden plants by suburban nectarivorous birds.Crossref | GoogleScholarGoogle Scholar |
Ganteaume A, Jappiot M, Lampin C, Guijarro M, Hernando C (2013) Flammability of some ornamental species in wildland–urban interfaces in Southeastern France: laboratory assessment at particle level. Environmental Management 52, 467–480.
| Flammability of some ornamental species in wildland–urban interfaces in Southeastern France: laboratory assessment at particle level.Crossref | GoogleScholarGoogle Scholar |
Gao X, Schwilk DW (2022) Burn hot or tolerate trees: flammability decreases with shade tolerance in grasses. Oikos 2022, e08930
| Burn hot or tolerate trees: flammability decreases with shade tolerance in grasses.Crossref | GoogleScholarGoogle Scholar |
Gibbons P, Gill AM, Shore N, Moritz MA, Dovers S, Cary GJ (2018) Options for reducing house-losses during wildfires without clearing trees and shrubs. Landscape and Urban Planning 174, 10–17.
| Options for reducing house-losses during wildfires without clearing trees and shrubs.Crossref | GoogleScholarGoogle Scholar |
Gill AM, Stephens SL (2009) Scientific and social challenges for the management of fire-prone wildland–urban interfaces. Environmental Research Letters 4, 034014
| Scientific and social challenges for the management of fire-prone wildland–urban interfaces.Crossref | GoogleScholarGoogle Scholar |
Gill AM, Zylstra P (2005) Flammability of Australian forests. Australian Forestry 68, 87–93.
| Flammability of Australian forests.Crossref | GoogleScholarGoogle Scholar |
Hammill K, Tasker E (2010) ‘Vegetation, fire and climate change in the greater Blue Mountains world heritage area.’ (NSW Department of Environment, Climate Change and Water: Sydney, NSW, Australia)
Hawthorne T (2021) Patterns in plant flammability at a fire-prone wildland-urban interface in eastern Australia. MSc Thesis, University of Technology Sydney, Ultimo, NSW, Australia.
He T, Lamont BB, Pausas JG (2019) Fire as a key driver of earth’s biodiversity. Biological Reviews 94, 1983–2010.
| Fire as a key driver of earth’s biodiversity.Crossref | GoogleScholarGoogle Scholar |
IPBES (2019) ‘Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services.’ (IPBES Secretariat: Bonn, Germany)
Jauni M, Ramula S (2015) Meta-analysis on the effects of exotic plants on the fitness of native plants. Perspectives in Plant Ecology, Evolution and Systematics 17, 412–420.
| Meta-analysis on the effects of exotic plants on the fitness of native plants.Crossref | GoogleScholarGoogle Scholar |
Jaureguiberry P, Bertone G, Díaz S (2011) Device for the standard measurement of shoot flammability in the field. Austral Ecology 36, 821–829.
| Device for the standard measurement of shoot flammability in the field.Crossref | GoogleScholarGoogle Scholar |
Jin Y, Qian H (2019) V.PhyloMaker: an R Package that can generate very large phylogenies for vascular plants. Ecography 42, 1353–1359.
| V.PhyloMaker: an R Package that can generate very large phylogenies for vascular plants.Crossref | GoogleScholarGoogle Scholar |
Jir-Ming C, Jun-Hsien Y (1996) The measurement of open propane flame temperature using infrared technique. Journal of Quantitative Spectroscopy and Radiative Transfer 56, 133–144.
| The measurement of open propane flame temperature using infrared technique.Crossref | GoogleScholarGoogle Scholar |
Keszei A, Brubaker CL, Foley WJ (2008) A molecular perspective on terpene variation in Australian Myrtaceae. Australian Journal of Botany 56, 197–213.
| A molecular perspective on terpene variation in Australian Myrtaceae.Crossref | GoogleScholarGoogle Scholar |
Khan N, Moinuddin K (2021) The role of heat flux in an idealised firebreak built in surface and crown fires. Atmosphere 12, 1395
| The role of heat flux in an idealised firebreak built in surface and crown fires.Crossref | GoogleScholarGoogle Scholar |
Kraaij T, Msweli ST, Potts AJ (2022) Fuel trait effects on flammability of native and invasive alien shrubs in coastal fynbos and thicket (Cape Floristic Region). PeerJ 10, e13765
| Fuel trait effects on flammability of native and invasive alien shrubs in coastal fynbos and thicket (Cape Floristic Region).Crossref | GoogleScholarGoogle Scholar |
Krix DW, Murray BR (2018) Landscape variation in plant leaf flammability is driven by leaf traits responding to environmental gradients. Ecosphere 9, e02093
| Landscape variation in plant leaf flammability is driven by leaf traits responding to environmental gradients.Crossref | GoogleScholarGoogle Scholar |
Krix DW, Murray BR (2022) A predictive model of leaf flammability using leaf traits and radiant heat flux for plants of fire-prone dry sclerophyll forest. Forests 13, 152
| A predictive model of leaf flammability using leaf traits and radiant heat flux for plants of fire-prone dry sclerophyll forest.Crossref | GoogleScholarGoogle Scholar |
Krix DW, Phillips ML, Murray BR (2019) Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland-urban interface. International Journal of Wildland Fire 28, 295–307.
| Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland-urban interface.Crossref | GoogleScholarGoogle Scholar |
Krix DW, Murray ML, Murray BR (2022) Increasing radiant heat flux affects leaf flammability patterns in plant species of eastern Australian fire-prone woodlands. Plant Biology 24, 302–312.
| Increasing radiant heat flux affects leaf flammability patterns in plant species of eastern Australian fire-prone woodlands.Crossref | GoogleScholarGoogle Scholar |
Lenth RV (2022) emmeans: Estimated Marginal Means, aka Least-Squares Means. Available at https://CRAN.R-project.org/package=emmeans [verified 12 August 2022]
Madrigal J, Marino E, Guijarro M, Hernando C, Díez C (2012) Evaluation of the flammability of gorse (Ulex europaeus L.) managed by prescribed burning. Annals of Forest Science 69, 387–397.
| Evaluation of the flammability of gorse (Ulex europaeus L.) managed by prescribed burning.Crossref | GoogleScholarGoogle Scholar |
Martin RE, Gordon DA, Gutierrez MA, Lee DS, Molina DM, Schroeder RA, Sapsis DB, Stephens SL, Chambers M (1994) Assessing the Flammability of Domestic and Wildland Vegetation. In ‘Proceedings of the 12th conference on fire and forest meteorology’. pp. 130–137. (Society of American Foresters: Bethesda, MD, USA)
McWethy DB, Schoennagel T, Higuera PE, Krawchuk M, Harvey BJ, Metcalf EC, Schultz C, Miller C, Metcalf AL, Buma B, Virapongse A, Kulig JC, Stedman RC, Ratajczak Z, Nelson CR, Kolden C (2019) Rethinking resilience to wildfire. Nature Sustainability 2, 797–804.
| Rethinking resilience to wildfire.Crossref | GoogleScholarGoogle Scholar |
Mell WE, Manzello SL, Maranghides A, Butry D, Rehm RG (2010) The wildland–urban interface fire problem – current approaches and research needs. International Journal of Wildland Fire 19, 238–251.
| The wildland–urban interface fire problem – current approaches and research needs.Crossref | GoogleScholarGoogle Scholar |
Molina JR, Martín T, Rodríguez Y Silva F, Herrera MÁ (2017) The ignition index based on flammability of vegetation improves planning in the wildland-urban interface: a case study in southern Spain. Landscape and Urban Planning 158, 129–138.
| The ignition index based on flammability of vegetation improves planning in the wildland-urban interface: a case study in southern Spain.Crossref | GoogleScholarGoogle Scholar |
Moritz MA, Hazard R, Johnston K, Mayes M, Mowery M, Oran K, Parkinson A-M, Schmidt DA, Wesolowski G (2022) Beyond a focus on fuel reduction in the WUI: the need for regional wildfire mitigation to address multiple risks. Frontiers in Forests and Global Change 5, 848254
| Beyond a focus on fuel reduction in the WUI: the need for regional wildfire mitigation to address multiple risks.Crossref | GoogleScholarGoogle Scholar |
Msweli ST, Potts AJ, Fritz H, Kraaij T (2020) Fire weather effects on flammability of indigenous and invasive alien plants in coastal fynbos and thicket shrublands (Cape Floristic Region). PeerJ 8, e10161
| Fire weather effects on flammability of indigenous and invasive alien plants in coastal fynbos and thicket shrublands (Cape Floristic Region).Crossref | GoogleScholarGoogle Scholar |
Munger GT (2005) Melaleuca quinquenervia. In ‘Fire effects information system’. (US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory) Available at https://www.fs.usda.gov/database/feis/plants/tree/maggra/all.html [verified 20, April 2023].
Murray BR, Hardstaff LK, Phillips ML (2013) Differences in leaf flammability, leaf traits and flammability-trait relationships between native and exotic plant species of dry sclerophyll forest. PLoS One 8, e79205
| Differences in leaf flammability, leaf traits and flammability-trait relationships between native and exotic plant species of dry sclerophyll forest.Crossref | GoogleScholarGoogle Scholar |
Murray BR, Martin LJ, Brown C, Krix DW, Phillips ML (2018) Selecting low-flammability plants as green firebreaks within sustainable urban garden design. Fire 1, 15
| Selecting low-flammability plants as green firebreaks within sustainable urban garden design.Crossref | GoogleScholarGoogle Scholar |
Murray BR, Brown C, Murray ML, Krix DW, Martin LJ, Hawthorne T, Wallace MI, Potvin SA, Webb JK (2020) An integrated approach to identify low-flammability plant species for green firebreaks. Fire 3, 9
| An integrated approach to identify low-flammability plant species for green firebreaks.Crossref | GoogleScholarGoogle Scholar |
Mutch RW (1970) Wildland fires and ecosystems: a hypothesis. Ecology 51, 1046–1051.
| Wildland fires and ecosystems: a hypothesis.Crossref | GoogleScholarGoogle Scholar |
Nguyen KQ, Cuneo P, Cunningham SA, Krix DW, Leigh A, Murray BR (2016) Ecological effects of increasing time since invasion by the exotic African olive (Olea europaea ssp. cuspidata) on leaf-litter invertebrate assemblages. Biological Invasions 18, 1689–1699.
| Ecological effects of increasing time since invasion by the exotic African olive (Olea europaea ssp. cuspidata) on leaf-litter invertebrate assemblages.Crossref | GoogleScholarGoogle Scholar |
Nolan RH, Bowman DMJS, Clarke H, Haynes K, Ooi MKJ, Price OF, Williamson GJ, Whittaker J, Bedward M, Boer MM, Cavanagh VI, Collins L, Gibson RK, Griebel A, Jenkins ME, Keith DA, Mcilwee AP, Penman TD, Samson SA, Tozer MG, Bradstock RA (2021) What do the Australian black summer fires signify for the global fire crisis? Fire 4, 97
| What do the Australian black summer fires signify for the global fire crisis?Crossref | GoogleScholarGoogle Scholar |
Orme D, Freckleton R, Thomas G, Petzoldt T, Fritz S, Isaac N, Pearse W (2018) caper: Comparative Analyses of Phylogenetics and Evolution in R_. R package version 1.0.1. Available at https://CRAN.R-project.org/package=caper [verified 25 November 2022]
Padullés Cubino J, Buckley HL, Day NJ, Pieper R, Curran TJ (2018) Community‐level flammability declines over 25 years of plant invasion in grasslands. Journal of Ecology 106, 1582–1594.
| Community‐level flammability declines over 25 years of plant invasion in grasslands.Crossref | GoogleScholarGoogle Scholar |
Paradis E, Schliep K (2019) ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528.
| ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R.Crossref | GoogleScholarGoogle Scholar | [verified 25 November 2022]
Pausas JG, Alessio GA, Moreira B, Segarra-Moragues JG (2016) Secondary compounds enhance flammability in a Mediterranean plant. Oecologia 180, 103–110.
| Secondary compounds enhance flammability in a Mediterranean plant.Crossref | GoogleScholarGoogle Scholar |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, Urcelay C, Veneklaas EJ, Reich PB, Poorter L, Wright IJ, Ray P, Enrico L, Pausas JG, de Vos AC, Buchmann N, Funes G, Quétier F, Hodgson JG, Thompson K, Morgan HD, ter Steege H, van der Heijden MGA, Sack L, Blonder B, Poschlod P, Vaieretti MV, Conti G, Staver AC, Aquino S, Cornelissen JHC (2013) New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany 61, 167–234.
| New handbook for standardised measurement of plant functional traits worldwide.Crossref | GoogleScholarGoogle Scholar |
Perry GLW, Wilmshurst JM, Ogden J, Enright NJ (2015) Exotic mammals and invasive plants alter fire-related thresholds in southern temperate forested landscapes. Ecosystems 18, 1290–1305.
| Exotic mammals and invasive plants alter fire-related thresholds in southern temperate forested landscapes.Crossref | GoogleScholarGoogle Scholar |
Plucinski MP, Anderson WR, Bradstock RA, Gill AM (2010) The initiation of fire spread in shrubland fuels recreated in the laboratory. International Journal of Wildland Fire 19, 512–520.
| The initiation of fire spread in shrubland fuels recreated in the laboratory.Crossref | GoogleScholarGoogle Scholar |
Popović Z, Bojović S, Marković M, Cerdà A (2021) Tree species flammability based on plant traits: a synthesis. Science of the Total Environment 800, 149625
| Tree species flammability based on plant traits: a synthesis.Crossref | GoogleScholarGoogle Scholar |
Potts E, Tng D, Apgaua D, Curran TJ, Engert J, Laurance SGW (2022) Growth form and functional traits influence the shoot flammability of tropical rainforest species. Forest Ecology and Management 522, 120485
| Growth form and functional traits influence the shoot flammability of tropical rainforest species.Crossref | GoogleScholarGoogle Scholar |
Prince D, Shen C, Fletcher T (2017) Semi-empirical model for fire spread in shrubs with spatially-defined fuel elements and flames. Fire Technology 53, 1439–1469.
| Semi-empirical model for fire spread in shrubs with spatially-defined fuel elements and flames.Crossref | GoogleScholarGoogle Scholar |
Radeloff VC, Hammer RB, Stewart SI, Fried JS, Holcomb SS, Mckeefry JF (2005) The wildland–urban interface in the United States. Ecological Applications 15, 799–805.
| The wildland–urban interface in the United States.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2022) ‘R: A language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/ [Accessed 12 August 2022]
Revelle W (2022) psych: Procedures for Personality and Psychological Research. Available at https://CRAN.R-project.org/package=psych, version=2.2.3 [Accessed 12 August 2022]
Romero B, Ganteaume A (2021) Effect of fire frequency on the flammability of two Mediterranean pines: link with needle terpene content. Plants 10, 2164
| Effect of fire frequency on the flammability of two Mediterranean pines: link with needle terpene content.Crossref | GoogleScholarGoogle Scholar |
Romero B, Fernandez C, Lecareux C, Ormeño E, Ganteaume A (2019) How terpene content affects fuel flammability of wildland–urban interface vegetation. International Journal of Wildland Fire 28, 614–627.
| How terpene content affects fuel flammability of wildland–urban interface vegetation.Crossref | GoogleScholarGoogle Scholar |
Santacruz-García AC, Bravo S, del Corro F, Ojeda F (2019) A comparative assessment of plant flammability through a functional approach: the case of woody species from Argentine Chaco Region. Austral Ecology 44, 1416–1429.
| A comparative assessment of plant flammability through a functional approach: the case of woody species from Argentine Chaco Region.Crossref | GoogleScholarGoogle Scholar |
Santamarta-Cerezal JC, Guzmán J, Neris J, Arraiza MP, Ioras F (2012) Forest hydrology, soil conservation and green barriers in Canary Islands. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 40, 9–13.
| Forest hydrology, soil conservation and green barriers in Canary Islands.Crossref | GoogleScholarGoogle Scholar |
Schwilk DW (2003) Flammability is a niche construction trait: canopy architecture affects fire intensity. The American Naturalist 162, 725–733.
| Flammability is a niche construction trait: canopy architecture affects fire intensity.Crossref | GoogleScholarGoogle Scholar |
Schwilk DW (2015) Dimensions of plant flammability. New Phytologist 206, 486–488.
| Dimensions of plant flammability.Crossref | GoogleScholarGoogle Scholar |
Signorell A, Aho K, Alfons A, Anderegg N, Aragon T, Arachchige C, Arppe A, Baddeley A, Barton K, Bolker B, Borchers HW, Caeiro F, Champely S, Chessel D, Chhay L, Cooper N, Cummins C, Dewey M, Doran HC, Dray S, Dupont C, Eddelbuettel D, Ekstrom C, Elff M, Enos J, Farebrother RW, Fox J, Francois R, Friendly M, Galili T, Gamer M, Gastwirth JL, Gegzna V, Gel YR, Graber S, Gross J, Grothendieck G, Harrell Jr FE, Heiberger R, Hoehle M, Hoffmann CW, Hojsgaard S, Hothorn T, Huerzeler M, Hui WW, Hurd P, Hyndman RJ, Jackson C, Kohl M, Korpela M, Kuhn M, Labes D, Leisch F, Lemon J, Li D, Maechler M, Magnusson A, Mainwaring B, Malter D, Marsaglia G, Marsaglia J, Matei A, Meyer D, Miao W, Millo G, Min Y, Mitchell D, Mueller F, Naepflin M, Navarro D, Nilsson H, Nordhausen K, Ogle D, Ooi H, Parsons N, Pavoine S, Plate T, Prendergast L, Rapold R, Revelle W, Rinker T, Ripley BD, Rodriguez C, Russell N, Sabbe N, Scherer R, Seshan VE, Smithson M, Snow G, Soetaert K, Stahel WA, Stephenson A, Stevenson M, Stubner R, Templ M, Lang DT, Therneau T, Tille Y, Torgo L, Trapletti A, Ulrich J, Ushey K, VanDerWal J, Venables B, Verzani J, Villacorta Iglesias PJ, Warnes GR, Wellek S, Wickham H, Wilcox RR, Wolf P, Wollschlaeger D, Wood J, Wu Y, Yee T, Zeileis A (2021) DescTools: tools for descriptive statistics. R package version 0.99.44.
Smith SA, Brown JW (2018) Constructing a broadly inclusive seed plant phylogeny. American Journal of Botany 105, 302–314.
| Constructing a broadly inclusive seed plant phylogeny.Crossref | GoogleScholarGoogle Scholar |
Snyder JR (1984) The role of fire: much ado about nothing. Oikos 43, 404–405.
| The role of fire: much ado about nothing.Crossref | GoogleScholarGoogle Scholar |
Souza MAd, Vale ATd (2019) Survey of low flammability plants in burned areas of cerrado in the federal district and analysis of their physical properties. Ciência Florestal 29, 191–192.
Tumino BJ, Duff TJ, Goodger JQD, Cawson JG (2019) Plant traits linked to field-scale- flammability metrics in prescribed burns in Eucalyptus forest. PLoS One 14, e0221403
| Plant traits linked to field-scale- flammability metrics in prescribed burns in Eucalyptus forest.Crossref | GoogleScholarGoogle Scholar |
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecology Letters 14, 702–708.
| Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems.Crossref | GoogleScholarGoogle Scholar |
Wang D, Guan D, Zhu S, Kinnon MM, Geng G, Zhang Q, Zheng H, Lei T, Shao S, Gong P, Davis SJ (2021) Economic footprint of California wildfires in 2018. Nature Sustainability 4, 252–260.
| Economic footprint of California wildfires in 2018.Crossref | GoogleScholarGoogle Scholar |
Webster J (2000) ‘The complete bushfire safety book.’ (Random House: NSW, Australia)
Weise DR, White RH, Beall FC, Etlinger M (2005) Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation. International Journal of Wildland Fire 14, 321–338.
| Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation.Crossref | GoogleScholarGoogle Scholar |
Whiffin T, Ladiges PY (1992) Patterns of variation and relationships in the Eucalyptus alpina–E. baxteri complex (Myrtaceae) based on leaf volatile oils. Australian Systematic Botany 5, 695–709.
| Patterns of variation and relationships in the Eucalyptus alpina–E. baxteri complex (Myrtaceae) based on leaf volatile oils.Crossref | GoogleScholarGoogle Scholar |
White RH, Zipperer WC (2010) Testing and Classification of individual plants for fire behaviour: plant selection for the wildland–urban interface. International Journal of Wildland Fire 19, 213–227.
| Testing and Classification of individual plants for fire behaviour: plant selection for the wildland–urban interface.Crossref | GoogleScholarGoogle Scholar |
Wyse SV, Perry GLW, O’Connell DM, Holland PS, Wright MJ, Hosted CL, Whitelock SL, Geary IJ, Maurin KJL, Curran TJ (2016) A quantitative assessment of shoot flammability for 60 tree and shrub species supports rankings based on expert opinion. International Journal of Wildland Fire 25, 466–477.
| A quantitative assessment of shoot flammability for 60 tree and shrub species supports rankings based on expert opinion.Crossref | GoogleScholarGoogle Scholar |
Wyse SV, Perry GLW, Curran TJ (2018) Shoot-level flammability of species mixtures is driven by the most flammable species: implications for vegetation-fire feedbacks favouring invasive species. Ecosystems 21, 886–900.
| Shoot-level flammability of species mixtures is driven by the most flammable species: implications for vegetation-fire feedbacks favouring invasive species.Crossref | GoogleScholarGoogle Scholar |
Zylstra P, Liow LH (2021) Linking fire behaviour and its ecological effects to plant traits, using FRaME in R. Methods in Ecology and Evolution 12, 1365–1378.
| Linking fire behaviour and its ecological effects to plant traits, using FRaME in R.Crossref | GoogleScholarGoogle Scholar |
Zylstra P, Bradstock RA, Bedward M, Penman TD, Doherty MD, Weber RO, Gill AM, Cary GJ (2016) Biophysical mechanistic modelling quantifies the effects of plant traits on fire severity: species, not surface fuel loads, determine flame dimensions in eucalypt forests. PLoS One 11, e0160715
| Biophysical mechanistic modelling quantifies the effects of plant traits on fire severity: species, not surface fuel loads, determine flame dimensions in eucalypt forests.Crossref | GoogleScholarGoogle Scholar |