

Restoration of burned and post-fire logged Austrocedrus chilensis stands in Patagonia: effects of competition and environmental conditions on seedling survival and growth
M. Florencia Urretavizcaya A B D and Guillermo E. Defossé A CA Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Centro de Investigación y Extensión Forestal Andino Patagónico (CIEFAP), CC 14 Esquel (9200) Chubut, Argentina.
B Ministerio de la Producción, Provincia de Chubut, Argentina.
C Universidad Nacional de la Patagonia San Juan Bosco, Facultad de Ingeniería, Esquel, Chubut, Argentina.
D Corresponding author. Email: mfurretavizcaya@ciefap.org.ar
International Journal of Wildland Fire 28(5) 365-376 https://doi.org/10.1071/WF18154
Submitted: 18 September 2018 Accepted: 21 January 2019 Published: 28 February 2019
Journal compilation © IAWF 2019 Open Access CC BY-NC-ND
Abstract
In Andean Patagonia, Argentina, severe wildfires produce high mortality in Austrocedrus chilensis forests. Owing to its high timber quality, A. chilensis trees are generally salvage logged right after wildfires. Post-logged areas result in open, denuded stands with low herbaceous or shrub cover, which precludes natural A. chilensis regeneration. In severely burned A. chilensis stands, we determined how different site conditions (salvage logging, SL, and without logging, WL), and the combination of different methods of control of above- and belowground competition and micro-environmental factors (incident radiation, soil temperature and soil moisture) affected survival, growth and hydric status of planted A seedlings. Two growing seasons after plantation, seedling survival was lower than 10% at SL sites, whereas it was near 90% at WL sites. Four seasons after establishment, and regardless of competition treatments, no seedlings survived at SL sites, whereas over 75% survived at WL sites. Radiation attenuation by canopy of WL sites benefitted A. chilensis seedling survival, and no additional control of early successional herbaceous species should be necessary to attain high seedling survival 4 years after restoration. Selective logging, by leaving ~50% of burned snags, and active restoration practices, may help reconcile economic needs of society and ecological requirements of A. chilensis for recovering former structure and functions.
Additional keywords: Andean cypress, facilitation, seedling performance, temperate forests.
Introduction
Wildfires have shaped the structure and functioning of most terrestrial ecosystems (Wright and Bailey 1982; Crisp et al. 2011). Every ecosystem, however, has its own fire history, specifically defined as its fire regime. The fire regime is composed of different elements, identified as (1) fire type and intensity, (2) size of typical significant fires and (3) frequency or return intervals for specific land units (Heinselman 1981). In forest ecosystems, long fire return intervals may produce high severity fires, whereas short fire return intervals may lead to low severity fires (Pyne 1984; Agee 1993; Beschta et al. 2004). High severity wildfires usually produce major changes in community and population structure, modifying resources availability and altering the physical environment (Agee 1993). These changes favour the establishment of early successional species, also creating suitable conditions for invasion by new species. High severity fires may also have long lasting effects on low fire-tolerant forests, especially in those whose most prevalent species reproduce strictly by seed, which are killed during, or because of, the fire disturbance. Moreover, if remaining burned snags are removed from stands after high severity fires, additional changes are created in the post-fire denuded environment, increasing solar radiation, and modifying soil temperature and wind speed (McIver and Starr 2000; Beschta et al. 2004). In other cases, in which seeds of forest species are slightly or not damaged by fire, the rapid growth of herbaceous species right after the fire disturbance may constrain their germination and establishment, by out-competing for water and nutrients (Rose and Rosner 2005). The combination of these factors generally delay the natural process of forest recovery and, in extreme situations, preclude burned forests from recovering to pre-disturbance conditions. Under these situations, the fire-disturbed stand needs active restoration practices to return to its normal successional path, by sowing or planting former species. However, in order to guarantee the establishment of the target species, restoration practices sometimes require additional microsite manipulation to avoid or diminish any deleterious biotic or micro-environmental effects on planted seedlings (Whisenant 1999). These effects are mainly caused by competition for soil water or nutrients by early successional herbaceous vegetation, excessive soil heating by higher incoming radiation reaching denuded soils, or by the lack of nurse plants (generally shrubs or half-shrubs) that protect seedlings from excessive temperature fluctuations (Padilla and Pugnaire 2006; Urretavizcaya et al. 2017). Forest-productivity research has identified interactions between competing vegetation and logging debris left after forest harvesting. In general, retention of logging debris in temperate zones inhibits development of herbaceous and sometimes woody species. Retention of logging debris has also been associated with mulching effects. These effects imply conservation of soil water near the soil surface by reducing evaporation and topsoil temperatures (Harrington et al. 2013).
In restoration projects, controlling competing species may be sometimes necessary for diminishing their deleterious effects on planted seedlings (Jobidon et al. 1998; Löf et al. 1998; Dinger and Rose 2009; Maguire et al. 2009). This control may involve using herbicides, manual removal of competing vegetation or the application of mulch (Löf et al. 1998; Harper et al. 2005; Jylhä and Hytönen 2006). Removing competing vegetation by chemical or mechanical methods reduces soil cover and modifies some topsoil physical conditions, increasing incoming radiation and soil-surface temperature, and also influencing soil-water content (Defossé and Robberecht 1996; Harrington et al. 2013). For some forest species at seedling stages, high topsoil temperatures at stem-collar level may negatively affect their survival (Kolb and Robberrecht 1996). However, the use of mulch around seedlings may increase survival by reducing temperature variations, conserving soil moisture and diminishing plant evapotranspiration (Flint and Childs 1987). Although mulching is widely used in horticulture, it has scarcely been used in restoration projects aimed at recovering forest species under field conditions (Blanco-García and Lindig-Cisneros 2005).
The forests of Austrocedrus chilensis (D.Don) Pic. Ser et Bizarri (commonly known as cypress), are part of the Andean–Patagonian temperate forests that run along the Cordillera de los Andes in southern Chile and Argentina (Cabrera 1976). In Argentina, A. chilensis is located in a narrow belt that goes from 37°08′ to 43°44′S in the eastern slope of the Andes (Pastorino et al. 2006). Austrocedrus chilensis forms either pure of mixed stands. Pure stands occupy ~95 500 ha, whereas those in which A. chilensis grows intermixed with Nothofagus dombeyi Mirb. Oerst. (locally named as coihue) cover ~32 800 ha. National fire statistics show that during the last 20 years, an average of 3000 ha of pure or mixed A. chilensis stands have been annually affected by wildfires (Secretaría de Ambiente y Desarrollo Sustentable de la Nación 2013; Mohr Bell 2015).
After wildfires, the main factors that influence recovery of affected forests are intensity and severity of each particular fire, size or area covered, and environmental conditions prevailing during and after the fire event (Connell and Slatyer 1977; Veblen et al. 1999). For A. chilensis forests, the lack of a soil-seed bank after severe wildfires is the initial limiting factor for natural stand recovery (Urretavizcaya and Defossé 2004). Likewise, severe wildfires produce very high A. chilensis tree mortality because its very thin bark is rapidly affected by the fire and, unlike other Patagonian species, it does not re-sprout after fires (Gallo et al. 2004). These conditions mean that active-restoration practices, such as seedling planting or seed sowing, are often necessary for restoring A. chilensis stands affected by severe fires (Urretavizcaya et al. 2012).
Most burned A. chilensis forests are logged right after fire events because of their high timber quality and market demands (Perdomo et al. 2007). Once burned-log extraction is finished, logged areas result in open, denuded stands with very low herbaceous or shrub cover, similar to what happens in other postfire-logged conifer forests elsewhere (McIver and Starr 2000; Beschta et al. 2004). However, for A. chilensis forests, there are no studies that analyse these conditions. The new macro- and micro-environmental conditions created in these post-burning logged areas may preclude survival and establishment of planted A. chilensis seedlings, sometimes making any restoration plan unsuccessful. Similar to what was stated in Castro et al. (2011) and Harrington et al. (2013, 2018), we hypothesise that, for A. chilensis forests, the presence of burned logs act as structural nurses that, without competing with established vegetation, help reduce incoming radiation and maintain topsoil moisture, crucial for seedling survival during the summer drought. In the case of salvage logging, mulch attenuates the desiccant effects of high levels of direct solar radiation and help reduce evaporative loss by diminishing high topsoil temperatures that are usual in this region during the summer. We also postulate that mulch, as well as a reduction in nearby vegetation competition, has the potential to benefit A. chilensis seedling performance by increasing resource availability and modifying microclimate conditions.
The objective of this study was to determine how different site conditions (salvage logging, SL, and without logging, WL), microsite interventions (different methods of control of above- and belowground competition), and micro-environmental factors (incident radiation, soil temperature and moisture) affect survival, growth and hydric status of A. chilensis seedlings planted in severely burned stands. The outcomes of this study not only cast light on the effects of these factors on A. chilensis seedling establishment, but also help to design appropriate protocols for successfully restoring fire-damaged A. chilensis forests. Additionally, this study provides some guidelines that should be followed for sustainable managing post-fire A. chilensis stands.
Materials and methods
Study site
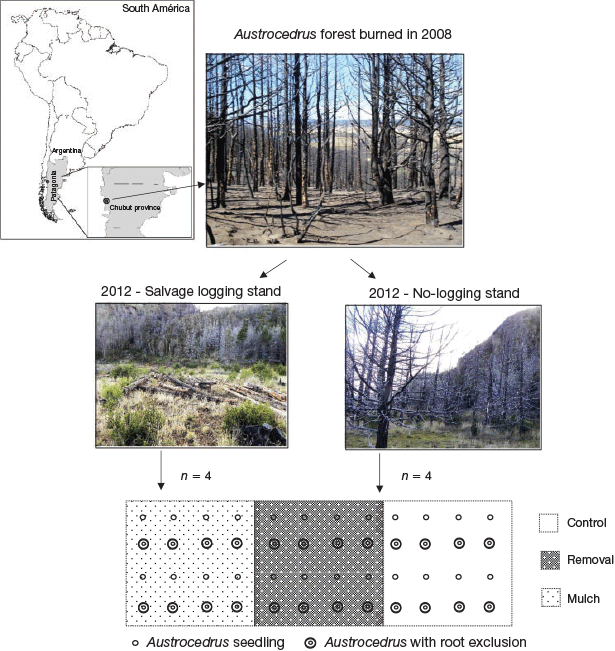
The study area was located near the town of Corcovado (43°32′S, 71°33′W), in Chubut province, Patagonia, Argentina (Fig. 1). Its climate is “mediterranean-temperate”, with cold and rainy winters and dry and mild-to-hot summers and early autumn. Mean annual precipitation is 800 mm, 75% of which falls during the winter months. Average annual temperature is 9.3°C. Soils of the region derive from volcanic ashes (Etchevehere 1972) and correspond to Andisols in the study area. These are rich in nutrients (except phosphorous), and have high soil-water retention (Colmet Dâage et al. 1993). This area represents the southernmost location of pure stands of A. chilensis in the Patagonian region of Argentina (Dezzotti and Sancholuz 1991). At the end of February 2008 (summer in the southern hemisphere), a wildfire categorised as extreme behaviour and high severity ravaged 80 ha within this area. The categorisation of this fire was based on regional observations and by applying the classification proposed by Mutch and Swetnam (1995). Under this classification, the studied fire falls in the category 5: very high severity; most sub-canopy trees killed and >50% of canopy trees killed (Fig. 1).
|
Experimental design and treatments
In an area representative of the burned forest, we selected two adjacent and similarly burned A. chilensis stands (Fig. 1). One of them (~4 ha) was salvage cut in 2010 (i.e. all remaining A. chilensis snags removed), whereas the other (2 ha) conserved all the standing burned trees. Both stands presented similar aspect and 0 to 5% slope. In August 2012 (winter in the southern hemisphere), a split–split plot experiment was installed. Site condition or logging was the main-effect plot and had two levels (SL and undisturbed or WL). Aboveground competition was the second-effect plot and had three levels (aboveground vegetation manually removed (RE), mulch application (MU) and no aboveground removal, or control (CO)). Root competition was the third-effect plot and had two levels (root exclusion (EX), and root control or without exclusion (RC)).
The experiment was installed in eight plantation blocks of 8 × 24 m (4 in SL and 4 in WL conditions) (Fig. 1). In every block, 48 A. chilensis seedlings were planted at a distance of 2 × 2 m each. At the time of planting, seedlings were 3 years old and were produced at Centro de Investigación y Extensión Forestal Andino Patagónico nursery (Esquel, Chubut, Argentina) from seeds of the same genetic zone of the study site (Pastorino 2015). Each seedling was manually planted in similarly shovel-made holes. Before planting, a sample composed of 20 randomly chosen seedlings were tested to determine their morphological attributes. These attributes (mean ± standard error) were: seedling height (21 ± 0.5 cm); collar diameter (4.2 ± 0.1 mm), aerial dry weight (3.1 ± 0.2 g), root dry weight (2.5 ± 0.2 g), aerial to root dry weight ratio (1.3) and sturdiness index (50) (Hasse 2007). These values are within the range of customary morphological attributes used for commercial A. chilensis seedlings. At planting, plastic root-exclusion tubes 25 cm in diameter and 25 cm long (Cook and Ratcliff 1984; Defossé 1995) were installed in half of the planted seedlings to isolate their root system from those of neighbouring herbaceous species. Each block was divided into three plots (sample units) containing 16 seedlings each. The three aerial competition treatments (RE, MU and CO) were randomly assigned to one of these plots (Fig. 1). In total, 456 A. chilensis seedlings were used in the experiment (228 per stand and 48 per block). Right after plantation, all seedlings were identified and their specific height and collar diameter measured for determining further survival and growth. Each seedling was protected by a metallic mesh for avoiding foraging by the exotic European hare (Lepus europaeus Pallas).
The mulch came from chipped A. chilensis branches and slash residues remaining after the wildfire. Mulch cover was 3–5-cm high in a 40-cm radius around treated seedlings. Aerial vegetation was manually removed with a hoe and a rake from each removal treatment in a 40-cm radius around each seedling, trying not to disturb the topsoil around it (Fig. 2).

|
Characterisation of the vegetation of the study site
At 3 and 6 months after setting plantation (November 2012, spring in the southern hemisphere, and February 2013, summer in the southern hemisphere), early successional species grown in both SL and WL stands were cut, taken to the laboratory, identified and their dry weight determined. The sampling procedure involved collection of all biomass located within eight 0.5-m2 plots randomly distributed within Control plots of SL (n = 4) and WL blocks (n = 4).
Environmental variables measured
Photosynthetically active radiation (PAR), and soil temperature (ST) and moisture (SM) were measured three times (mid-spring, 11 November 2012; mid-summer, 6 February 2013; and late summer, 22 March 2013) during the first growing season after the start of the experiment. These measurements were performed to determine if there were significant differences among sites (SL and WL) and aerial competition (RE, MU and CO) treatments. For measuring PAR, a Cavadevices Ceptometer (Cavadevices.com, Buenos Aires, Argentina) with Photosynthetic Photon Flux method was used. Topsoil temperature was measured with an infrared laser thermometer Cole–Parmer Model 39650-02 (Instrument Company, Chicago, IL, USA) and at 10-cm soil depth with a Digi-Sense DualLogR thermometer and thermocouples Model 91100-50 (Cole-Parmer Instrument Company Vernon Hills, IL, USA). Soil samples from 0 to 10 cm (2 per plot in each block, 48 per date) and 20 to 30 cm (1 per plot in each block, 24 per date) were taken and their water content determined gravimetrically. At the beginning of the second growing season (October 2013), automatic data loggers Decagon Em5b (Decagon Devices, Pullman, USA) with moisture sensors ECH2O (Decagon Devices, Pullman, USA) at 10-cm soil depth were installed in randomly selected SL and WL blocks and each aerial competition treatment. These measurements allowed continuous recording of soil moisture data to the end of the growing season (March 2014).
Seedling survival, growth and hydric conditions measurements
We recorded seedling survival and height growth at the end of the growing season in 2013, 2014 and 2016. In April 2014 and 2016, seedling-collar diameter was also measured. Survival, height and collar-diameter measurements were performed in all seedlings. In mid-summer (February 2013), the hydric status of seedlings was determined both at predawn and midday at both SL and WL sites, and also in aerial (RE, MU and CO) and belowground (EX and RC) competition treatments. For these measurements, we used a Scholander pressure chamber (BioControl Model 4, Buenos Aires, Argentina). The measuring technique involved cutting of a small portion of a top A. chilensis seedling branch, using 24 seedlings at predawn and 24 at midday. Two blocks were randomly selected for each WL and SL site treatment and 4 seedlings were sampled per each aerial (RE, MU and CO) treatment, 2 in EX and 2 in the RC treatments.
Analyses
Vegetation biomass was analysed by analysis of variance (ANOVA) of one factor: site (WL and SL), and independently for each measuring date. PAR, ST and SM determined during the first growing season, were analysed by ANOVA with two factors: site (WL and SL) and aerial competition (RE, MU and CO) treatments, and independently for each measuring date. Soil moisture, which started to be continuously measured from the beginning of the second growing season, was analysed through mixed lineal models. Soil-moisture values were averaged at monthly intervals for each treatment, and significant differences among treatments determined for each month during the growing season.
Austrocedrus chilensis seedling survival was evaluated as a function of the first (SL and WL), second (RE, MU and CO) and third (EX and RC) effect treatments. The analyses involved repeated measures through lineal mixed models by considering the three effects as fixed, and blocks and time as random factors. These analyses were performed after the first summer, second winter and second summer (April and October 2013 and April 2014 respectively). Survival at the fourth summer (April 2016) was only measured in aerial (RE, MU, CO) and root (EX and RC) competition in WL treatment, because no survival was registered at SL plots and blocks. In these analyses, P-values with Bonferroni correction and a P ≤ 0.05 were used. When significant differences were found, mean comparison was made by using Fisher’s least significant difference. Height growth after the first growing season (2012–2013) was analysed with the same models used for survival. After that date, the same models were used but only for one site (WL), because most seedlings at SL plots had already died. Height and collar-diameter growth was analysed for the periods 2012–2014 and 2014–2016 and then for the entire study period (2012–2016). Seedling hydric status was analysed by ANOVA with three factors: site (SL and WL), aerial (RE, MU and CO) and root (EX and RC) competition treatments. For all analyses, we used INFOSTAT Statistical Package (Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Córdoba, Argentina, www.infostat.com.ar, accessed 7 February 2019) (Balzarini et al. 2008; Di Rienzo et al. 2017) and their interface with the R Foundation for Statistical Computing Platform(R version 3.4.0 The R-CoreTeam 2017, https://www.r-project.org/, accessed 7 February 2019). Unless stated otherwise, the probability level was set at P < 0.05.
Results
Vegetation
At the beginning of the experiment, herbaceous plant cover was similar at all treatment plots and blocks, and the predominant species were Holcus lanatus, Acaena ovalifolia, Carduus thoermeri, Rumex acetosella and other grasses. Total herbaceous (grasses and herbs) aboveground biomass was not significantly different at both SL and WL plots. Three and six months after plantation (4.5 and 5 years after the fire), the biomass averaged 272 ± 38 and 414 ± 25 g m−2 at SL plots and 395 ± 44 and 431 ± 34 g m−2 at WL plots.
Environmental variables
Photosynthetically active radiation
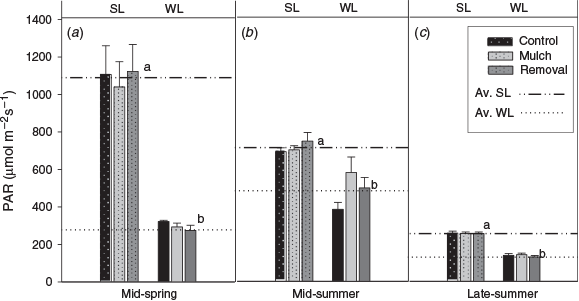
Photosynthetically active radiation was significantly higher in SL treatments as compared with WL treatments, but was not significantly different among aerial (RE, MU and CO) treatments. Within sites (SL and WL), the higher PAR differences were registered during mid-spring 2012. Salvage logging showed PAR values above 1000 µmol m−2 s−1, almost four times the PAR registered (290 µmol m−2 s−1 ) in WL treatments. Photosynthetically active radiation differences started to decline towards mid- and late summer (Fig. 3).
|
Soil temperature
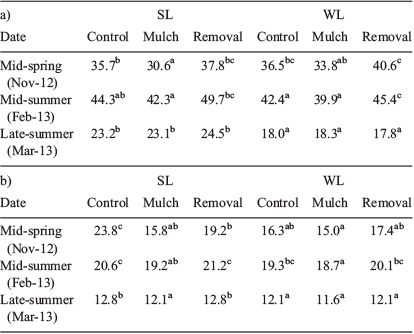
Topsoil temperature in mid-spring and mid-summer of the first growing season was not significantly different between sites (SL and WL), but it was significantly different among competition treatments (Table 1a). In both SL and WL plots, topsoil temperature varied between 30 and 40°C in mid-spring, increasing to 40 to 50°C by mid-summer. The MU treatment significantly reduced topsoil temperature from mid-spring to mid-summer as compared with the other aerial competition treatments. During the summer, SL plots showed slightly higher topsoil temperatures as compared with WL sites, although they were significantly different only at the end of the summer (18.0 and 23.4°C for WL and SL respectively, Table 1a).
|
Soil temperature at 10 cm belowground was not significantly different between SL and WL plots, but it was significantly different among aerial competition treatments (Table 1b). The MU treatment also showed a significantly lower 10-cm soil depth temperature as compared with the other aerial competition treatments. Temperature at 10 cm soil depth was, in average, 18, 24 and 8°C lower than topsoil temperature in mid-spring, and mid- and late summer respectively.
Soil moisture
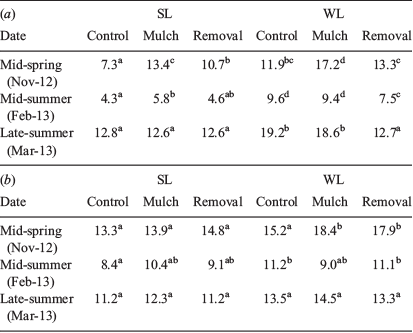
During the first growing season after seedling plantation, soil moisture in the first 10-cm soil depth was not significantly different between SL and WL plots in mid-spring, but it was significantly higher at WL plots as compared with SL sites as the growing season advanced (Table 2a). Soil moisture was also significantly different among aerial competition treatments (CO < RE < MU) in mid-spring and mid-summer. In mid-summer, soil moisture was significantly lower in RE as compared with MU treatments (Table 2a). In late summer (end of the first growing season), soil moisture was significantly lower in all competition treatments at SL sites as compared with those located at WL sites. At 20- to 30-cm depth, soil moisture was not significantly different at SL and WL sites in the mid-spring measurement, although it was significantly lower in CO as compared with the other aerial competition treatments (Table 2b). During mid-summer, interactions between sites (SL and WL) and aerial competition treatments were marginally significant (P = 0.0522). As it was expected, soil moisture in the CO treatments at SL sites was significantly lower than that at WL sites, whereas CO in this treatment showed the higher moisture values (Table 2b). At the end of the summer, no significant differences in moisture values were recorded between sites or among aerial competition treatments.
|
Dynamics of soil moisture at 10-cm soil depth
Soil moisture in the CO and RE aerial competition treatments at WL site was highest during mid-spring, with 25.6 and 23.5% respectively. Aerial competition treatments at the same date but in SL showed intermediate soil moisture values, ~20%, while MU treatment showed the lowest soil moisture values 4.7 and 14.4% in SL and WL, respectively. Soil-moisture dynamics showed a constant decline from the beginning of the spring towards the summer, with some upward pulses that represent mid-summer precipitation events (Fig. 4). Application of the MU treatments appears to have a deleterious effect in both SL and WL site treatments, although the WL treatment seems to rapidly recover moisture values after two rainfall events. After three consecutive rainfall events in mid-summer (February), all treatment effects (i.e. sites and aerial competition) disappeared. Soil moisture steadily diminished in all treatments towards the end of the summer (Fig. 4).

|
Austrocedrus chilensis performance
Seedling survival
Seedling survival was significantly affected by sites (SL and WL) during the whole study period, and by aerial competition treatments (RE, MU and CO) during the second winter and the second summer analysed (Fig. 5). However, during the first 2 years after starting the experiment, survival was not significantly different in both root (EX and RC) competition treatments.

|
Survival slowly diminished at SL sites as compared with WL sites after the first summer, and did not present significant differences related to aerial (RE, MU and CO) or root (EX and RC) competition treatments (Fig. 5). After the second winter, however; seedling survival steadily diminished at SL sites for all aerial competition treatments. At the beginning of the second summer, most seedlings at SL treatment had died, and only 20% of seedlings in the aerial competition control (CO) remained alive. No seedlings survived at SL sites by the fourth summer after plantation, whereas for all competition treatments of WL, survival remained above 85%, with the only exception of the combination of EX and MU treatments, which was 75% (Table 3). At the end of the fourth growing season, significant interaction was found between aerial (RE, MU, and CO) and root competition (EX and RC) treatments. The highest survival was registered in the combination between CO and EX treatments, and in MU and RC treatments. Unexpectedly, the MU and EX treatments showed the lowest survival. The other treatments presented intermediate survival values (Table 3).

|
Seedling growth
During the first summer after plantation, seedling height growth was significantly different in all treatments: site (SL and WL), aerial (RE, MU and CO) and root competition (EX and RC), although it was not significant with regard to any treatment interactions. Seedlings grew more at WL sites (P < 0.0001) and in RE and MU (P = 0.0099) and EX (P = 0.0022) treatments. Growth was minimal, with absolute values that varied from 0.25 to 1.11 cm at SL sites, although they varied from 1.25 to 2.56 cm at WL sites. The combination of WL with RE and EX treatments showed significantly higher growth as compared with all SL treatments and to WL-CO-RC treatment combination (Table 4).
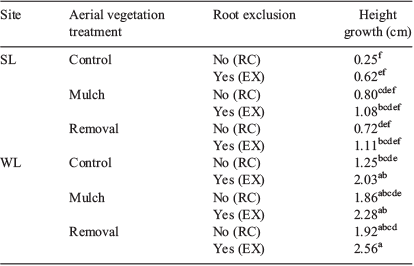
|
From the first summer up to the end of the experiment, seedling growth was only measured at WL sites because all seedlings at SL sites had already died. Within WL treatment, seedling height growth was similar in all aerial and root competition treatments, and averaged 5.6 ± 1.4 cm during the period 2012–2014 (Fig. 6a). From 2014 to the end of the experiment in 2016, seedlings grew at an average of 19.3 ± 1.5 cm in the CO treatment, significantly higher than the RE and MU treatments, which grew 14.1 ± 0.5 and 16.3 ± 1.5 cm respectively (Fig. 6a). No significant differences in height growth were found between EX and RC root competition treatments during this period. By considering the whole study period (2012–2016), all seedlings in this WL site treatment grew 21.0 ± 1.2 cm on average, presenting no significant differences among all aerial treatments (RE, MU and CO) or between root competition (EX and RC) treatments (Fig. 6a). Seedling collar diameter was not significantly different for all aerial and root competition treatments, and grew on average 1.9 ± 0.3 mm for the 2014–2016 period and 3.4 ± 0.4 mm for the 2014–2016 period. For the complete 2012–2016 period, collar diameter grew on average 5.1 ± 0.5 mm (Fig. 6a).

|
Hydric status of A. chilensis seedlings
In mid-summer of the first growing season, predawn and midday hydric status of cypress seedling branches was associated with site (SL and WL) but not to aerial or root competition treatments. At predawn, seedlings grown at SL sites showed significantly (P < 0001) lower (more negative) water potential average values (−1.6 MPa) than at WL sites (−1.2 MPa) (Fig. 7a). At midday, when the were higher evapotranspiration rates, average values were −2.2 MPa for SL plots and −1.6 MPa for WL plots (Fig. 7b).

|
Discussion
This study showed that, in A. chilensis post-burned stands, salvage logging significantly affected survival and growth of restored A. chilensis seedlings. Results showed that two growing seasons after plantation, seedling survival was <10% at SL sites, whereas it was near 90% at WL sites. Four seasons after establishment, and regardless of sub-treatments, no seedlings survived at SL sites, whereas over 75% survived at WL plots. These findings are useful to help alleviate the lack of reforestation typically observed at burned and logged sites.
Tree seedlings, in general, experience transplantation shock during the first growing season after planting, while they try to acclimate to their new site conditions (Hobbs 1984; Ouzts et al. 2015). During their acclimation period, A. chilensis seedlings showed a little shoot growth, while post-fire herbaceous vegetation established vigorously. Competition by neighbouring species is one of the main interactions affecting vegetation growth in early post-fire successional stages (Jobidon et al. 1998; Löf et al. 1998; Dinger and Rose 2009; Maguire et al. 2009). However, competition might affect growth in a great variety of ways, and it may differ among communities (Reader et al. 1994). In a mesic area similar to our study site, A. chilensis seedling survival and growth were positively associated with post-fire cover during the 2 years after plantation (Urretavizcaya et al. 2017). In our study, however, this nursing effect appeared two seasons after acclimation. Neighbouring plants seemed to exert a nursing rather than a competing effect, as A. chilensis seedlings grew vigorously from the third season onwards. These results agree with the study of Holmgren and Scheffer (2010), who reported results for a similar mesic site, and with other authors (Veblen et al. 1995; Gobbi 1999) dealing with other pure and fire-undisturbed A. chilensis stands. These studies showed that facilitation overcomes other interaction effects, and occurs in small to intermediate canopy gaps (Veblen et al. 1995) associated with high herbaceous and shrub cover (Gobbi 1999). Our study also showed that the use of root-exclusion tubes neither benefited nor harmed seedling performance.
At the beginning of the first summer after seedling plantation, PAR at SL sites was almost 4 times higher than at WL plots, significantly increasing topsoil temperature at the end of that period. These factors created a more stressful microenvironment at SL sites, perhaps causing its high seedling mortality as compared with WL plots by the end of the second summer. By contrast, at WL sites, standing snags reduced PAR reaching the soil surface, attenuating topsoil heating and improving overall seedling micro-environmental conditions. This situation is similar to what Castro et al. (2011) reported for plants grown in other Mediterranean environments. Although PAR and topsoil temperature at the end of the summer were significantly higher at SL sites as compared with WL plots, soil moisture was similar in both sites. Why, then, when having similar soil moisture levels, was higher mortality produced at SL sites as compared with WL plots? The answer could be found by considering the way A. chilensis developed physiological mechanisms to avoid hydric stress. Gyenge et al. (2005) demonstrated that A. chilensis avoids water stress by strong stomatal control. In our study, A. chilensis seedlings were able to tolerate soil-water deficit in both SL and WL treatments, but were unable to cope with high radiation levels at SL sites as compared with WL plots. This inference is supported by other studies with the same species, which showed that, even with adequate soil moisture, high levels of incident radiation increase the risk of photo inhibition and high cell damage because of overheating, causing high A. chilensis seedling mortality (Gyenge et al. 2007; Caselli et al. 2019). Additionally at WL sites, shade protection exerted by standing snags not only reduced PAR and diurnal topsoil temperatures, but also attenuated low night temperatures especially during the winter, thus creating a better microclimate for planted seedlings. This inference is supported by Castro et al. (2011), whose study showed that stands with snags maintain higher topsoil temperatures in winter as compared with nearby bare-tree areas.
After the first dry summer season, soil water returned to field capacity in all treatments except in those covered by mulch. This MU treatment may have obstructed normal water infiltration into the soil profile, being more noticeable at SL sites than in WL plots, because of its higher levels of radiation and the desiccant effect of winds. This may help explain the poor performance of seedlings grown in this MU treatment as compared with other treatments. Mulch is a mechanical barrier that may protect the seedling collar from high PAR and high topsoil temperatures, but also intercepting precipitation and dew condensation. This may be the reason why, in other study dealing with A. chilensis seedlings (Lallement et al. 2010), no differences in survival and growth were detected by using 2-cm shredded wood as mulch, as compared with unprotected control.
Conclusions
Severe fires kill A. chilensis trees and seeds, producing drastic modifications in stand structure. Post-fire macro- and micro-environmental conditions, particularly high PAR levels, may limit seedling survival and growth if salvage logging is applied to remaining A. chilensis snags, making any restoration efforts futile. In burned sites without logging, however; PAR attenuation by a canopy of remaining snags benefitted A. chilensis seedling survival, and no additional control of early successional herbaceous species should be necessary to attain high seedling survival. Also at SL sites, diminution of aerial or belowground competition did not ameliorate the deleterious effects of PAR. The addition of mulch as it was applied and, contrary to what was expected, did not improve seedling survival or growth. Our results agree with other studies (i.e. McIver and Starr 2000; Castro et al. 2011), which showed that post-fire standing snags provide significant shade, diminishing soil-surface heating and modifying surface micro-environmental conditions as compared with nearby salvage logging areas. However, standing post-fire snags are often logged for economic reasons (Castro et al. 2011), posing socio-ecological conflicts should the stand need to be actively restored, because seedling survival under this situation is almost nil (Keyser et al. 2009).
Taking into account these results, and to achieve high seedling establishment rates, limited logging is suggested in burned forest, such as the one presented in this case. One generally accepted recommendation is to leave ~50% of the snags of each diameter class, and all snags whose diameter at breast height is >50 cm or were >150 years old (Beschta et al. 2004). This recommendation may also be compatible with objectives related to the many other ecological roles of snags in the recovering landscape, including the provision of habitat for a variety of species and playing a key role in biological and physical processes (Thomas 1979; Donato et al. 2013). In burned A. chilensis stands, if salvage logging is the only alternative, a restoration plan should consider waiting for seedling plantation till the lower strata of plant cover and some shrubs has recovered. These shrubs may act as nurse plants, facilitating seedling survival and assuring further establishment (Kitzberger et al. 2000; Urretavizcaya et al. 2012). Aerial devices could also be used to attenuate direct radiation during early stages of seedling establishment (Urretavizcaya and Defossé 2013). Finally, post-fire management of burned forests should consider a variety of options to help reconcile and better balance the competing society’s economical needs with the benefits of sustainable management through ecological restoration.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This project was partly financed by a Grant (PIP173-CONICET) from the National Research Council of Argentina.
Acknowledgements
We gratefully acknowledge Luis Villegas and other personnel of Santa Teresita ranch, for the interest and collaboration during the development of this study. We also owe special thanks to Oscar De Knollseisen, M. Florencia Oyharçabal and Juan Monges for their collaboration in many stages of the study while carrying out this Project, and to Lucas Díaz and Gabriela Tavella for their participation in the installation of the experiment.
References
Agee JK (Ed.) (1993) ‘Fire Ecology of Pacific Northwest Forests.’ (Island Press: Washington, DC, USA)Balzarini MG, Gonzalez L, Tablada M, Casanoves F, Di Rienzo JA, Robledo CW (2008). Manual del Usuario, Editorial Brujas, Córdoba, Argentina.
Beschta RL, Rhodes JJ, Kauffman JB, Gresswell RE, Minshall GW, Karr JR, Parry DA, Hauer ER, Frissell CA (2004) Postfire management on forested public lands of the western United States. Conservation Biology 18, 957–967.
| Postfire management on forested public lands of the western United States.Crossref | GoogleScholarGoogle Scholar |
Blanco-García A, Lindig-Cisneros R (2005) Incorporation in sustainable forestry management: using Pine-Bark to improve native species establishment on tephra deposits. Ecology 13, 703–709.
Cabrera AL (Ed.) (1976) ‘Las Regiones Fitogeográficas Argentinas.’ (Editorial ACME: Buenos Aires, Argentina)
Caselli M, Urretavizcaya MF, Loguercio GÁ, Defossé GE (2019) Light and moisture conditions suitable for establishing andean cypress and coihue beech seedlings in Patagonia: a nursery approach. Forest Science 65, 27–39.
Castro J, Allen CD, Molina-Morales M, Marañón-Jiménez S, Sánchez-Miranda Á, Zamora R (2011) Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment. Restoration Ecology 19, 537–544.
| Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment.Crossref | GoogleScholarGoogle Scholar |
Colmet Dâage F, Mazzarino MJ, Lanciotti AA (1993) Características de los suelos volcánicos en el S.O. del Chubut. Instituto Nacional de Tecnología Agropecuaria (INTA), Centro Regional Patagonia Norte (Bariloche) INTA Comunicación Técnica N° 22, Recursos naturales-suelos.
Connell JH, Slatyer RO (1977) Mechanisms of sucesión in natural communities and their role in community stability and organization. American Naturalist 111, 1119–1144.
| Mechanisms of sucesión in natural communities and their role in community stability and organization.Crossref | GoogleScholarGoogle Scholar |
Cook SG, Ratcliff D (1984) A study of root and shoot competition on the growth of green panic (Panicum maximum var trichoglume) seedlings in an existing grassland using root exclusion tubes. Journal of Applied Ecology 21, 971–982.
| A study of root and shoot competition on the growth of green panic (Panicum maximum var trichoglume) seedlings in an existing grassland using root exclusion tubes.Crossref | GoogleScholarGoogle Scholar |
Crisp MD, Burrows GE, Cook LG, Thornhill AH, Bowman DM (2011) Flammable biomes dominated by eucalypts originated at the Cretaceous–Palaeogene boundary. Nature Communications 2, 193
| Flammable biomes dominated by eucalypts originated at the Cretaceous–Palaeogene boundary.Crossref | GoogleScholarGoogle Scholar | 21326225PubMed |
Defossé GE (1995) Germination, emergence, and survival of Festuca spp. seedlings in a steppe of Patagonia, Argentina. PhD thesis, University of Idaho, Moscow, ID, USA.
Defossé GE, Robberecht R (1996) Effects of competition on the postfire recovery of 2 bunchgrass species. Journal of Range Management 49, 137–142.
| Effects of competition on the postfire recovery of 2 bunchgrass species.Crossref | GoogleScholarGoogle Scholar |
Dezzotti A, Sancholuz L (1991) Los bosques de Austrocedrus chilensis en Argentina: ubicación, estructura y crecimiento. Bosque 12, 43–52.
| Los bosques de Austrocedrus chilensis en Argentina: ubicación, estructura y crecimiento.Crossref | GoogleScholarGoogle Scholar |
Dinger EJ, Rose R (2009) Integration of soil moisture, xylem water potential, and fall–spring herbicide treatments to achieve the maximum growth response in newly planted Douglas-fir seedlings. Canadian Journal of Forest Research 39, 1401–1414.
| Integration of soil moisture, xylem water potential, and fall–spring herbicide treatments to achieve the maximum growth response in newly planted Douglas-fir seedlings.Crossref | GoogleScholarGoogle Scholar |
Donato DC, Fontaine JB, Kauffman JB, Robinson WD, Law BE (2013) Fuel mass and forest structure following stand-replacement fire and post-fire logging in a mixed-evergreen forest. International Journal of Wildland Fire 22, 652–666.
| Fuel mass and forest structure following stand-replacement fire and post-fire logging in a mixed-evergreen forest.Crossref | GoogleScholarGoogle Scholar |
Etchevehere PH (1972) Los suelos de la Región Andino Patagónica. In ‘La Región de los Bosques Andino-Patagónicos, Sinopsis General’. (Ed. M. J. Dimitri) pp. 83–95. (Colección Científica del INTA: Buenos Aires, Argentina)
Flint L, Childs S (1987) Effect of shading, mulching and vegetation control on Douglas-fir seedling growth and soil water supply. Forest Ecology and Management 18, 189–203.
| Effect of shading, mulching and vegetation control on Douglas-fir seedling growth and soil water supply.Crossref | GoogleScholarGoogle Scholar |
Gallo L, Pastorino MJ, Donoso C (2004) Variación en Austrocedrus chilensis (D.don) Pic. Ser et Bizzarri (Ciprés de la Cordillera). In ‘Variación Intraespecífica en las especies arbóreas de los bosques templados de Chile y Argentina’. (Eds C Donoso, A Premoli, L Gallo, R Ipinza) pp. 233–250. (Marisa Cuneo: Valdivia, Chile)
Gobbi M (1999) Austrocedrus chilensis management: effects on microsites and regeneration. International Journal of Ecology and Environmental Sciences 25, 71–83.
Gyenge JE, Fernández ME, Dalla Salda G, Schlichter T (2005) Leaf and whole-plant water relations of thee Patagonian conifer Austrocedrus chilensis (D.Don) Pic. Ser. et Bizzarri: implications on its drought resistance capacity. Annals of Forest Science 62, 297–302.
| Leaf and whole-plant water relations of thee Patagonian conifer Austrocedrus chilensis (D.Don) Pic. Ser. et Bizzarri: implications on its drought resistance capacity.Crossref | GoogleScholarGoogle Scholar |
Gyenge JE, Fernández ME, Schlichter T (2007) Influence of radiation and drought on gas exchange of Austrocedrus chilensis seedlings. Bosque 28, 220–225.
| Influence of radiation and drought on gas exchange of Austrocedrus chilensis seedlings.Crossref | GoogleScholarGoogle Scholar |
Harper GJ, Comeau PG, Biring BS (2005) A comparison of herbicide and mulch mat treatments for reducing grass, herb, and shrub competition in the BC interior Douglas-fir zone – ten-year results. Western Journal of Applied Forestry 20, 167–176.
Harrington TB, Slesak RA, Schoenholtz SH (2013) Variation in logging debris cover influences competitor abundance, resource availability, and early growth of planted Douglas-fir. Forest Ecology and Management 296, 41–52.
| Variation in logging debris cover influences competitor abundance, resource availability, and early growth of planted Douglas-fir.Crossref | GoogleScholarGoogle Scholar |
Harrington TB, Peter DH, Slesak RA (2018) Logging debris and herbicide treatments improve growing conditions for planted Douglas-fir on a droughty forest site invaded by Scotch broom. Forest Ecology and Management 417, 31–39.
| Logging debris and herbicide treatments improve growing conditions for planted Douglas-fir on a droughty forest site invaded by Scotch broom.Crossref | GoogleScholarGoogle Scholar |
Hasse DL (2007) Morphological and Physiological Evaluations of Seedling Quality. In ‘National Proceedings: Forest and Conservation Nursery Associations 2006’. 19–22 June 2006, Oregon. (Eds LE Riley, RK Dumroese, TD Landis) pp. 3–8. (USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA)
Heinselman ML (1981) Fire and succession in the conifer forests of northern North America. In ‘Forest Succession: Concepts and Application’. (Eds DC West, HH Shugart, DB Botkin) pp. 374–405. (Springer-Verlag: New York, NY, USA)
Hobbs SD (1984) The influence of species and stocktype selection on stand establishment: an ecophysiological perspective. In ‘Seedling Physiology and Reforestation Success’. (Eds ML Duryea, GN Brown) pp. 179–224. (Martinus Nijhoff /Dr W. Junk Publishers: Dordrecht, The Netherlands)
Holmgren M, Scheffer M (2010) Strong facilitation in mild environments: the stress gradient hypothesis revisited. Journal of Ecology 98, 1269–1275.
| Strong facilitation in mild environments: the stress gradient hypothesis revisited.Crossref | GoogleScholarGoogle Scholar |
Jobidon R, Charette L, Bemier PY (1998) Initial size and competing vegetation effects on water stress and growth of Picea mariana (Mill.) BSP seedlings planted in three different environments. Forest Ecology and Management 103, 293–305.
| Initial size and competing vegetation effects on water stress and growth of Picea mariana (Mill.) BSP seedlings planted in three different environments.Crossref | GoogleScholarGoogle Scholar |
Jylhä P, Hytönen J (2006) Effect of vegetation control on the survival and growth of Scots pine and Norway spruce planted on former agricultural land. Canadian Journal of Forest Research 36, 2400–2411.
| Effect of vegetation control on the survival and growth of Scots pine and Norway spruce planted on former agricultural land.Crossref | GoogleScholarGoogle Scholar |
Keyser TL, Smith FW, Shepperd WD (2009) Short-term impact of post-fire salvage logging on regeneration, hazardous fuel accumulation, and understorey development in ponderosa pine forests of the Black Hills, SD, USA. International Journal of Wildland Fire 18, 451–458.
| Short-term impact of post-fire salvage logging on regeneration, hazardous fuel accumulation, and understorey development in ponderosa pine forests of the Black Hills, SD, USA.Crossref | GoogleScholarGoogle Scholar |
Kitzberger T, Steinaker DF, Veblen TT (2000) Effects of climatic variability on facilitation of tree establishment in northern Patagonia. Ecology 81, 1914–1924.
| Effects of climatic variability on facilitation of tree establishment in northern Patagonia.Crossref | GoogleScholarGoogle Scholar |
Kolb PF, Robberrecht R (1996) High temperature and drought stress effects on survival on Pinus ponderosa seedlings. Tree Physiology 16, 665–672.
| High temperature and drought stress effects on survival on Pinus ponderosa seedlings.Crossref | GoogleScholarGoogle Scholar | 14871688PubMed |
Lallement M, Tognetti C, Gobbi ME (2010) Post-fire restoration of native tree species: effects of wood shaving application. In ‘Principles and Practice of Forest Landscape Restoration. Case Studies from the Drylands of Latin America’. (Eds A Newton, N Tejedor) pp. 154–156. (IUCN: Gland, Switzerland).
Löf M, Gemmel P, Nilsson U, Welander NT (1998) The influence of site preparation on growth in Quercus robur L. seedlings in a southern Sweden clear-cut and shelterwood. Forest Ecology and Management 109, 241–249.
| The influence of site preparation on growth in Quercus robur L. seedlings in a southern Sweden clear-cut and shelterwood.Crossref | GoogleScholarGoogle Scholar |
Maguire DA, Mainwaring DB, Rose R, Garber SM, Dinger EJ (2009) Response of coastal Douglas-fir and competing vegetation to repeated and delayed weed control treatments during early plantation development. Canadian Journal of Forest Research 39, 1208–1219.
| Response of coastal Douglas-fir and competing vegetation to repeated and delayed weed control treatments during early plantation development.Crossref | GoogleScholarGoogle Scholar |
McIver JD, Starr L (2000) Environmental effects of postfire logging: literature review and annotated bibliography. General Technical Report PNW-GTR-486. USDA Forest Service (Portland, OR, USA)
Mohr Bell D (2015) Superficies afectadas por incendios en la región Bosque Andino Patagónico durante los veranos de 2013–2014 y 2014–2015 Nodo Regional Bosque Andino Patagónico (SAyDS – CIEFAP)
Mutch LS, Swetnam TW (1995) Effects of fire severity and climate on ring-width growth of giant sequoia after fire. In ‘Proceedings Symposium on Fire in Wilderness and Park Management: Past Lessons and Future Opportunities’, March 30-April 1 1993, Missoula MT (Eds JK Brown, RW Mutch, CW Spoon, RH Wakimoto) USDA Forest Service, Intermountain Research Station, General Technical Report INT-GTR-320, pp. 241–246. (Ogden, UT, USA)
Ouzts J, Kolb T, Huffman D, Meador AS (2015) Post-fire ponderosa pine regeneration with and without planting in Arizona and New Mexico. Forest Ecology and Management 354, 281–290.
| Post-fire ponderosa pine regeneration with and without planting in Arizona and New Mexico.Crossref | GoogleScholarGoogle Scholar |
Padilla FM, Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments. Frontiers in Ecology and the Environment 4, 196–202.
| The role of nurse plants in the restoration of degraded environments.Crossref | GoogleScholarGoogle Scholar |
Pastorino MJ, Aparicio A, Azpilicueta MM (2015). Regiones de Procedencia del Ciprés de la Cordillera y bases conceptuales para el manejo de sus recursos genéticos en Argentina. (Ediciones INTA: Buenos Aires, Argentina)
Pastorino MJ, Fariña MM, Bran D, Gallo LA (2006) Extremos geográficos de la distribución natural de Austrocedrus chilensis (Cupressaceae). Boletín de la Sociedad Argentina de Botánica 41, 307–311.
Perdomo M, De Agostini N, Cuevas J, Andenmatten E (2007) Reforestación con ciprés de la cordillera en la Reserva Forestal Loma del Medio-Río Azul (INTA-SFA). Avances logrados y desafíos futuros. In ‘EcoForestar 2007. Primera reunión sobre forestación en la patagonia. pp. 370–373. (Centro de Investigación y Extensión Forestal Andino Patagónico, Consejo Federal de Inversiones, Universidad Nacional de la Patagonia San Juan Bosco: Esquel, Argentina)
Pyne SJ (Ed.) (1984) ‘Introduction to Wildland Fire. Fire Management in the United States.’ (Wiley: New York, NY, USA)
Reader R, Wilson S, Belcher J, Wisheu I, Keddy P, Tilman D, Morris E, Grace J, McGraw J, Olff H (1994) Plant competition in relation to neighbor biomass: an intercontinental study with Poa pratensis. Ecology 75, 1753–1760.
| Plant competition in relation to neighbor biomass: an intercontinental study with Poa pratensis.Crossref | GoogleScholarGoogle Scholar |
Rose R, Rosner L (2005) Eighth-year response of Douglas-fir seedlings to area of weed control and herbaceus versus woody weed control. Annals of Forest Science 62, 481–492.
| Eighth-year response of Douglas-fir seedlings to area of weed control and herbaceus versus woody weed control.Crossref | GoogleScholarGoogle Scholar |
Secretaría de Ambiente y Desarrollo Sustentable de la Nación (2013) ‘Estadística de Incendios Forestales.’ (SAyDS: Buenos Aires, Argentina)
Thomas JW (Ed.) (1979) Wildlife habitats in managed forests – the Blue Mountains of Oregon and Washington. Agricultural Handbook 553 (USDA: Washington, DC, USA)
Urretavizcaya MF, Defossé G (2004) Soil seed bank of Austrocedrus chilensis (D.Don) Pic. Serm. et Bizarri related to different degrees of fire disturbance in two sites of southern Patagonia, Argentina. Forest Ecology and Management 187, 361–372.
| Soil seed bank of Austrocedrus chilensis (D.Don) Pic. Serm. et Bizarri related to different degrees of fire disturbance in two sites of southern Patagonia, Argentina.Crossref | GoogleScholarGoogle Scholar |
Urretavizcaya MF, Defossé G (2013) Effects of nurse shrubs and tree shelters on the survival and growth of two Austrocedrus chilensis seedling types in a forest restoration trial in semiarid Patagonia, Argentina. Annals of Forest Science 70, 21–30.
| Effects of nurse shrubs and tree shelters on the survival and growth of two Austrocedrus chilensis seedling types in a forest restoration trial in semiarid Patagonia, Argentina.Crossref | GoogleScholarGoogle Scholar |
Urretavizcaya MF, Defossé GE, Gonda HE (2012) Effect of sowing season, plant cover, and climatic variability on seedling emergence and survival in burned Austrocedrus chilensis forests. Restoration Ecology 20, 131–140.
| Effect of sowing season, plant cover, and climatic variability on seedling emergence and survival in burned Austrocedrus chilensis forests.Crossref | GoogleScholarGoogle Scholar |
Urretavizcaya MF, Gonda HE, Defossé GE (2017) Effects of post-fire plant cover in the performance of two cordilleran cypress (Austrocedrus chilensis) seedling stocktypes planted in burned forests of northeastern Patagonia, Argentina. Environmental Management 59, 419–430.
| Effects of post-fire plant cover in the performance of two cordilleran cypress (Austrocedrus chilensis) seedling stocktypes planted in burned forests of northeastern Patagonia, Argentina.Crossref | GoogleScholarGoogle Scholar | 27848000PubMed |
Veblen TT, Burns BR, Kitzberger T, Lara A, Villalba R (1995). The ecology of the conifers of southern South America. In ‘Ecology of the Southern Conifers’. (Eds NJ Enright, RS Hill.) pp. 120–129. (Melbourne University Press: Melbourne, Vic., Australia)
Veblen TT, Kitzberger T, Villalba R, Donnegan J (1999) Fire history in northern Patagonia: the roles of humans and climatic variation. Ecological Monographs 69, 47–67.
| Fire history in northern Patagonia: the roles of humans and climatic variation.Crossref | GoogleScholarGoogle Scholar |
Whisenant S (Ed.) (1999) ‘Repairing Damaged Wildlands: a Process-Orientated, Landscape-Scale Approach.’ (Cambridge University Press: Cambridge, UK)
Wright HA, Bailey AW (Eds) (1982) ‘Fire Ecology: United States and Southern Canada.’ (Wiley: New York, NY, USA)