

Body condition in an endangered minnow is variable but not strongly related to environmental conditions
Thomas P. Archdeacon A * and Paige M. Dunnum A
A * and Paige M. Dunnum A
A
Abstract
Seasonal patterns in body condition may help identify sensitive time periods or environmental stressors in fishes. Our objectives were to document spatiotemporal patterns of body condition of wild and hatchery-released Rio Grande silvery minnows (Hybognathus amarus), an endangered species endemic to the Rio Grande basin of North America. We collected fish weekly or bi-weekly from February to September, 2017 through 2019, in the Rio Grande, New Mexico, USA. Our time-series spanned an extreme drought year in 2018, allowing us to observe changes in body condition in relation to markedly different flow conditions. We collected body measurements from 2520 RGSM during the 3 years of our study. Spatiotemporal patterns in body condition were evident among and within years and sampling reaches, but within-sample variation in body condition was high. Annual patterns did not appear to be related to annual flow regime. Recaptured hatchery-reared fish were in slightly lower body condition, but followed the same seasonal patterns as did wild fish. Contrary to our expectations, body condition in both females and males was higher during the extreme drought year of 2018. Body condition may not be useful for identifying environmental stressors in small-bodied, short-lived fishes because individual variability is greater than spatial or temporal variation.
Keywords: breeding, drought, endangered species, fish, health, population supplementation, stream, stress.
Introduction
Studying how anthropogenic and natural stressors affect animal reproduction, health and overall well-being is important for reversing declining population trends (Edwards et al. 2019). In fishes, body condition is often linked to overall health. Many indices measure body ‘fatness’ for a given length (e.g. ‘body condition’) as a surrogate for measuring the overall health of the animal (Wuenschel et al. 2019). Body condition can be a proxy for lipid content and energy densities (Schloesser and Fabrizio 2017; Wuenschel et al. 2019; Brosset et al. 2023). In turn, lipid reserves are predictive of egg quality and condition of offspring (Muir et al. 2010; Brosset et al. 2016). Anthropogenic stressors can negatively affect body condition (Cavraro et al. 2019), which may reduce reproduction and recruitment (Gray et al. 2000; Balcombe et al. 2012) or survival (Pangle et al. 2004). Body condition indices can also be correlated with an array of traits, including movement behaviour (Kanno et al. 2023), temperature tolerance (Turko et al. 2020) and vigilance (Pollock et al. 2006).
Small-bodied fishes in the western United States are declining as water stress increases (Theis et al. 2023). Many fish species are facing multiple stressors, which can affect the outcome of flow restorations and hinder conservation efforts (Göthe et al. 2019). Basic biological and life-history information is lacking for many small-bodied fishes in the western United States, which is needed for designing conservation efforts (Theis et al. 2023). Quantifying seasonal variation in body condition over multiple years while accounting for reproductive development will be useful for identifying sensitive periods of time for the species, improving hatchery management and population supplementation practices, and identifying important environmental stressors.
The Rio Grande silvery minnow (Hybognathus amarus; hereafter RGS minnow) is a short-lived, small-bodied fish species endemic to the Rio Grande basin (Bestgen and Platania 1991). The Rio Grande silvery minnow belongs to a specialised reproductive guild of freshwater minnows characterised by pelagic broadcast-spawning of non-adhesive, nearly neutrally buoyant eggs (Platania and Altenbach 1998). Because they are small-bodied, RGS minnows are likely to use an income breeding strategy, in which available energy for reproduction coincides with daily foraging rather than stored energy reserves (McBride et al. 2015). Peak snowmelt spring flows usually coincide with RGS minnow spawning in April through early June, with a peak in May (Krabbenhoft et al. 2014; Archdeacon et al. 2024). However, the contemporary Rio Grande is flow-regulated and has both lower peak flows and total water volume compared with pre-1900 (Blythe and Schmidt 2018). Damming and flow regulation have influenced habitat, including channel narrowing (Swanson et al. 2011) and nearly annual channel drying overlapping the current distribution of RGS minnows (Archdeacon 2016; Archdeacon and Reale 2020). Water stress will increase with continued anthropogenic pressures and climate change (Mix et al. 2012; Holmes 2022), further threatening RGS minnows. Many minnows in the pelagic broadcast-spawning guild are found in the Great Plains of the western United States and are experiencing similar population declines (Dudley and Platania 2007; Worthington et al. 2018). Understanding how these pressures affect the well-being of fishes can help design mitigation practices.
Our objectives were to document temporal patterns of body condition in RGS minnows. We used weekly or biweekly body condition indices collected from both wild and hatchery-reared RGS minnows, from February through September, 2017–2019. Our time-series spanned an extreme drought year in 2018, allowing us to observe potential changes in body condition related to markedly different environmental conditions over the 3 years. We had the following two primary objectives: (1) to quantify spatiotemporal variability in body condition, including drought years, and (2) to compare differences in body condition between wild-born and hatchery-reared RGS minnows released and recaptured in Rio Grande, New Mexico. Our results may be informative for management of other small-bodied fishes with similar biology that are facing environmental threats.
Methods
The Middle Rio Grande (MRG) extends from downstream of Cochiti Dam to Elephant Butte Dam (Fig. 1), ~337 km, of which ~300 km is upstream of Elephant Butte Reservoir. The MRG is divided into the following four reaches by diversion dams: Cochiti Reach, Angostura Reach, Isleta Reach and San Acacia Reach (Fig. 1). Currently, the only extant population of RGS minnows occurs in the MRG (Bestgen and Platania 1991). Flows on the mainstem are heavily regulated for flood control, water storage and water deliveries. During our study, 2 years experienced annual flows well above median values (2017 and 2019) and 1 year had annual flows well below the median (Fig. 1). Estimated abundance of spawning adults was variable during the study; from spring of 2017 through spring 2018, abundance was high (>106) and the adult population was dominated by Age-1 fish in both 2017 and 2018 (Yackulic et al. 2022). Following a period of low spring runoff and extensive channel drying in summer of 2018, numbers fell two orders of magnitude by autumn, and the population was low in spring of 2019, composed mainly of hatchery and Age-2 fish (Archdeacon et al. 2020). Thus, we observed above-average adult density and high annual flows in 2017, high spring adult density and very low annual flows in 2018, and very high flows but low adult density from autumn 2018 through 2019. During the years of this study, all hatchery fish were given distinguishing marks (Archdeacon et al. 2023).
(a) Flow regime, (b) geographical locator, and Rio Grande silvery minnow collection locations in the Middle Rio Grande, New Mexico, 2017–2019.
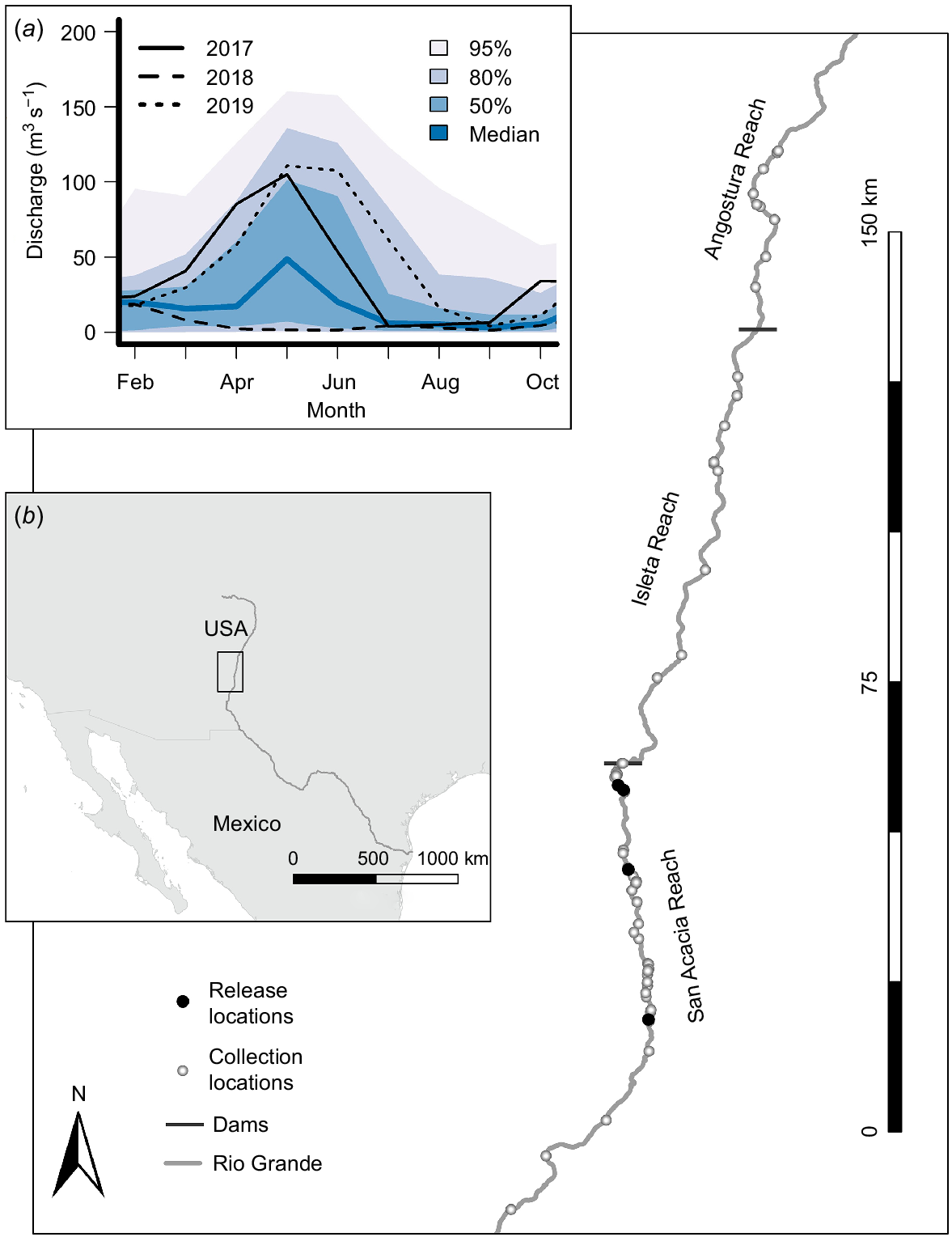
Fish were collected under Federal Permit TE676811-11 and New Mexico Permit 3764. We used several methods to collect fish depending on river discharge. We primarily used beach seines, but supplemented seining with fyke nets during high flows and backpack electrofishing during low flows. In 2017, we collected only wild fish from all three reaches to compare inter-reach differences. In 2018 and 2019, we focused on the San Acacia Reach and the comparison between hatchery and wild fish. Collections were made bi-weekly in 2017 and 2018. In 2019, we focused on the spawning season, with approximately monthly collections during February–April and July–September and weekly collections beginning in late April through early June. Isolated pools frequently form during flow-intermittency and are very shallow and have poor water quality, with fish crowding, high and fluctuating temperatures, and low dissolved oxygen concentration (Van Horn et al. 2022). Consequently, we did not collect fish from isolated pools for this study.
Rio Grande silvery minnow fish were euthanased with tricaine methanesulphonate and preserved in 10% zinc-buffered formalin during field collections, before storage in 70% ethanol following Grant and Archdeacon (2021). We measured standard length (to the nearest 0.01 mm) and blot-dried the fish prior to being weighed to the nearest 0.01 g. Standard length was corrected for shrinkage (Grant and Archdeacon 2021). Sex was determined by examination of gonads; ovaries from females were removed, weighed and subtracted from whole bodyweight to account for reproductive investment. Testes were not weighed for males because they make up only a small percentage of total bodyweight.
We used a scaled mass index (SMI) following Peig and Green (2009) as a proxy for overall health, calculated as follows:
where Wi is the weight of Fish i, Li is the length of Fish i, L0 is the average length of fish in all pooled samples, and bSMA is the slope of the standardised major regression from the log(weight)–log(length) relationship. In our study, the L0 for was 53.5 for males and 54.5 for females. For males, bSMA was 3.20 and for females, 3.24. We chose to use SMI because it is suitable for making comparisons across age classes, unlike the more traditional Fulton’s K (Peig and Green 2010).
To quantify seasonal trends, we used linear regressions including a third-order polynomial term for week of the year. To test our specific a priori hypotheses, we pooled data collected pre-spawn (April) each year and used a Student’s t-test or ANOVA to examine differences among reaches, years and wild-born or hatchery-reared fish. We used program R (ver. 4.3.2, R Foundation for Statistical Computing, Vienna, Austria, see https://www.r-project.org/) for model building. We used R package smatr (ver. 3.4.8, see https://cran.r-project.org/package=smatr/; Warton et al. 2012) to determine the bSMA.
Results
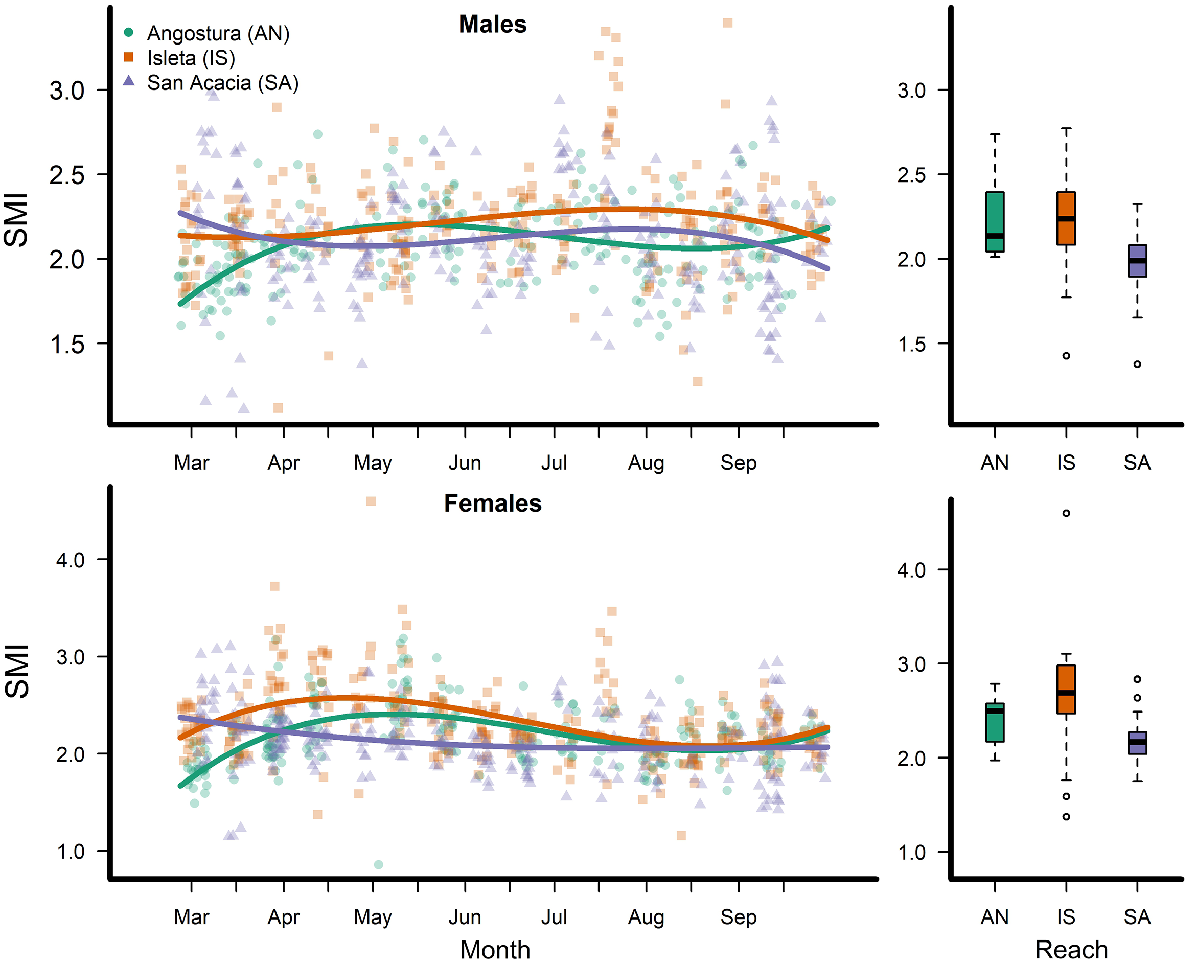
We collected 2520 total RGS minnows during the 3 years of our study (Table 1). Flow conditions in 2017 and 2019 were above median flow values, whereas flows in 2018 were near minimum observed values throughout the year (Fig. 1). Although spatiotemporal patterns in SMI were evident among and within years and sampling reaches, within-period variation in SMI was high (Fig. 2). Among the three reaches sampled in 2017, males from the San Acacia Reach had lower SMI than those from the other reaches during the pre-spawn period, with a SMI of 1.97 (s.e. = 0.037, P = 0.0009). Male SMI prior to spawning in the Angostura and Isleta Reaches were not statistically different, averaging 2.24 (s.e. = 0.068). Female SMI differed significantly in all three reaches during the pre-spawn period: in the Angostura Reach, mean SMI was 2.40 (s.e. = 0.084), in the San Acacia Reach, mean SMI was 2.18 (s.e. = 0.106, P = 0.0420), and in the Isleta Reach, mean SMI was 2.66 (s.e. = 0.106, P = 0.042). Length and mass of RGS minnows collected by year, reach, origin and sex are shown in Supplementary Fig. S1 and S2.
| Year | Reach | Males | Females | Origin | |
|---|---|---|---|---|---|
| 2017 | Angostura | 221 | 251 | Wild | |
| 2017 | Isleta | 243 | 279 | Wild | |
| 2017 | San Acacia | 270 | 287 | Wild | |
| 2018 | San Acacia | 150 | 147 | Wild | |
| 2018 | San Acacia | 187 | 199 | Hatchery | |
| 2019 | San Acacia | 27 | 45 | Wild | |
| 2019 | San Acacia | 78 | 136 | Hatchery | |
| Total | 1176 | 1344 |
Left column shows observations (points) and mean (lines) scaled mass index (SMI) of Rio Grande silvery minnows in three reaches of the Middle Rio Grande (MRG), New Mexico, in 2017. Right column shows boxplots of SMI from each reach of the MRG in the pre-spawning period (April) of 2017.
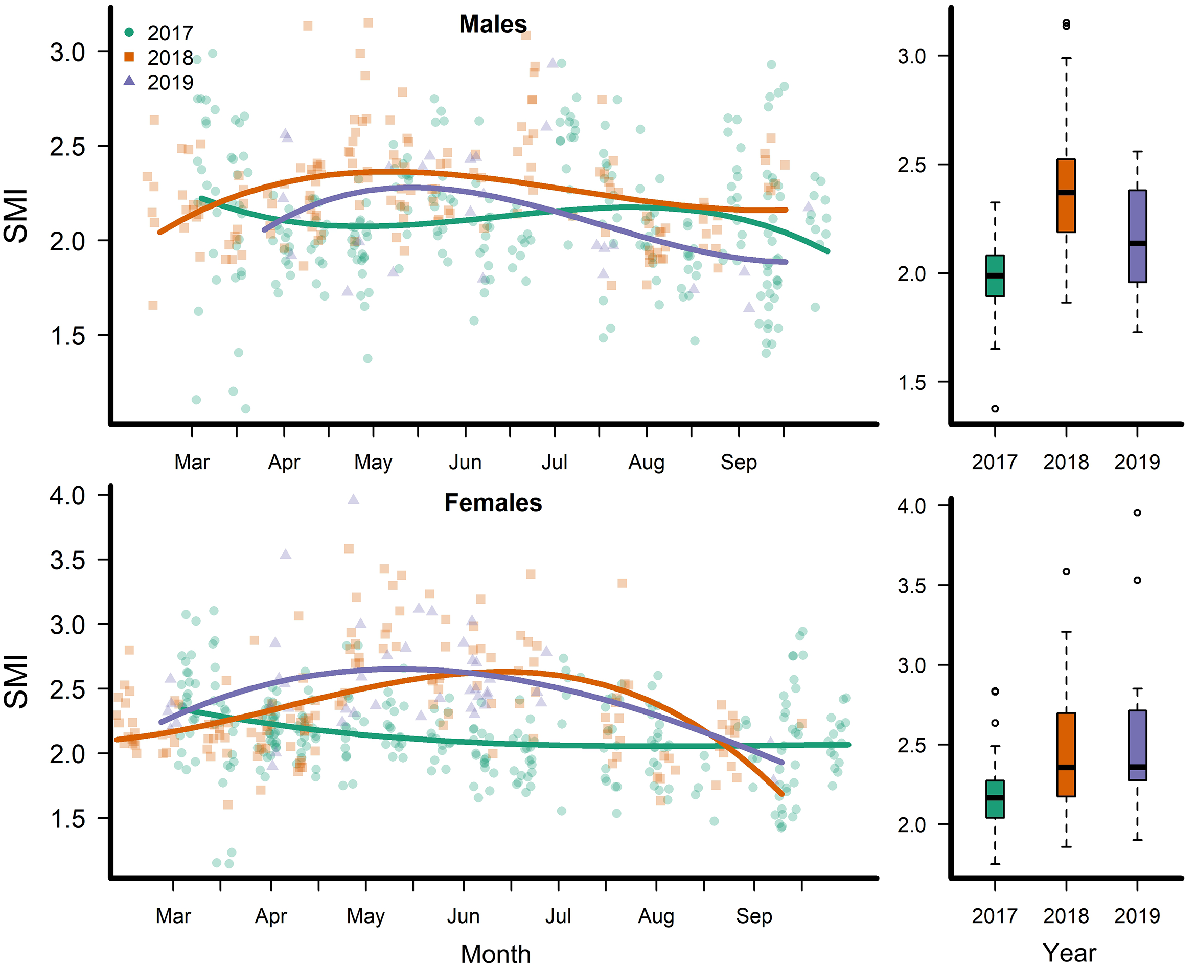
Despite different flow conditions among the 3 years, SMI in both females and males was lower during the high-flow year of 2017 and higher during the extreme drought year of 2018 (Fig. 3). During the pre-spawn period, male mean SMI in 2017 was 1.97 (s.e. = 0.042) and in 2019 it was not statistically different (P = 0.077), whereas in 2018 mean male SMI was 2.40 (s.e. = 0.060, P < 0.0001). Female SMI differed among all years and was highest in 2019. Female SMI was 2.18 (s.e. = 0.063) in 2017, 2.42 in 2018 (s.e. = 0.234, P = 0.014) and 2.58 (s.e. = 0.128, P = 0.003) in 2019.
Left column shows observations (points) and mean (lines) scaled mass index (SMI) of Rio Grande silvery minnows in the San Acacia Reach of the Middle Rio Grande, New Mexico, 2017–2019. Right column shows boxplots of SMI from the San Acacia Reach of the Middle Rio Grande in the pre-spawning period (April).
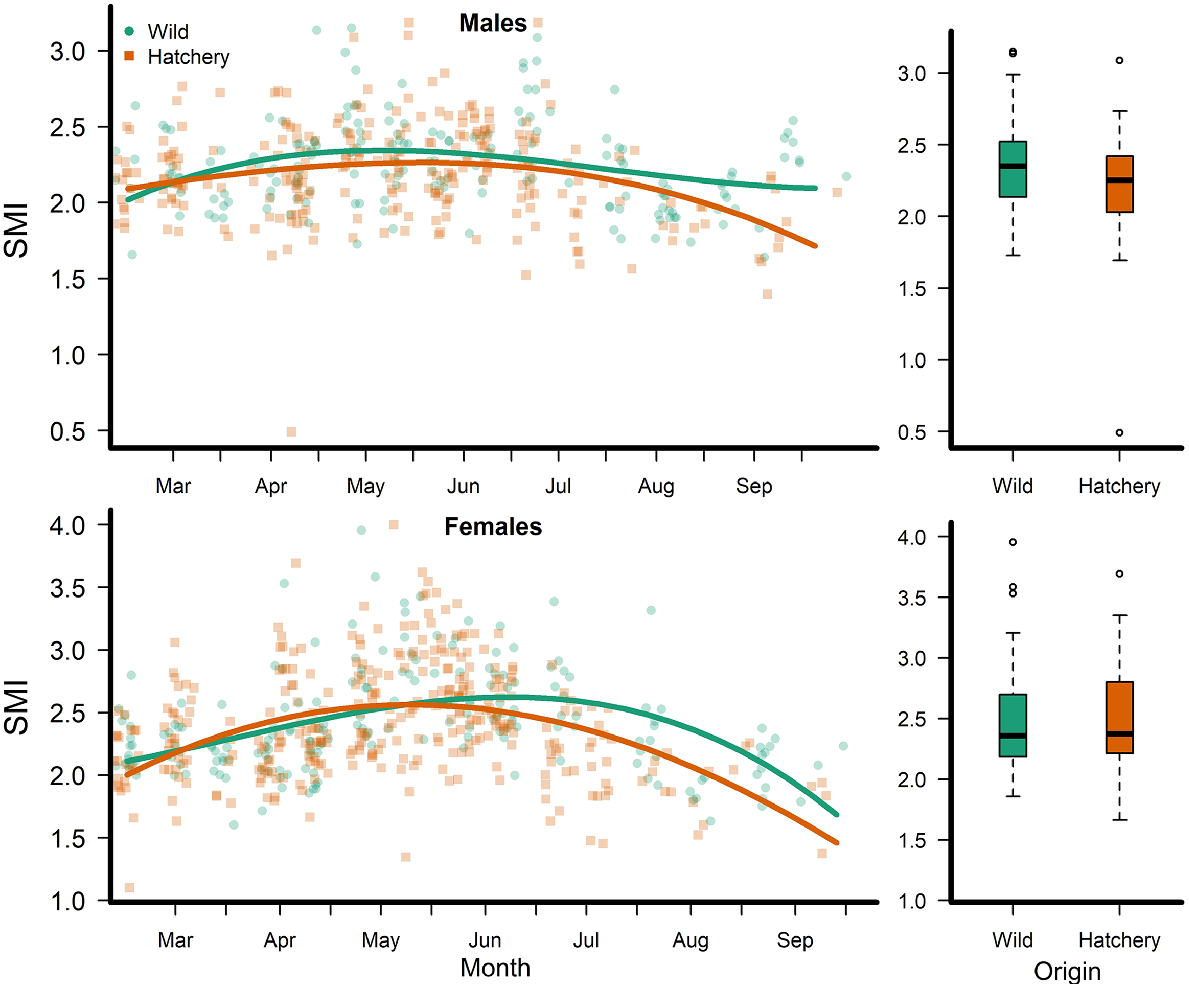
Both male and female hatchery fish had slightly lower overall body condition than did wild fish (Fig. 4). However, SMI during the pre-spawn period was 2.46 (s.e. = 0.064) and not statistically different between hatchery-reared and wild-born females (P = 0.639). Hatchery-reared males had lower SMI, with a mean of 2.21 (s.e. = 0.069, P = 0.036), than did the wild-born males, with a mean SMI of 2.36 (s.e. = 0.053).
Left column shows observations (points) and mean (lines) scaled mass index (SMI) of wild-born and hatchery Rio Grande silvery minnows in the San Acacia Reach of the Middle Rio Grande, New Mexico, 2018–2019. Right column shows boxplots of SMI from wild-born and hatchery Rio Grande silvery minnows during the pre-spawning period (April).
Discussion
Surprisingly, we observed only weak spatial and temporal variation in body condition. Although annual flow regimes were markedly different, both males and females had lower body condition in 2017 than in other years, despite high-flow conditions. Individual variation in body condition within a sampling period was generally much greater than was variation across seasonal or spatial trends. Fish in both very poor and very good relative condition were collected during almost every sampling event. Thus, time-series of average body condition may not be useful for identifying environmental stressors in small-bodied fishes because individual variability is greater than spatial or temporal variation.
Contrary to our expectations of lowered body condition during low-flow years, we observed higher SMI during the drought year of 2018. Because water quality, habitat quality and anthropogenic stressors can negatively affect body condition (Oliva-Paterna et al. 2003; Maceda-Veiga et al. 2014), we assumed RGS minnow body condition would have a negative response to lower annual flow regime. Other studies have found similar positive responses in intermittent streams (Cruz et al. 2022), or no difference in body condition between perennial and intermittent sites potentially because feeding became more efficient (Glarou et al. 2019). For RGS minnows, channel intermittency can result in catastrophic population losses when fish are trapped in isolated pools, regardless of body condition of individual fish, because of complete desiccation of pools (Archdeacon and Reale 2020). Recruitment is also limited during low-flow years (Yackulic et al. 2022). Therefore, body condition may not be as important of a predictor of stress in RGS minnows as are other indicators.
A possible explanation for the lack of seasonal variation in condition during pre-spawning is that the RGS minnow is likely to use an income breeding strategy rather than a capital breeding strategy. Income breeding is a strategy often used by small fish that is characterised by a prolonged spawning season driven by local resource availability (McBride et al. 2015). Capital-breeding fishes store energy during pre-spawning seasons and use body reserves during a short spawning season, whereas income-breeding fishes acquire and allocate energy to reproduction more immediately. Small-bodied fishes may not show strong population-level changes in body condition because energy reserves are relatively small compared with those of large-bodied, capital-breeding fishes. Changes in body condition may be reflecting conditions individual fish are experiencing in the days prior, and not necessarily reflecting long-term stressors. Thus, body condition prior to spawning may affect individual fish, but is unlikely to be a driver of recruitment for RGS minnows.
Post-release monitoring of hatchery-reared fish is critical for evaluating conservation efforts, but is often overlooked (Rytwinski et al. 2021). Our study was possible because nearly all hatchery RGS minnows have been given identifying marks prior to release (Archdeacon et al. 2023). We were able to determine that hatchery fish had trends in body condition similar to those of wild-born fish. Our observations suggest that after high initial mortality following release into the river (75–85%, Yackulic et al. 2022), hatchery fish behave similarly to wild fish. Stress associated with release into the wild is likely to act as a harsh filter for fish that are already in poor condition or are poorly adapted to riverine conditions. This is an important post-release observation because poor body condition can reduce reproductive success (Gray et al. 2000). Current augmentation guidelines are based on length and Fulton’s K and fish from distinct hatcheries must average at least 35-mm standard length and average Fulton’s K of >0.80. Up to 300,000 RGS minnows may be released annually, depending on numbers of wild fish. We suggest that increasing length and condition criteria for hatchery-reared RGS minnows could improve post-release survival and reproductive fitness (e.g. 75% must meet minimum criteria), as observed for other species (Fonken et al. 2023). Doing this should lessen genetic drift associated with hatchery augmentation (Osborne et al. 2024) by increasing the number of females contributing to reproductive efforts.
We acknowledge several limitations in our study. First, although we corrected for the length of fish in our study, mass increases after preservation and can introduce a positive bias in condition in freshwater fishes (Nordeide 2020). Our estimates of L0 are representative of fish lengths during other years, but the estimate of BSMA may not be, owing to changes in shape and weight of fish after preservation. Therefore, comparison with other observations should be cautious, and development of SMI from live specimens may be beneficial. Second, although condition and lipid content are correlated in RGS minnows (Powell et al. 2017), other lethal sampling techniques more accurately represent overall health and may be better predictors of fitness, such as hepatosomatic index, lipid content, or dry-matter content (Brosset et al. 2023; Kanno et al. 2023), although lipid content may not reflect evolutionary fitness (Wilder et al. 2016). Last, our study was focused on adult fish prior to spawning through summer low flows. Our samples were representative of the age structure of adult fish, composed almost exclusively of Age-1 fish in 2017 and 2018, with a small number of Age-2 fish appearing in 2019 (Archdeacon et al. 2020). However, future studies of Age-0 fish from late summer through winter may help explain observations in adult fish.
Although inter- and intra-seasonal patterns in RGS minnow body condition were evident, relationships did not appear to be related to flow conditions. Contrary to our expectations, body condition was not negatively affected by drought. Importantly, we found only small differences in body condition during the pre-spawn period between hatchery-reared and wild-born RGS minnows. Flow management actions should focus on improving recruitment and summer survival for RGS minnows, because body condition is highly variable prior to spawning and does not appear to be strongly linked to flow conditions. Nonetheless, non-flow management actions related to hatchery practices should aim to improve size and condition of RGS minnows prior to release, to improve efficacy of hatchery fish for improving demographic and genetic resilience.
Data availability
Data that support the findings of this study may be requested from the corresponding author.
Disclaimer
The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the US Fish and Wildlife Service.
Acknowledgements
We thank Josh Grant, Eileen Henry, Mo Hobbs, Scott Mullner and Lyle Thomas for field and laboratory assistance. We thank two anonymous reviewers for constructive comments that improved the manuscript.
References
Archdeacon TP (2016) Reduction in spring flow threatens Rio Grande silvery minnow: trends in abundance during river intermittency. Transactions of the American Fisheries Society 145, 754-765.
| Crossref | Google Scholar |
Archdeacon TP, Reale JK (2020) No quarter: lack of refuge during flow intermittency results in catastrophic mortality of an imperiled minnow. Freshwater Biology 65, 2108-2123.
| Crossref | Google Scholar |
Archdeacon TP, Diver-Franssen TA, Bertrand NG, Grant JD (2020) Drought results in recruitment failure of Rio Grande silvery minnow (Hybognathus amarus), an imperiled, pelagic broadcast-spawning minnow. Environmental Biology of Fishes 103, 1033-1044.
| Crossref | Google Scholar |
Archdeacon TP, Dudley RK, Remshardt WJ, Knight W, Ulibarri M, Gonzales EJ (2023) Hatchery supplementation increases potential spawning stock of Rio Grande silvery minnow after population bottlenecks. Transactions of the American Fisheries Society 152, 187-200.
| Crossref | Google Scholar |
Archdeacon TP, Diver TA, Grey RE (2024) Reproductive phenology of wild and hatchery-reared Rio Grande silvery minnow. Report submitted to the US Bureau of Reclamation. (Albuquerque Area Office: Albuquerque, NM, USA) doi:10.13140/RG.2.2.29854.92485
Balcombe SR, Lobegeiger JS, Marshall SM, Marshall JC, Ly D, Jones DN (2012) Fish body condition and recruitment success reflect antecedent flows in an Australian dryland river. Fisheries Science 78, 841-847.
| Crossref | Google Scholar |
Bestgen KR, Platania SP (1991) Status and conservation of the Rio Grande silvery minnow, Hybognathus amarus. The Southwestern Naturalist 36(2), 225-232.
| Crossref | Google Scholar |
Blythe TL, Schmidt JC (2018) Estimating the natural flow regime of rivers with long-standing development: the northern branch of the Rio Grande. Water Resources Research 54, 1212-1236.
| Crossref | Google Scholar |
Brosset P, Lloret J, Muñoz M, Fauvel C, Van Beveren E, Marques V, Fromentin J-M, Ménard F, Saraux C (2016) Body reserves mediate trade-offs between life-history traits: new insights from small pelagic fish reproduction. Royal Society Open Science 3, 160202.
| Crossref | Google Scholar | PubMed |
Brosset P, Averty A, Mathieu-Resuge M, Schull Q, Soudant P, Lebigre C (2023) Fish morphometric body condition indices reflect energy reserves but other physiological processes matter. Ecological Indicators 154, 110860.
| Crossref | Google Scholar |
Cavraro F, Bettoso N, Zucchetta M, D’Aietti A, Faresi L, Franzoi P (2019) Body condition in fish as a tool to detect the effects of anthropogenic pressures in transitional waters. Aquatic Ecology 53, 21-35.
| Crossref | Google Scholar |
Cruz A, Llinares C, Martín-Barrio I, Castillo-García G, Arana P, García-Berthou E, Fletcher DH, Almeida D (2022) Comparing morphological, parasitological, and genetic traits of an invasive minnow between intermittent and perennial stream reaches. Freshwater Biology 67, 2035-2049.
| Crossref | Google Scholar |
Dudley RK, Platania SP (2007) Flow regulation and fragmentation imperil pelagic-spawning riverine fishes. Ecological Applications 17, 2074-2086.
| Crossref | Google Scholar | PubMed |
Fonken DR, Conner MM, Walsworth TE, Thompson PD (2023) Benefits of stocking fewer but larger individuals with implications for native fish recovery. Canadian Journal of Fisheries and Aquatic Sciences 80, 439-450.
| Crossref | Google Scholar |
Glarou M, Vourka A, Vardakas L, Andriopoulou A, Skoulikidis N, Kalogianni E (2019) Plasticity in life history traits of a cyprinid fish in an intermittent river. Knowledge & Management of Aquatic Ecosystems 420, 25.
| Crossref | Google Scholar |
Grant JD, Archdeacon TP (2021) Shrinkage of Rio Grande silvery minnow after preservation in formalin and storage in ethanol. Western North American Naturalist 81, 544-549.
| Crossref | Google Scholar |
Gray SC, De Silva SS, Ingram BA, Gooley GJ (2000) Effects of river impoundment on body condition and reproductive performance of the Australian native fish, Macquarie perch (Macquaria australasica). Lakes & Reservoirs: Science, Policy and Management for Sustainable Use 5, 281-291.
| Crossref | Google Scholar |
Göthe E, Degerman E, Sandin L, Segersten J, Tamario C, Mckie BG (2019) Flow restoration and the impacts of multiple stressors on fish communities in regulated rivers. Journal of Applied Ecology 56, 1687-1702.
| Crossref | Google Scholar |
Holmes R (2022) Water woes worsen: Middle Rio Grande reservoir modeling projects declining water availability under climate change simulations. MSc thesis, Michigan Technological University, Houghton, MI, USA. Available at https://doi.org/10.37099/mtu.dc.etdr/1351
Kanno Y, Locklear ML, Platis NM, Lewis ST (2023) Body condition metrics explain fish movement in experimental streams. Journal of Zoology 320, 18-28.
| Crossref | Google Scholar |
Krabbenhoft TJ, Platania SP, Turner TF (2014) Interannual variation in reproductive phenology in a riverine fish assemblage: implications for predicting the effects of climate change and altered flow regimes. Freshwater Biology 59, 1744-1754.
| Crossref | Google Scholar |
Maceda-Veiga A, Green AJ, De Sostoa A (2014) Scaled body-mass index shows how habitat quality influences the condition of four fish taxa in north-eastern Spain and provides a novel indicator of ecosystem health. Freshwater Biology 59, 1145-1160.
| Crossref | Google Scholar |
McBride RS, Somarakis S, Fitzhugh GR, Albert A, Yaragina NA, Wuenschel MJ, Alonso-Fernández A, Basilone G (2015) Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish and Fisheries 16, 23-57.
| Crossref | Google Scholar |
Mix K, Lopes VL, Rast W (2012) Environmental drivers of streamflow change in the Upper Rio Grande. Water Resources Management 26, 253-272.
| Crossref | Google Scholar |
Muir AM, Sutton TM, Arts MT, Claramunt RM, Ebener MP, Fitzsimons JD, Johnson TB, Kinnunen RE, Koops MA, Sepúlveda MM (2010) Does condition of lake whitefish spawners affect physiological condition of juveniles? Journal of Great Lakes Research 36(Suppl. 1), 92-99.
| Crossref | Google Scholar |
Nordeide JT (2020) Accuracy of body mass estimates of formalin-preserved fish – a review. Journal of Fish Biology 96, 288-296.
| Crossref | Google Scholar | PubMed |
Oliva-Paterna FJ, Miñnano PA, Torralva M (2003) Habitat quality affects the condition of Barbus sclateri in Mediterranean semi-arid streams. Environmental Biology of Fishes 67, 13-22.
| Crossref | Google Scholar |
Osborne MJ, Archdeacon TP, Yackulic CB, Dudley RK, Caeiro-Dias G, Turner TF (2024) Genetic erosion in an endangered desert fish during a megadrought despite long-term supportive breeding. Conservation Biology 38, e14154.
| Crossref | Google Scholar | PubMed |
Pangle KL, Sutton TM, Kinnunen RE, Hoff MH (2004) Overwinter survival of juvenile lake herring in relation to body size, physiological condition, energy stores, and food ration. Transactions of the American Fisheries Society 133, 1235-1246.
| Crossref | Google Scholar |
Peig J, Green AJ (2009) New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos 118, 1883-1891.
| Crossref | Google Scholar |
Peig J, Green AJ (2010) The paradigm of body condition: a critical reappraisal of current methods based on mass and length. Functional Ecology 24, 1323-1332.
| Crossref | Google Scholar |
Platania SP, Altenbach CS (1998) Reproductive strategies and egg types of seven Rio Grande Basin cyprinids. Copeia 1998(3), 559-569.
| Crossref | Google Scholar |
Pollock MS, Pollock RJ, Chivers DP (2006) Effects of body size, body condition, and breeding state on responses to alarm cues by fathead minnows. Canadian Journal of Zoology 84, 1351-1357.
| Crossref | Google Scholar |
Powell MS, Hardy RW, Hutson AM, Toya LA, Tave D (2017) Comparison of body composition and fatty acid profiles between wild and cultured Rio Grande silvery minnows. Journal of Fish and Wildlife Management 8, 487-496.
| Crossref | Google Scholar |
Rytwinski T, Kelly LA, Donaldson LA, Taylor JJ, Smith A, Drake DAR, Martel AL, Geist J, Morris TJ, George AL, Dextrase AJ, Bennett JR, Cooke SJ (2021) What evidence exists for evaluating the effectiveness of conservation-oriented captive breeding and release programs for imperilled freshwater fishes and mussels? Canadian Journal of Fisheries and Aquatic Sciences 78(9), 1332-1346.
| Crossref | Google Scholar |
Schloesser RW, Fabrizio MC (2017) Condition indices as surrogates of energy density and lipid content in juveniles of three fish species. Transactions of the American Fisheries Society 146, 1058-1069.
| Crossref | Google Scholar |
Swanson BJ, Meyer GA, Coonrod JE (2011) Historical channel narrowing along the Rio Grande near Albuquerque, New Mexico in response to peak discharge reductions and engineering: magnitude and uncertainty of change from air photo measurements. Earth Surface Processes and Landforms 36, 885-900.
| Crossref | Google Scholar |
Theis S, Castellanos-Acuña D, Hamann A, Poesch MS (2023) Small-bodied fish species from the western United States will be under severe water stress by 2040. Conservation Science and Practice 5(1), e12856.
| Crossref | Google Scholar |
Turko AJ, Nolan CB, Balshine S, Scott GR, Pitcher TE (2020) Thermal tolerance depends on season, age and body condition in imperilled redside dace Clinostomus elongatus. Conservation Physiology 8, coaa062.
| Crossref | Google Scholar |
Van Horn DJ, Reale JK, Archdeacon TP (2022) Water quality in three potential drought refuges in an arid-land river: assessing habitat suitability for at-risk fish species. Knowledge & Management of Aquatic Ecosystems 423, 7.
| Crossref | Google Scholar |
Warton DI, Duursma RA, Falster DS, Taskinen S (2012) smatr 3 – an R package for estimation and inference about allometric lines. Methods in Ecology and Evolution 3, 257-259.
| Crossref | Google Scholar |
Wilder SM, Raubenheimer D, Simpson SJ (2016) Moving beyond body condition indices as an estimate of fitness in ecological and evolutionary studies. Functional Ecology 30, 108-115.
| Crossref | Google Scholar |
Worthington TA, Echelle AA, Perkin JS, et al. (2018) The emblematic minnows of the North American Great Plains: a synthesis of threats and conservation opportunities. Fish and Fisheries 19, 271-307.
| Crossref | Google Scholar |
Wuenschel MJ, McElroy WD, Oliveira K, McBride RS (2019) Measuring fish condition: an evaluation of new and old metrics for three species with contrasting life histories. Canadian Journal of Fisheries and Aquatic Sciences 76, 886-903.
| Crossref | Google Scholar |
Yackulic CB, Archdeacon TP, Valdez RA, Hobbs M, Porter MD, Lusk J, Tanner A, Gonzales EJ, Lee DY, Haggerty GM (2022) Quantifying flow and nonflow management impacts on an endangered fish by integrating data, research, and expert opinion. Ecosphere 13, e4240.
| Crossref | Google Scholar |