

Two new genera of land snail from dry subtropical forests of eastern Australia: Brigaladra gen. nov. and Euryladra gen. nov. (Eupulmonata: Camaenidae)
Lorelle Stanisic A B * , Frank Köhler C and Carmel McDougall A
A B * , Frank Köhler C and Carmel McDougall A
A School of Environment and Science, Australian Rivers Institute, Nathan Campus, Griffith University, 170 Kessels Road, Nathan, Qld 4111, Australia.
B Queensland Museum, PO Box 3300, South Brisbane, Qld 4101, Australia.
C Australian Museum Research Institute, 1 William Street, Sydney, NSW 2010, Australia.
Handling Editor: Nerida Wilson
Invertebrate Systematics 36(6) 506-532 https://doi.org/10.1071/IS21075
Submitted: 5 November 2021 Accepted: 15 February 2022 Published: 29 June 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Figuladra is a poorly understood genus of camaenid land snail endemic to subtropical eastern Australia, comprising species that inhabit either dry rainforests in the coastal hinterland or dry sclerophyll forests further inland. Given that the genus occurs in areas that have seen extensive land clearing, such as the inland brigalow scrub (savannah) and coastal vine thickets (dry rainforest), resolution of this genus is required to inform conservation efforts within these critical landscapes. Here we use an integrative approach based on comparative analyses of mitochondrial DNA sequences and key morphological features to review the systematic relationships within the group. Specifically, we performed Maximum Likelihood and Bayesian analyses of concatenated DNA sequences of two partial mitochondrial genes (16S and COI) to reconstruct the phylogenetic relationships of Figuladra. We subsequently examined features of the reproductive system that are considered to be useful in diagnosing genera (whole genitalia) and species (penial architecture) within the Camaenidae. Based on patterns of molecular differentiation and an assessment of the anatomy, we propose a more restrictive definition of Figuladra and describe two new monotypic genera for species that were previously assigned to Figuladra; Euryladra gen. nov. for Varohadra incei mattea Iredale, 1933 and Brigaladra gen. nov. for Varohadra volgiola Iredale, 1933. These two new genera differ from Figuladra sensu stricto in exhibiting distinctive epiphallic coiling patterns and anatomical differences of the penis–epiphallus configuration. The study shows that these three genera, and allied taxa, reveal contrasting patterns of diversity and distribution in two neighbouring habitat types: high levels of diversity and endemism in fire-sensitive seasonal subtropical forests juxtaposed against low diversity in large intervening areas of regularly burnt savannah and open woodlands.
Keywords: anatomy, camaenid, land snail, mitochondrial DNA, morphology, open woodland, Queensland, savannah, vine thicket.
Introduction
The Camaenidae is a family of rock, forest and woodland snails that are widespread across Australia, absent only from south-western Western Australia, Tasmania and the sandy deserts of central Australia (Stanisic et al. 2010, 2018; Hugall and Stanisic 2011). There are two centres of diversity for camaenids, the Kimberley region of the far north-west and the ranges of eastern Australia. Australian camaenids inhabit rainforests (moist tropical, sub-tropical and vine thickets) and woodlands (open eucalyptus forests) to drier savannahs (brigalow, scrub) (e.g. Hugall et al. 2002; Stanisic et al. 2010, 2018; Köhler 2011a; Parkyn and Newell 2013).
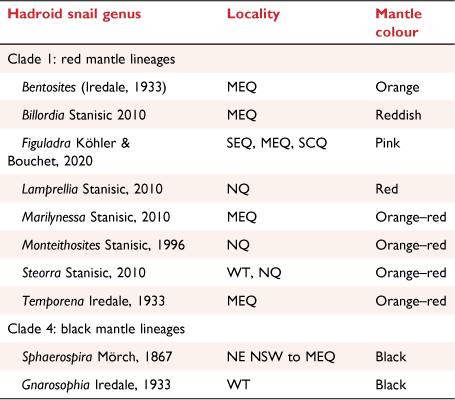
In eastern Australia, there are several genera of so-called ‘large forest hadroid snails’ (large, often striped camaenids). Figuladra Köhler & Bouchet, 2020 is one of these genera from north-eastern Australia (Queensland) with a distribution from the moister coastal areas to the drier interior west of the Great Dividing Range, mostly inhabiting dry vine thickets and open woodlands. Most species occur close to the coast, but two additional species inhabit dry vine thickets in mid-eastern Queensland to the west of the mountains that run along Australia’s east coast. Most Figuladra species are considered to be narrow range endemics (Stanisic et al. 2010). Currently, the identification and delineation of species in this genus rests predominantly on shell characters such as colour, size and banding patterns, but details of the reproductive anatomy and phylogenetic relationships are largely unknown. Figuladra taxonomy is complicated by a high intraspecific variation in shell colour and banding pattern in some of the species. Given that the genera occur in areas that have seen extensive land clearing, such as the inland brigalow scrub (savannah) and coastal vine thickets (dry rainforest), resolution of this genus is required to inform conservation efforts within these critical landscapes.
Figuladra has a particularly convoluted taxonomic history. The eastern Australian hadroids were once all placed in a single genus, Sphaerospira Mörch by Smith (1992). However, Stanisic et al. (2010) restricted Sphaerospira to some species more closely associated with rainforests along the eastern seaboard and allocated many species to other genera. This subdivision of Sphaerospira was made with main emphasis on animal colour, particularly that of the mantle, and shell morphology. Unpublished details of the reproductive anatomy formed an additional tier for the separation of these genera (Stanisic et al. 2010, p. 7). Figuladra in the current configuration was delineated by Stanisic et al. (2010) using a name first introduced by Iredale (1933). However, Köhler and Bouchet (2020) found that Iredale’s original name was unavailable due to the lack of a formal diagnosis (Art. 13.4 of the Code; International Trust for Zoological Nomenclature 2012) and that this name had not been made available subsequently. To maintain the previous treatment of Stanisic et al. (2010), these authors described Figuladra as a new genus with the type species Helix curtisiana Pfeiffer, 1864. Prior to Stanisic’s treatment, Iredale (1937, p. 30) placed Figuladra Iredale, 1933 in synonymy of Varohadra Iredale, 1933. However, Varohadra Iredale, 1933 is also an unavailable name (Köhler and Bouchet 2020). Smith (1992) correctly recognised Figuladra and Varohadra as nomina nuda and placed all species previously classified under these names as members of Sphaerospira Mörch, 1867 citing Bishop’s (1978) anatomical study of S. fraseri Griffith and Pidgeon, 1833 as the reason for this taxonomic decision. Stanisic and Stanisic (2020) resolved historical taxonomic confusion surrounding Figuladra species described before 1900 thus allowing for revisions of Figuladra to be undertaken unencumbered by taxonomic ambiguity.
Molecular phylogenetic data for eastern Australian camaenids were limited until the landmark study by Hugall and Stanisic (2011). The phylogenetic tree of Hugall and Stanisic (2011, fig. 7) covered ~90% of the eastern Australian camaenid species accepted at the time and demonstrated that the traditional, largely shell-based classification was misled by convergence in shell characters. Shell morphology has proven to be of limited value as a taxonomic marker when used to delimit taxa in camaenids without regard to other sources of evidence (Köhler and Johnson 2012; Perez et al. 2014; Köhler and Criscione 2015). Similarly, Hugall and Stanisic (2011) concluded that the taxonomic classification of east coast camaenids had been misdirected by parallelism in shell form, but also stated that comparative shell morphology has a place within the framework of an integrative taxonomy. In contrast to the limited use of shell morphology, comparative anatomy of the terminal reproductive organs has been shown to be taxonomically informative in distinguishing genera (whole genitalia) and species (penial architecture) within the Camaenidae (e.g. Solem 1979, 1985, 1992; Stanisic 1996, 2013, 2016; Köhler 2011a, 2011b; Criscione and Köhler 2014a).
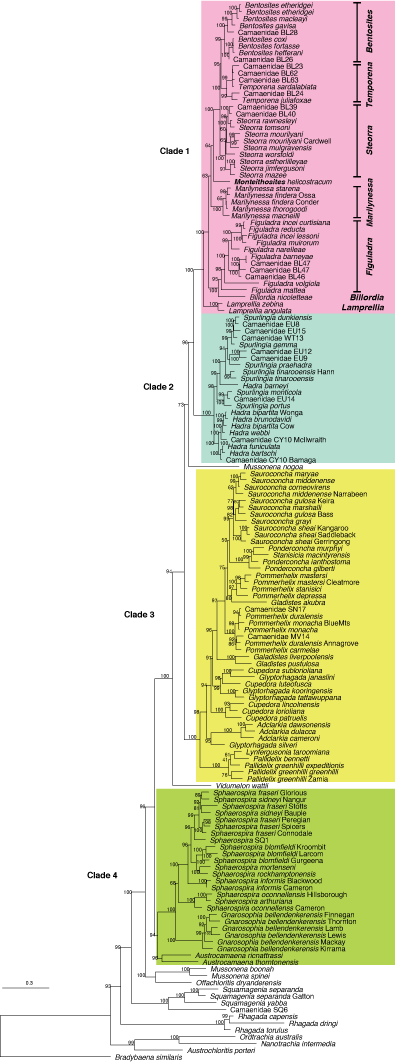
Hugall and Stanisic (2011) showed that there were four major clades within the ‘hadroid complex’ of camaenids (Clades 1–4). Clades 1 and 4 have overlapping distributions from far north-eastern Australia down to north-eastern New South Wales. However, the distributions are somewhat complementary, with Clade 1 taxa tending to occur in dry seasonal forest habitats (often referred to as vine thickets, scrub or dry rainforest in Australia), whereas Clade 4 taxa tend to occur in wetter rainforest habitats. Clade 2 comprised species that are concentrated in north-eastern Australia whereas Clade 3 comprised species in south-eastern Australia. Sphaerospira, as previously construed, represented a polyphyletic group spanning lineages in both Clade 1 and Clade 4 (Fig. 1). Hugall and Stanisic subsequently confirmed that the type species, Sphaerospira fraseri (Griffith & Pidgeon, 1833), belonged to Clade 4. This phylogeny illustrated that many species from Clade 1, thought to belong to Sphaerospira, were no longer in that genus. Figuladra was applied to the lineage containing mattea, volgiola and incei curtisiana (see Fig. 1) within Clade 1 by Stanisic et al. (2010). The apparent closest relative to this clade is the genus Billordia that comprises two species that occur in vine thickets on rocky screes in south-central Queensland.
|
The taxonomy of Figuladra clearly needs revision. The phylogeny of Hugall and Stanisic (2011) provides the basis for further investigations and a starting point for an in-depth examination of the Figuladra lineage containing the species F. mattea, F. volgiola and F. incei curtisiana (type species), and the related genus Billordia. The aim of this study was to test the taxonomic status of the Figuladra lineage by means of molecular phylogenetic and comparative morphological analyses. We use an integrative approach initially based on comparative analyses of mitochondrial phylogenetics by analysing fragments of two partial mitochondrial genes, 16S rDNA (16S) and cytochrome c oxidase subunit I (COI). We used key morphological features (shell, radular morphology and reproductive anatomy) to review the systematic relationships of Figuladra more comprehensively.
Abbreviations used
General: SEM, scanning electron microscopy; SC, spirit collection; RC, dry collection.
Institutions: AMS, Australian Museum, Sydney; QM, Queensland Museum, Brisbane.
Geography: BL, Brigalow Lands; Ck, Creek; I, Island; MEQ, mid-eastern Queensland; Mt, Mount; NP, National Park; NQ, North Queensland; NSW, New South Wales; R, River; Ra, Range; SCQ, southern-central Queensland; SEQ, south-eastern Queensland; SF, State Forest, WA, Western Australia; WT, Wet Tropics.
Molecular phylogenetics: 16S, 16S rRNA gene; BI: Bayesian Inference; COI, cytochrome c oxidase subunit I gene; ML, Maximum Likelihood.
Shell features: D, shell diameter; H, shell height; H/D, ratio of shell height to shell diameter; W, number of whorls.
Genital anatomy: BC, bursa copulatrix; DG, prostate; E, epiphallus; EF, epiphallic flagellum; EP, epiphallic pore; GD, hermaphroditic duct; GG, albumen gland; P, penis; PPL, penial pilasters; PRM, penial retractor muscle; PS, penial sheath; PV, penial verge; UT, uterus; UV, free oviduct; V, vagina; VD, vas deferens; Y, atrium.
Materials and methods
Specimens and sampling
This work is based on examinations of dry shells and ethanol preserved samples deposited in the Queensland Museum, Brisbane and the Australian Museum, Sydney. This includes material collected recently during field trips in eastern Queensland. Live specimens were transported from the field to the laboratory, euthanased by immersing in water, usually overnight, fixed in 95% ethanol and subsequently stored in 70% ethanol for use in anatomical and molecular studies. All newly collected specimens have been lodged in the Queensland Museum (Supplementary Table S1).
Molecular phylogenetics
Genomic DNA was extracted from small pieces of foot muscle using the QIAGEN DNeasy tissue kit following the Spin Column protocol. Fragments of two partial mitochondrial genes (16S and COI) were amplified by PCR using the primers 16SCS1 (Chiba 1999) and 16SBD1 (Sutcharit et al. 2007) for 16S and LCO1490 and HCO2198 (Folmer et al. 1991) for COI. Standard M13F and M13R primer sequences were appended to the COI primers to assist sequencing. PCR reactions (Supplementary Table S2) were performed under the following conditions: initial denaturation at 98°C for 2 min, followed by 35 cycles of 98°C for 30 s, 48°C for 30 s and 72°C for 30 s, with a final elongation step at 72°C for 5 min for 16S; initial denaturation at 94°C for 2 min, followed by 45 cycles of 95°C for 60 s, 50°C for 60 s and 72°C for 60 s, with a final elongation step at 68°C for 10 min for COI. The size of the PCR products was checked using UV transillumination after gel electrophoresis in a 1.5% agarose gel and TAE buffer solution. The PCR products were purified and sequenced at Macrogen Inc. (South Korea).
Sequences generated (from 34 individuals) have been deposited in GenBank under the accession numbers OL753690–OL753711 (COI) and OL839880–OL839914 (16S). Using the camaenid phylogeny of eastern Australia published by Hugall and Stanisic (2011) as a guide, several additional camaenid sequences were downloaded from GenBank and included in our analysis. Clades 1–4 from Hugall and Stanisic (2011) were defined as the hadroid clades and we have used this framework to anchor our study (Fig. 1). Accordingly, we included species shown to be more closely related to Figuladra (Clade 1 of the tree from Hugall and Stanisic 2011) and species that often occur in sympatry with Figuladra species considered to belong to a distantly related clade (Clade 4 of the tree from Hugall and Stanisic 2011). Sequences of the Western Australian camaenid Rhagada dampierana and Austrochloritis porteri from SEQ (Clade 9 from Hugall and Stanisic 2011) were used to root the tree. Details of the sequences used in the analysis can be found in Table 1.

|
The mtDNA sequences were aligned using the MAFFT (ver. 7.308, see https://mafft.cbrc.jp/alignment/software/; Katoh and Standley 2013) plug-in within Geneious (ver. 9.1.8, see https://www.geneious.com/; Kearse et al. 2012), with default settings. We used Guidance2 (ver. 2.02, see http://guidance.tau.ac.il/; Sela et al. 2015) to identify and remove unreliably aligned regions in the 16S alignment by employing default settings. COI sequences were translated into amino acid sequences to ensure that possible pseudogenes were not included. For phylogenetic analyses, 16S and COI nucleotide sequences were concatenated into one partitioned dataset. Four partitions were designated, one for each of the three codon positions of the COI and one for the 16S gene fragment.
Phylogenetic relationships were estimated by employing a Maximum Likelihood-based method of tree reconstruction and Bayesian Inference. A Maximum Likelihood (ML) phylogeny was reconstructed by using the software IQ-TREE (ver. 1.6, see http://iqtree.cibiv.univie.ac.at/; Nguyen et al. 2015). We used the integrated modellfinder under the option ‘merge’ to identify the best-fit model of sequence evolution and merge partitions with similar models before the analysis (Kalyaanamoorthy et al. 2017). Nodal support of the best ML tree was estimated by performing 1000 ultra-fast bootstrap replicates. Bayesian posterior probabilities were estimated by running a 10 000 000 generation Metropolis-coupled Markov chain Monte Carlo (four chains, one heated, sampling rate 1000 generations) with a burn-in of 100 000 as implemented by the MrBayes (ver. 2.2.4, see https://github.com/NBISweden/MrBayes/; Ronquist and Huelsenbeck 2003) plug-in in Geneious (ver. 9.1.8).
Mean genetic p-distances within and among groups were calculated using MEGA6 (ver. 10.2.6, see www.megasoftware.net; Tamura et al. 2013).
Comparative morpho-anatomy
Details of the samples used for analysis of morpho-anatomy are provided in Supplementary Table S1. Characters investigated included shell shape, size (height and diameter), whorl count and colour patterns of both shell and mantle tissue (shell secreting organ). Standard definitions for conchological characters were used (Solem and van Bruggen 1984). Shell dimensions (height, diameter, whorl count) were measured using callipers with a precision of 0.1 mm. Differences in shell size (diameter) are indicated in the diagnoses using the following terms: medium (10–20 mm), large (21–40 mm), very large (>40 mm). Whorls were counted to the nearest 1/8 whorl following Clark (2009). The microsculpture of the teleoconch was examined using a TM-1000 Tabletop Scanning Electron Microscope in topo image mode. For SEM, shells were cleaned by gently brushing in warm soapy water followed by ultrasound treatment. Samples were mounted on sticky tabs, sputter-coated with gold, and imaged under high vacuum.
Bodies were removed from the shells and dissected to extract the reproductive system that was submerged in a shallow container of 75% ethanol and pinned to a black latex base using very fine ‘Austerlitz’ entomological pins. Photographs of the reproductive system were taken using a NIKON Coolpix 4500 camera mounted on a WILD M5 stereo microscope.
Radulae and jaws were extracted from preserved specimens and cleaned by soaking in 10% KOH solution overnight followed by rinsing in water and ethanol. Specimens were mounted on aluminium stubs, coated with gold and studied under scanning electron microscopy.
High resolution images of shells and penial anatomy (260–600 MB) were obtained using a Visionary Digital BK-Plus laboratory system camera set-up located in the Queensland Museum’s Digital Imaging Unit.
Results
Molecular phylogeny
We analysed two different datasets. First, we used Hugall and Stanisic’s (2011) original sequences and re-calculated relationships among the hadroid forest snails (Clade 1) but updated the taxa following the classification of Stanisic et al. (2010). We did this because Hugall and Stanisic (2011) used many placeholder names that cannot be translated to currently accepted taxonomy without expert knowledge.
We analysed a restricted sequence dataset containing new and previously reported (GenBank) sequences of Figuladra and some more closely related taxa to resolve the relationships of Figuladra in more detail. Phylogenetic analyses were based on 25 newly produced COI and 34 new 16S sequences of Queensland camaenids, and several GenBank sequences (2 for COI and 18 for 16S). These sequences represented 33 species from 11 genera (Table 1). We found that the topology of our tree was consistent with the one of Hugall and Stanisic (Fig. 2). However, we note that many of the key nodes had posterior probability/ultrafast bootstrap values below 95 and are thus only moderately supported. According to this tree, Figuladra is most closely related to Lamprellia, Monteithosites, Marilynessa and members of a clade containing Steorra, Temporena and Bentosites all from Clade 1 as first identified by Hugall and Stanisic (2011). Billordia is nested within the Figuladra clade. The Figuladra clade contained three major lineages, Figuladra mattea, Figuladra volgiola and a lineage including three subclades with a geographical structuring – one lineage from MEQ, another from the coastal area of SEQ and a third from the montane areas of the Boyne Range, Mount Biggenden and Mount Mudlo.

|
This dataset contained sequences of 48 individuals. All COI sequences had a length of 655 bp after pruning of the primer sites, whereas the multiple 16S sequence alignment had a length of 770 bp after trimming of ends and removing ambiguously aligned sections. For the phylogenetic analysis, a data partition was applied that allowed parameters for each codon position of the COI fragment and the 16S fragment to be modelled independently. The model test implemented in IQ-Tree identified the following models of sequence evolution as the best-fit models for the different partitions by means of the Bayesian information criterion: TIM2 + F + I + G4 for 16S, TN + F + I + G4 for the 1st and 2nd and TPM3 + F + I for 3rd codon positions in COI. These models were applied in the partitioned Maximum Likelihood analysis. The concatenated alignment had 1425 columns, of which 546 were parsimony-informative, 136 were singleton sites and 743 were constant sites.
The best Maximum Likelihood tree (Fig. 2a) was generally consistent with the phylogeny produced by Hugall and Stanisic (2011) placing Figuladra within ‘Clade 1’ and separate from the Sphaerospira species in Clade 4. The bootstrap consensus tree of the ML analysis confirmed all species of Figuladra formed a clade that also contained Billordia nicoletteae. Nodal support was high for three major clades within Figuladra, however support for higher-level relationships was moderate. The Bayesian consensus phylogram (Fig. 2b) exhibited a largely identical topology with the exception of the placement of Billordia and the Sphaerospira clade that was shown to be unresolved with respect to the seven species included in the tree.
We recognise three major clades within the Figuladra lineage (Fig. 2). The first major clade, Clade A, comprised several Figuladra species. The clade revealed a bifurcation between two sub-clades that contained the MEQ (F. incei incei, F. challisi, F. barneyae and Camaenid BL47) and SEQ species respectively. A third sub-clade contained two species occupying upland habitats, F. narelleae (Bouldercombe Gorge, Boyne Range and related drainage lines) and F. bayensis (Mount Mudlo and Mount Biggenden). The second major clade, Clade B, corresponded to F. volgiola from MEQ and SCQ. Branches within this clade corresponded to geographically isolated sub-populations of F. volgiola in the Peak Range, Broadsound Range and Expedition Range respectively. Clade C corresponded to Figuladra mattea, a species with a broad SEQ and SCQ distribution.
Genetic distances
p-distances within clades ranged from 0.1 to 32.6% for 16S and from 0.4 to 26.1% for COI (Supplementary Table S3). The interspecific distances within F. mattea were below 5.4% across the entire range in SEQ and SCQ for both genes. F. volgiola showed low divergence within 16S (0.25–0.43%) and higher divergence within COI (7.8%).
Intraspecific p-distances ranged from 1.3 to 19.8% in Clade A, from 0.2 to 7.8% in Clade B and from 0.1 to 3.6% in Clade C (Supplementary Table S3). These intraspecific distances were considerably lower than the mean interclade p-distances ranging from 10.0 to 23.6%. Although the p-distance within species was low, the p-distance over sequence pairs among groups suggested by the ML and BL tree topology averaged 13% for the COI gene and 19% for the 16S gene (Table 2). Differences among genera ranged between 10% (Clade C and Billordia) and 23.6% (Clade A and Billordia).

|
Morphological variation
Genital morphology
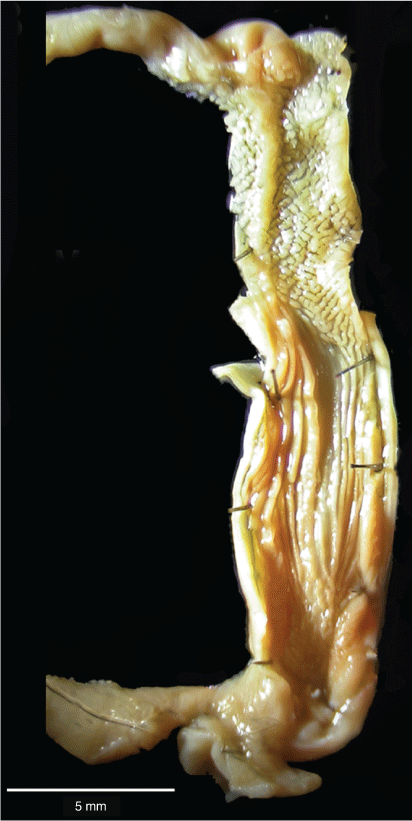
A total of 39 specimens of Figuladra mattea (Clade B) and F. volgiola (Clade C) from SEQ, MEQ and SCQ and 6 specimens of Billordia species were dissected. Although 202 specimens of Figuladra (Clade A) have been dissected for comparison, only 3 representative specimens of Figuladra from this set are featured herein. Other specimens from Clade 1 were dissected for genital anatomical comparison (see Supplementary Table S1). There was a clear distinction between the genital anatomies of Figuladra mattea, F. volgiola and all other species of Figuladra. Billordia Stanisic, 2010, Denhamiana Stanisic, 2016 and Bentosites Iredale, 1933 were also noticeably dissimilar in gross anatomical structure (Fig. 3). A shorter, thicker epiphallus and bands of muscular tissue at the epiphallic-penial junction are characteristics of all species of Clade A (=Figuladra sensu stricto; Fig. 3b). This muscular tissue connecting the apical portion of the penis with the epiphallus is lacking in both F. mattea, F. volgiola and Billordia (Fig. 4). Furthermore, the long, twisted and looped epiphallus of F. volgiola (Clade B), the long, coiled descending arm of the epiphallus of F. mattea (Clade C) and the knotted epiphallus of Billordia are distinguishing characters from all other Figuladra species (Clade A; Fig. 4). The ratio between the length of the penis and the length of the vagina is much lower in F. mattea (mean 1.6) and F. volgiola (mean 1.7) than in Figuladra s.s. (mean 2.4) (Supplementary Table S4).

|

|
The penial interior of F. volgiola (Clade B) was characterised by one fleshy longitudinal pilaster running the full length of the penis descending into the atrium, and an apical region with basal ridges forming thin, chevron-like thickenings for one-third of the penial length (Fig. 5). This pattern was consistent across F. volgiola specimens from different localities including the Peak, Expedition, Boomer and Broadsound Ranges. All had chevron pilasters in the apical region and the penial-length longitudinal plaster (Supplementary Fig. S1).

|
The penial interior of F. mattea (Clade C) is characterised by an upper penial chamber with numerous irregular pustules giving rise to a chevron-like pattern of pustules; a single longitudinal pilaster in the upper penial chamber; and several regular longitudinal thickenings that descend into the atrium (Fig. 6). This pattern of pilasters in the penial chamber architecture of all F. mattea specimens examined from within the broad range in SEQ and SCQ was consistent, with only minimal variations in the number of pilasters in the atrium (Supplementary Fig. S2).
|
Billordia bridgettae has a double longitudinal thickening descending into the atrium and an apical region with basal ridges forming thin, chevron-like thickenings for more than one-half of the penial length (Fig. 7a). Typical Figuladra species (Clade A) have a single or double longitudinal thickening in the apical region that also has rugose, crenelated or spade-like pilasters in various combinations generally for one-third of the penial length and bifurcating before descending into the atrium (Fig. 7b–d).

|
Shell morphology
In total, 635 shells of F. mattea and F. volgiola, and 34 shells of Billordia nicoletteae and B. bridgettae were examined and measured (Supplementary Table S1). Variation in shape, colour patterns and shell morphometry (shell parameters H, D, H/D, W) was minimal within F. mattea and F. volgiola. Shells of F. volgiola (Clade B) and F. mattea (Clade C) were similar in size and whorl count (Fig. 8, Table 3) compared to other Figuladra species. F. volgiola differed from all other species by having a fully closed umbilicus (Fig. 9). With the exception of F. volgiola with a closed umbilicus, the shells of all Figuladra species exhibit only minor differences among species. Both Billordia species were smaller and flatter than any Figuladra species in Clade A, B and C and have a significantly lower H/D ratio and whorl count. The sub-discoidal shell shape with a more open umbilicus in Billordia species differed from the depressedly globose to trochoidally globose shell shape of Clades A, B and C (see Fig. 12e). We observed variations in lip colour and suffusion: F. mattea, F. volgiola, B. nicoletteae and B. bridgettae had white lips with no suffusion behind the lip unlike all other Figuladra species that d varying levels of brown suffusion behind the lip. All Figuladra species exhibited a variety of colour forms ranging from solid dark to banded. One species, F. pallida, also exhibited a ‘blond’ form. Figuladra volgiola was always banded (Fig. 9). Shells of F. mattea were generally banded but the species also encompassed a heavily banded, darker form in Dulacca, SCQ and a ‘blond’ form in Ipswich, SEQ (Fig. 10). Neither F. mattea, F. volgiola nor Billordia species had solid dark, unbanded shells.

|

|

|

|
SEM investigation of teleoconch microsculpture revealed similarities among all specimens. Shell sculpture consisted of fine, irregularly disposed microradical, periostrical threads. The exception was Billordia with B. bridgettae and B. nicollettae, both having a pustulose teleoconch microsculpture (Fig. 11).

|
Mantle colour
Differences were observed in the colour of the mantle (Fig. 12a–d, f). The orange mantle of Figuladra volgiola was similar to the mantle colour in Bentosites, but differed from that of Figuladra species in Clade A, that had a strong pink mantle colour (Stanisic et al. 2010). The mantle of F. mattea was consistently orange–pink across the species’ entire range. This feature distinguished these groups from the Sphaerospira clade that had a black mantle and other genera belonging to Clade 1 (Table 4).

|
|
Jaw and radula
No significant differences in jaw and radular morphology were detected among any of the examined species (Fig. 13). The radulae were typically camaenid with a unicuspid central tooth, laterals gradually becoming bicuspid and multicuspid marginals (Solem 1973).

|
Discussion
Phylogenetic relationships
Current knowledge of the phylogenetic relationships of the eastern Australian Camaenidae essentially rests on the phylogenetic study of Hugall and Stanisic (2011). This study presented a mitochondrial phylogeny based on the analysis of three partial mitochondrial genes (12S, 16S, COI), in which species now placed in Figuladra were shown as members of Clade 1 (labelled as the mattea, volgiola, BL4 and appendiculata lineages).
Our new phylogeny included samples from the Hugall and Stanisic’s original tree. This phylogeny is consistent with Hugall and Stanisic (2011) in the placement of Bentosites, Steorra, Monteithosites, Marilynessa, Figuladra, Billordia and Lamprellia in Clade 1 and the grouping of Sphaerospira, Austrocamaena and Gnarosophia, all Queensland hadroid snails, in a separate clade (Clade 4). However, the placement of Billordia nicoletteae within the Figuladra clade differed from the Hugall and Stanisic’s (2011) phylogeny. The implication is that Figuladra as circumscribed by Stanisic et al. (2010) is not monophyletic. To maintain monophyletic taxa, the following options exist: synonymising Billordia with Figuladra, with Billordia as the older name having priority, or alternatively, describing new monotypic genera for F. mattea and F. volgiola thus maintaining Billordia and Figuladra as distinct genera. As the placement of Billordia nicolettaea had very low support within the maximum likelihood analysis, and was unresolved by the Bayesian analysis, a taxonomic decision as to how best to delineate genera requires an evaluation of comparative morpho-anatomy in addition to the phylogenetic inference.
Genetic distances
Davison et al. (2009) found a large overlap between intra- and interspecific divergence in mitochondrial DNA in stylommatophoran land snails with intraspecific distances sometimes extreme, reaching 20–30%. In Asian camaenids, Kameda et al. (2007) reported uncorrected pairwise sequence divergences of 0–7.3% within and 0.3–20.5% among traditionally circumscribed species. In our study, the observed p-distances among clades A, B and C (Table 2) are comparable with the genetic divergence observed among genera in other studies of Australian camaenids (Köhler and Johnson 2012; Taylor et al. 2015). Similar genetic distances are observed among genera of other taxa. Oba et al. (2015) in their study of 275 click beetles, stated an average p-distance among genera using the COI gene as 11.6%. In the study of sea snails (Neogastropoda), Zou et al. (2011) established that the mean pairwise divergence among specimens of different sea snail genera using both the 16S and COI genes was 18.4% (range 6.3–24.8%). However, although no such benchmark is available for delimitation of land snail genera, sea snails do show very similar generic p-distances to those observed here within the hadroid camaenids.
Morphological variation
Using COI and 16S genes, Köhler and Johnson (2012) showed an association between a genetic distance greater than 6% and the presence of distinct genital morphology in snails. With all comparative p-distances greater than 10% for both 16S and COI genes among F. mattea, F. volgiola, Billordia and Figuladra s.s. lineages, this substantial mitochondrial differentiation is not unexpectedly complemented by a significant difference in penial anatomy (Fig. 3, Table 2). Solem (1997) proposed that the gross genital anatomy of adult camaenids is informative for delimitation of genera and as such, this anatomical feature is an important aspect of this study. The separation of F. mattea and F. volgiola from Billordia and Figuladra s.s. is reinforced by differences in gross genital features. The genital anatomy of Billordia has a knotted epiphallus and lacks the uncoiled epiphallus of Figuladra in Clade A, and the coiled, looped and twisted epiphalluses of F. mattea (Clade B) and F. volgiola (Clade C) respectively. The muscular tissue at the penial–epiphallic junction that characterises Figuladra species is absent in Billordia, F. mattea and F. volgiola. The penial–vagina ratio is significantly different among Figuladra s.s., F. mattea and F. volgiola.
Radulae and jaws have previously been shown to be of little use in generic or species delimitation in camaenids (Solem 1984; Köhler 2010b) and this study reveals similar results. However, comparisons of the mantle tissue that generally extends to the foot and body in hadroid snails, shows a difference from the pink mantle and pinkish grey animal of Figuladra s.s. to pinkish-orange to orange forms in both F. mattea and F. volgiola respectively. This difference in colour pattern is consistent with the differences among other hadroid genera (see Table 4).
Shell characters have been shown to typically offer little value to the delimitation of these genera. All the hadroid species in Clade 1 and 4 display banding patterns in various combinations. However, the lack of umbilicus in Figuladra volgiola is a shell character separating this species from Billordia species and any species in Figuladra s.s. Unlike Figuladra species that have sub-globose to globose shells, Billordia is typified by a subdiscoidal shell with a low spire (Fig. 12e) and is distinguished from Figuladra species by the pustulose microsculpture on the teleoconch. Bentosites, Monteithosites, Denhamiana and Figuladra species all have a relatively smooth teleoconch with a microsculpture of fine periostracal microradial threads.
New genera
We have taken a multidisciplinary approach and have considered not only the phylogenetic evidence and genetic distances but also morphological evidence. As most of the key nodes from the molecular phylogenetic analysis had posterior probability/ultrafast bootstrap values below 95, our proposed classification rests predominantly on our morphological analysis. We note that the molecular phylogenetic analysis is congruent with our proposed classification. The use of additional markers may improve support values in future studies. From our assessment of the observable differences, we consider the best option to be to not to subsume Clades A to C within the genus Billordia given the extent of anatomical and morphological differentiation but to recognise the mattea and volgiola lineages as distinct genera as these represent evolutionarily distinct lineages that are characterised by unique anatomical features. Accordingly, we support Clades A, B, C and Billordia as separate lineages within the Queensland hadroid group. Two new monotypic genera, Euryladra, gen. nov. (Clade B, the mattea lineage) and Brigaladra gen. nov. (Clade C, the volgiola lineage), are therefore diagnosed by the combination of shell and penial morphology supported by phylogenetic data. Updated diagnoses of Figuladra and Billordia are therefore included.
These phylogenetic reconstructions indicate that Euryladra and Brigaladra are both monophyletic and monotypic. Although highly unusual for single camaenid species to persist across such a vast region as SEQ, SCQ and MEQ, there is no evidence to support multiple species within these two lineages. Monotypic genera in land snails are not unknown with 50 monotypic camaenid genera in Australia including 23 from Queensland (Köhler 2010a, 2010b; Stanisic et al. 2010, 2018; Stanisic 2016). This scenario is reflective of the complex history of land snail evolution in eastern Australia whereby mesic communities have undergone a long history of climate-induced fragmentation and isolation.
Biogeography: diversification across habitats in Clade 1 camaenid land snails
Land snails are dependent on mesic conditions for activity and many groups have retained a preference for rainforest or other environments that are at least seasonally mesic (Solem and van Bruggen 1984; Stanisic 1994; Stanisic et al. 2007). Land snails also show an excellent capacity to persist in relatively localised areas, especially in association with limestone outcrops or other rocky ranges that provide microrefugia from key potential threats such as fire and aridity (Couper and Hoskin 2008). Accordingly, land snails are potentially an excellent group for understanding biotic patterns in response to broadscale climatic change, such as the ongoing and increasing aridification of the Australian continent (Byrne et al. 2008, 2011).
Unlike some other lineages of eastern Australian camaenids that have distributions centred on wet evergreen rainforests (for example: Hugall et al. 2003), Clade 1 land snails are strongly associated with relatively drier closed forest (Stanisic et al. 2010; Hugall 2011), referred to in Australia as vine-thickets and scrub, but on a global scale consistent with what is referred to as dry seasonal tropical forests or subtropical dry forest (Webb and Tracey 1981; Neldner et al. 2019). Six of the nine genera recognised in Clade 1 have distributions centred on these vine-thicket habitats and three have diversified into other habitats in addition to vine thicket (see Table 5). This diversity in dry closed forest habitats suggests that this clade adapted to these habitats early in the evolutionary history and has subsequently undergone ecological and evolutionary diversification. One putative driver of this diversification may have been the spread of open grassy and fire-affected woodland or savannah habitats that are generally depauperate in land snails (Stanisic 1994; Stanisic and Ponder 2004). Reflecting this trend, within Clade 1 only two lineages (Lamprellia and the newly recognised Euryladra) regularly occur in more open woodland habitats, despite these habitats covering vast areas of eastern Australia.

|
Within this broader evolutionary and paleo-environmental context the four focal genera for study show contrasting contemporary distributions that likely reflect differences in habitat and ecology. Figuladra s.s. is the only clade among these four genera that is restricted to an area east of the Great Dividing Range. This genus is distributed in near-coastal dry vine thickets with nine species now distributed from north of the Mary River, Gympie, SEQ to slightly south of the O’Connell River, south of Proserpine, MEQ (Stanisic et al. 2010). Records from both QM and AM, and our observations from field work show that species appear to be primarily vine thicket dwellers, but can occur in gallery forests along adjacent or emanating drainage lines.
Brigaladra also appears to be an obligate vine thicket occupant that is represented by a single species, B. volgiola. Although this species occurs west of the Great Dividing Range, records have only been made in ranges (Peak, Broadsound, Boomer and Expedition Ranges) living in vine thickets on rocky scree. Within these ranges B. volgiola is closely associated with vine thickets, although the species also occasionally occurs in woodland along emanating drainage lines, for example in Charlevue Creek, Expedition Range, SCQ; in Roper Creek, Peak Range, MEQ; and in Apis Creek, Boomer Range, MEQ (Fig. 14, 15a, b). These ranges are located on either side of the Isaac-Comet Downs Bioprovince that has been largely cleared for farming. The Isaac-Comet Downs, a lowland region of brigalow and open savannah woodland, is home to camaenids such as Pallidelix potteri Stanisic, 2018. Museum records (AM and QM) show that no B. volgiola were found in this lowland area in early collections made before much of the subsequent land clearing. The apparent disjunct distribution of the B. volgiola populations in scattered vine thickets of rock-strewn ranges appears to mirror the fragmented contemporary distribution of these vine thickets.

|

|
Like Brigaladra, Billordia is also associated with vine thickets in the drier ranges (Peak, Broadsound) west of the Great Dividing Range on either side of the Isaac-Comet Downs. The two species, B. bridgettae and B. nicoletteae live among vine thicket on rocky scree and, unlike Brigaladra, have not been found in vine thickets on other substrates. The former has been found sympatrically with Brigaladra in the Peak Range, MEQ, but museum records show that each occur within different vine thickets in the same locality. B. nicoletteae occurs in the vicinity of Lotus Creek at the southern end of the Connors Range, MEQ. Billordia species have a flattened shell and correlated open umbilicus (see Fig. 12e). These features are hypothesised to be adaptations to living among rocky scree where more globose shell shapes impede access to movement in these refugia (Criscione and Köhler 2013; Hamilton 2021). The pustulose teleoconch and sharply reflected lip represent a marked divergence from the shells of the other members of Clade 1. Given the very limited wet material available in QM, molecular data from both Billordia species would enhance future comparative study.
Euryladra is represented by a single species, E. mattea, that has a very wide distribution in the eucalypt woodland east and west of the Great Dividing Range (Fig. 15c, d). Occasionally, but relatively infrequently, E. mattea occupies semi-deciduous vine thicket. Euryladra appears to be one of only two lineages in Clade 1 that can persist in the evolving drier eucalypt-dominated landscape. The physiological, behavioural or morphological adaptations that have enabled this taxon to occur across a vast area outside of vine thickets remain unknown, but could be an interesting area for further investigation.
Systematics
Class GASTROPODA
Order EUPULMONATA
Superfamily HELICOIDEA
Family CAMAENIDAE
Brigaladra L. Stanisic, gen. nov.
Etymology
Named for the brigalow lands and a contraction of Figuladra.
Diagnosis
Shell large, sutures weakly impressed; protoconch with weak, crowded radial ridges, teleoconch smooth, microsculpture of fine microradial periostracal threads; lip reflected, umbilicus closed. Shell yellowish with white suffusion behind lip, white with brown banding continuing onto the lip; brown circum-umbilical patch. Animal orange–grey with an orange mantle. Reproductive system with long epiphallus looped and twisted bound in connective tissue; penis approximately the same length as the vagina.
Range
Boomer Range and Broadsound Range, SEQ; Expedition Range and Peak Range, SCQ.
Remarks
Brigaladra is distinguished from Billordia, Figuladra and Euryladra by having a fully closed umbilicus and a long twisted epiphallus. Figuladra has a shorter, thicker epiphallus and muscular tissue at the epiphallic–penial junction. Billordia has a knotted epiphallus and Euryladra has a coiled epiphallus encased in connective tissue at the epiphallic–penial junction.
Brigaladra volgiola (Iredale, 1933), comb. nov.
(Fig. 3a, 4b, 5, 8, 10, 11b, 13d–f, 14, 15a,b, Tables 1–4)
Common name
Boomer Range Banded Snail.
Material examined
Lectotype. NHMUK 1880.12.11.14 [N end of Expedition Range. W of Rockhampton. Height of shell 22.64 mm, diameter 30.62 mm, H/D 0.739, whorls 5.875].
Paralectotypes. AMSC107629 [6RC, N end Expedition Range, W of Rockhampton, 23°42′S, 148°59′E, 1872, J.C. Cox collection]; AMSC107630 [6RC, N end Expedition Range, W of Rockhampton, 23°42′S, 148°59′E, 1872, J.C. Cox collection].
Other material. (All SCQ and MEQ) Boomer Range. QMMO46286, 9SC/RC; QMMO23495, 3SC/RC; QMMO44110, 13SC; QMMO47961, 4RC; Broadsound Range, MEQ. AMSC107641, 4RC; AMSC386835, 1RC, QMMO45967, 77SC/RC; QMMO45973, 18SC/RC; QMMO46282, 27SCRC; Expedition Range, SCQ: AMSC5579, 1RC; AMSC471624, 2RC; QMMO46300, 1 RC; QMMO87091, 6SC; QMMO87092, 6SC; QMMO87093, 12SC; QMMO87094, 12SC; QMMO87095, 12SC; QMMO87096, 11SC; Peak Range. QMMO78066, 85SC/RC; QMMO78080, 3RC, QMMO78082, 6RC, QMMO78113, 12SC/RC; QMMO78117, 3SC/RC; QMMO78118, 22SC; QMMO78123, 10SC; QMMO78133, 18 RC; QMMO78134, 25SC/RC; QMMO78145, 1RC; QMMO78148, 2RC; QMMO78155, 4 RC; QMMO78170, 10SC/RC.
Taxonomic note
When Cox (1872) described Helix andersoni, a holotype was not explicitly nominated and mention was made that the shell was one of ‘many fine specimens that Mr Anderson had collected’ (p. 654). This demonstrates the existence of a type series. Smith (1992) labelled a specimen 1880.12.11.4 as a holotype and listed the two paratype lots but did not acknowledge the set as part of a type series. The NHMUK has the collection log showing 14 ‘species’ (1800.12.11.5–21) presented to the museum by Dr Cox. This set includes the single specimen of H. andersoni considered to be part of this series. Two paratype lots, each with 6 specimens labelled as being from Cox’s collection, are considered to be part of this type series and are located in the Australian Museum. Consultation with the NHMUK proved that Smith’s voucher number was printed incorrectly (J. Ablett, pers.obs.). To correct this error, the specimen with registration number NHMUK1880.12.11.14 is redesignated as a lectotype, and AMSC107629 and AMSC107630 are redesignated as paralectotypes.
Diagnosis
As for genus.
Description
Shell large, diameter 26.91–34.13 mm (mean 30.33 mm), height 19.64–26.96 mm (mean 23.25 mm), H/D ratio 0.716–0.841 (mean 0.766), whorls 5.750–6.250 (mean 6.000); subglobose with a moderately elevated spire; whorls rounded, sutures impressed; protoconch with weak crowded radial ridges, teleoconch smooth, microsculpture of fine periostracal microradial threads; shell yellow with numerous narrow brown spiral bands; lip reflected, white with brown banding continuing onto the lip and clearly visible in the aperture; umbilicus closed with brown circum-umbilical patch; animal orange–grey with an orange mantle. Based on 211 measured adult specimens (see Supplementary Table S1).
Genitalia. Penis long and tapered with slightly expanded apical bulb, penial sheath present; penial chamber with one fleshy longitudinal pilaster descending into the atrium, apical region with basal ridges forming thin chevron-like thickenings for one-third of the penial length; penial retractor muscle inserted at the junction of the two arms of the epiphallus; epiphallus with short, thin muscular ascending arm, descending arm relatively longer, more expanded; epiphallus entering penis through a simple pore; vas deferens thin and attached to penial sheath with connective tissue; short, vestigial epiphallic flagellum present at the epiphallus–vas deferens junction, tightly bound to the vas deferens; vagina short but more than half the length of the penis, PVR 1.052–2.051 (mean 1.709); free oviduct shorter than vagina; bursa copulatrix situated at the base of the albumen gland, with a slender stalk; atrium simple. Prostate, uterus and hermaphroditic duct without unusual features. Based on 19 dissected adult specimens (see Supplementary Tables S1, S4).
Distribution and habitat
Boomer Range, Broadsound Range, SEQ; Expedition Range, Peak Range, SCQ; semi-evergreen vine thicket to adjoining dry woodland, living under rocks and inside logs.
Euryladra L. Stanisic, gen. nov.
Etymology
From the New Latin prefix eurys, which means widespread and a contraction of Figuladra.
Diagnosis
Shell large, sutures weakly impressed; protoconch with weak crowded radial ridges, teleoconch smooth, microsculpture of extremely fine periostracal microradial threads; lip expanded and moderately flared; umbilicus open. Shell yellowish with white suffusion behind lip and dark brown circum-umbilical patch; animal pinkish-orange–grey with a pinkish-orange mantle. Reproductive system with long, coiled epiphallus entwined in connective tissue at the epiphallic-penial junction; penis approximately the same length as the vagina.
Range
From Tweed River, NE NSW to Rockhampton, SEQ and inland as far west as Carnarvon NP, SCQ.
Remarks
Euryladra differs from Brigaladra, Billordia and Figuladra by genital characteristics such as a strongly coiled epiphallus and the configuration of the penial pilasters. Euryladra lacks the muscular tissue connecting the penis to the epiphallus that is present in Figuladra. The single longitudinal pilaster in the upper penial chamber of Euryladra differs from the longitudinal thickenings in Brigaladra, Billordia and Figuladra.
Euryladra mattea (Iredale, 1933), comb. nov.
(Fig. 3b, 6, 8, 10, 11a, 12c,d, 13a–c, 14, 15c,d, 16, Tables 1–4)
Common name
Pale Banded Snail.
Material examined
Holotype. AMSC100642 [Rockhampton, SEQ, 23°22′S, 152° 32′E, coll. H. Bernhard, ii. 1932. Diameter 2.04 mm, height 2.92 mm, H/D 0.699, number of whorls 6].
Other Material. (All SEQ and SCQ) Ambrose limestone quarry: QMMO8158; Archookoora Forest Stn, via Kingaroy: QMMO4262; Auburn R NP, via Mundubbera: QMMO64432; Ban Springs, QMMO21151; Banana Ra, QMMO50489, QMMO50640, QMMO50657, QMMO50658; Baralaba, MO23499; Bauhinia Downs Stn: QMMO35820, QMMO5858; Beaudesert: QMMO68572; Big Woody I: QMMO65483; Biggenden: QMMO47478, QMMO47483, AMSC136406, AMSC434601; Biloela: QMMO76378, QMMO76379, QMMO76380, QMMO80514, AMSC158652; Blackwater: QMMO67777, AMSC444386; Boomer Ra: QMMO47963, QMMO50487, QMMO50488; Bouldercombe: QMMO29869, QMMO31296: Boyne Range: QMMO80364; Bracewell: QMMO72175, QMMO79346, QMMO8135; Bribie Is: QMMO32359, QMMO9822; Brisbane: QMMO75436, QMMO76682, QMMO77887, AMSC158527, AMSC434606, SAMAD14513, MCZ315889; Broadsound Ra: QMMO45979, QMMO47847; Builyan Railway Stn: QMMO20317; Bundaberg: QMMO6775, AMSC92299; Burnett R: QMMO31420, AMSC136724; Burrum Heads: QMMO52122, AMSC467373; Byfield: AMSC471706; Calliope: QMMO72201, QMMO72203, QMMO72204, QMMO72285, QMMO72316, QMMO72331, AMSC444295; Cania Gorge: QMMO23133, QMMO35092; Cannindah: MO24559; Carnarvon NP: QMMO56643; Clifton Ra: QMMO22384, QMMO22994; Comet: QMMO44100, AMSC400913; Cooloola SF: QMMO5106, QMMO5254, QMMO5857; Coomba Falls: QMMO5002, QMMO5003; Coringa: QMMO24180; Cracow: QMMO79549, QMMO79555, QMMO79560; Curtis Island: QMMO4222, QMMO4267, QMMO4907; Dallarnil: QMMO35093; Dan Scrub: QMMO80301; Dawes Ra: QMMO22640, AMSC136769; Dawson Ra: QMMO44136, AMSC137865; Dingo: QMMO46309, QMMO55291; Dulacca: QMMO86812; Dululu: QMMO45532; Eidsvold: QMMO49081, QMMO55229, AMSC31654, AMSC80633; Emerald: QMMO57096; Emu Park: QMMO6801; Esk: QMMO6783, QMMO77842; Gayndah: QMMO76041, QMMO76062, QMMO76125, AMSC137799; Gin: QMMO38277, AMSC471651; Gladstone: QMMO72181, QMMO72215, QMMO72255, AMSC444319; Glastonbury SF, QMMO21591; Gogango Ra: QMMO49020, QMMO60906; Goodnight Scrub: QMMO21800, QMMO23439; Goomeri: QMMO51966, QMMO52024, AMSC434603; Goondiwindi: QMMO76043, QMMO76067, QMMO76068; Granite Ck: QMMO43549; Gumdale: QMMO62313; Gympie: QMMO51937, QMMO55771; Henickes Hill: QMMO86916; Horrigan Ck: MO23558; Injune: QMMO79093, QMMO86640, QMMO86643, AMSC478030; Ipswich: QMMO55362, QMMO62694, QMMO6774, QMMO77877, AMSC113806, AMSC444317; Isla Gorge NP: QMMO73099, QMMO76050, QMMO86908; Kalpowar SF: QMMO21069; Keppel Bay: AMSC434528; Kilcoy: QMMO70157; Kilkivan: QMMO86935; Kroombit Tops: QMMO80556, QMMO80564, QMMO80592, AMSC434529, AMSC468221; Laidley: QMMO77848; Lake Weyba: QMMO6788; Lawgi Hall: QMMO80578, QMMO80585; Limestone Ck: QMMO5114; Many Peaks: QMMO66635, QMMO67031, QMMO6782; Marlborough: QMMO34413, QMMO46280; Maryborough: QMMO4879, QMMO6778, QMMO6787, AMSC136495, AMSC444349, AMSC444364; Miriam Vale: QMMO64065; Monto: QMMO79759, QMMO79779, QMMO79827, AMSC101254, AMSC434599; Montville: QMMO6784, AMSC137863; Morganville: DM13834941; Morinish: QMMO5123; Moura: QMMO76124, QMMO76400, QMMO76402, QMMO76681, QMMO76781, AMSC79101; Mt Aldis: QMMO79420; Mt Gayndah: MO20171; Mt Kelly: MO24547; Mt Larcom: QMMO72178, QMMO72212, QMMO79341, QMMO79348, QMMO80406, AMSC444335; Mt Misery: QMMO26439, QMMO74666; Mt Morgan: QMMO29770, QMMO21498, MO23484, QMMO4618, QMMO61898, AMSC157846, AMSC444318, AMSC467372; Mt Ox: QMMO79356; Mt Perry: QMMO86814, QMMO86836; Mt Scoria: QMMO80448, QMMO80461; Mt Tamborine: QMMO3967; Mt Walsh NP: QMMO66840, QMMO71480; Mt Woowoonga: QMMO22440, QMMO86675, AMSC136717; Mudlo Gap: QMMO22990, QMMO39435; Mulgildie Plateau: QMMO70171; Mundubbera: QMMO75425, QMMO76436; Murgon: QMMO22988; N Stradbroke I: QMMO66654; Nambour: QMMO56696; Nanango: QMMO70503; Noosa: QMMO62242; Noosa, QMMO64296; Noosa, QMMO71569; Nuga Nuga NP: QMMO79448, QMMO85015; Oakey Ck: QMMO35104; Palmgrove NP: QMMO79416; Pialba: AMSC79125; Planet Downs: QMMO35829, QMMO44123; Port Curtis: AMSC17593, AMSC79083; Raglan: QMMO21483, QMMO72172, QMMO7294; Redcliffe: QMMO37947; Ridgelands: QMMO5130, AMSC444329; Rockhampton: QMMO15091, QMMO77035, QMMO78027, QMMO78061, QMMO78090, QMMO78100, AMSC158986, AMSC434605, AMSC444308; Somerset Dam: QMMO62243; Springsure: QMMO79964, QMMO85045; St Helena I:, QMMO67338, QMMO69417, AMSC172702; Targinnie: QMMO72332, QMMO86799; Taroom: QMMO79553, QMMO79569, QMMO86895; Thangool: MO28447; Theodore: QMMO60849, QMMO60851, QMMO64382, AMSC444457; Toogoom: QMMO39448; Warwick:, QMMO64531; Weyba NP: QMMO47501; Woodgate NP: QMMO63797; Woody I: QMMO5856; Woolooga: QMMO71212, QMMO74836; Wowan: QMMO54777, QMMO60709, AMSC471659, AMSC75111. (Other records are available from the Australian and Queensland Museums).
Diagnosis
As for genus.
Description
Shell large, diameter of shell 18.50–27.47 mm (mean 22.68 mm), height 25.28–34.48 mm (mean 30.23 mm), H/D 0.683–0.820 (mean 0.749); evenly coiled, depressedly globose with a low domed spire; whorls rounded 5.750–6.750 (mean 6.00); sutures weakly impressed; protoconch with weak crowded radial ridges, teleoconch smooth, microsculpture of extremely fine periostracal microradial threads; shell yellowish with prominent broad subsutural and peripheral bands and numerous narrower dark brown spiral lines and bands; dark brown circum-umbilical patch; lip white, expanded and moderately flared, reflected over umbilicus and partially occluding this; umbilicus narrowly open to almost closed, excavate. Animal pinkish-orange–grey with pinkish-orange mantle. Based on 434 measured adult specimens (see Supplementary Table S1).
Genitalia. Penis with thin sheath, long, cylindrical; walls of upper penial chamber with numerous irregular pustules giving rise to a chevron-like pattern of pustules; a single longitudinal pilaster in the upper penial chamber and several regular longitudinal thickenings that descend into the atrium. Epiphallus with a muscular ascending arm partially coiled and entwined in connective tissue at the epiphallic–penial junction, when unravelled, 2–3 times the length of a thinner descending arm; epiphallus entering penis through a simple pore. Penial retractor muscle inserted at the junction of two arms of the epiphallus. Vas deferens attached to penial sheath with connective tissue; vestigial epiphallic flagellum situated at epiphallus–vas deferens junction, tightly bound to vas deferens; vagina short almost the same length as the penis, PVR 1.293–1.958 (mean 1.616); free oviduct much shorter than vagina, swollen at the junction to the vagina; bursa copulatrix with a slender stalk and clavate head located at the base of the albumen gland. Ovotestis and albumen gland, hermaphroditic duct typical. Based on 20 dissected adult specimens (see Supplementary Tables S1, S4).
Distribution and habitat
From Tweed River, NE NSW to Rockhampton, SEQ and inland as far west as Carnarvon NP, SCQ; open woodland and dry vine thicket living under rocks and logs.
Remarks
Euryladra mattea exhibits a range of banding patterns and colour suffusions. Specimens from Taroom, Dulacca and Planet Downs, SCQ have a darker shell than most E. mattea from the extensive range. A very rare ‘blonde’ form with no banding occurs at Bergins Hill near Ipswich, SEQ. All forms however, have a pale base. Flattening of the shell in some forms also results in a wider umbilicus (Fig. 10). In the past, E. mattea has been confused with Figuladra challisi (Cox 1873). E. mattea species are only found south of Rockhampton whereas F. challisi is a species described from Keswick I, MEQ. The true F. challisi lacks the excavated umbilical area of E. mattea and has a much more solid, porcelain-like shell (Stanisic et al. 2010; Stanisic and Stanisic 2020).

|
Figuladra Köhler & Bouchet, 2020
Diagnosis
Shell large to very large (diameter 20–41 mm, height 20–34 mm), loosely to tightly coiled; depressedly globose to trochoidally globose with a dome-shaped spire; whorls rounded and often laterally compressed with weakly impressed sutures; protoconch features weak crowded radial ridges, teleoconch has a smooth microsculpture of extremely fine periostracal microradial threads; lip is reflected, white with a dark brown suffusion behind this and at the top of the columella; umbilicus narrowly open; yellow to dark brown circum-umbilical patch; shell colour variable even within species from dark brown with few to numerous brown spiral bands to monochrome brown or yellow; animal dark grey with a pinkish tinge and a pinkish-red mantle. Reproductive system with a sheath of muscular tissue connecting the epiphallus to the top of the penis; vas deferens thin and attached to penial sheath with connective tissue; short, thin epiphallic flagellum is present, situated near the epiphallus–vas deferens junction, tightly bound to the vas deferens; vagina short, generally less than half the length of the penis; atrium is simple; free oviduct is shorter than vagina; bursa copulatrix simple with a slender stalk, situated at the base of the albumen gland; prostate, uterus and hermaphroditic duct without unusual features.
Range
Eastern Queensland from Maryborough to St Helen’s Beach, on off-lying islands from the South Cumberland Island group to Fraser Island, and west to the Great Dividing Range.
Remarks
Among eastern Australian camaenids, Figuladra is distinguished by the almost smooth microsculpture and pinkish mantle. Genital anatomy separates Figuladra from other hadroid snail genera by having a band of muscular tissue connecting the penis and epiphallus. This feature is not present in any other hadroid genus. Figuladra is also separated from Euryladra and Brigaladra by the penial–vaginal ratio in which the vagina is considerably shorter than the penis.
Billordia Stanisic, 2010
Diagnosis
Shell medium to large (diameter 15–26 mm), subdiscoidal with a low spire; whorls rounded (average 5.250), sutures weakly impressed; protoconch with weak radial ridges, teleoconch sculpture of minute scattered pustules overlain by crowded periostracal scales; lip sharply reflected; umbilicus open, partly covered by the reflected columella. Colour yellowish-brown with numerous prominent brown spiral bands of varying widths, with or without a brown circum-umbilical patch, and a white lip. Reproductive system with long, knotted epiphallus entwined in connective tissue at the epiphallic–penial junction; penis approximately twice the length of the vagina.
Range
Western foothills of the southern Connors Range and Peak Range, SCQ and MEQ (inland).
Remarks
Billordia is distinguished from Figuladra, Euryladra and Brigaladra by the subdiscoidal shell with pustulose microsculpture on the teleoconch, shell coiling pattern and knotted epiphallus in the genitalia.
Data availability
All data generated or analysed during this study are included in this article and the Supplementary Information. Sequence information generated in this study are available at GenBank, accession numbers OL753690–OL753711 (COI) and OL839880–OL839914 (16S). Zoobank: urn:lsid:zoobank.org:pub:DEA7E10A-8708-4E8C-8F90-4A66682D2727.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This research was funded by HDR candidate support funding from Griffith University to L. Stanisic and C. McDougall.
Supplementary material
Supplementary material is available online.
Acknowledgements
We acknowledge Griffith University, the Queensland Museum and Australian Museum for allowing us to access facilities and collections for this study. We sincerely thank Paul Oliver from Queensland Museum and Griffith University for continued support and input into the biogeography story and Andrew Hugall for sharing the sequences used in this study. We would like to express our deep gratitude to Dr John Stanisic, former Curator of Malacology and honorary researcher at Queensland Museum for support during field work and helpful comments during the study. A special thanks to Craig Eddie, environmental consultant and malacologist in Roma, for assistance with the collection of some specimens from SCQ for this project in times when fieldwork was unable to be conducted. Thank you to Geoff Thompson from the Queensland Museum Digital Imaging Unit for providing high resolution images of some of the genital anatomy and improving the quality of several other images, and to Kim Maxwell for cartographic expertise. We thank the reviewers for constructive comments.
References
Bishop, MJ (1978). A revision of the genus Thersites Pfeiffer (Pulmonata: Camaenidae). Journal of the Malacological Society of Australia 4, 9–21.| A revision of the genus Thersites Pfeiffer (Pulmonata: Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Byrne, M, Yeates, D, Joseph, L, Kearney, M, Bowler, J, Williams, M, Cooper, S, Donnellan, S, Keogh, J, and Leys, R (2008). Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota. Molecular Ecology 17, 4417.
| Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota.Crossref | GoogleScholarGoogle Scholar | 18761619PubMed |
Byrne, M, Steane, DA, Joseph, L, Yeates, DK, Jordan, GJ, Crayn, D, Aplin, K, Cantrill, DJ, Cook, LG, Crisp, MD, Keogh, JS, Melville, J, Moritz, C, Porch, N, Sniderman, JMK, Sunnucks, P, and Weston, PH (2011). Decline of a biome: evolution, contraction, fragmentation, extinction and invasion of the Australian mesic zone biota. Journal of Biogeography 38, 1635–1656.
| Decline of a biome: evolution, contraction, fragmentation, extinction and invasion of the Australian mesic zone biota.Crossref | GoogleScholarGoogle Scholar |
Chiba, S (1999). Accelerated evolution of land snails Mandarina in the oceanic Bonin Islands: evidence from mitochondrial DNA sequences. Evolution 53, 460–471.
| Accelerated evolution of land snails Mandarina in the oceanic Bonin Islands: evidence from mitochondrial DNA sequences.Crossref | GoogleScholarGoogle Scholar | 28565404PubMed |
Clark, SA (2009). A review of the land snail genus Meridolum (Gastropoda: Camaenidae) from central New South Wales, Australia. Molluscan Research 29, 61–120.
Couper, PJ, and Hoskin, CJ (2008). Litho‐refugia: the importance of rock landscapes for the long‐term persistence of Australian rainforest fauna. Australian Zoologist 34, 554–560.
Cox, JC (1872). Description of a new volute and twelve new species of land‐shells from Australia and the Solomon Islands. Proceedings of the Zoological Society of London 1871, 643–647.
Cox, JC (1873). Descriptions of new species of land and marine shells from Australia and the Solomon and Louisiade Islands. Proceedings of the Zoological Society of London 1873, 564–569.
Criscione, F, and Köhler, F (2013). Conserved shell disguises diversity in Mesodontrachia land snails from the Australian Monsoon Tropics (Gastropoda: Camaenidae). Zoologica Scripta 42, 389–405.
| Conserved shell disguises diversity in Mesodontrachia land snails from the Australian Monsoon Tropics (Gastropoda: Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Criscione, F, and Köhler, F (2014a). Molecular phylogenetics and comparative anatomy of Kimberleytrachia Köhler, 2011 – a genus of land snail endemic to the coastal Kimberley, Western Australia with description of new taxa (Gastropoda, Camaenidae). Contributions to Zoology 83, 245–267.
| Molecular phylogenetics and comparative anatomy of Kimberleytrachia Köhler, 2011 – a genus of land snail endemic to the coastal Kimberley, Western Australia with description of new taxa (Gastropoda, Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Criscione, F, and Köhler, F (2014b). Cardiotrachia and Rachita – two new land snail genera from the East Kimberley, Western Australia (Eupulmonata: Camaenidae) Systematics and Biodiversity 12, 55–64.
| Cardiotrachia and Rachita – two new land snail genera from the East Kimberley, Western Australia (Eupulmonata: Camaenidae)Crossref | GoogleScholarGoogle Scholar |
Davison, A, Blackie, RLE, and Scothern, GP (2009). DNA barcoding of stylommatophoran land snails: a test of existing sequences. Molecular Ecology Resources 9, 1092–1101.
| DNA barcoding of stylommatophoran land snails: a test of existing sequences.Crossref | GoogleScholarGoogle Scholar | 21564847PubMed |
Folmer, O, Black, M, Hoeh, W, Lutz , R, and Vrijenhoek, R (1991). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3, 294–299.
Hamilton, ZR (2021). Repeated evolution of an undescribed morphotype of Rhagada (Gastropoda: Camaenidae) from the inland Pilbara, Western Australia. Invertebrate Systematics 35, 203–215.
| Repeated evolution of an undescribed morphotype of Rhagada (Gastropoda: Camaenidae) from the inland Pilbara, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Hugall, A, Moritz, C, Moussalli, A, and Stanisic, J (2002). Reconciling paleodistribution models and comparative phylogeography in the Wet Tropics rainforest land snail Gnarosophia bellendenkerensis (Brazier 1875). Proceedings of the National Academy of Sciences of the United States of America 99, 6112–6117.
| Reconciling paleodistribution models and comparative phylogeography in the Wet Tropics rainforest land snail Gnarosophia bellendenkerensis (Brazier 1875).Crossref | GoogleScholarGoogle Scholar | 11972064PubMed |
Hugall A, Stanisic J, Moritz C (2003) Phylogeography of terrestrial gastropods: the case of the Sphaerospira lineage and history of Queensland rainforests. In ‘Molecular systematics and phylogeography of Mollusks’. (Eds L Lydeard C, R David) pp. 270–301. (Smithsonian Institution Press: Washington, DC, USA)
Hugall AF (2011) Phylogeny and biogeography of the camaenid land snails of eastern Australia. PhD dissertation, University of Adelaide. Available at https://hdl.handle.net/2440/119079
Hugall, AF, and Stanisic, J (2011). Beyond the prolegomenon: a molecular phylogeny of the Australian camaenid land snail radiation. Zoological Journal of the Linnean Society 161, 531–572.
| Beyond the prolegomenon: a molecular phylogeny of the Australian camaenid land snail radiation.Crossref | GoogleScholarGoogle Scholar |
International Trust for Zoological Nomenclature (2012) ‘International Code of Zoological Nomenclature’, 4th edn. (ICZN: London, UK) Available at https://www.iczn.org/the-code/the-code-online/ [Verified 17 August 2021]
Iredale, T (1933). Systematic notes on Australian land shells. Records of the Australian Museum 19, 37–59.
| Systematic notes on Australian land shells.Crossref | GoogleScholarGoogle Scholar |
Iredale, T (1937). A basic list of the land Mollusca of Australia. Part II. Australian Zoologist 9, 1–39.
Kalyaanamoorthy, S, Minh, BQ, Wong, TKF, von Haeseler, A, and Jermiin, LS (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nature Methods 14, 587–589.
| ModelFinder: fast model selection for accurate phylogenetic estimates.Crossref | GoogleScholarGoogle Scholar | 28481363PubMed |
Kameda, Y, Kawakita, A, and Kato, M (2007). Cryptic genetic divergence and associated morphological differentiation in the arboreal land snail Satsuma (Luchuhadra) largillierti (Camaenidae) endemic to the Ryukyu Archipelago, Japan. Molecular Phylogenetics and Evolution 45, 519–533.
| Cryptic genetic divergence and associated morphological differentiation in the arboreal land snail Satsuma (Luchuhadra) largillierti (Camaenidae) endemic to the Ryukyu Archipelago, Japan.Crossref | GoogleScholarGoogle Scholar | 17500012PubMed |
Katoh, K, and Standley, DM (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30, 772–780.
| MAFFT multiple sequence alignment software version 7: improvements in performance and usability.Crossref | GoogleScholarGoogle Scholar | 23329690PubMed |
Kearse, M, Moir, R, Wilson, A, et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649.
| Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data.Crossref | GoogleScholarGoogle Scholar | 22543367PubMed |
Köhler, F (2010a). Three new species and two new genera of land snails from the Bonaparte Archipelago in the Kimberley, Western Australia (Pulmonata, Camaenidae). Molluscan Research 30, 1–16.
Köhler, F (2010b). Uncovering local endemism in the Kimberley, Western Australia: description of new species of the genus Amplirhagada Iredale, 1933 (Pulmonata, Camaenidae). Records of the Australian Museum 62, 217–284.
| Uncovering local endemism in the Kimberley, Western Australia: description of new species of the genus Amplirhagada Iredale, 1933 (Pulmonata, Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Köhler, F (2011a). The camaenid species of the Kimberley Islands, Western Australia (Stylommatophora: Helicoidea). Malacologia 54, 203–406.
| The camaenid species of the Kimberley Islands, Western Australia (Stylommatophora: Helicoidea).Crossref | GoogleScholarGoogle Scholar |
Köhler, F (2011b). Australocosmica, a new genus of land snails from the Kimberley, Western Australia (Eupulmonata, Camaenidae). Malacologia 53, 199–216.
| Australocosmica, a new genus of land snails from the Kimberley, Western Australia (Eupulmonata, Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Köhler, F, and Bouchet, P (2020). On unavailable genus-group names introduced by Tom Iredale for Australian non-marine gastropods: nomenclatural clarifications and descriptions of new genera. Molluscan Research 40, 150–159.
| On unavailable genus-group names introduced by Tom Iredale for Australian non-marine gastropods: nomenclatural clarifications and descriptions of new genera.Crossref | GoogleScholarGoogle Scholar |
Köhler, F, and Criscione, F (2015). A molecular phylogeny of camaenid land snails from north-western Australia unravels widespread homoplasy in morphological characters (Gastropoda, Helicoidea). Molecular Phylogenetics and Evolution 83, 44–55.
| A molecular phylogeny of camaenid land snails from north-western Australia unravels widespread homoplasy in morphological characters (Gastropoda, Helicoidea).Crossref | GoogleScholarGoogle Scholar | 25463754PubMed |
Köhler, F, and Johnson, MS (2012). Species limits in molecular phylogenies: a cautionary tale from Australian land snails (Camaenidae: Amplirhagada Iredale, 1933). Zoological Journal of the Linnean Society 165, 337–362.
| Species limits in molecular phylogenies: a cautionary tale from Australian land snails (Camaenidae: Amplirhagada Iredale, 1933).Crossref | GoogleScholarGoogle Scholar |
Köhler, F, Criscione, F, and Shea, M (2020). A mitochondrial phylogeny uncovers taxonomic ambiguity and complex phylogeographic patterns in the eastern Australian land snail Austrochloritis (Stylommatophora, Camaenidae). Journal of Zoological Systematics and Evolutionary Research 58, 1005–1020.
| A mitochondrial phylogeny uncovers taxonomic ambiguity and complex phylogeographic patterns in the eastern Australian land snail Austrochloritis (Stylommatophora, Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Neldner VJ, Butler DW, Guymer GP (2019) ‘Queensland’s regional ecosystems: building a maintaining a biodiversity inventory, planning framework and information system for Queensland. Version 2.1.’ (Queensland Herbarium, Department of Environment and Science: Brisbane, Qld, Australia)
Nguyen, L-T, Schmidt, HA, von Haeseler, A, and Minh, BQ (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating Maximum Likelihood phylogenies. Molecular Biology and Evolution 32, 268–274.
| IQ-TREE: a fast and effective stochastic algorithm for estimating Maximum Likelihood phylogenies.Crossref | GoogleScholarGoogle Scholar | 25371430PubMed |
Oba, Y, Ôhira, H, Murase, Y, Moriyama, A, and Kumazawa, Y (2015). DNA barcoding of Japanese click beetles (Coleoptera, Elateridae). PLoS One 10, e0116612.
| DNA barcoding of Japanese click beetles (Coleoptera, Elateridae).Crossref | GoogleScholarGoogle Scholar | 25636000PubMed |
Parkyn, J, and Newell, DA (2013). Australian land snails: a review of ecological research and conservation approaches. Molluscan Research 33, 116–129.
| Australian land snails: a review of ecological research and conservation approaches.Crossref | GoogleScholarGoogle Scholar |
Perez, KE, Defreitas, N, Slapcinsky, J, Minton, RL, Anderson, FE, and Pearce, TA (2014). Molecular phylogeny, evolution of shell shape, and DNA barcoding in Polygyridae (Gastropoda: Pulmonata), an endemic North American clade of land snails. American Malacological Bulletin 32, 1–31.
| Molecular phylogeny, evolution of shell shape, and DNA barcoding in Polygyridae (Gastropoda: Pulmonata), an endemic North American clade of land snails.Crossref | GoogleScholarGoogle Scholar |
Ronquist, F, and Huelsenbeck, JP (2003). MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574.
| MrBayes 3: Bayesian phylogenetic inference under mixed models.Crossref | GoogleScholarGoogle Scholar | 12912839PubMed |
Sela, I, Ashkenazy, H, Katoh, K, and Pupko, T (2015). GUIDANCE2: accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Research 43, W7–W14.
| GUIDANCE2: accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters.Crossref | GoogleScholarGoogle Scholar | 25883146PubMed |
Smith BJ (1992) Non-marine Mollusca. In ‘Zoological Catalogue of Australia’. (Ed. WWK Houston) Vol. 8, p. 405. (Australian Government Publishing Service)
Solem, A (1973). Convergence in pulmonate radulae. The Veliger 15, 165–170.
Solem A (1979) A theory of land snail biogeographic patterns through time. In ‘Pathways in Malacology’. (Eds S Van der Spoel, AC Van Bruggen, J Lever) pp. 225–249. (Scheltema & Holkema: Utrecht, Netherlands)
Solem, A (1984). Camaenid land snails from Western and central Australia (Mollusca: Pulmonata: Camaenidae). IV. Taxa from the Kimberley, Westraltrachia Iredale, 1933 and related genera. Records of the Western Australian Museum Supplements 17, 427–705.
Solem, A (1985). Camaenid land snails from Western and central Australia (Mollusca: Pulmonata: Camaenidae). V. Remaining Kimberley genera and addenda to the Kimberley. Records of the Western Australian Museum Supplements 20, 707–981.
Solem, A (1992). Camaenid land snails from Southern and eastern South Australia, excluding Kangaroo Island. Records of the South Australian Museum. Monograph Series 2, 1–338.
Solem, A (1997). Camaenid land snails from western and central Australia (Mollusca: Camaenidae). VII. Taxa from Dampierland through the Nullarbor. Records of the Western Australian Museum 50, 1461–1906.
Solem A, van Bruggen AC (1984) World-wide snails: biogeographical studies on non-marine Mollusca. In ‘Biogeographical Studies on Non-marine Mollusca’. (Eds A Solem, AC van Bruggen) pp. 6–22. (E.J. Brill/W. Backhuys: Leiden, Netherlands)
Stanisic, J (1994). The distribution and patterns of species diversity of land snails in eastern Australia. Memoirs of the Queensland Museum 36, 207–214.
Stanisic, J (1996). A new camaenid land snail from the Wet Tropics Biogeographic Region, northeastern Queensland (Eupulmonata, Camaenidae. Memoirs of the Queensland Museum 39, 355–363.
Stanisic, J (2013). A new genus, Denhamiana gen. nov., and two new species of land snail from inland central Queensland (Eupulmonata, Camaenidae). Memoirs of the Queensland Museum 58, 367–374.
Stanisic, J (2016). A new genus and two new species of land snail from geographically remote lithorefugial habitats in Queensland including a remarkable range extension for Quistrachia Iredale, 1939 (Gastropoda: Eupulmonata: Camaenidae). Memoirs of the Queensland Museum, Nature 60, 43–51.
| A new genus and two new species of land snail from geographically remote lithorefugial habitats in Queensland including a remarkable range extension for Quistrachia Iredale, 1939 (Gastropoda: Eupulmonata: Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Stanisic J, Ponder WF (2004) Forest snails in eastern Australia – one aspect of the other 99%. In ‘Conservation of Australia’s Forest Fauna’. (Ed. D Lunney) pp. 127–149. (Royal Zoological Society of New South Wales)
Stanisic, L, and Stanisic, J (2020). Reconciling nomenclature and type localities of some Queensland hadroid camaenids: a prelude to a revision of Figuladra Köhler & Bouchet, 2020 (Gastropoda: Eupulmonata: Camaenidae). Memoirs of the Queensland Museum, Nature 62, 1–24.
Stanisic, J, Cameron, RAD, Pokryszko, BM, and Nekola, JC (2007). Forest snail faunas from S.E. Queensland and N.E. New South Wales (Australia): patterns of local and regional richness and differentiation. Malacologia 49, 445–462.
| Forest snail faunas from S.E. Queensland and N.E. New South Wales (Australia): patterns of local and regional richness and differentiation.Crossref | GoogleScholarGoogle Scholar |
Stanisic J, Shea M, Potter D, Griffiths O (2010) ‘Australian land snails. Volume 1. A field guide to eastern Australian species.’ (Bioculture Press: Mauritius)
Stanisic J, Shea M, Potter D, Griffiths O (2018) ‘Australian land snails. Volume 2. A field guide to southern, central and western Australian species.’ (Bioculture Press: Singapore)
Sutcharit, C, Asami, T, and Panha, S (2007). Evolution of whole‐body enantiomorphy in the tree snail genus Amphidromus. Journal of Evolutionary Biology 20, 661–672.
| Evolution of whole‐body enantiomorphy in the tree snail genus Amphidromus.Crossref | GoogleScholarGoogle Scholar | 17305832PubMed |
Tamura, K, Stecher, G, Peterson, D, Filipski, A, and Kumar, S (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution 30, 2725–2729.
| MEGA6: Molecular Evolutionary Genetics Analysis version 6.0.Crossref | GoogleScholarGoogle Scholar | 24132122PubMed |
Taylor, JPA, Johnson, MS, and Stankowski, S (2015). Molecular phylogenetics and complementary geographical distributions of species of the Western Australian land snail genera Plectorhagada Iredale, 1933 and Strepsitaurus Solem, 1997 (Gastropoda: Camaenidae). Zoological Journal of the Linnean Society 174, 305–321.
| Molecular phylogenetics and complementary geographical distributions of species of the Western Australian land snail genera Plectorhagada Iredale, 1933 and Strepsitaurus Solem, 1997 (Gastropoda: Camaenidae).Crossref | GoogleScholarGoogle Scholar |
Webb LJ, Tracey JG (1981) Australian rainforests: patterns and change. In ‘Ecological Biogeography of Australia’. (Ed. A Keast) pp. 605–694. (W. Junk: The Hague, Netherlands)
Zou, S, Li, Q, Kong, L, Yu, H, and Zheng, X (2011). Comparing the usefulness of distance, monophyly and character-based DNA barcoding methods in species identification: a case study of neogastropoda. PLoS One 6, e26619.
| Comparing the usefulness of distance, monophyly and character-based DNA barcoding methods in species identification: a case study of neogastropoda.Crossref | GoogleScholarGoogle Scholar | 22039517PubMed |