

Economic effects of nutritional constraints early in life of cattle
A. R. Alford A B C , L. M. Cafe A B , P. L. Greenwood A B and G. R. Griffith A BA Cooperative Research Centre for Beef Genetic Technologies, University of New England, Armidale, NSW 2351, Australia.
B New South Wales Department of Primary Industries, Armidale, NSW 2351, Australia.
C Corresponding author. Email: andrew.alford@dpi.nsw.gov.au
Animal Production Science 49(6) 479-492 https://doi.org/10.1071/EA08266
Submitted: 29 October 2008 Accepted: 16 February 2009 Published: 13 May 2009
Abstract
An experiment was conducted at the Grafton Agricultural Research Station on the northern coast of New South Wales whereby low and high pasture nutritional systems were imposed on a herd of Hereford cows during pregnancy and from birth to weaning in a factorial design. Offspring representing extremes of growth to birth and/or weaning were then selected for study of long-term consequences of growth early in life. Implications of the nutritional treatments of cows on subsequent weaning rates were also tested with data from previous studies. The extent to which these extreme maternal nutritional and offspring growth scenarios affected herd profitability was tested with the Beef-N-Omics decision support package. For the representative cattle enterprise modelled, gross margin per hectare ranged from $A114 to $A132. In all cases, the gross margin for those groups with fetal growth based on a higher level of nutrition exceeded that of their peers on a lower level of nutrition. It is more profitable for cows and calves to have access to a high standard of nutrition during pregnancy and up to weaning than for them to have access only to a poor standard of nutrition. Incorporating differential weaning rates following maternal nutritional treatments reduced gross margins per hectare by up to 30%. On average, a 1% reduction in weaning rate resulted in a 4.5% reduction in gross margin. Restricted cow–calf nutrition affects the future cow fertility, as well as the current calf progeny, economically.
Additional keywords: Australia, beef, birthweight, economics, evaluation, fetal programming, nutrition.
Introduction
Australian beef-cattle producers are increasingly managing their production systems to target specific markets better for their stock. They aim to grow cattle efficiently on pasture during the early phases of their lives, followed by the use of high-quality feedstuffs during later growth phases to reduce the risk of not meeting the targeted market specifications. However, pasture-reliant growth of cattle is typically a prolonged process during which cattle experience widely differing nutrition levels owing to variable pasture quality and availability, climatic extremes and constraints on optimal management of cattle and pastures. Finishing cattle on grain may not be sufficient to overcome early life nutritional deficiencies. Hence, it is important to understand the influences of different patterns of nutrition and growth during pregnancy and lactation on the performance of the offspring. This requirement has assumed greater importance in recent times with the ongoing drought and the prospect of even greater variability in climate because of global warming.
In relation to the underlying biological relationships, fetal growth of cattle has been extensively studied in relation to calf survival. It has been shown that growth of the fetal calf can be slowed during the latter half of pregnancy by severely restricted maternal nutrition (Holland and Odde 1992). Similarly, during late pregnancy, the size of the dam can restrict the growth of fetuses with high prenatal growth potential (Joubert and Hammond 1958; Ferrell 1991). However, until recently the consequences of fetal growth for subsequent performance, particularly in relation to carcass and eating-quality characteristics at market weights, have been less well understood (Greenwood et al. 2006, 2009; Greenwood and Cafe 2007). Although severe growth retardation during fetal life has been shown to reduce muscle growth and increase fatness later in life in sheep (Villette and Theriez 1981; Greenwood et al. 1998, 2000), fetal growth retardation appears to have lesser effects on body composition in cattle (Tudor et al. 1980; Greenwood et al. 2006, 2009; Greenwood and Cafe 2007). Recent research has also established that cattle severely growth-restricted during fetal life consume less feed during feedlotting later in life than do their counterparts well grown during fetal life (Cafe et al. 2009). However, when compared at the same feedlot entry weight, significant differences in feed intake were not apparent, and the low- and high-birthweight cattle did not differ in feed efficiency (Cafe et al. 2009).
Influences of preweaning nutrition on postweaning growth and composition of cattle at market weights are better described (Berge 1991; Hearnshaw 1997; Greenwood and Cafe 2007). Nutritional restriction early in postnatal life can have prolonged effects on subsequent growth of cattle (Reardon and Everitt 1973), although severe restriction before weaning results in similar carcass fatness or slightly leaner cattle when recovered for prolonged periods on pasture (Tudor et al. 1980; Berge 1991; Hearnshaw 1997; Cafe et al. 2006a; Greenwood et al. 2006, 2009; Greenwood and Cafe 2007). As with fetal growth, cattle severely restricted in growth to weaning also consumed less feed during feedlotting during later life than did cattle grown well from birth to weaning (Cafe et al. 2009). Again, however, these differences in feed intake were due to differences in weight of the cattle at the same age and were not evident when the cattle were compared at the same feedlot entry weight. Further, differences in feed efficiency, owing to growth to weaning, were not evident (Cafe et al. 2009).
Commercially, information on consequences of maternal nutrition and offspring performance is important for beef-cattle producers to assess whether it is economically worthwhile to invest in different nutritional systems. Hence, in the present paper a farm-level modelling system is described that allows an economic evaluation of experimental results of maternal nutritional systems and offspring performance reported by Greenwood et al. (2006, 2009) and Cafe et al. (2006b, 2009), and the economic outcomes of applying this system in two different ways are reported. The economic analysis also examines the implications of lower cow fertility and subsequent weaning rates that would be expected (Barlow et al. 1994) to result from the varying nutritional treatments of cows reported in the study. Implications are then drawn for beef-cattle producers in the region where the study was undertaken.
Materials and methods
Overview of the animal experiment
The present experiment was conducted as a part of the research program of the Cooperative Research Centre for Cattle and Beef Quality (Beef CRC). The objective was to study the long-term consequences of maternal nutrition and different patterns of early life growth on subsequent growth to heavy market weights, and on the resulting feedlot performance, carcass, yield and eating-quality characteristics. The experiment is detailed fully in Cafe et al. (2006b) and Greenwood et al. (2006) and in the associated papers in this special issue of Animal Production Science (Cafe et al. 2009; Greenwood et al. 2009).
The dams used in the experiment were Hereford cows and heifers (360 in total), and the sires were Piedmontese (high muscling) and Wagyu (high marbling), eight of each sire breed. The experiment was conducted at Grafton Agricultural Research and Advisory Station, on the northern coast of New South Wales (NSW), across two breeding cycles. Mating commenced in November 2000, with calves being born in the winter–spring period of 2001 and again in the winter–spring period of 2002. Cows were managed within high and low nutritional systems at Grafton from diagnosis of pregnancy during the first trimester to parturition. Lactating cows and their calves were similarly managed until weaning at ~7 months of age. This produced calves with high or low birthweights, and high or low growth to weaning. Half of the cows and calves swapped nutritional treatments at birth, resulting in progeny with the following combinations of early life nutrition:
-
LL, low during pregnancy, and low during lactation;
-
LH, low during pregnancy, and high during lactation;
-
HL, high during pregnancy, and low during lactation; and
-
HH, high during pregnancy, and high during lactation.
The number of progeny reaching weaning from the two breeding cycles was 534. Of these, 240 core animals were selected for further study, representing extremes of birthweight and growth to weaning. Following weaning, the core animals were backgrounded on improved temperate pastures at Glen Innes Agricultural Research and Advisory Station on the New England Tablelands, in steer- or heifer-cohort groups until they reached a weight of ~500 kg. They were then grain-fed for 120 days at the Tullimba research feedlot, west of Armidale. They were ~30 months of age at feedlot exit, when they were slaughtered at the John Dee Abattoir near Warwick in south-eastern Queensland, where their carcass attributes were assessed. The final progeny group was slaughtered in March 2005. This core group of 240 cattle has been used in the economic analysis as they represent the most extreme early life growth scenarios likely to occur and because there is a complete dataset relating to each of these animals.
This experimental protocol approximated common production systems in the region, where weaners are bred on the northern coast of NSW and then grown out on the better-quality pastures of the NSW Northern Tablelands, before feedlot finishing. The management events for the cows and their progeny within the experiment were detailed by Cafe et al. (2006b) and Greenwood et al. (2006).
Nutritional treatments at Grafton
The high and low nutritional treatments for the cows and their offspring were pasture-based, with the use of supplementation as required to meet the experimental growth targets and to ensure animal-welfare requirements were maintained within the low nutritional system (Cafe et al. 2006b).
The low nutritional system was based on poor-quality native and naturalised pastures on unfertilised duplex soils. The target growth rate for these calves from birth to weaning was 500–600 g/day. The high nutritional system was based on improved-pasture species on heavy alluvial flats and higher red alluvium. Irrigated ryegrass was used to fill the winter feed gap that prevails in this region, and to ensure continued high nutrition. The target growth rate for these calves from birth to weaning was 800–900 g/day.
During both breeding cycles, some strategic feed supplementation was required, and this was more significant in 2002–2003 because of very severe drought. Lactating cows and calves were fed a pelleted, complete ration for most of the preweaning period because there was very little dry matter available. The supplementation allowed the research growth targets to be met, while ensuring welfare standards were maintained (Cafe et al. 2006b).
The growth parameters for each early life growth group selected for the study of their subsequent performance are shown in Table 1. On average, the calves selected for the low-birthweight groups were 10 kg lighter than their high-birthweight counterparts. Subsequent high nutrition up until weaning significantly raised daily gain and weaning weights, although those calves on high nutrition during pregnancy (HL and HH) were always heavier than their respective low-nutrition counterparts (LL and LH).

|
Backgrounding at Glen Innes
Three cohorts of progeny (two steer cohorts and one heifer cohort) were sent to Glen Innes to start pasture backgrounding soon after weaning. They grazed as cohort groups (n = 80 per cohort) for ~18 months. The pastures at Glen Innes comprised good-quality temperate improved species (i.e. fescue, phalaris and white clover), producing large cattle-weight gain during spring and summer, and little gain in the cold winter period.
Some supplementation was also required at Glen Innes during the 2002–2003 drought to ensure animals reached their target feedlot entry weights by ~26 months of age. The growth of the early life treatment groups during backgrounding is shown in Table 2. Calves with high growth rate during pregnancy (HL and HH) were always ahead of their low-growth counterparts (LL and LH). However, there was also some compensatory gain evident, with those calves that entered backgrounding on a slower growth path between birth and weaning (LL and HL) having faster backgrounding growth rates than their LH and HH peers.

|
Feedlot finishing at Tullimba
The 240 core cattle were grain-fed at Tullimba feedlot for ~120 days after backgrounding. The feedlot growth and exit weights are shown in Table 3. Because of the experimental protocols, feedlot entry was based on cohort age, and not on individual weights as would be done commercially. At the completion of feedlotting, the cattle were ~30 months of age. There was no evidence of compensatory gain in the feedlot – those animals that went in lighter grew more slowly than those that went in heavier.

|
Carcass characteristics
Following feedlotting, the cattle were transported to John Dee Abattoir at Warwick in southern Queensland for slaughter and assessment of carcass attributes. Detailed carcass, yield and meat-quality data were collected, as reported by Greenwood et al. (2006, 2009). Importantly for the economic analysis, the carcasses from all four treatments met the market specifications for the grain-fed export market that the abattoir supplies. Average carcass weights for the treatment groups were 353 kg, 376 kg, 383 kg and 409 kg for the LL, LH, HL and HH treatment groups, respectively.
Restricted early life growth resulting in differences in the weight of calves at weaning persisted until 30 months of age. Animals that were smaller at weaning remained smaller at slaughter. Some compensation occurred following restriction of growth during lactation, but not following restriction of growth during pregnancy.
Carcass quality of these animals was not adversely affected by growth restriction during early life, under present carcass specifications. Smaller animals had smaller carcasses, and tended to yield a higher proportion of meat and a lower proportion of fat trim than did the larger animals (Greenwood et al. 2006, 2009). This was mainly due to the weight differences placing them at different stages of the growth curve, so that the larger animals had undergone more fattening than the smaller ones. Subcutaneous- and intramuscular-fat measures were not affected to the extent of influencing the value of the carcass. These findings have subsequently been confirmed in cattle grown slowly or rapidly to weaning and backgrounded to the same feedlot entry weights (Cafe et al. 2006a).
Similarly, eating quality of beef was unaffected by growth during early life (see Greenwood et al. 2006); hence, no differences in returns per unit of beef would occur if an objective beef-marketing system, such as Meat Standards Australia, was used.
An important aspect of the experimental design that needs to be considered when interpreting the outcomes of the present study is that nutritional treatments and offspring studied were deliberately selected to represent extremes of maternal nutrition and offspring growth. This necessarily introduces a bias against the low nutritional groups and a bias towards the high nutritional groups. In this sense, the experiment, and the analyses that follow, represent best-case v. worst-case scenarios.
Overview of the economic analyses
The experimental design necessitated various management decisions to be made that would be uneconomic in a context of commercial farm business, e.g. the level of supplementary feeding undertaken and feedlot entry criteria for animals. Therefore, as described in Davies et al. (2009), key results from the experiment were applied to a representative cattle-enterprise model. Thus, an enterprise-level economic evaluation of the experimental outcomes was undertaken, following the three production phases outlined previously. This would be consistent with a representative farmer having a cow–calf breeding enterprise on the northern coast of NSW as well as a growing–backgrounding enterprise in the adjacent Northern Tablelands region. Traditionally, northern coast beef production systems have focussed on store-cattle breeding (Davies et al. 1999), whereas a broader range of enterprises such as breeding and growing out of beef cattle as well as the transfer in of store cattle from coastal regions for growing and finishing is typical of beef production in the Northern Tablelands (Alford et al. 2003). The farmer is also assumed to custom finish his cattle in a local feedlot.
The farmer is assumed to have 200 ha of mixed pasture available on the northern coast, and another 200 ha of mixed pasture available on the Northern Tablelands.
The economic evaluation is based on 2006 average prices and costs, where all dollar values are expressed in Australian dollars.
Beef-N-Omics model
The Beef-N-Omics software package (Dobos et al. 2006) was used in this evaluation. This package has been selected for all farm-level modelling of new technologies relevant to southern Australian beef production systems within the Beef CRC. It has been used by beef extension officers and some commercial producers during several years, enabling practical validation of the model, including the herd-dynamics and metabolic energy-demand algorithms derived from MAFF (1984) standards. The Beef-N-Omics program incorporates feed budgets and financial gross-margin budgets for static herds. It is sufficiently flexible to allow for the input of various ages and liveweights for growing stock from weaning to turn-off.
To reduce the complexity of the economic analysis of the early life nutritional study, it was decided to use Beef-N-Omics assuming the same land resource and associated pasture resource for each of the growth treatments. This avoids confounding effects of different capital investment in land and associated pasture management among the nutritional treatments, confining the analysis to the major focus of the study. Thus, varying areas to improved pastures was not considered. Rather, pasture types and areas for each of the two different regions were held fixed across each growth path, whereas energy available for the cow herd was varied by altering the stocking rate to avoid the use of supplementary feeding and still provide sufficient metabolisable energy (ME) to meet the four sets of cattle-growth rates. For example, for the cow–calf phase at Grafton, 150 breeding cows could be run on the standard 200 ha of pasture to achieve the LL growth path, whereas only 128 breeding cows could be run on the same pasture to reach the HH growth path. Limitations of this approach are recognised, given the simple ME approach used by Beef-N-Omics and the associated pasture modelling; however, the methodology allows for a consistent approach across all four experimental treatments.
Pasture types
An accurate representation of pasture types and growth rates during the year is a crucial input into a Beef-N-Omics analysis. The Beef-N-Omics package contains an extensive pasture library that has been built up from various NSW Agriculture trial datasets that were collated for use in the Prograze program (NSW Agriculture 1996). New pasture types can be added at any time.
Several pasture-growth profiles provided in the Beef-N-Omics library were tested for their similarity to the various pasture types implied by the cow–calf growth profiles for the various treatments. The different pasture mixes modelled in the representative enterprises (described below) were considered to be consistent with the pasture types and relative proportions available to the experimental treatments.
There are three pasture types assumed for the Grafton phase of the experiment, including the following:
-
setaria (Setaria sphacelata), rhodes grass (Chloris gayana) and white clover (Trifolium repens) (100 ha);
-
kikuyu (Pennisetum clandestinum), dominant pasture (60 ha); and
-
short-term ryegrass (Lolium multiflorum), with periodic nitrogen application (40 ha).
The growth rates for these three pasture types are detailed in Alford et al. (2007). They are consistent with those identified by Ashwood et al. (1992).
In relation to the Glen Innes phase of the experiment, introduced pasture mixes are commonly utilised in the Northern Tablelands grazing systems to produce feeder cattle for the feedlot market or to finish beef cattle (Davies and Llewelyn 2006). Such pastures include introduced temperate grasses [e.g. phalaris (Phalaris spp.), fescue (Festuca arundinacea), cocksfoot (Dactylis glomerata), ryegrasses (Lolium spp.)] and legumes [e.g. white clover (Trifolium repens) and subterranean clover (T. subterraneum)]. These pasture types and associated agronomic practices are detailed by Lodge and Whalley (1989), Lowien et al. (1997) and NSW Agriculture (1996).
The Beef-N-Omics model assumes an introduced grass-dominated pasture of fescue (Festuca arundinacea) and phalaris (Phalaris spp.), with at least 20% base dry matter (DM) present as white clover (T. repens) or subterranean clover (T. subterraneum) and with annual-maintenance fertiliser applications. This mix is assumed to cover the entire 200 ha of available grazing land. The growth rates for this pasture mix are shown in Alford et al. (2007).
Cattle prices and costs
Prices and costs used in the analysis are for 2006 and are detailed in Alford et al. (2007). Herd costs and returns for the cow–calf activity at Grafton were derived from a standard NSW Department of Primary Industries budget (NSW DPI 2006), whereas costs for the Glen Innes backgrounding phase were developed independently and detailed in Alford et al. (2007). The costing of the feedlot phase of the experiment was derived from the feedlot operator by using commercial values. These feedlot costs and income for 2006 are also provided in Alford et al. (2007). Aside from fixed per cattle charges, the cost of feedlotting is based on feed consumed at an average price of $260 per tonne as fed assuming 85% DM. Combined feed-intake data for each of the four treatments were used to derive average feed consumed per animal in the feedlot. Average intakes (t/head) during the 120-day feedlotting phase were 1.51, 1.61, 1.68 and 1.76 t/animal for the LL, LH, HL and HH treatments, respectively. Because the carcasses from all four treatments met the market specifications for the grain-fed export market that the abattoir supplies, a common price of $4.00/kg carcass weight was applied to all fed cattle.
Budgets for the development and annual maintenance of the various pastures assumed in the analysis are provided in Alford et al. (2007). The cost used for the Grafton phase in the cost and return calculations is a weighted average based on the relative areas of the two pasture types. Because these pasture costs are calculated on an annual basis, in the cost and return calculations outlined below, the pasture costs are each multiplied by 1.5 to account for the 18-month period that the Grafton pastures are feeding the cows and offspring to weaning, and the 18-month period that the Glen Innes pastures are feeding the backgrounding offspring to feedlot entry.
Cattle weights and growth rates
Given the basic pasture, cost and return data, the economic implications of the various early life growth treatments can be investigated using two approaches, the ‘common backgrounding’ approach and the ‘common feedlot weight’ approach.
In the ‘common backgrounding’ approach, all animals are backgrounded for the same period of time and they all enter the feedlot at the same time but at different weights (up to 80-kg difference across the treatments). They spend the same amount of time in the feedlot but grow at different rates, therefore exiting the feedlot at different weights (up to 100-kg difference). The group weights and growth rates used are those described in Tables 2 and 3. Although this approach would not be regarded as normal commercial practice, there were no adverse commercial implications as the carcasses from all four treatments met the market specifications for the grain-fed export market that the abattoir supplies. On the basis of these assumptions, the costs and returns for each of the four growth paths, by phase and in aggregate, are shown below in Table 11.
In the ‘common feedlot weight’ approach, the farm manager is assumed to adopt different production practices. The animals are backgrounded for different periods of time so as to achieve a more common feedlot entry weight. They spend the same amount of time in the feedlot, grow at more similar rates and exit the feedlot at more similar weights. This approach is more like normal commercial practice, and has been followed in other analyses of cattle experimental data (Davies et al. 2007) where there have been more marked feedlot exit-weight differentials and substantial penalties for non-compliance with market specifications.
As there are no actual data for this type of management for the representative farm, it has to be simulated on the basis of assumptions. The following two main issues arise:
-
whether animals that are held for longer or shorter periods on backgrounding would have the same backgrounding average daily gain (ADG) as that measured in the experiment; and
-
whether animals that enter the feedlot at heavier or lighter weights would have the same average feedlot ADG as that measured in the experiment (with the same carcass specifications).
In relation to the first issue, any longer or shorter period on backgrounding required to achieve a group weight close to the average weight of all groups (~512 kg) would be relatively short in relation to the overall backgrounding period of 18 months. For example, the LL group entered the feedlot at an average weight of 473 kg (Table 3). To put on the additional 39 kg at their average backgrounding ADG of 0.6 kg/day would take ~65 days – or ~10% of the total backgrounding period. This is a relatively minor additional period and is also at the end of the backgrounding period, so growth rates during these extra days should be similar to the average across the whole period. It is assumed therefore, that the same backgrounding ADG would apply across a shorter or longer period for any early life treatment group.
In relation to the second issue, there is a more direct relationship between the entry weight and ADG in the feedlot, and the feedlot ADG (Table 3) are almost three times those during the backgrounding period (Table 2). Data (L. M. Cafe, unpubl. data) on some of these animals, detailing individual feed intakes, provided the basis for estimating adjusted ADG and exit weights for the economic analysis. Of the 240 core progeny, 154 animals were subjected to a detailed feed-intake analysis at the Tullimba feedlot during a 70-day period. These were the heifer cohort born in 2001 and the steer cohort born in 2002. The actual recorded data for these animals are shown in Table 4. The mean entry weight of these 154 animals and the weight for each of the early life treatment groups is less than the weight for all of the core animals reported in Table 3 above (since there are a greater proportion of heifers in this smaller subset). However, their feedlot exit weights were generally greater than those of the core animals.

|
Statistical models were run on this dataset, adjusting for a common feedlot entry weight of 490 kg (the mean weight of all 154 animals).1 The model-predicted adjusted ADG and exit weights for each treatment are shown in Table 5. As expected, the daily gain and exit weights are higher for the LL and LH groups than actually recorded, and lower for the HL and HH groups.

|
The differences in weights and ADG between the actual recorded data for these 154 animals and the model predictions for the common entry weight were calculated and are reported in Table 6. For the two extreme treatment groups, there are differences of >30 kg for entry liveweight, >40 kg for exit liveweight and >0.06 kg/day for ADG.

|
The results from the feed-intake trial animals above are then used to predict the growth of the core animals at a common entry weight, adjusting for the differences in mean weights and ADG of each group by applying proportional weight differences as shown in Table 6. These results are shown in Table 7.

|
The calculated feedlot entry weights are not all the same, with the spread in weights, intakes and ADG being much reduced from the raw data. The implications of these new weights for the backgrounding phase are shown in Table 8.

|
The cost, revenue and profit calculations were adjusted to include the new feedlot exit weights and feed intakes, and an allowance was also made for the extra backgrounding days required by the LL and LH groups, or the fewer backgrounding days required by the HL and HH groups, to achieve the predicted ‘common’ feedlot entry weight. On the basis of these assumptions, differences in costs and returns for each of the growth paths are reported below in Table 13.
Cow reproductive performance
An additional economic analysis was conducted to examine the economic implications of the effect of the nutritional treatments described above on subsequent fertility of the cows. It is well established that cow nutrition has a major influence on reproduction performance, with factors such as cow liveweight, body-condition score and plane of nutrition potentially affecting subsequent pregnancy rates. Although the current experimental design did not address these issues, an attempt was made to account for this influence on reproduction in the economic analysis.
The treatment groups in the current experiment were balanced at prejoining for cow liveweight and condition score and the subsequent cow weights at weaning were found to be significantly different among the groups, ranging from 409 kg for the LL to 510 kg for the HH treatment group (Table 9) (Cafe et al. 2006b). Associated cow condition scores (L. M. Cafe, unpubl. data) also showed some differences among the treatments at weaning (Table 9), with the LL and HL treatments having the lowest condition scores of 4.1 and 4.3, respectively, whereas the LH and HH treatment-group cows had significantly higher condition scores of 5.3 and 5.8, respectively.

|
In previous experiments at the Grafton site, Barlow et al. (1994) studied a variety of cross-breeding strategies, including Bos taurus × B. indicus breeds. Although the Hereford × Simmental and Hereford × Friesian crosses used by Barlow et al. (1994) were not of the extreme type used in the Cafe et al. (2006b, 2009) and Greenwood et al. (2006, 2009) trials, they do provide an approximation of the mature cow size and the impact of the level of nutrition on the subsequent fertility of cows. The Barlow et al. (1994) results are summarised in Table 10.
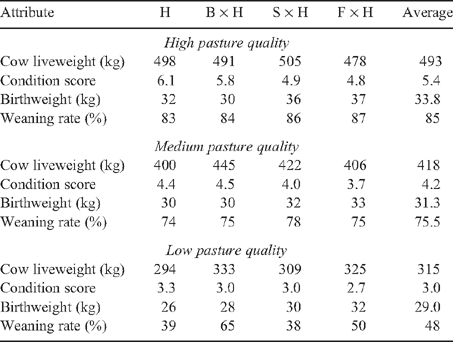
|
A comparison of the prejoining liveweights and associated condition scores of cows reported in Barlow et al. (1994) with those measured in the present study suggests that the average values for the medium pasture-nutrition treatment of Barlow et al. (1994) closely approximates the LL treatment reported for the present study, and the high pasture-nutrition treatment of Barlow et al. (1994) most closely matches the HH treatment. Taking the range of average weaning rates reported by Barlow et al. (1994) for the medium- and high-nutrition groups, it was therefore assumed that the weaning rates in the present study would take the values of 75% for LL and 85% for HH. Given the larger influence of preweaning nutrition treatment on the liveweight and condition score of cows (the HL cows having significantly lower condition score and liveweight at weaning than the LH cows), the HL group were assumed to have a weaning rate of 78% and LH 81%. These rates were approximately equidistant from the respective weaning-rate extremes.
These differing weaning rates were then applied to the enterprise models, with the same cost and price assumptions for each of the breeding, growing out and feedloting phases. The impacts of these potential differences in reproductive efficiency on the economic outcomes of the different nutritional treatments are reported in Table 14 below.
Results
Common backgrounding and feedlotting periods
The gross margins for each of the four growth paths, in aggregate and per breeding cow and per hectare, are shown in Table 11 for the ‘common backgrounding’ approach (not yet including variation in reproductive rates). The three phases are up to weaning, backgrounding and finishing, respectively.

|
Under the LL scenario, where nutrition is restricted both during pregnancy and before weaning, 150 breeding cows can be run on the 200 ha of northern coast pasture. Total costs across the two pasture phases are the highest under this scenario ($90 900), as most of the costs are on a per head basis or are related to the number of breeders. Even though numbers in the feedlot are high, the costs of feedlotting are relatively low because these animals are growing more slowly and they require less feed – $393/cow across the feeding period or $3.28/cow.day. These are the lightest animals coming out of the feedlot (353 kg carcass weight); however, there are 121 of them, so feedlot income is almost $171 000. The net effect is an enterprise gross margin of $45 500, which translates to $114/ha or $303/cow.
In the LH scenario, where nutrition is restricted during pregnancy but not before weaning, only 132 cows can be run on the standard pasture area. Because there are fewer cows and calves, the costs during the two pasture phases are substantially lower ($85 200) than for the LL scenario. Feedlot costs are much the same as for the LL scenario. Although there are fewer animals, they are growing faster and are therefore eating more – $418/cow or $3.48/cow.day. They are slightly heavier than the LL animals but there are fewer of them, so feedlot income is quite a bit lower at $165 400. Overall, the enterprise gross margin is higher than for the LL scenario, by about $1000, which equates to $116/ha but $352/cow, owing to the lower numbers of cows.
In the case of the HL treatment, where nutrition is not restricted during pregnancy but is restricted before weaning, 144 breeding cows can be run on the 200 ha of pasture. Total costs during the two pasture phases ($88 100) are the second highest, and the costs of feedlotting are the highest because there are almost as many animals as in the LL scenario and they are growing more rapidly and they need more feed than those in the LL treatment – $437/cow over the feeding period or $3.97/cow.day. These animals are heavier (383 kg) coming out of the feedlot than those in the LL and LH treatments and there are 118 of them, so feedlot income ($179 200) is the highest of any scenario. The overall result is an enterprise gross margin of almost $52 600, which translates to $132/ha or $365/cow.
Finally, in the HH scenario, nutrition is not restricted either during pregnancy or before weaning. At these consistent high-energy intakes and growth rates, only 128 cows can be run on the standard pasture area. Given these low numbers of animals, the total costs during the two pasture phases are the lowest of any scenario ($84 000) and the total feedlot costs are also low, even though the animals are growing faster and are therefore eating more – $459/cow or $3.83/cow.day. These cattle are heavier than all the other animals but there are fewer of them, so feedlot income is the second lowest at $170 100. Overall, the enterprise gross margin is $49 600, or at $124/ha and $387/cow.
A sensitivity analysis of these results to feed-cattle prices was undertaken (Table 12). It was found that the HH treatment was the most economic treatment at very low cattle prices ($2/kg carcass weight) (50% reduction), whereas the HL treatment achieved the highest enterprise gross margin across the range of cattle prices tested from $3 to $6/kg carcass weight.

|
Common feedlot entry liveweights
The gross margins for each of the four growth paths, in aggregate and per breeding cow and per hectare, were recalculated for the ‘common feedlot liveweight’ approach (again not yet accounting for the different reproductive performances). In particular, the cost, revenue and profit calculations were adjusted to include the new feedlot exit weights and feed intakes (Table 7), and allowance was made for the extra backgrounding days required by the LL and LH groups, or the fewer backgrounding days required by the HL and HH groups, in order to achieve a predicted ‘common’ feedlot entry liveweight (Table 8). The changes in the major values compared with Table 11 are given in Table 13.

|
The gross margin values for the ‘common feedlot weight’ approach are all lower than those for the ‘common backgrounding’ approach. Therefore, given the particular set of experimental data analysed here, it costs more to manage different groups of animals growing at different rates to achieve common feedlot entry weights than to manage animals to enter the feedlot at the same time. The fact that all the animals were able to meet market specifications, even though they had quite different final weights, plays a large role in this outcome. Had there been a large number of animals discounted for not reaching specification, e.g. owing to the adoption of a different carcass-pricing system, the impact of the second method may have been quite different.
The difference in the gross margins for each nutritional group is very small between the two approaches. The largest difference for the ‘common feedlot weight’ approach is for the HH group, and it is only a 3.5% change from the gross margin calculated in the ‘common backgrounding’ approach (Table 11).
Finally, the groups are ranked in the same order for both approaches: the HL group is the most profitable, followed by the HH group, the LH group and then the LL group. It can be concluded therefore, that whichever approach is used, the same implications for producers would hold.
Potential effect of nutritional treatments on subsequent fertility
Because the different feedlot entry weights did not have a major impact on the relative economic outcomes of the different nutritional treatments in the present experiment, either modelling approach could be used to investigate the economic implications of cow fertility as a consequence of the different nutritional treatments. Therefore, the initial Beef-N-Omics models were rerun assuming the adjusted weaning rates (on the basis of Table 10), and the new results were compared with the results previously reported in Table 11. The decreases in gross margin for the LL-, LH- and HL-treatment groups as a consequence of the decrease in subsequent fertility are given in Table 14.

|
The LL treatment modelled with a 75% weaning rate resulted in a 30% decrease in gross margin relative to the LL result, assuming an 85% weaning rate. The LH- and HL-treatment groups were found to have lower gross margins of 14% and 19%, assuming weaning rates of 81% and 78%, respectively. Since the HH group was based on the 85% weaning rate there was no change to this gross margin. The economic ranking of the treatments from most profitable to least profitable are HH ($387/cow), LH ($302/cow), HL ($295/cow) and LL ($213/cow).
Discussion
The experiment analysed here was conducted on the northern coast of New South Wales. Low and high pasture nutritional systems were imposed on a herd of Hereford cows during pregnancy, and then again from birth to weaning, with a factorial design also imposed to select offspring with extremes of growth to birth and/or weaning. Thus, the analyses are essentially best-case and worst-case scenarios because of this selection of extremes.
The results for the ‘common backgrounding’ approach show that it is more profitable for cows and calves to have access to a high standard of nutrition during pregnancy and up to weaning than for them to have access only to a poor standard of nutrition during this time period. The enterprise gross margin for the HH group comes to $49 600 ($124/ha and $387/cow), compared with the enterprise gross margin for the LL group of $45 500 ($114/ha or $303/cow). The net benefit is more than $4000 to the enterprise, $10/ha or $84/cow. The value of the product from the 22 extra cows able to be run on the standard 200 ha of pasture does not compensate for the extra costs of backgrounding and finishing those animals and for their lower carcass weights.
Analysis of costs, returns and profits by the ‘common feedlot weight’ approach gave roughly the same numerical results, and in both cases the groups are ranked in the same order, with the HL group being the most profitable, followed by the HH group, the LH group and then the LL group.
However, another important finding is that the economic implications are not large. There is a 15% difference in the enterprise gross margin between the best-case (HL) and worst-case (LL) scenarios, an 8% difference between the HL and HH groups and only a 2% difference between the LL and LH groups. Given that extremes in offspring growth were selected for the study, this provides more evidence of the relatively flat profit surfaces found in many agricultural industries (Alford et al. 2003; Farquharson 2006; Pannell 2006).
Commercially, the information is important for beef-cattle producers to assess whether it is economically feasible to invest in different genetics and/or different nutrition systems. For example, given the calculated net benefit of $10/ha or $84/cow between the LL and HH groups, this is the upper bound on the amount of money the typical producer could invest each year in pasture improvement to shift them from a LL-growth path to a HH-growth path. However, these conclusions need to be tempered with up to date information on both seasonal conditions and potential supplementary feed costs. It is also planned that this type of information will be used to enhance models for phenotypic prediction of beef-cattle performance. This is the subject of current work in the Beef CRC.
Another outcome examined was the impact of maternal nutritional setbacks on subsequent reproductive performance. It would be expected that cows that suffered a nutritional setback would take longer to reconceive and/or would exhibit a lower reconception rate than cows that had a high standard of nutrition during their previous pregnancy or before reconception. For example, there is some recent work in the USA that indicates economic benefits of improved nutrition of cows pre- and postpartum (Stalker et al. 2006). Although not designed as a component of the experiments reported here, previous studies examining the impact on cow nutrition on subsequent weaning rates were applied to the models used in the present study.
When the economic analysis accounted for the potential impact of nutritional treatments on herd fertility, assuming a steady-state gross-margin analysis, the continuously high nutrition treatment (HH) achieved the highest gross-margin return and the LL treatment the lowest. The impact of weaning rate on the economic outcome of the beef enterprises is in magnitude consistent to that reported by Barlow et al. (1989) who found that a 1% reduction in calving rate translated to a 1.3% reduction in beef-herd gross margin. In the present study, between the HH and LL treatments a 1% reduction in weaning rate resulted in an average of 4.5% reduction in beef-herd gross margin. The major factors influencing the potential economic impact of fertility on a beef enterprise would be whether the herd is self-replacing or buys in replacement females, and the management strategies with respect to females that fail to rebreed as well as the final value of the sale progeny. In the present study all progeny including females are highly valued grain-fed stock and are sold, with replacement females purchased.
Sire-breed effects have not been discussed, although these aspects of the overall experimental design are presented in Cafe et al. (2006b) and in Greenwood et al. (2006). For the core dataset, sire breed and early life nutrition or growth interactions were not evident for the commercial characteristics of interest and, therefore, the economic analyses did not compare sire genotypes.
Conclusions
The economic analysis using the ‘common backgrounding’ approach used in the present study showed that for the representative cattle enterprise modelled, total gross margins ranged from $45 500 for the LL nutritional system to $52 600 for the HL system. Gross margin per hectare ranged from $114 for the LL system to $132 for the HL system, whereas gross margin per breeding cow ranged from $303 for the LL system to $387 for the HH system. In all cases, the gross margin for those groups that started life on a higher level of nutrition exceeded their peers on a lower level of nutrition.
The analysis, when repeated by using the ‘common feedlot weight’ approach showed that it costs more to manage different groups of animals growing at different rates to achieve common feedlot entry weights than to manage animals to enter the feedlot at the same time. However, the differences in gross margins between the two methods are small in all cases. The largest difference is for the HH group, which shows only a 3.5% difference among the results from each approach. By using both approaches, the nutritional groups are ranked in the same order. Thus, whichever method is used, the overall findings are robust and the same implications for producers would hold.
Although the economic implications are not large, it is more profitable for cows and calves to have access to a high standard of nutrition during pregnancy and up to weaning than for them to have access only to a poor standard of nutrition during this time period. This is particularly the case if cow nutrition after calving affects the subsequent reproductive performance, with the LL- and HL-treatment groups showing the largest declines in gross margin per cow when lower potential weaning rates were accounted for. Given that this analysis is undertaken using enterprise gross margins, care is required in translating the results to individual whole-farm businesses, where there is a need to account for all farm resources to determine the profitability of a particular technology or management strategy.
Acknowledgements
Financial and in-kind support of the Cooperative Research Centre for Cattle and Beef Quality, NSW Department of Primary Industries, CSIRO Livestock Industries and the University of New England is gratefully acknowledged. We also acknowledge the considerable research efforts of Ms Helen Hearnshaw and Dr David Hennessy, technical and farm staff of NSW Department of Primary Industries at Grafton, Glen Innes and Armidale, Beef CRC staff at ‘Tullimba’ feedlot, and management and staff at John Dee abattoir, Warwick. Leanne Orr and Kirrily Pollock provided constructive comments on an earlier draft.
Barlow R,
Hearnshaw H,
Arthur PF, Darnell RE
(1994) Evaluation of Hereford and first-cross cows on three pasture systems. I. Calf growth and reproductive performance of young cows. Journal of Agricultural Science 122, 121–129.
| Crossref |
[Verified 23 March 2009]
Davies BL,
Alford AR, Griffith GR
(2009) Economic effects of alternate growth path, time of calving and breed type combinations across southern Australian beef cattle environments: feedlot finishing at the New South Wales experimental site. Animal Production Science 49, 535–541.
| Crossref | GoogleScholarGoogle Scholar |
[Verified 23 March 2009]
Ferrell CL
(1991) Maternal and fetal influences on uterine and conceptus development in the cow: I. Growth of the tissues of the gravid uterus. Journal of Animal Science 69, 1945–1953.
|
CAS |
PubMed |
[Verified 17 October 2008]
Pannell D
(2006) Flat earth economics: the far-reaching consequences of flat payoff functions in economic decision making. Review of Agricultural Economics 28, 553–566.
| Crossref | GoogleScholarGoogle Scholar |

Reardon TF, Everitt GC
(1973) Effects of preweaning nutrition on subsequent growth rate, feed conversion efficiency and carcass composition of identical twin steers. Proceedings of the New Zealand Society of Animal Production 33, 26–38.
Stalker LA,
Adams DC,
Klopfenstein TJ,
Feuz DM, Funston RN
(2006) Effects of pre- and postpartum nutrition on reproduction in spring calving cows and calf feedlot performance. Journal of Animal Science 84, 2582–2589.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Tudor GD,
Utting DW, O’Rourke PK
(1980) The effect of pre- and post-natal nutrition on the growth of beef cattle. III. The effect of severe restriction in early postnatal life on the development of the body components and chemical composition. Australian Journal of Agricultural Research 31, 191–204.
| Crossref | GoogleScholarGoogle Scholar |
Villette Y, Theriez M
(1981) Effect of birth weight on lamb performances. II. Carcass and chemical composition of lambs slaughtered at the same weight. Annales de Zootechnie 30, 169–182.
| Crossref | GoogleScholarGoogle Scholar |
1 These were REML analyses, including effects of birthweight, preweaning nutrition, sex by year cohort, sire genotype and their interactions, with feedlot entry weight as a covariate (linear, and where significant, quadratic) to predict means at equivalent feedlot entry weight.