

Recent advances to improve nitrogen efficiency of grain-finishing cattle in North American and Australian feedlots
Frances Cowley A E , Jenny Jennings B , Andy Cole C and Karen Beauchemin D
A E , Jenny Jennings B , Andy Cole C and Karen Beauchemin D
A School of Environmental and Rural Science, University of New England, Elm Avenue, Armidale, NSW 2350, Australia.
B Texas AgriLife, Texas A&M University, 6500 Amarillo Boulevard West, Amarillo, TX 79106, USA.
C US Department of Agriculture, Agricultural Research Service, 2300 Experiment Road, Bushland, TX 79012, USA.
D Lethbridge Research and Development Centre, Agriculture and Agri-Food Canada, 5403 1st Avenue South, Lethbridge, AB T1J 4B1, Canada.
E Corresponding author. Email: fcowley@une.edu.au
Animal Production Science 59(11) 2082-2092 https://doi.org/10.1071/AN19259
Submitted: 2 May 2019 Accepted: 7 June 2019 Published: 16 September 2019
Journal Compilation © CSIRO 2019 Open Access CC BY-NC-ND
Abstract
Formulating diets conservatively for minimum crude-protein (CP) requirements and overfeeding nitrogen (N) is commonplace in grain finishing rations in USA, Canada and Australia. Overfeeding N is considered to be a low-cost and low-risk (to cattle production and health) strategy and is becoming more commonplace in the US with the use of high-N ethanol by-products in finishing diets. However, loss of N from feedlot manure in the form of volatilised ammonia and nitrous oxide, and nitrate contamination of water are of significant environmental concern. Thus, there is a need to improve N-use efficiency of beef cattle production and reduce losses of N to the environment. The most effective approach is to lower N intake of animals through precision feeding, and the application of the metabolisable protein system, including its recent updates to estimation of N supply and recycling. Precision feeding of protein needs to account for variations in the production system, e.g. grain type, liveweight, maturity, use of hormonal growth promotants and β agonists. Opportunities to reduce total N fed to finishing cattle include oscillating supply of dietary CP and reducing supply of CP to better meet cattle requirements (phase feeding).
Additional keywords: ammonia volatilisation, cattle feedlot, proteins, ruminant metabolism.
Introduction
Feeding to meet or exceed crude-protein (CP) requirement, and maintaining a fixed CP inclusion throughout the finishing period, are long-standing and common practices in formulating grain-based feedlot finishing rations in the dominant feedlot countries of USA, Canada and Australia. Overfeeding nitrogen (N) is considered to be a low-risk diet strategy, if for no other reason than to cover within-pen and across-pen variation in genetic potential. In particular, overfeeding rumen-degradable protein (RDP) is common, because of the perception that the negatives are few, while the cost of non-protein nitrogen (NPN) is low. Overfeeding rumen-undegraded protein (RUP) is uncommon in Australia, as sourcing true RUP is expensive. However, the feeding of steam-flaked corn supplemented with high-RUP corn-processing co-products (various forms of distiller grains (DGs) and sweet bran) in North America has significantly increased RUP intake and overfeeding of N (May et al. 2010; Hales et al. 2012). The utilisation of large quantities of these by-products in finishing diets leads to greater intakes of CP and metabolisable protein (MP) than is required by the animal. There has been little effort to reduce or manipulate the feeding of N in feedlots, in part due to logistics (such as labour, feedmill and feed-delivery management) and to the low purchase cost of urea and high-protein by-products. The impacts of excessive dietary N on the animal are equivocal (Gleghorn et al. 2004; Jennings et al. 2018), but the excreted excess N poses important environmental costs, and the energetic costs to the animal of excess protein may be underestimated. This review discusses recent advances in the protein nutrition of lot-fed cattle, and their application in Australia, Canada and the USA, and examines the need, scope and considerations for improved N efficiency in grain-finished beef production.
Review of current feeding practices in North America, Canada and Australia
While the USA, Canada and Australia all have large feedlot industries, feedlot cattle nutrition recommendations are largely based on research from North America, and the USA in particular. Nevertheless, there are important differences in the diets, protein sources and rumen degradabilities (Table 1), and feeding periods and target markets for feedlot cattle in these countries, which interact with supply and requirements for N.

|
United States
With over 11 million head of beef cattle on feed at any one time in the United States, maintaining and efficiently utilising feed resources is challenging. Average days on feed in the USA is 201, with a minimum of 150 days (Samuelson et al. 2016c). Receiving weight is mostly 272–363 kg, and shipping weight 590–680 kg (Samuelson et al. 2016c). Almost 90% of beef is consumed domestically, and the target carcass is highly marbled, but not over-fattened (Drouillard 2018). Sources of CP have evolved over time and substantially changed in the late 1990s with an increase in grain-based ethanol production. Finishing diets that previously contained costly cottonseed meal, soybean meal and alfalfa (lucerne) as protein sources, now contain cheaper wet-corn distiller-grain by-products (97% of finishing diets contain DGs; Samuelson et al. 2016c). Due to their high availability and low cost, DGs have been incorporated into finishing diets at much higher rates and even used as an energy source as well as the primary protein source (Samuelson et al. 2016c). This, in turn, substantially increased protein content of finishing diets, with nutritionist-recommended ranges of 13.0–14.3% and recommended maximum CP of 15.9% (Samuelson et al. 2016c). However, the low RDP content of corn means that NPN supplementation may still be required, even in DG-supplemented diets (Ponce et al. 2019). As a result, Samuelson et al. (2016c) indicated that many feedlot diets contain greater concentrations of CP than is recommended, and survey respondents still recommended 1.16% DM urea on average, with a maximum NPN inclusion of 2.82% DM. New technologies, such as β agonists and improved genetics, also allow cattle to be fed longer and carcass weights to increase by 50 kg or more. Recently, improved processing of distiller grains has reduced fat content compared with that 10 years ago. This change has lowered the incentive to use these products as an energy source. With current ethanol subsidies diminishing, the US feedlot industry is once again seeking traditional protein sources. By-products that were once highly available have become limited, and resources previously used may have found a different place in the market. Furthermore, migration of dairies into the southern plains of the USA has increased competition for protein sources.
Canada
Canada grain-finishes ~2.5 million cattle per year, with almost 50% of beef being exported, mainly to the USA. Target carcasses, therefore, reflect those in the USA. The industry exclusively uses Bos taurus breeds, unlike Australia and USA, which rely on both B. taurus and B. indicus breeds. Two main feedlot-finishing approaches are used for slaughter cattle in Canada (Legesse et al. 2016). In the calf-fed system, weaned calves that may be backgrounded for a short time are fed a finishing diet for 140–240 days (slaughtered at 15–16 months of age, weighing 590–650 kg). In the yearling-fed system, weaned calves are fed forage over the winter, pastured the following spring and sold to feedlots in the autumn where they are finished for 2–4 months (20–22 months of age and 630–700 kg at slaughter). Regardless of the system, the finishing diet is formulated with few ingredients and only low-level grain processing, with the main grain being dry-rolled or temper-rolled barley. Barley is sometimes partially replaced by wheat or corn, depending on price. In most cases, diets are formulated with minimal consideration for RDP and RUP, to supply a CP content of 12–13.5%. Urea and canola meal are the most common protein sources included, but dry-corn DGs and other meals enter least-cost diet formulation when the economics are favourable. Similarly to the USA, corn DGs were used extensively in the past as an energy and protein source, but recent increases in price currently limit their widespread use in cattle diets. Because the CP content of barley is relatively high compared with that of corn, minimal protein supplementation is generally used. A small amount of forage (mainly barley silage, corn silage, straw), representing ~5–12% of dietary DM, is included as a source of physically effective fibre.
Australia
In 2018, Australia fed 3 million grain-finished cattle. Thirty per cent of Australian beef supplies the domestic market, with the remainder being exported, mainly to the USA, Japan, Korea and China. The domestic market is characterised by heifers and steers fed for 60–80 days, with entry liveweights of 300–350 kg and exit liveweights of 450–550 kg. Export market requirements vary, but target longer feeding (100–120 days, up to >220 days for Japan) and higher receiving (400–450 kg) and final (600–750 kg) liveweights than the domestic market. Dry-matter (and thus, CP) intakes and average daily gains of Australian short-fed cattle are higher than those of Australian export cattle, and many North American cattle. A characteristic distinguishing the typical Australian feedlot-fed animal from those in the USA and Canada is the relatively late entry to the feedlot (commonly 12–18 months of age, 280–450 kg), with an extended forage- or pasture-based growing phase. Seasonal pasture variability and market requirements (age, weight, fatness) require grain finishing. While there are no published surveys of Australian feedlot-diet practices, it is possible to glean some idea of current feeding practices from the limited published research conducted in Australian commercial feedlots. We searched the Google Scholar and Scopus databases for research published between 2010 and 2019, reporting commercial feedlot diets from Australia. This yielded five publications providing information on the composition of 11 feedlot diets (Gaughan et al. 2010; Klieve et al. 2012; Croft et al. 2014; Bai et al. 2015; Goddard et al. 2015). All published diets were based on wheat or barley as their main grain source, and although, before 2005, many Australian feedlot diets were based on sorghum, this is not currently common practice due to export of sorghum to China, and its low feeding value for cattle. The published receiving rations included CP at 13.7–14.3% of DM. The most common protein source in finishing rations was whole cottonseed, with canola meal and lupins also being used. Reported finisher rations included CP at 12.6–14.4% (mean = 13.6%) of DM. Only one research publication has documented a progressive decrease in CP content through the adaptation phase (phase feeding). Few research publications on feedlot cattle nutrition have been published in Australia, and, so, the present review relies heavily on North American research.
The need for improved N efficiency
Environmental issues related to cattle feedlot production are a concern for the cattle industry, consumers and policy makers. Of particular importance is the loss of N from feedlot manure in the form of volatilised ammonia (NH3) and nitrous oxide (N2O). Atmospheric NH3 forms secondary products that contribute to fine particulate matter (PM) and reduced air quality, and deposition of atmospheric NH3 to land can increase N2O emission via nitrification and denitrification, as well as soil acidification, eutrophication of ecosystems, and loss of biodiversity (Beusen et al. 2008). It is estimated that the global NH3-N emission from agricultural systems is ~32 Tg/year, with livestock production accounting for 59–71% (Beusen et al. 2008). The main sources are intensive animal production and spreading of animal manure.
Concentrated animal-feeding operations are ‘hot spots’ of NH3 emissions because manure is left to accumulate in pens or stockpiled (anaerobic) until it can be applied to soils (McGinn et al. 2016). Only 5–20% of N consumed during the feeding period is retained in the carcass (Koenig and Beauchemin 2013), with the remainder being excreted in urine and manure. The proportion of N consumed by feedlot cattle that is excreted in manure is relatively high compared with that for other livestock. For example, Groenestein et al. (2016) estimated that the quantity of feed N required to produce meat is greatest for beef (~5 kg N/kg meat) and least for broiler chickens (~2 kg N/kg meat). However, in contrast to non-ruminants, because most of the protein in the diets of the cow herd, grazing stocker cattle and finishing cattle is from human-non-edible protein sources, cattle production actually increases the value of low-quality grass, grain and by-product proteins to high-quality human-edible protein sources (Baber et al. 2018). Approximately 40–60% of the N consumed daily by feedlot cattle is excreted in urine with 20–30% of N consumed being excreted in faeces (Koenig et al. 2013). Nitrogen in urine is mainly (60–80% of urine N) in the form of urea, which is mostly derived from catabolism of exogenous proteins, with the remaining urine N being derived from catabolism of endogenous proteins (Jones et al. 1990). Urinary urea is rapidly converted to ammonium (NH4+) and NH3 (collectively ammoniacal N given as NHx) by microbial ureases. Consequently, in open-cattle feedlots, ~40–60% of the N consumed is lost to the atmosphere as NH3 (Flesch et al. 2007; Erickson and Klopfenstein 2010). McGinn et al. (2016) calculated that a 22 500-head feedlot emits ~2.5 Mg/day of NH3-N, with a daily economic value of over US$3500 if it were to be replaced with urea fertiliser.
Thus, there is a need to improve N-use efficiency of beef-cattle production and reduce losses of N to the environment. In general, as N intake decreases, N excretion also decreases (Angelidis et al. 2019), thereby reducing the potential for N pollution (Waldrip et al. 2013; Dong et al. 2014). For example, reducing dietary N content from 13.5% to a fixed level closer to animal requirements (11.6%), resulted in a reduction of daily ammonia emissions of 21–40% (Galles et al. 2011). A 1-g reduction in daily N supply in beef diets would decrease manure N excretion by 0.761 g (Angelidis et al. 2019).
Although some loss of N is unavoidable, certain management practices have been shown to improve N-use efficiency by cattle. The most effective approach is to lower N intake of animals through precision feeding, phase feeding and diet formulation (Erickson and Klopfenstein 2010; NASEM 2016). Likewise, reducing the level of inclusion of DGs in diets helps lower dietary CP content and N intake, as diets containing a high inclusion of DGs typically exceed the N requirement of animals (Koenig et al. 2018). Recent studies have shown the potential of incorporating condensed tannins into cattle diets to shift the route of N excretion from urine (more labile) to faeces (Koenig and Beauchemin 2018; Koenig et al. 2018). Condensed tannins can form complexes with dietary proteins, thereby decreasing degradability of protein in the rumen. Feed intake of receiving cattle is often low, yet requirements for MP are high. Thus, there is an opportunity to supplement receiving diets with protected amino acids such as lysine and methionine, so as to supply MP without increasing dietary CP content (NASEM 2016). While targeted supplementation of amino acids may reduce N excretion, its application on-farm is limited due to the costs. Any reduction in N supply must be applied carefully so that production is not compromised; a reduced rate of growth may increase the days on feed required to meet market specifications, which could increase total N emissions per kilogram of beef (Vasconcelos et al. 2009). In addition to diet optimisation, management strategies can help minimise the loss of N from manure. For example, bedding pens with straw or sawdust and feeding less-digestible diets help increase the carbon : N ratio of manure (Erickson and Klopfenstein 2010). Cleaning pens more frequently, especially during summer months when volatilisation is greatest, helps reduce N losses (Erickson and Klopfenstein 2010). Given the tremendous implication for the environment, development of strategies to improve N-use efficiency of cattle and retention of N in manure are areas of research with potentially high rewards, and, therefore, require additional investment.
Estimating N requirement and supply
So as to achieve these reductions in feedlot N excretion and emission, greater precision is required in the estimation of N requirement and supply to finishing cattle. The 7th edition of the Nutrient Requirements of Beef Cattle (NRC 2000) moved from a CP approach to one of MP, which considered not just the N supplied in the diet (RDP and RUP), but also the true protein digested in the small intestine, accounting for both dietary RUP and microbial crude protein (MCP). There appears to be little application of the MP system by feedlot nutritionists in the time since the release of the 7th edition, and most US nutritionists are not considering RUP in formulating for protein, although many add urea (Samuelson et al. 2016c).
The 8th edition of the Nutrient Requirements of Beef Cattle (NASEM 2016) has made major changes to the beef cattle nutrient-requirement model (BCNRM) consideration of MP. Requirements for MP are unchanged, but there are significant modifications to the prediction of MP supply from the diet. Undegraded protein supply from the forage component of the diet has been reduced to account for the N content of completely undigested cell walls; this may affect estimates of N supply in receiving and step-up diets for finishing cattle, which may need additional RUP. MCP production from silages in step-up diets is less that than from hays because of the fermentation of non-structural carbohydrates during ensiling.
One important new consideration in BCNRM is the prediction of MCP synthesis (NASEM 2016). Where the model previously assumed a 13% efficiency of synthesis of MCP from total digestible nutrients (TDN) in concentrate diets, BCNRM adopts two equations for the prediction of MCP supply, depending on dietary ether extract (EE) content, and these reduce the estimate of MCP synthesis. Degradable protein is assumed to be converted to MCP with 100% efficiency. For diets containing <3.9% EE (this distinction was applied arbitrarily), MCP is now predicted by the following equation:
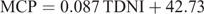
For diets containing EE ≥ 3.9%, MCP is predicted by
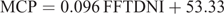
where FFTDNI is fat-free TDN intake (g/day).
The second equation is of relevance to finishing diets in Australia and the southern plains of the USA, which frequently contain up to 7% added fat on a DM basis.
Another important consideration in BCNRM is urea recycling. The unique ability of ruminants to recycle N to the gastrointestinal tract instead of excreting it to the environment is key to reducing dietary N while maintaining performance (Marini and Van Amburgh 2005). The analyses conducted in the 2016 revision (NASEM 2016) generated a quadratic relationship for the estimate of recycled urea available for anabolism on the basis of N intake, primarily from research using forage-based diets (Eisemann and Tedeschi 2016). Formerly, NRC (2000) estimated urea N recycling on the basis of relationships between ruminal NH3 and plasma urea N concentrations. In general, the new predictions of urea N used for anabolism (Eisemann and Tedeschi 2016; NASEM 2016) are lower than the predictions of the NRC (2000) across N intake. In feedlot finishing diets, RDP requirements are easily met, even before the increased estimate of N recycling is applied. However, NASEM (2016) identified that more complex modelling will be needed to integrate RDP supply with absorption of NH3 from the rumen, leading to an allowance for urea recycling in only the mechanistic level of solution of the model, and further research is required to better estimate urinary urea-N excretion for concentrate diets. Eisemann and Tedeschi (2016) speculated that the increase in rumen fermentation rates and products on concentrate diets would maximise the efficiency of MCP synthesis, and hence, the use, and perhaps transfer, of recycled N to the rumen for microbial anabolism. Overall, this new approach lowers the need for urea or RDP, which would decrease N excretion. Adoption of the updated MP system, formulated properly, could permit a significant reduction of the intake of protein in grain-finishing diets; however, to achieve this, greater confidence in its predictions and application on-farm using concentrate diets is needed.
Interaction of growth promotants with dietary N requirements
Growth promotants change the normal pattern of maturation, and the rate and composition of liveweight gain. These increase the deposition of lean muscle and, therefore, the requirements for MP and, so, interact with phase-feeding strategies. Hormonal growth-promotant implants (HGPs), such as oestrogens and androgens (e.g. trenbolone acetate), have been long used in the US, Canadian and Australian feedlot industries, and act to increase protein accretion by via IGF-1 and growth hormone-mediated cellular proliferation and reduced protein degradation (Dayton and White 2014). Galyean (1996) reviewed research on the interaction between HGPs and supplemental protein, and concluded that multiple use of high-potency (trenbolone acetate) implant programs increased the demand for protein, particularly in the early release phase of the implant, although CP concentrations could be reduced later in the feeding period without detriment. In Australia, where cattle enter a short finishing phase direct from a pasture-based grower period, many cattle undergo compensatory muscle gain during the early finishing period. The additional muscle accretion from HGPs may exacerbate demand for protein in the early finishing period in Australia, compared with heavier cattle in North America.
The rapid adoption of β-adrenergic agonist growth promoters (βAAs) during the last 20–45 days on feed in US and Canadian lot-feeding in the past decade has raised the question of whether there is an interaction of these with protein metabolism, similar to that observed for HGPs. The action of βAAs, including zilpaterol hydrochloride (ZH) and ractopamine hydrochloride (RH), to directly alter protein metabolism at a cellular level (Johnson et al. 2014; Walter et al. 2016) has been hypothesised to increase cattle requirements for dietary CP, or to reduce urea N available for recycling (Brake et al. 2011). Administration of ZH to cattle on maintenance diets decreased total and urinary N excretion without change in N intake, apparent digestion or faecal N excretion (Walter et al. 2016). Meanwhile, N-retention rate increased by 57% and skeletal muscle fractional protein-accretion rate increased by 299% in ZH-treated cattle, as a result of upregulation of protein synthesis, rather than downregulated degradation (Walter et al. 2016), which suggests that requirement for dietary N may be increased. When finishing diets were supplemented with protein in the form of DGs or urea (Brake et al. 2011), ZH increased DM and N intake, but still increased N retention (by 63% on an ad libitum-intake basis and by 36% as a proportion of N intake, across all diets), but with no effect on urea entry rate. It was concluded that the effect was likely to be mediated via reduction in urea production and recycling to the rumen (Brake et al. 2011). Recent studies have also demonstrated that a new β-agonist (Experior (lubabegron), Elanco Animal Health, Greenfield, IN, USA) can decrease NH3 emissions (Brown et al. 2019). These combined results may suggest N-retention as a mechanism for increased muscling, gain : feed ratio (G : F), carcass yield and dressing percentage in ZH-treated cattle (Scramlin et al. 2010; Lean et al. 2014; Samuelson et al. 2016a; Walter et al. 2016).
There is evidence that βAAs also act on the metabolism of N in the rumen, as well as in skeletal-muscle protein metabolism. Ruminal NH3 and amino acid concentrations decreased when cattle were fed RH, and when RH was added in vitro to rumen-fluid fermentation tubes along with casein, so that N was not limiting (Walker and Drouillard 2010). It appeared that RH reduced microbial proteolysis and deamination of dietary protein to NH3, by mechanisms unknown (Walker and Drouillard 2010).
Results such as these prompted the hypothesis that demand for dietary protein, and specifically NPN, may be increased under βAA administration (Walker and Drouillard 2010; NASEM 2016), as it is for the HGPs. However, large-scale performance trials (Hales et al. 2016; Samuelson et al. 2016a, 2016b) have found no interaction of the CP content or source (RDP or RUP) with administration of the βAAs RH or ZH. Daily gain, G : F and lean-tissue deposition were increased with RH, but not to an extent that protein-metabolism requirements were affected by the concentration of RUP (6.1–10.4%) or RDP (7.3–13.4%; Samuelson et al. 2016b). No interactions between ZH and dietary urea concentration were found for feedlot performance or carcass characteristics (Samuelson et al. 2016a). Increasing RDP, while maintaining CP at 13.7% and decreasing dietary MP supply, did not affect cattle performance (Walker et al. 2006). Supplementation of RH-treated feedlot cattle with soybean meal, thus increasing dietary CP from 13.5% to 17.5% (Hales et al. 2016), and from 12.5 to 14.7% (Cônsolo et al. 2016), has not been found to affect animal performance or carcass characteristics. We, therefore, conclude that there is insufficient evidence to support an increase in dietary MP requirement in (at least corn-fed) cattle treated with βAAs; however, further clarification may come from systemic studies testing MP requirements (NASEM 2016).
The additional protein for skeletal muscle-tissue accretion under the effects of the βAAs appears to not come from the diet; thus, it has been hypothesised that N is repartitioned from the liver and gastrointestinal tissues (Montgomery et al. 2009; Holland et al. 2010). Walter et al. (2018) reported decreased total splanchnic tissue mass under the effects of ZH that was not caused by a reduction in DM intake (DMI) or an improvement in total tract digestibility. Reduced N deposition in non-skeletal muscle tissue has also been reported in pigs treated with RH (Dunshea et al. 1998). This would reduce the maintenance protein requirements of the splanchnic tissues, resulting in decreased maintenance energy requirements and decreased protein turnover (due to smaller mass). Walter et al. (2018) proposed that this may explain the reduced urea entry rate observed with ZH by Brake et al. (2011), which could also have a circular effect by further limiting the nutrition, and, therefore, size, of the splanchnic tissues.
Animal costs of excess protein
Meeting RDP requirements promotes rumen microbial fermentation, DMI and nutrient flow from the rumen. Conventional wisdom is that provision of RDP in excess of microbial protein-synthesis requirement has no value, being absorbed into plasma NH3, converted into urea and excreted in the urine (NRC 2000; Erickson et al. 2016; Klatt et al. 2018). However, there may be an energetic cost of ureagenesis for the body to detoxify this excess RDP-derived NH3 (Lobley et al. 1995), and the N excreted in faeces and urine has a significant impact on the environment. In the body, excess N is converted to urea in the liver and then excreted in the urine. Jennings et al. (2018) conducted two experiments to determine the effects of feeding excess CP and MP (using corn gluten feed and corn gluten meal) on the maintenance-energy requirements of steers fed a finishing diet at 1 or 2 × maintenance-energy intake. Maintenance-energy requirements, estimated from linear and quadratic regression of energy retention on metabolisable-energy intake, were 4–6% greater for steers overfed CP (19.5% CP with 14.14% MP) than for control steers receiving 13.8% CP with 9.63% MP. Jennings et al. (2018) suggested that feeding excess CP and MP from a protein source that is high in RDP and low in protein quality will increase maintenance-energy requirements of finishing steers.
Benefits of high protein inclusion
There are some benefits of high concentrations of dietary CP beyond covering among-animal variation in requirements. Immune responses to inflammation decrease N retention and increase plasma concentrations of essential and non-essential amino acids, suggesting that immunocompromised cattle have an increased metabolic amino acid demand, at a time when DMI may be depressed (Waggoner et al. 2009). Supplying CP at high inclusion rates (16%) reduced the N-retention depression in response to a lipopolysaccharide challenge. However, the relationship between dietary N and morbidity in receival cattle is unclear and requires further research (Galyean et al. 1999).
Trenkle (1979) proposed that blood NH3 served as a systemic buffer in cattle fed high-concentrate diets. Owens et al. (2014) proposed a role for high NH3 concentrations in cattle fed high-energy diets, on the basis of the properties of urea and NH3 as weak acids and osmolytes. Ammonia often functions as a weak base, and, therefore, has some buffering properties when present in the rumen, which may be valuable in energy-dense diets based on highly fermentable grains (Scott 1972; Heitmann and Bergman 1978; Trenkle 1979; Zinn et al. 2003). When absorbed across the rumen wall and converted to urea, the consequent increase in blood urea N will act to increase blood osmolality. This can lead to increased saliva production and flow to the rumen, improving the rumen buffering capacity, and increasing rumen dilution rate (Owens et al. 2014). Blood urea N cycled to the large intestine increases the postruminal and faecal pH (Owens et al. 2014). It is speculated that this could increase compensatory digestion of organic matter at the large intestine (Owens et al. 2014). Overfeeding N as a buffer in the rumen may not be acceptable in the future because of the unintended consequences of excess N excretion. Optimising diets by incorporating physically effective fibre and grain processing to reduce fines and rate of fermentability to control rumen acidosis might be better approaches in the future.
Recent work by researchers at Kansas State University has suggested that increasing post-ruminal flows of protein and amino acids can improve enzymatic small-intestinal starch digestion (SISD). Zinn et al. (2002) reported that infusing casein into the abomasum increased starch digestion in the small intestine. Although, under normal circumstances, only a small proportion of dietary starch is digested post-ruminally, SISD to glucose yields 42% more energetic value per gram than does starch fermented to volatile fatty acids in the rumen (Owens et al. 1986), and, so, there are considerable efficiency benefits to be gained from moving starch digestion from the rumen to the small intestine. It has been proposed that pancreatic α-amylase secretion is a limiting constraint to SISD, and that this can be increased with greater post-ruminal flows of high-quality protein. Duodenal infusion of casein and casein-like non-essential amino acids increased pancreatic α-amylase secretion linearly, and increased SISD, by, on average, 0.31 for each gram of duodenally infused casein (Brake et al. 2014a, 2014b; Blom et al. 2016). Rumen protection of non-essential amino acids and glutamate may, therefore, increase SISD in feedlot cattle fed corn-based diets. This increased amino acid flow to the small intestine, and subsequent increase in SISD (and, therefore, energy availability) can lead to increased N balance, N retention and, therefore, lean-tissue accretion (Blom et al. 2016). Shifting the site of starch digestion is more easily achieved in corn diets than barley or wheat diets because of the very low ruminal escape starch in the latter, even with minimal processing (Theurer 1986).
Potential strategies for better meeting N requirements with supply
The perceived low cost, with few apparent negative effects on animal performance, promotes the feeding of N in surplus. However, when we account for the environmental costs of excreted surplus N on emissions of NH3, and increasing costs of high RUP feeds, then pursuing a reduction in N intake assumes new importance. Potential strategies for reducing total N to finishing cattle include better synchronisation of N and energy supply, intermittent supply of dietary CP, and reducing supply of CP to better meet cattle requirements, according to the stage of growth (phase feeding).
Synchronisation of nutrients
It is suggested that synchronisation of the ruminal degradation rate of carbohydrates and CP could increase ruminal microbial protein synthesis, improve N-use efficiency, decrease urinary N excretion, and, subsequently, improve animal performance. While several studies have used the synchrony index of Sinclair et al. (1993) and other methods in an attempt to ‘synchronise’ the fermentation rates of dietary organic matter (OM) and CP within the rumen, the results have been equivocal (Herrera-Saldana et al. 1990; Robinson and McQueen 1994; Shabi et al. 1998; Dewhurst et al. 2000; Valkeners et al. 2004, 2006; Rotger et al. 2006).
All published studies of nutrient synchrony have been conducted with moderate- to high-roughage diets and, generally, in animals fed once per day. In a study that used high-concentrate diets, Rotger et al. (2006) noted no effect of synchrony on ruminal NH3 or volatile fatty acid concentrations, or on diet digestibility. Valkeners et al. (2004, 2006) modified ruminal N–OM synchrony by changing the pattern of feeding diets that were either low or high in RDP and noted no effect of dietary synchrony on N retention, ruminal MCP synthesis, or ruminal microbial efficiency (g microbial protein/kg OM fermented). Cole and Todd (2008) calculated the synchrony index of diets fed in nine performance and seven metabolism studies that used various grain-processing methods and protein sources. Results of that analysis suggested that N–carbohydrate synchronisation was of minimal importance in ruminants fed complete diets and had little or no effect on average daily gain, duodenal N flow, or MCP synthesis. In contrast to the synchronisation theory, the synchrony index was negatively correlated with G : F and microbial protein-production efficiency (g microbial protein/kg digested OM). Thus, the limited data available with moderate- to high-concentrate diets suggest that ruminal synchrony has little, if any, effect on ruminal or whole-animal metabolism.
Cole and Todd (2008) and Reynolds and Kristensen (2008) proposed several reasons for the lack of importance of N–carbohydrate synchrony in finishing cattle. First, feedlot cattle tend to eat small meals frequently within a diurnal feeding period. Thus, microbial fermentation in the rumen comprises N and carbohydrate from several different meals. Second, N recycling from the lower gut to the rumen (and vice versa) may help synchronise N and carbohydrate availability.
Synchronised oscillations: oscillating dietary CP
In contrast to the synchrony theory, it has been demonstrated that cattle fed protein-deficient forages can be supplemented with natural protein sources at 48-h intervals without adversely affecting animal performance compared with 24-hourly protein supplementation (Collins and Pritchard 1992). Cole (1999) hypothesised that if sufficient fermentable carbohydrate is available within the ruminal contents, N utilisation could be improved via increased or optimised N recycling to the rumen in grain-fed animals. He theorised that by oscillating dietary CP at a rate similar to the rate of digesta passage, the overall quantity of CP fed could be decreased without adversely affecting animal performance. The objective was to ‘synchronise’ a ruminal N deficiency with a N excess in the lower gut, and vice versa, by oscillating the dietary CP percentages between deficient and adequate or excessive concentrations, thereby optimising N recycling.
Reported responses to oscillating dietary CP concentrations are summarised in Table 2. In initial studies (Cole 1999), oscillating dietary CP concentrations of sheep at 24-h intervals did not affect N retention; however, oscillating at 48-h intervals, a value closer to the estimated rate of passage was effective. In all subsequent studies, N retention has been increased (Table 2). In agreement with the theory of Cole (1999), Archibeque et al. (2007b) noted that oscillating dietary CP tended (P < 0.06) to increase uptake of urea by the portal drained viscera (i.e. N recycling).

|
Oscillating CP appears to improve N metabolism, in part, by an increase in ruminal MCP production (Kiran and Mutsvangwa 2009; Doranalli et al. 2011). Archibeque et al. (2007b), Kiran and Mutsvangwa (2009) and Doranalli et al. (2011) noted that oscillating dietary CP increased urea recycling to the rumen. Cole (1999) theorised that, for an oscillating CP regimen to increase N recycling to the rumen, compared with a static CP concentration, one of the oscillated diets would need to be deficient in RDP, and the average CP or RDP concentration would have to be near, or below, animal requirements. Capture of recycled N in MCP should be greater with more fermentable, grain-based high-concentrate diets than with high-forage diets. Shawn Archibeque at Colorado State University (unpubl. data) fed finishing steers a 10% or 13.5% CP diet or oscillated protein by deleting urea from the diets for 2 days, of each four days. By oscillating CP, the total N intake was decreased by 7 kg, and there was no effect on animal performance or carcass characteristics. However, with irregular (3 and 4 days of 7) oscillations between 13.5% and 11.6% CP, Galles et al. (2011) could not achieve the reduction in NH3 emissions from a 13.5% control diet that were recorded on a reduced, 11.6% CP diet. Crude protein oscillation has several challenges in practice, such as, for one, increased complexity of ration manufacture and feeding management.
Phase-feeding protein
Typically, feed-yard diets change little during a feeding period. Because N retention, and, thus, protein requirements, decrease as animals mature, dietary CP concentrations may exceed cattle requirements late in the feeding period. As days on feed increases, DMI and, therefore, N intake increase. As time on feed increases, urinary urea N excretion increases for the same N intake (Vasconcelos et al. 2009). However, apparent N absorption (% N intake) changes little, and N retention (g/day) decreases (Vasconcelos et al. 2009), suggesting that there is an opportunity to reduce N supply later in the feeding period.
Although common in the swine and poultry industries, changing nutrient concentrations in the diet to more accurately meet animal requirements (referred to as phase-feeding) has rarely been used in cattle feeding. Studies with dry-rolled corn-based diets (Erickson et al. 1999; Cooper et al. 2000; Trenkle 2002) have demonstrated that dietary CP concentrations can be substantially decreased late (i.e. last 28–56 days) in the feeding period, with little or no effect on cattle performance. By reducing the CP content of steam-flaked corn (SFC)-based diets from 13.0% to 11.5% or 10.0% either in the last 47 days on feed, or after mean pen liveweight reached 477 kg, Vasconcelos et al. (2006) were able to reduce N wastage and excretion, without any reduction in rate of gain or G : F. However, in a larger study with SFC-based diets, the phase-feeding of protein has appeared less promising (Cole et al. 2006). Cole et al. (2006) noted that average daily gain (ADG), DMI, and G : F during the last 56 days on feed were similar in steers continuously fed an 11.5% CP diet and steers continuously fed a 13.0% CP diet. However, steers changed from the 13.0% CP diet to the 11.5% CP diet with 56 days left on feed had lower (P < 0.05) ADG and DMI than those continuously fed the 13% CP diet, suggesting that an adjustment period may be required when changing diets late in the feeding period. Although Cole et al. (2006) noted depressed performance (ADG and G : F) in phase-fed cattle during the last 56 days on feed, they also noted that phase-feeding decreased N intake by 1.4–3.8 kg/steer, decreased N excretion by 0.1–2.9 kg/steer, and decreased estimated N volatilisation by ~4.4 kg/steer, or 22% compared with continuous feeding of the 13.0% CP diet. Similar decreases in N excretion (Cooper et al. 2000; Vasconcelos et al. 2009) and estimated N volatilisation losses (Erickson et al. 1999; Cole et al. 2005; Todd et al. 2006) have been reported in beef cattle when dietary CP concentrations are decreased. Cole et al. (2006) also noted that decreasing dietary CP during the last 28 days on feed (vs the last 56 days) significantly decreased animal performance. Differences in results of phase-feeding between studies using dry-rolled corn- and SFC-based diets are likely to be attributable to differences in RDP requirements (Cooper et al. 2002a, 2002b; Gleghorn et al. 2004). Differences in the results of the studies of Vasconcelos et al. (2006) and Cole et al. (2006) when SFC-based diets were fed may be the result of differences in implanting strategies.
Cattle arriving in a low body condition (rib cage visible) have shrunken liver and digestive tract (the splanchnic tissues; Ryan et al. 1993). These tissues require significant protein accretion to regain their weight relative to bodyweight during realimentation (Ryan et al. 1993). Therefore, in such cattle, the maintenance energy requirement is decreased, leaving more energy for liveweight gain, but the availability of intake protein for skeletal muscle deposition may be restricted as it is redirected to growth of the splanchnic tissues to accommodate high DMI. The splanchnic tissues are not only a significant deposit of protein mass in the body, but are also a location of high protein turnover. Further research on timing of phase feeding on such cattle is required.
If a β-agonist is fed during the last 20–45 days on feed, it would be feasible to apply phase-feeding reduction in dietary CP at the same time. However, further research is needed to clarify how phase-feeding of CP could be used with β-agonists late in the feeding period. In addition, although the focus of N phase feeding research has been on reducing N supply at the end of the finishing period, recent changes in estimates of RDP supply from forages in the BCNRM may be a cause for a reassessment of the application of the MP system in step-up diets.
Conclusions
Currently, N formulation of grain finishing diets for cattle is based on conservative estimation of minimum CP requirements, resulting in overfeeding of N as a perceived low-risk, low-cost strategy. However, the high CP content of finishing diets, particularly due to the use of low-cost DG by-products in the USA, not only has measurable effects on energy requirements to excrete the excess N, but also increases N output into the environment. The future is likely to present new challenges to N feeding in feedlots, as increasing cost of land and commodities will increase the cost of dietary N. Increasing pressure is also being placed on cattle producers to monitor and reduce N emissions and pollution.
Recent research on practical feeding strategies to decrease N supply has produced mixed results. Reducing dietary N in the late finishing period is one of the most promising and adoptable strategies to improve N efficiency in grain-finishing cattle. The recent adoption of βAAs in North America does not appear to have increased N requirements, but may have reduced N excretion rates.
Changes in the BCNRM 8th edition are an initial step towards better prediction of the MP supply. There has been little adoption so far of the MP system to improve precision of dietary N formulation in grain-finishing diets. This suggests that researchers need to provide greater confidence for industry in the ability of the MP system to predict and refine formulations for MCP, RDP and RUP in grain-finishing diets, improving precision and reducing waste. Options to reduce total N intake and N wastage are available, but given that the current balance of risk and cost steer feedlots towards over-feeding of N, it is possible that only a regulatory or market-mechanism that targets N emissions and pollution, thus changing this equation, will cause a reduction in N overuse.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
The authors thank the reviewers who provided feedback and suggestions on this manuscript. This research did not receive any specific funding.
References
Angelidis A, Crompton L, Misselbrook T, Yan T, Reynolds CK, Stergiadis S (2019) Evaluation and prediction of nitrogen use efficiency and outputs in faeces and urine in beef cattle. Agriculture, Ecosystems & Environment 280, 1–15.| Evaluation and prediction of nitrogen use efficiency and outputs in faeces and urine in beef cattle.Crossref | GoogleScholarGoogle Scholar |
Archibeque S, Freetly H, Cole N, Ferrell C (2007a) The influence of oscillating dietary protein concentrations on finishing cattle. II. Nutrient retention and ammonia emissions. Journal of Animal Science 85, 1496–1503.
| The influence of oscillating dietary protein concentrations on finishing cattle. II. Nutrient retention and ammonia emissions.Crossref | GoogleScholarGoogle Scholar | 17264236PubMed |
Archibeque S, Freetly H, Ferrell C (2007b) Net portal and hepatic flux of nutrients in growing wethers fed high-concentrate diets with oscillating protein concentrations. Journal of Animal Science 85, 997–1005.
| Net portal and hepatic flux of nutrients in growing wethers fed high-concentrate diets with oscillating protein concentrations.Crossref | GoogleScholarGoogle Scholar | 17145976PubMed |
Baber JR, Sawyer JE, Wickersham TA (2018) Estimation of human-edible protein conversion efficiency, net protein contribution, and enteric methane production from beef production in the United States. Translational Animal Science 2, 439–450.
| Estimation of human-edible protein conversion efficiency, net protein contribution, and enteric methane production from beef production in the United States.Crossref | GoogleScholarGoogle Scholar |
Bai M, Flesch TK, McGinn SM, Chen D (2015) A snapshot of greenhouse gas emissions from a cattle feedlot. Journal of Environmental Quality 44, 1974–1978.
| A snapshot of greenhouse gas emissions from a cattle feedlot.Crossref | GoogleScholarGoogle Scholar | 26641350PubMed |
Beusen AHW, Bouwman AF, Heuberger PSC, Van Drecht G, Van Der Hoek KW (2008) Bottom-up uncertainty estimates of global ammonia emissions from global agricultural production systems. Atmospheric Environment 42, 6067–6077.
| Bottom-up uncertainty estimates of global ammonia emissions from global agricultural production systems.Crossref | GoogleScholarGoogle Scholar |
Blom EJ, Brake DW, Anderson DE (2016) Increases in duodenal glutamic acid supply linearly increase small intestinal starch digestion but not nitrogen balance in cattle. Journal of Animal Science 94, 5332–5340.
| Increases in duodenal glutamic acid supply linearly increase small intestinal starch digestion but not nitrogen balance in cattle.Crossref | GoogleScholarGoogle Scholar | 28046181PubMed |
Brake DW, Titgemeyer EC, Jones ML (2011) Effect of nitrogen supplementation and zilpaterol–HCl on urea kinetics in steers consuming corn-based diets. Journal of Animal Physiology and Animal Nutrition 95, 409–416.
| Effect of nitrogen supplementation and zilpaterol–HCl on urea kinetics in steers consuming corn-based diets.Crossref | GoogleScholarGoogle Scholar | 21039927PubMed |
Brake DW, Bailey EA, Titgemeyer EC, Anderson DE (2014a) Small intestinal digestion of raw cornstarch in cattle consuming a soybean hull-based diet is improved by duodenal casein infusion. Journal of Animal Science 92, 4047–4056.
| Small intestinal digestion of raw cornstarch in cattle consuming a soybean hull-based diet is improved by duodenal casein infusion.Crossref | GoogleScholarGoogle Scholar | 25023803PubMed |
Brake DW, Titgemeyer EC, Anderson DE (2014b) Duodenal supply of glutamate and casein both improve intestinal starch digestion in cattle but by apparently different mechanisms. Journal of Animal Science 92, 4057–4067.
| Duodenal supply of glutamate and casein both improve intestinal starch digestion in cattle but by apparently different mechanisms.Crossref | GoogleScholarGoogle Scholar | 25057031PubMed |
Brown MS, Cole NA, Gruber S, Kube J, Teeter JS (2019) Modeling and prediction accuracy of ammonia gas emissions from feedlot cattle. Applied Animal Science 35, 347–356.
| Modeling and prediction accuracy of ammonia gas emissions from feedlot cattle.Crossref | GoogleScholarGoogle Scholar |
Cole NA (1999) Nitrogen retention by lambs fed oscillating dietary protein concentrations. Journal of Animal Science 77, 215–222.
| Nitrogen retention by lambs fed oscillating dietary protein concentrations.Crossref | GoogleScholarGoogle Scholar | 10064047PubMed |
Cole NA, Todd RW (2008) Opportunities to enhance performance and efficiency through nutrient synchrony in concentrate-fed ruminants. Journal of Animal Science 86, E318–E333.
| Opportunities to enhance performance and efficiency through nutrient synchrony in concentrate-fed ruminants.Crossref | GoogleScholarGoogle Scholar | 17940155PubMed |
Cole NA, Greene LW, McCollum FT, Montgomery TH, McBride KW (2003) Influence of oscillating dietary crude protein concentration on performance, acid-base balance, and nitrogen excretion of steers. Journal of Animal Science 81, 2660–2668.
| Influence of oscillating dietary crude protein concentration on performance, acid-base balance, and nitrogen excretion of steers.Crossref | GoogleScholarGoogle Scholar | 14601868PubMed |
Cole NA, Clark RN, Todd RW, Richardson CR, Gueye A, Greene LW, Mcbride K (2005) Influence of dietary crude protein concentration and source on potential ammonia emissions from beef cattle manure. Journal of Animal Science 83, 722–731.
| Influence of dietary crude protein concentration and source on potential ammonia emissions from beef cattle manure.Crossref | GoogleScholarGoogle Scholar | 15705770PubMed |
Cole NA, Defoor PJ, Galyean ML, Duff GC, Gleghorn JF (2006) Effects of phase-feeding of crude protein on performance, carcass characteristics, serum urea nitrogen concentrations, and manure nitrogen of finishing beef steers. Journal of Animal Science 84, 3421–3432.
| Effects of phase-feeding of crude protein on performance, carcass characteristics, serum urea nitrogen concentrations, and manure nitrogen of finishing beef steers.Crossref | GoogleScholarGoogle Scholar | 17093237PubMed |
Collins RM, Pritchard RH (1992) Alternate day supplementation of corn stalk diets with soybean meal or corn gluten meal fed to ruminants. Journal of Animal Science 70, 3899–3908.
| Alternate day supplementation of corn stalk diets with soybean meal or corn gluten meal fed to ruminants.Crossref | GoogleScholarGoogle Scholar | 1474027PubMed |
Cônsolo NRB, Mesquita BS, Rodriguez FD, Rizzi VG, Silva LFP (2016) Effects of ractopamine hydrochloride and dietary protein content on performance, carcass traits and meat quality of Nellore bulls. Animal 10, 539–546.
| Effects of ractopamine hydrochloride and dietary protein content on performance, carcass traits and meat quality of Nellore bulls.Crossref | GoogleScholarGoogle Scholar |
Cooper R, Milton T, Klopfenstein TJ (2000) Phase-feeding metabolizable protein for finishing steers. Nebraska Beef Cattle Reports 366. Available at http://digitalcommons.unl.edu/animalscinbcr/366 [Verified 1 May 2019]
Cooper RJ, Milton C, Klopfenstein T, Jordon D (2002a) Effect of corn processing on degradable intake protein requirement of finishing cattle. Journal of Animal Science 80, 242–247.
| Effect of corn processing on degradable intake protein requirement of finishing cattle.Crossref | GoogleScholarGoogle Scholar | 11831524PubMed |
Cooper RJ, Milton CT, Klopfenstein TJ, Scott TL, Wilson CB, Mass RA (2002b) Effect of corn processing on starch digestion and bacterial crude protein flow in finishing cattle. Journal of Animal Science 80, 797–804.
| Effect of corn processing on starch digestion and bacterial crude protein flow in finishing cattle.Crossref | GoogleScholarGoogle Scholar | 11890417PubMed |
Croft IA, Clayton EH, Cusack PMV (2014) Health and production of feedlot cattle following supplementation with urea–molasses in starter pens. Australian Veterinary Journal 92, 166–170.
| Health and production of feedlot cattle following supplementation with urea–molasses in starter pens.Crossref | GoogleScholarGoogle Scholar | 24766047PubMed |
Dayton WR, White ME (2014) Meat science and muscle biology symposium: role of satellite cells in anabolic steroid-induced muscle growth in feedlot steers. Journal of Animal Science 92, 30–38.
| Meat science and muscle biology symposium: role of satellite cells in anabolic steroid-induced muscle growth in feedlot steers.Crossref | GoogleScholarGoogle Scholar | 24166993PubMed |
Dewhurst RJ, Davies DR, Merry RJ (2000) Microbial protein supply from the rumen. Animal Feed Science and Technology 85, 1–21.
| Microbial protein supply from the rumen.Crossref | GoogleScholarGoogle Scholar |
Dong Rl, Zhao G, Chai Ll, Beauchemin KA (2014) Prediction of urinary and fecal nitrogen excretion by beef cattle. Journal of Animal Science 92, 4669–4681.
| Prediction of urinary and fecal nitrogen excretion by beef cattle.Crossref | GoogleScholarGoogle Scholar | 25149338PubMed |
Doranalli K, Penner GB, Mutsvangwa T (2011) Feeding oscillating dietary crude protein concentrations increases nitrogen utilization in growing lambs and this response is partly attributable to increased urea transfer to the rumen. The Journal of Nutrition 141, 560–567.
| Feeding oscillating dietary crude protein concentrations increases nitrogen utilization in growing lambs and this response is partly attributable to increased urea transfer to the rumen.Crossref | GoogleScholarGoogle Scholar | 21310865PubMed |
Drouillard JS (2018) Current situation and future trends for beef production in the United States of America: a review. Asian-Australasian Journal of Animal Sciences 31, 1007–1016.
| Current situation and future trends for beef production in the United States of America: a review.Crossref | GoogleScholarGoogle Scholar | 29973030PubMed |
Dunshea F, Eason P, Campbell R, King R (1998) Interrelationships between dietary ractopamine, energy intake, and sex in pigs. Australian Journal of Agricultural Research 49, 565–574.
| Interrelationships between dietary ractopamine, energy intake, and sex in pigs.Crossref | GoogleScholarGoogle Scholar |
Eisemann J, Tedeschi L (2016) Predicting the amount of urea nitrogen recycled and used for anabolism in growing cattle. The Journal of Agricultural Science 154, 1118–1129.
| Predicting the amount of urea nitrogen recycled and used for anabolism in growing cattle.Crossref | GoogleScholarGoogle Scholar |
Erickson G, Klopfenstein T (2010) Nutritional and management methods to decrease nitrogen losses from beef feedlots. Journal of Animal Science 88, E172–E180.
| Nutritional and management methods to decrease nitrogen losses from beef feedlots.Crossref | GoogleScholarGoogle Scholar | 20081072PubMed |
Erickson, GE, Klopfenstein, TJ, Milton, T, Herold, D (1999) Effects of matching protein to requirements on performance and waste management in the feedlot. Nebraska Beef Cattle Reports 402. Available at http://digitalcommons.unl.edu/animalscinbcr/402 [Verified 1 May 2019]
Erickson GE, Watson A, MacDonald J, Klopfenstein T (2016) Protein nutrition evaluation and application to growing and finishing cattle. In ‘27th annual Florida ruminant nutrition symposium’, 15–17 February 2016, Gainesville, Florida. pp. 91–102. (Department of Animal Sciences, University of Florida Institute of Food and Agricultural Sciences: Gainesville, FL) Available at http://dairy.ifas.ufl.edu/rns/2016/Proceedings%20FL%20RNS%202016.pdf [Verified 2 August 2019]
Flesch TK, Wilson JD, Harper LA, Todd RW, Cole NA (2007) Determining ammonia emissions from a cattle feedlot with an inverse dispersion technique. Agricultural and Forest Meteorology 144, 139–155.
| Determining ammonia emissions from a cattle feedlot with an inverse dispersion technique.Crossref | GoogleScholarGoogle Scholar |
Galles K, Ham J, Westover E, Stratton J, Wagner J, Engle T, Bryant TC (2011) Influence of reduced nitrogen diets on ammonia emissions from cattle feedlot pens. Atmosphere 2, 655–670.
| Influence of reduced nitrogen diets on ammonia emissions from cattle feedlot pens.Crossref | GoogleScholarGoogle Scholar |
Galyean ML (1996) Protein levels in beef cattle finishing diets: industry application, university research, and systems results1. Journal of Animal Science 74, 2860–2870.
| Protein levels in beef cattle finishing diets: industry application, university research, and systems results1.Crossref | GoogleScholarGoogle Scholar | 8923202PubMed |
Galyean ML, Perino LJ, Duff GC (1999) Interaction of cattle health/immunity and nutrition. Journal of Animal Science 77, 1120–1134.
| Interaction of cattle health/immunity and nutrition.Crossref | GoogleScholarGoogle Scholar | 10340578PubMed |
Gaughan JB, Bonner S, Loxton I, Mader TL, Lisle A, Lawrence R (2010) Effect of shade on body temperature and performance of feedlot steers1. Journal of Animal Science 88, 4056–4067.
| Effect of shade on body temperature and performance of feedlot steers1.Crossref | GoogleScholarGoogle Scholar | 20709874PubMed |
Gleghorn JF, Elam NA, Galyean ML, Duff GC, Cole NA, Rivera JD (2004) Effects of crude protein concentration and degradability on performance, carcass characteristics, and serum urea nitrogen concentrations in finishing beef steers. Journal of Animal Science 82, 2705–2717.
| Effects of crude protein concentration and degradability on performance, carcass characteristics, and serum urea nitrogen concentrations in finishing beef steers.Crossref | GoogleScholarGoogle Scholar | 15446487PubMed |
Goddard A, Gardner GE, McGilchrist P (2015) The effect of supplementing beef cattle with crude glycerol on the pH of the M. Longissimus thoracis. In ‘61st international congresses of meat science and technology’, Clermont-Ferrand, France, 23–28 August 2015. pp. 5–38. Available at http://icomst-proceedings.helsinki.fi/papers/2015_05_38.pdf [Verified 2 August 2019]
Groenestein CM, Hutchings NJ, Haenel HD, Amon B, Menzi H, Mikkelsen MH, Misselbrook TH, van Bruggen C, Kupper T, Webb J (2016) How do NH3 emissions relate to nitrogen use efficiency of livestock production? In ‘Proceedings of the 2016 international nitrogen initiative conference. Solutions to improve nitrogen use efficiency for the world’, 4–8 December 2016, Melbourne, Australia. Available at www.ini2016.com [Verified 2 August 2019]
Hales KE, Cole NA, MacDonald JC (2012) Effects of corn processing method and dietary inclusion of wet distillers grains with solubles on energy metabolism, carbon− nitrogen balance, and methane emissions of cattle. Journal of Animal Science 90, 3174–3185.
| Effects of corn processing method and dietary inclusion of wet distillers grains with solubles on energy metabolism, carbon− nitrogen balance, and methane emissions of cattle.Crossref | GoogleScholarGoogle Scholar | 22585790PubMed |
Hales K, Shackelford S, Wells J, King D, Pyatt N, Freetly H, Wheeler T (2016) Effects of dietary protein concentration and ractopamine hydrochloride on performance and carcass characteristics of finishing beef steers. Journal of Animal Science 94, 2097–2102.
| Effects of dietary protein concentration and ractopamine hydrochloride on performance and carcass characteristics of finishing beef steers.Crossref | GoogleScholarGoogle Scholar | 27285706PubMed |
Heitmann R, Bergman E (1978) Glutamine metabolism, interorgan transport and glucogenicity in the sheep. American Journal of Physiology. Endocrinology and Metabolism 234, E197
| Glutamine metabolism, interorgan transport and glucogenicity in the sheep.Crossref | GoogleScholarGoogle Scholar |
Herrera-Saldana R, Gomez-Alarcon R, Torabi M, Huber J (1990) Influence of synchronizing protein and starch degradation in the rumen on nutrient utilization and microbial protein synthesis. Journal of Dairy Science 73, 142–148.
| Influence of synchronizing protein and starch degradation in the rumen on nutrient utilization and microbial protein synthesis.Crossref | GoogleScholarGoogle Scholar | 2312880PubMed |
Holland BP, VanOverbeke DL, Stein DR, Hilton GG, Shook JN, Burciaga-Robles LO, Krehbiel CR, Yates DA, Montgomery JL, Hutcheson JP, Streeter MN, Nichols WT, Step DL (2010) Effect of extended withdrawal of zilpaterol hydrochloride on performance and carcass traits in finishing beef steers1. Journal of Animal Science 88, 338–348.
| Effect of extended withdrawal of zilpaterol hydrochloride on performance and carcass traits in finishing beef steers1.Crossref | GoogleScholarGoogle Scholar | 19749012PubMed |
Jennings JS, Meyer BE, Cole NA, Guiroy PJ (2018) Energy costs of feeding excess protein from corn-based by-products to finishing cattle. Journal of Animal Science 96, 653–669.
| Energy costs of feeding excess protein from corn-based by-products to finishing cattle.Crossref | GoogleScholarGoogle Scholar | 29390094PubMed |
Johnson BJ, Smith SB, Chung KY (2014) Historical overview of the effect of β-adrenergic agonists on beef cattle production. Asian-Australasian Journal of Animal Sciences 27, 757–766.
| Historical overview of the effect of β-adrenergic agonists on beef cattle production.Crossref | GoogleScholarGoogle Scholar | 25050012PubMed |
Jones SJ, Starkey D, Calkins CR, Crouse J (1990) Myofibrillar protein turnover in feed-restricted and realimented beef cattle. Journal of Animal Science 68, 2707–2715.
| Myofibrillar protein turnover in feed-restricted and realimented beef cattle.Crossref | GoogleScholarGoogle Scholar | 2211400PubMed |
Kiran D, Mutsvangwa T (2009) Nitrogen utilization in growing lambs fed oscillating dietary protein concentrations. Animal Feed Science and Technology 152, 33–41.
| Nitrogen utilization in growing lambs fed oscillating dietary protein concentrations.Crossref | GoogleScholarGoogle Scholar |
Klatt BJ, Harsh BN, McCann JC (2018) Determining rumen degradable protein requirements in growing beef cattle. Journal of Animal Science 96, 73–74.
| Determining rumen degradable protein requirements in growing beef cattle.Crossref | GoogleScholarGoogle Scholar |
Klieve AV, McLennan SR, Ouwerkerk D (2012) Persistence of orally administered Megasphaera elsdenii and Ruminococcus bromii in the rumen of beef cattle fed a high grain (barley) diet. Animal Production Science 52, 297–304.
| Persistence of orally administered Megasphaera elsdenii and Ruminococcus bromii in the rumen of beef cattle fed a high grain (barley) diet.Crossref | GoogleScholarGoogle Scholar |
Koenig KM, Beauchemin KA (2013) Nitrogen metabolism and route of excretion in beef feedlot cattle fed barley-based finishing diets varying in protein concentration and rumen degradability. Journal of Animal Science 91, 2310–2320.
| Nitrogen metabolism and route of excretion in beef feedlot cattle fed barley-based finishing diets varying in protein concentration and rumen degradability.Crossref | GoogleScholarGoogle Scholar | 23478813PubMed |
Koenig KM, Beauchemin KA (2018) Effect of feeding condensed tannins in high protein finishing diets containing corn distillers grains on ruminal fermentation, nutrient digestibility, and route of nitrogen excretion in beef cattle. Journal of Animal Science 96, 4398–4413.
| Effect of feeding condensed tannins in high protein finishing diets containing corn distillers grains on ruminal fermentation, nutrient digestibility, and route of nitrogen excretion in beef cattle.Crossref | GoogleScholarGoogle Scholar | 30032241PubMed |
Koenig K, McGinn S, Beauchemin K (2013) Ammonia emissions and performance of backgrounding and finishing beef feedlot cattle fed barley-based diets varying in dietary crude protein concentration and rumen degradability. Journal of Animal Science 91, 2278–2294.
| Ammonia emissions and performance of backgrounding and finishing beef feedlot cattle fed barley-based diets varying in dietary crude protein concentration and rumen degradability.Crossref | GoogleScholarGoogle Scholar | 23478824PubMed |
Koenig KM, Beauchemin KA, McGinn SM (2018) Feeding condensed tannins to mitigate ammonia emissions from beef feedlot cattle fed high-protein finishing diets containing distillers grains. Journal of Animal Science 96, 4414–4430.
| Feeding condensed tannins to mitigate ammonia emissions from beef feedlot cattle fed high-protein finishing diets containing distillers grains.Crossref | GoogleScholarGoogle Scholar | 30032212PubMed |
Lean IJ, Thompson JM, Dunshea FR (2014) A meta-analysis of zilpaterol and ractopamine effects on feedlot performance, carcass traits and shear strength of meat in cattle. PLoS One 9, e115904
| A meta-analysis of zilpaterol and ractopamine effects on feedlot performance, carcass traits and shear strength of meat in cattle.Crossref | GoogleScholarGoogle Scholar | 25548908PubMed |
Legesse G, Beauchemin KA, Ominski KH, McGeough EJ, Kroebel R, MacDonald D, Little SM, McAllister TA (2016) Greenhouse gas emissions of Canadian beef production in 1981 as compared with 2011. Animal Production Science 56, 153–168.
| Greenhouse gas emissions of Canadian beef production in 1981 as compared with 2011.Crossref | GoogleScholarGoogle Scholar |
Lobley G, Connell A, Lomax MA, Brown DS, Milne E, Calder AG, Farningham D (1995) Hepatic detoxification of ammonia in the ovine liver: possible consequences for amino acid catabolism. British Journal of Nutrition 73, 667–685.
| Hepatic detoxification of ammonia in the ovine liver: possible consequences for amino acid catabolism.Crossref | GoogleScholarGoogle Scholar | 7626587PubMed |
Marini JC, Van Amburgh ME (2005) Partition of nitrogen excretion in urine and the feces of Holstein replacement heifers. Journal of Dairy Science 88, 1778–1784.
| Partition of nitrogen excretion in urine and the feces of Holstein replacement heifers.Crossref | GoogleScholarGoogle Scholar | 15829671PubMed |
May M, DeClerck J, Quinn M, DiLorenzo N, Leibovich J, Smith D, Hales K, Galyean M (2010) Corn or sorghum wet distillers grains with solubles in combination with steam-flaked corn: feedlot cattle performance, carcass characteristics, and apparent total tract digestibility. Journal of Animal Science 88, 2433–2443.
| Corn or sorghum wet distillers grains with solubles in combination with steam-flaked corn: feedlot cattle performance, carcass characteristics, and apparent total tract digestibility.Crossref | GoogleScholarGoogle Scholar | 20407069PubMed |
McGinn S, Janzen H, Coates T, Beauchemin K, Flesch T (2016) Ammonia emission from a beef cattle feedlot and its local dry deposition and re-emission. Journal of Environmental Quality 45, 1178–1185.
| Ammonia emission from a beef cattle feedlot and its local dry deposition and re-emission.Crossref | GoogleScholarGoogle Scholar | 27380065PubMed |
Montgomery JL, Krehbiel CR, Cranston JJ, Yates DA, Hutcheson JP, Nichols WT, Streeter MN, Swingle RS, Montgomery TH (2009) Effects of dietary zilpaterol hydrochloride on feedlot performance and carcass characteristics of beef steers fed with and without monensin and tylosin. Journal of Animal Science 87, 1013–1023.
| Effects of dietary zilpaterol hydrochloride on feedlot performance and carcass characteristics of beef steers fed with and without monensin and tylosin.Crossref | GoogleScholarGoogle Scholar | 18997069PubMed |
NASEM (2016) ‘Nutrient requirements of beef cattle.’ 8th revised edn. (The National Academies Press: Washington, DC)
NRC (Ed. Subcommittee on Beef Cattle Nutrition) (2000) ‘Nutrient requirements of beef cattle.’ (The National Academies Press: Washington, DC)
Owens F, Zinn R, Kim Y (1986) Limits to starch digestion in the ruminant small intestine. Journal of Animal Science 63, 1634–1648.
| Limits to starch digestion in the ruminant small intestine.Crossref | GoogleScholarGoogle Scholar | 3539905PubMed |
Owens FN, Qi S, Sapienza DA (2014) Invited review: applied protein nutrition of ruminants: current status and future directions. The Professional Animal Scientist 30, 150–179.
| Invited review: applied protein nutrition of ruminants: current status and future directions.Crossref | GoogleScholarGoogle Scholar |
Ponce C, Cole N, Sawyer J, da Silva J, Smith D, Maxwell C, Brown M (2019) Effects of wet corn distillers grains with solubles and non-protein nitrogen on feeding efficiency, growth performance, carcass characteristics, and nutrient losses of yearling steers. Journal of Animal Science 97, 2609–2630.
| Effects of wet corn distillers grains with solubles and non-protein nitrogen on feeding efficiency, growth performance, carcass characteristics, and nutrient losses of yearling steers.Crossref | GoogleScholarGoogle Scholar | 30985872PubMed |
Reynolds CK, Kristensen NB (2008) Nitrogen recycling through the gut and the nitrogen economy of ruminants: an asynchronous symbiosis. Journal of Animal Science 86, E293–E305.
| Nitrogen recycling through the gut and the nitrogen economy of ruminants: an asynchronous symbiosis.Crossref | GoogleScholarGoogle Scholar | 17940161PubMed |
Robinson P, McQueen R (1994) Influence of supplemental protein source and feeding frequency on rumen fermentation and performance in dairy cows. Journal of Dairy Science 77, 1340–1353.
| Influence of supplemental protein source and feeding frequency on rumen fermentation and performance in dairy cows.Crossref | GoogleScholarGoogle Scholar | 8046074PubMed |
Rotger A, Ferret A, Calsamiglia S, Manteca X (2006) Effects of nonstructural carbohydrates and protein sources on intake, apparent total tract digestibility, and ruminal metabolism in vivo and in vitro with high-concentrate beef cattle diets. Journal of Animal Science 84, 1188–1196.
| Effects of nonstructural carbohydrates and protein sources on intake, apparent total tract digestibility, and ruminal metabolism in vivo and in vitro with high-concentrate beef cattle diets.Crossref | GoogleScholarGoogle Scholar | 16612021PubMed |
Ryan W, Williams I, Moir R (1993) Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights. Australian Journal of Agricultural Research 44, 1623–1633.
| Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights.Crossref | GoogleScholarGoogle Scholar |
Samuelson K, Hubbert M, Löest C (2016a) Effects of dietary urea concentration and zilpaterol hydrochloride on performance and carcass characteristics of finishing steers. Journal of Animal Science 94, 5350–5358.
| Effects of dietary urea concentration and zilpaterol hydrochloride on performance and carcass characteristics of finishing steers.Crossref | GoogleScholarGoogle Scholar | 28046136PubMed |
Samuelson K, Hubbert M, Oosthuysen E, Bester Z, Löest C (2016b) Effects of protein concentration and degradability on performance and carcass characteristics of finishing heifers receiving 0 or 400 mg ractopamine hydrochloride. In ‘Proceedings of the American Society of Animal Science, Western Section, Vol. 67’, 19–23 July 2016, Salt Lake City, UT, USA. pp. 200–204. (American Society of Animal Science: Champaign, IL) Available at https://www.asas.org/docs/default-source/western-section/wsasas_book_2016_ver4-1.pdf?sfvrsn=2 [Verified 2 August 2019]
Samuelson KL, Löest CA, Hubbert ME, Galyean ML (2016c) Nutritional recommendations of feedlot consulting nutritionists: the 2015 New Mexico State and Texas Tech University survey. Journal of Animal Science 94, 2648–2663.
| Nutritional recommendations of feedlot consulting nutritionists: the 2015 New Mexico State and Texas Tech University survey.Crossref | GoogleScholarGoogle Scholar | 27285940PubMed |
Scott D (1972) Excretion of phosphorus and acid in the urine of sheep and calves fed either roughage or concentrate diets. Quarterly Journal of Experimental Physiology and Cognate Medical Sciences 57, 379–392.
| Excretion of phosphorus and acid in the urine of sheep and calves fed either roughage or concentrate diets.Crossref | GoogleScholarGoogle Scholar | 4484585PubMed |
Scramlin SM, Platter WJ, Gomez RA, Choat WT, McKeith FK, Killefer J (2010) Comparative effects of ractopamine hydrochloride and zilpaterol hydrochloride on growth performance, carcass traits, and longissimus tenderness of finishing steers. Journal of Animal Science 88, 1823–1829.
| Comparative effects of ractopamine hydrochloride and zilpaterol hydrochloride on growth performance, carcass traits, and longissimus tenderness of finishing steers.Crossref | GoogleScholarGoogle Scholar | 20042550PubMed |
Shabi Z, Arieli A, Bruckental I, Aharoni Y, Zamwel S, Bor A, Tagari H (1998) Effect of the synchronization of the degradation of dietary crude protein and organic matter and feeding frequency on ruminal fermentation and flow of digesta in the abomasum of dairy cows. Journal of Dairy Science 81, 1991–2000.
| Effect of the synchronization of the degradation of dietary crude protein and organic matter and feeding frequency on ruminal fermentation and flow of digesta in the abomasum of dairy cows.Crossref | GoogleScholarGoogle Scholar | 9710769PubMed |
Sinclair LA, Garnsworth PC, Newbold JR, Buttery PJ (1993) Effect of synchronizing the rate of dietary energy and nitrogen release on rumen fermentation and microbial protein synthesis in sheep. The Journal of Agricultural Science 120, 251–263.
| Effect of synchronizing the rate of dietary energy and nitrogen release on rumen fermentation and microbial protein synthesis in sheep.Crossref | GoogleScholarGoogle Scholar |
Theurer CB (1986) Grain processing effects on starch utilization by ruminants. Journal of Animal Science 63, 1649–1662.
| Grain processing effects on starch utilization by ruminants.Crossref | GoogleScholarGoogle Scholar | 3539906PubMed |
Todd RW, Cole NA, Clark RN (2006) Reducing crude protein in beef cattle diet reduces ammonia emissions from artificial feedyard surfaces mention of trade or manufacturer names is made for information only and does not imply endorsement, recommendation, or exclusion by USDA–ARS. Journal of Environmental Quality 35, 404–411.
| Reducing crude protein in beef cattle diet reduces ammonia emissions from artificial feedyard surfaces mention of trade or manufacturer names is made for information only and does not imply endorsement, recommendation, or exclusion by USDA–ARS.Crossref | GoogleScholarGoogle Scholar | 16455840PubMed |
Trenkle A (1979) The relationship between acid-base balance and protein metabolism in ruminants. In ‘Regulation of acid–base balance’. (Eds WH Hales, P Meinhardt) pp. 149–157. (Church and Dwight Co. Inc.: Piscataway, NJ)
Trenkle A (2002) ‘Programmed feeding of protein to finishing beef steers. A.S. Leaflet R1774.’ Available at http://iowabeefcentre.org [Verified 1 September 2005].
Valkeners D, Théwis A, Piron F, Beckers Y (2004) Effect of imbalance between energy and nitrogen supplies on microbial protein synthesis and nitrogen metabolism in growing double-muscled Belgian Blue bulls1. Journal of Animal Science 82, 1818–1825.
| Effect of imbalance between energy and nitrogen supplies on microbial protein synthesis and nitrogen metabolism in growing double-muscled Belgian Blue bulls1.Crossref | GoogleScholarGoogle Scholar | 15217010PubMed |
Valkeners D, Théwis A, Amant S, Beckers Y (2006) Effect of various levels of imbalance between energy and nitrogen release in the rumen on microbial protein synthesis and nitrogen metabolism in growing double-muscled Belgian Blue bulls fed a corn silage-based diet1. Journal of Animal Science 84, 877–885.
| Effect of various levels of imbalance between energy and nitrogen release in the rumen on microbial protein synthesis and nitrogen metabolism in growing double-muscled Belgian Blue bulls fed a corn silage-based diet1.Crossref | GoogleScholarGoogle Scholar | 16543565PubMed |
Vasconcelos JT, Greene LW, Brown MS, Tedeschi LO, McCollum FT, Cole NA (2006) Effects of phase feeding of protein on performance, blood urea nitrogen concentration, manure nitrogen : phosphorus ratio, and carcass characteristics of feedlot cattle. Journal of Animal Science 84, 3032–3038.
| Effects of phase feeding of protein on performance, blood urea nitrogen concentration, manure nitrogen : phosphorus ratio, and carcass characteristics of feedlot cattle.Crossref | GoogleScholarGoogle Scholar | 17032797PubMed |
Vasconcelos JT, Cole NA, McBride KW, Gueye A, Galyean ML, Richardson CR, Greene LW (2009) Effects of dietary crude protein and supplemental urea levels on nitrogen and phosphorus utilization by feedlot cattle1. Journal of Animal Science 87, 1174–1183.
| Effects of dietary crude protein and supplemental urea levels on nitrogen and phosphorus utilization by feedlot cattle1.Crossref | GoogleScholarGoogle Scholar | 19028866PubMed |
Waggoner J, Löest C, Turner J, Mathis C, Hallford D (2009) Effects of dietary protein and bacterial lipopolysaccharide infusion on nitrogen metabolism and hormonal responses of growing beef steers. Journal of Animal Science 87, 3656–3668.
| Effects of dietary protein and bacterial lipopolysaccharide infusion on nitrogen metabolism and hormonal responses of growing beef steers.Crossref | GoogleScholarGoogle Scholar | 19648488PubMed |
Waldrip H, Todd R, Cole N (2013) Prediction of nitrogen excretion by beef cattle: a meta-analysis. Journal of Animal Science 91, 4290–4302.
| Prediction of nitrogen excretion by beef cattle: a meta-analysis.Crossref | GoogleScholarGoogle Scholar | 23825341PubMed |
Walker CE, Drouillard JS (2010) Effects of ractopamine hydrochloride are not confined to mammalian tissue: evidence for direct effects of ractopamine hydrochloride supplementation on fermentation by ruminal microorganisms1. Journal of Animal Science 88, 697–706.
| Effects of ractopamine hydrochloride are not confined to mammalian tissue: evidence for direct effects of ractopamine hydrochloride supplementation on fermentation by ruminal microorganisms1.Crossref | GoogleScholarGoogle Scholar | 19820054PubMed |
Walker D, Titgemeyer E, Drouillard J, Loe E, Depenbusch B, Webb A (2006) Effects of ractopamine and protein source on growth performance and carcass characteristics of feedlot heifers. Journal of Animal Science 84, 2795–2800.
| Effects of ractopamine and protein source on growth performance and carcass characteristics of feedlot heifers.Crossref | GoogleScholarGoogle Scholar | 16971581PubMed |
Walter LJ, Cole NA, Jennings JS, Hutcheson JP, Meyer BE, Schmitz AN, Reed DD, Lawrence TE (2016) The effect of zilpaterol hydrochloride supplementation on energy metabolism and nitrogen and carbon retention of steers fed at maintenance and fasting intake levels1. Journal of Animal Science 94, 4401–4414.
| The effect of zilpaterol hydrochloride supplementation on energy metabolism and nitrogen and carbon retention of steers fed at maintenance and fasting intake levels1.Crossref | GoogleScholarGoogle Scholar | 27898856PubMed |
Walter L-AJ, Schmitz AN, Nichols WT, Hutcheson JP, Lawrence TE (2018) Live growth performance, carcass grading characteristics, and harvest yields of beef steers supplemented zilpaterol hydrochloride and offered ad libitum or maintenance energy intake. Journal of Animal Science 96, 1688–1703.
| Live growth performance, carcass grading characteristics, and harvest yields of beef steers supplemented zilpaterol hydrochloride and offered ad libitum or maintenance energy intake.Crossref | GoogleScholarGoogle Scholar |
Zinn RA, Owens FN, Ware RA (2002) Flaking corn: processing mechanics, quality standards, and impacts on energy availability and performance of feedlot cattle. Journal of Animal Science 80, 1145–1156.
| Flaking corn: processing mechanics, quality standards, and impacts on energy availability and performance of feedlot cattle.Crossref | GoogleScholarGoogle Scholar | 12019600PubMed |
Zinn R, Barrajas R, Montano M, Ware R (2003) Influence of dietary urea level on digestive function and growth performance of cattle fed steam-flaked barley-based finishing diets. Journal of Animal Science 81, 2383–2389.
| Influence of dietary urea level on digestive function and growth performance of cattle fed steam-flaked barley-based finishing diets.Crossref | GoogleScholarGoogle Scholar | 14552362PubMed |
* Retired.