

Piglet birthweight and sex affect growth performance and fatty acid composition in fatty pigs
Marta Vázquez-Gómez A , Consolación García-Contreras B , Susana Astiz B , Laura Torres-Rovira B , Eugenio Fernández-Moya C , Álvaro Olivares A , Argimiro Daza D , Cristina Óvilo B , Antonio González-Bulnes B and Beatriz Isabel A E
A , Consolación García-Contreras B , Susana Astiz B , Laura Torres-Rovira B , Eugenio Fernández-Moya C , Álvaro Olivares A , Argimiro Daza D , Cristina Óvilo B , Antonio González-Bulnes B and Beatriz Isabel A E
A Faculty of Veterinary Medicine, Universidad Complutense de Madrid, Avenida Puerta de Hierro s/n, 28040 Madrid, Spain.
B Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Carretera de La Coruña km 7.5, 28040 Madrid, Spain.
C Ibéricos de Arauzo 2004 S.L., Calle Larga 33, 37408 Zorita de la Frontera, Spain.
D ETSIA, Universidad Politécnica de Madrid, Calle Ramiro de Maeztu 7, 28040 Madrid, Spain.
E Corresponding author. Email: bisabelr@ucm.es
Animal Production Science 60(4) 573-583 https://doi.org/10.1071/AN18254
Submitted: 18 April 2018 Accepted: 24 June 2019 Published: 17 January 2020
Journal compilation © CSIRO 2020 Open Access CC BY-NC-ND
Abstract
This study aimed to determine the effects of piglet birthweight (BIW) and sex, and within-litter BIW variation, on postnatal growth traits and meat quality in fatty breeds of pig. In total, 406 crossbred piglets (half male, half female) born to Iberian sows were studied during their postnatal development until slaughter. After birth, piglets were classified into four BIW categories: very low, low, medium and high. There was a negative effect of low BIW on growth patterns and fatty acid (FA) composition, but effects of litter size and within-litter BIW variation were not found. The very low BIW piglets underwent a period of significant catch-up growth (P < 0.005) relative to high BIW piglets during the early postnatal phase, but also showed a higher feed conversion rate and lower average daily weight gain (P < 0.05 for both measures) throughout the study period. BIW affected development during the entire productive life, and the sex effect increased with age. As a result, the period to reach market weight was longer in very low BIW piglets, by 43 days for females and 15 days for males, compared with their high BIW counterparts. BIW and sex also influenced amount of intramuscular fat, n-3 FA content and monounsaturated FA composition. The study indicates that BIW, modulated by sex, is a critical point for productive traits in fatty pigs. These results provide a basis for future strategies to enhance productive efficiency and meat quality of traditional swine breeds.
Additional keywords: carcass traits, Iberian pig, IUGR, lipids, meat quality.
Introduction
The increasing demand for high-quality, dry-cured products from fatty breeds of pig has moved management practices toward the more intensive rearing regimes used for lean breeds. Current reproductive strategies for swine production aim to increase prolificacy. However, there is evidence in lean breeds that greater litter size (LS) generates higher within-litter variation in birthweight (BIW) (Quiniou et al. 2002; Quesnel et al. 2008) and a higher incidence of piglets with low and very low BIW ( Foxcroft et al. 2006; Milligan et al. 2002). Available uterine space is decreased by increased prolificacy and leads to intra-uterine growth restriction (IUGR) processes, which may cause lower BIW in piglets (Wu et al. 2006). Intra-uterine growth restriction may be more severe in fatty breeds than in lean breeds because of the lower prolificacy and uterine capacity of the sows.
Lower BIW is associated with higher morbidity, different developmental patterns, and changes in body composition and homeostasis, due to prenatal programming through epigenetic changes (Gondret et al. 2005b, 2006; Gonzalez-Bulnes et al. 2016; Ji et al. 2017). Within-litter BIW variation (BIWV) may be associated with a high variability in carcass and meat quality in the same feedlot, which affects profitability and the production of dry-cured products (Andretta et al. 2016). Moreover, sex effects contribute to reducing feedlot homogeneity (Egea et al. 2016). Impaired postnatal growth patterns have been described for fatty breeds under experimental management (Gonzalez-Bulnes et al. 2012, 2014; Barbero et al. 2013). However, there is a lack of data under farm conditions, despite the high economic value of these breeds and their main differences from lean breeds (Nieto et al. 2012). Fatty pig breeds such as the Iberian breed produce well-recognised, dry-cured meat products of high quality (Lopez-Bote 1998). Homogeneity of growth patterns and meat quality within a feedlot are the primary goals in fatty pig production. However, fatty pigs show a lower weight homogeneity than lean breeds (Arévalo Mozos and Palomo Yagüe 2008; Soto et al. 2010).
The present study of fatty pigs under farm conditions aimed to determine the effects of BIW, within-litter BIWV and sex on postnatal development and carcass and meat quality at slaughter, including the fatty acid (FA) profile.
Materials and methods
Animals and diets
The study was performed according to the Spanish Policy for Animal Protection RD53/2013, which meets the European Union Directive (Directive 2010/63/EU revising Directive 86/609/EEC) on the protection of animals used for research. The experiment was specifically assessed and approved (report CEEA 2012/036) by the INIA Committee of Ethics in Animal Research. Animals were housed at a commercial farm, Ibéricos de Arauzo 2004 S.L. (Zorita de la Frontera, Salamanca, Spain).
In total, 406 crossbred piglets (~50% females and 50% males) were used in this study. They were born alive after insemination of 47 third-parity Iberian sows (Retinto strain) with cooled semen from Duroc boars (PIC (Genus plc), Stapeley, UK). The sows and offspring were managed in accordance with standard practices in commercial farms, including identification with electronic chips and housing indoors under controlled temperatures. Sows were allocated into groups for the first 101 days of pregnancy, and then placed in individual pens until the end of the suckling phase. Newborns were immediately sexed, measured, weighed and tagged. These piglets were allocated to mothers until weaning and then placed in collective pens. Males were surgically castrated within 2 days of birth. Sows and piglets were fed with standard, grain-based diets specific for Iberian pigs (diets are shown in Table S1, available as Supplementary Material to this paper), based on data from De Blas et al. (2013).
For experimental purposes, piglets were classified into four BIW categories: very low (VLBIW), low (LBIW), medium (MBIW) and high (HBIW). The classification was based on the mean and the standard deviation (s.d.) of the BIW of the study group of 406 piglets of 1.319 ± 0.313 kg (Blomberg et al. 2010; Vazquez-Gomez et al. 2016). The VLBIW group included piglets with a bodyweight below the mean minus 1 s.d. (BIW ≤0.99 kg); the LBIW group included piglets at the 30th percentile (after excluding VLBIW piglets, BIW 1.00–1.19 kg); the HBIW group included piglets from the 75th percentile (BIW ≥1.54 kg); and the MBIW group included piglets of BIW 1.20–1.50 kg.
At average age of 24 days (Day 24), 184 female and 175 male piglets were weaned. Of these, 132 females and 132 males (16 VLBIW, 24 LBIW, 55 MBIW and 37 HBIW pigs for each sex) were randomly selected and housed during the transition phase in groups of 12 piglets per pen, distributed by sex and BIW. During the growing–fattening phase (from Day 72 to slaughter), groups were randomly equalised to 120 males and 120 females (16 VLBIW, 24 LBIW, 44 MBIW and 36 HBIW pigs for each sex). Finally, 117 males and 115 females were slaughtered.
Evaluation of litter and piglet data at birth
Three litter-size categories were defined: small (total 3–6 piglets), medium (total 7–9 piglets), and large (total 10–13 piglets). The coefficient of variation (CV) and s.d. of BIW were calculated using the BIW of all piglets born alive for each litter.
Evaluation of growth pattern and fatness
Pigs were weighed from birth to slaughter at the following time points: birth, weaning (Day 24), at six points during postnatal growth (Days 71, 110, 150, 180, 215 and 240), and at slaughter, whenever they reached the minimum market weight (115 kg carcass weight; between Day 240 and Day 340). Bodyweight was determined individually at all these time points. On Day 340, the remaining pigs were sent to market regardless of their weight.
Average daily weight gain (ADWG) was calculated individually for the suckling phase, the transition phase (Days 25–71), five periods in the growing–fattening phase (Days 72–110, 111–150, 151–180, 181–215 and 216–240, with each period being labelled with the number of its last day), and the whole productive life. Feed conversion rate (FCR) for the five selected periods of the growing–fattening phase was calculated by pen, using the formula: daily feed intake block mean/ADWG of the period.
At birth and weaning, morphological measurements were recorded with a measuring tape. At weaning and at Days 110 and 215, backfat depth was determined at the level of the head of the last rib (P2 point) with an ultrasound machine (SonoSite, Bothell, WA, USA). At the slaughterhouse, the length of carcasses (from the posterior edge of the symphysis pubica to the anterior edge of the first rib) and the backfat thickness (at the last rib) were recorded. Carcass yield was calculated individually, using the formula: carcass weight/bodyweight at slaughter.
Tissue sample collection and drip-loss analysis
Samples of longissimus dorsi, the right lateral lobe of the liver and subcutaneous fat (SCF) at the measurement level were biobanked at −20°C until FA analysis. On the day of sampling, a sample of longissimus dorsi muscle was used for the drip-loss analysis (Calvo et al. 2016).
Evaluation of metabolic status
Metabolic status was assessed at slaughtering from blood plasma samples obtained by jugular puncture with 5 mL sterile heparin vacuum tubes (Vacutainer; BD, Franklin Lakes, NJ, USA). Plasma was separated and stored in vials at –20°C until assayed. Parameters for glucose (glucose and fructosamine) and lipid profiles (total cholesterol, high-density lipoprotein cholesterol, low-density lipoprotein cholesterol and triglycerides) were assessed with a clinical chemistry analyser (Crony Instruments, Rome). Plasma insulin concentrations in slaughter samples were also determined by using a Porcine ELISA kit (Mercodia, Uppsala, Sweden).
Evaluation of the fatty acid composition of diets
The one-step procedure proposed by Sukhija and Palmquist (1988) was used for the extraction and methylation of dietary FA. FA methyl esters were identified with a gas chromatograph (HP-6890; Hewlett Packard, Palo, Alto, CA, USA) with a flame ionisation detector and a capillary column (HP-Innowax: 30 m length, 0.32 mm i.d., 0.25 µm polyethylene glycol film thickness) (Lopez-Bote et al. 1997).
Evaluation of fat content and fatty acid composition of tissue samples
The lipids from intramuscular fat at the longissimus dorsi muscle (IMF) and liver fat were extracted as described by Segura and Lopez-Bote (2014). Total lipids in IMF and liver fat were separated into the main fractions: neutral lipids (NL) and polar lipids (PL) (Ruiz et al. 2004). SCF was separately analysed in outer and inner layers. Extracts were methylated (Segura et al. 2015b) and analysed using protocols developed at our laboratory (Lopez-Bote et al. 1997). Individual FA percentages for saturated, monounsaturated and polyunsaturated FA (SFA, MUFA and PUFA) were calculated. Total n-3, total n-6 FA, n-6/n-3 ratio and the unsaturated index were also calculated (Hulbert et al. 2007). The activity of stearoyl-CoA desaturase enzyme 1 (SCD-1) was estimated as C18:1/C18:0 and MUFA/SFA ratios (desaturation indices) (Hulver et al. 2005).
Statistical analyses
Data were analysed by using the GLM procedure in SAS version 9.4 (SAS Institute, Cary, NC, USA) with orthogonal contrasts (five contrasts). The first contrast was between sexes, the second between VLBIW pigs and other BIW groups, the third between VLBIW and LBIW pigs, the fourth between LBIW pigs and the sum of MBIW and HBIW pigs, and the fifth between MBIW and HBIW pigs. Changes over time in bodyweight and ADWG were assessed by a repeated-measures test with the Greenhouse–Geisser correction. BIW groups and sex were considered the main effects. Statistically significant interactions are presented. Sow was used as random effect in the birth and weaning analysis to account for the common maternal environment. Litter size was categorised into the three groups previously described and used as a random effect for birth data. For performance parameters, the respective age was used as a covariate. The Student’s t-test was used to identify differences between sexes in each BIW group.
The Chi-squared test was used to assess the mortality data and the percentage of VLBIW piglets, as described above. Pearson’s correlations were analysed using the PROC CORR procedure and regressions using the PROC REG procedure of SAS. The pig was the experimental unit for all variables studied except for the CV and s.d. of BIW and LS, where the sow was the unit, and for FCR data, with the pen. Tabulated results are expressed as mean ± root-mean-square error (RMSE), whereas graphical and text results are expressed as mean ± s.d. Statistical significance was accepted where P ≤ 0.05.
Results
Litter size and morphometric data of newborns
The mean LS was 8.9 ± 2.6 piglets born per sow (total), or 8.6 ± 2.6 piglets born alive. VLBIW and LBIW piglets born alive represented 15.8% and 14% of the total piglets. All morphological measurements were affected by BIW (P < 0.0001; Table 1), and the only sex-related effect was a longer (P < 0.005) thoracic perimeter in female than in male piglets (23.9 ± 2.1 cm vs 23.4 ± 2.5 cm). There were no significant differences between sexes for mean BIW or incidence of smaller piglets (male vs female: 13.3% vs 18.3% for VLBIW and 15.3% vs 12.8% for LBIW).

|
Almost half of the litters (48.9%) were classified as high LS, 32% medium LS, and 19.1% low LS. Moreover, the greater the LS, the more (P < 0.001) LBIW piglets were found for both sexes. Therefore, the mean value of BIW was lowered (P < 0.0001) with higher prolificacy (1.51 ± 0.26 kg for low LS vs 1.41 ± 0.28 kg for medium LS vs 1.23 ± 0.31 kg for high LS). Consequently, mean BIW decreased by ~43 g per pig for each unit increase in the total number of piglets born (P < 0.001; Fig. 1). However, the relationship between LS and BIW was weak because of the high BIWV. The greater the LS, the more within-litter BIWV (P < 0.005; Table 2) and differences were found between LS categories by s.d. (0.18 ± 0.10 for low LS vs 0.25 ± 0.08 for medium LS vs 0.27 ± 0.06 for high LS, P < 0.01). These data were reinforced by analysing BIWV by CV (12.1 ± 8.4% for low LS vs 18.4 ± 6.4% for medium LS vs 22.0 ± 4.7% for high LS, P < 0.001).
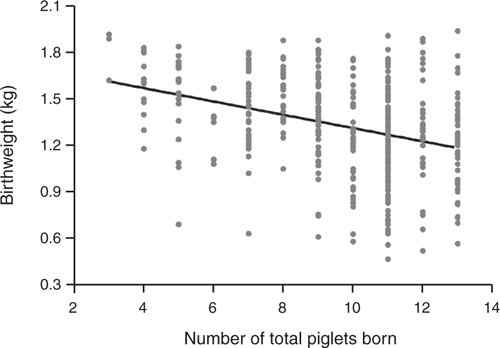
|

|
Effects of birthweight and sex on weaning traits
On average, 7.7 ± 0.7 piglets were weaned per sow, with no significant difference between sexes. Mortality was higher (P < 0.0001) in the VLBIW group (36%) than in heavier BIW groups (7%). Mean bodyweight at weaning was significantly correlated with BIW (r = 0.50, P < 0.0001; Table 2), and all morphological measurements were significantly different among BIW categories (P < 0.05 for all; Table 1). The lowest morphological measurements, weight and backfat depth (0.43 ± 0.11 cm vs 0.47 ± 0.12 cm, P < 0.05) were found in the VLBIW group, with significant differences relative to the other groups (P < 0.001 for all). Moreover, males showed greater backfat depth than females (0.48 ± 0.11 cm vs 0.44 ± 0.12 cm, P < 0.01).
Piglets in the VLBIW group also showed a lower (P < 0.0001) ADWG during the suckling phase (135 ± 38 g/day) than the other groups (mean 186 ± 47 g/day), and the LBIW group showed lower (P < 0.05) ADWG than piglets in groups MBIW and HBIW.
Effects of birthweight and sex on growing and fattening phases
At the end of the growth-transition phase (Day 71), VLBIW pigs continued showing lower (P < 0.01) ADWG (338 ± 76 g/day) than pigs from heavier BIW groups (381 ± 82 g/day). Moreover, LBIW pigs had lower (P < 0.05) weight and ADWG than the MBIW and HBIW pigs (Table 3).

|
At the beginning of the growing phase, at Day 110, LBIW pigs were heavier (P < 0.01) than VLBIW pigs (Table 3). Furthermore, HBIW and MBIW pigs showed lower ADWG and higher FCR than LBIW pigs (P < 0.005 for both; Table 3). There was a significant sex-related effect, with females showing higher weight and ADWG and lower FCR than males (P < 0.01 for both). Moreover, the changes in bodyweight and ADWG from birth to Day 240 were different among BIW groups and between sexes (P < 0.0001 for both).
At the following assessment, Day 150, VLBIW pigs remained lighter than the pigs in the other groups (54.0 ± 11.4 kg vs 58.7 ± 9.1 kg in the other groups, P < 0.001), mainly because of the lowest weight of VLBIW females at Day 150 (P < 0.0001; Fig. 2). At this age, the VLBIW category also showed lower ADWG and higher FCR than the other categories (P < 0.0005 for both; Table 3). At Day 180, VLBIW pigs continued to show lower (P < 0.05) weight and ADWG than pigs in the other groups, and FCR was lower (P < 0.001) with heavier BIW pigs. Regarding the sex effect, at Day 150, females had lower ADWG and higher FCR than males (P < 0. 0005 for both). At Day 180, males showed higher (P < 0.001) weight and ADWG than females.
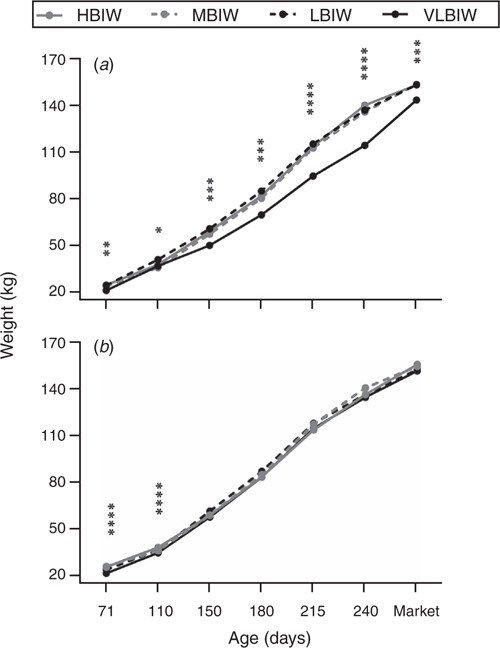
|
During the fattening phase, the VLBIW group continued to have lower weight and ADWG at both Day 215 and Day 240, and greater FCR at Day 215, than the other groups (P < 0.05 for all; Table 3). At Day 240, the LBIW group also showed lower ADWG and higher FCR than HBIW and MBIW groups (P < 0.005 for both; Table 3), mainly because LBIW males showed higher FCR. Moreover, males showed higher weight and lower ADGW than females at Day 240.
At slaughter, assessment of ADWG for the overall period revealed that the VLBIW group had lower (P < 0.001) values than the other groups (508 ± 74 g/day vs 568 ± 63 g/day). Moreover, LBIW pigs showed lower (P < 0.05) total ADWG than HBIW pigs (Table 3). The average number of days to market (DaM) linearly decreased (P < 0.05) with BIW increase (Table 4, Fig. 3), so that DaM was negatively correlated with birth and postnatal weights (Table 2). Hence, the VLBIW group was the oldest to be slaughtered and the lightest at slaughter. Furthermore, VLBIW females were the lightest (P < 0.0001) individuals at slaughter (Table 3).

|
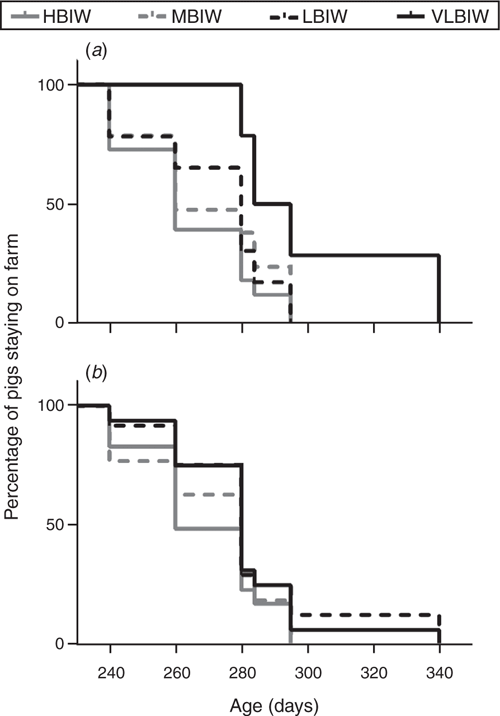
|
Regarding sex, in the simple linear regression between DaM and BIW, the intercepts and slopes of the relationship were different (P < 0.05; Table 4). DaM also accounted for 25% of the variation in weight at market, and the maximum point calculated was at Day 238, which means that beyond this time, weight at market decreased as DaM increased.
Effect of birthweight and sex on metabolic status
At slaughter, higher (P < 0.05) glucose concentrations were found in VLBIW pigs (127.3 ± 23.4 mg/dL) than in the other groups (mean value 107.6 ± 28.8 mg/dL), with a significant sex-related effect, VLBIW females showing the highest values (153.0 ± 14.1 mg/dL). On the other hand, plasma concentrations of insulin were highest in VLBIW males and lowest in VLBIW females (0.09 ± 0.05 µg/L vs 0.03 ± 0.00 µg/L; P < 0.05).
Effect of birthweight and sex on carcass traits and fatness
There were significant differences in fat accumulation between groups during postnatal development. The VLBIW group showed lower (P < 0.05) total backfat and outer layer depth than the other groups at Days 110 and 215 (Fig. 4). Moreover, the inner layer was thinner (P < 0.005) in the VLBIW group than in the other groups at Day 110. At Day 110, MBIW pigs had a thinner (P < 0.05) outer layer than HBIW pigs. The LBIW group showed thicker (P < 0.05) total backfat and inner layer than the heavier BIW groups at Day 215.
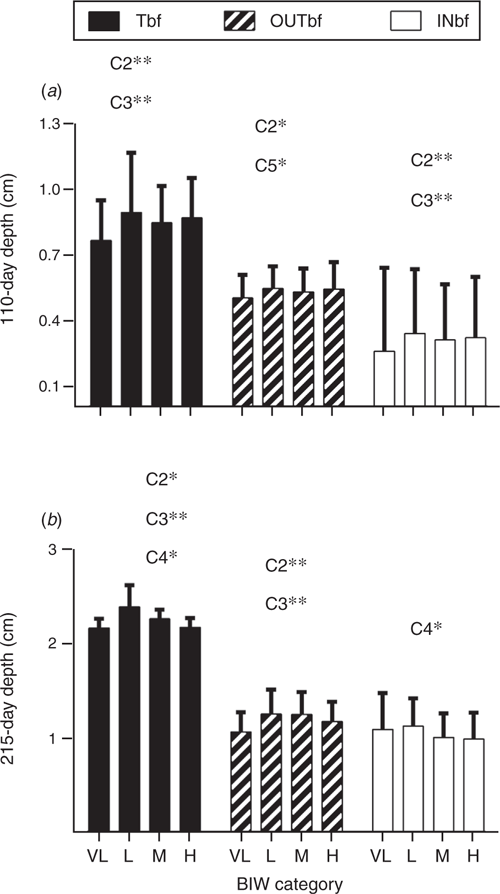
|
At Day 110, male pigs had a thicker (P < 0.05) inner layer than female pigs (0.33 ± 0.1 cm vs 0.30 ± 0.1 cm) but a thinner (P < 0.05) outer layer (0.52 ± 0.1 cm vs 0.56 ± 0.1 cm). At Day 215, males continued to have thicker (P < 0.0001) total backfat than females (2.36 ± 0.4 cm vs 2.13 ± 0.4 cm) and a thicker (P < 0.0001) inner layer (1.12 ± 0.3 cm vs 0.94 ± 0.3 cm). Furthermore, VLBIW females had the lowest values at Day 215 (P < 0.05). Conversely, at slaughter, there were no significant differences in backfat depths of carcasses between groups or between sexes (Table 5).

|
Regarding carcass traits at market, carcass length was shorter (P < 0.05) in the VLBIW group than in the other groups (Table 5), with VLBIW females having the lowest carcass length and weight. Carcass yield was positively correlated with age at market (Table 2), and the LBIW group showed higher (P < 0.05) IMF than the heavier BIW groups. Sex effects were found for IMF and liver fat, which were higher (P < 0.05 for both) in males than in females (Table 5).
Effects of birthweight and sex on tissue fatty acid composition
There were significant effects of both BIW category and sex on the FA profile of lipids from IMF (Table S2), SCF (Table S3) and liver fat (Table S4). The composition of lipids in IMF was influenced by both BIW and sex. Specifically, in the PL fraction, the VLBIW group showed higher SFA and MUFA and lower PUFA concentrations than the other groups (P < 0.05 for all). The main differences in the NL fraction were found in heavier BIW groups. The MBIW group showed lower (P < 0.01) concentrations of C18:1 n-9 and MUFA and lower (P < 0.05) desaturation indices (C18:1/C18:0 and MUFA/SFA) than the HBIW group, but a higher (P < 0.05) value of SFA. Sex-related effects in both NL and PL fractions were found. Considering only the NL fraction, males had higher (P < 0.05) MUFA concentration and unsaturated index, but lower (P < 0.05) SFA value than females. In the PL fraction, males showed higher (P < 0.05) C18:1 n-9 content.
The assessment of the FA profile of SCF (Table S3) addressed mostly sex-related differences, except that, independent of sex, the HBIW group showed higher (P < 0.05) n-3 FA values than the MBIW group in the outer layer of SCF. A similar FA profile to that of the IMF NL fraction was found in both the outer and inner layers for males and females. Moreover, females also showed a higher (P < 0.01) ∑n-6/∑n-3 ratio in the outer layer, with lower (P < 0.005) n-3 FA values at the inner layer.
Effects of BIW and sex were evident for liver fat (Table S4), with more significant differences between BIW groups in the NL fraction and more sex-related effects in the PL fraction. Assessment of the NL fraction showed that the VLBIW group had a higher SFA concentration and lower desaturation indices than the LBIW group (P < 0.05 for all). On the other hand, the LBIW group, compared with MBIW and HBIW groups, had higher (P < 0.05) MUFA concentrations and higher (P < 0.001) desaturation indices (C18:1/C18:0 and MUFA/SFA), and a lower (P < 0.05) SFA value. Sex-related effects were critical in the PL fraction, where C18:0 levels were higher (P < 0.05) for the VLBIW group than for the other groups; male and female VLBIW pigs were the most divergent in FA composition of the PL fraction, resulting in many significant global sex effects. Males showed higher C18:3 n-3 and MUFA concentrations and desaturation indices (P < 0.05 for all), but lower (P < 0.005) C18:0 and C18:2 n-6 values than females.
Discussion
The present study provides comprehensive evidence for the first time in a fatty breed of pig of the correlations between increases in LS and decreases in both BIW and within-litter BIW homogeneity. Subsequently, a low BIW penalised postnatal development and time to reach target weight, with carcass and meat quality decreasing within-feedlot homogeneity. These effects were also influenced by the sex of the offspring. Crossbred pigs were used in this trial, and similar results during postnatal development could be expected in purebred Iberian pigs. However, more experiments are needed to confirm this hypothesis.
Effects of litter size on birthweight and homogeneity
Larger LS was related to lower mean BIW and a higher within-litter BIWV and incidence of VLBIW piglets. Such effects have been described for lean breeds (Milligan et al. 2002; Foxcroft et al. 2006; Quesnel et al. 2008). The last of those studies stated that LS and BIWV are also affected by genotype, parity, management and nutritional (epigenetic) factors. Our results support that LS exerts a more pronounced effect on genotypes with a lower prolificacy. In the present study, the decrease in BIW with increasing LS seems greater in the Iberian breed (43 g of BIW per additional piglet born) than in lean genotypes (33–35 g of BIW; Quesnel et al. 2008; Beaulieu et al. 2010).
Effects of offspring birthweight and sex on postnatal development and fatness
A lower BIW was related to higher mortality during the suckling phase, as described in lean breeds (Quiniou et al. 2002; Wolf et al. 2008), and to significant differences in postnatal development patterns, as also described for lean breeds (Quiniou et al. 2002; Wu et al. 2006; Beaulieu et al. 2010). Overall, mean bodyweight and ADWG were highly influenced by BIW during suckling and transition phases. Data were not balanced by litter of origin but the statistical analysis ruled out any deviation caused by this effect.
We emphasise that LBIW, but not VLBIW, increased ADWG compared with heavier littermates during the period between weaning and the end of the transition phase. However, both groups increased ADWG at Day 110, and the weight of all BIW categories was more-or-less similar. These effects were more evident in females. Such data support the existence of a catch-up growth effect during the beginning of the growing phase (mainly evidenced in females) to increase the survival prospects of lighter piglets (Gonzalez-Bulnes and Ovilo 2012). Previous studies of our research group (Ayuso et al. 2015a, 2016) have related this catch-up growth to higher gene expression of pathways involved in cell growth and proliferation or protein turnover, which underlines the resilience of traditional breeds such as Iberian pigs.
Females showed decreased growth levels in the following growing periods. This development was confirmed by FCR, especially in VLBIW females, which did not maintain the expected growth rate during the growing or fattening phases. Hence, VLBIW females, despite catch-up growth at early postnatal stages, showed the lowest market weight and therefore the longest DaM value.
Overall, these results indicate that individuals with lower BIW do not compensate their low BIW during postnatal growth and are less efficient (higher FCR) than heavier littermates, which is similar to previous findings in lean breeds (Gondret et al. 2005b; Rehfeldt and Kuhn 2006; Bérard et al. 2008; Beaulieu et al. 2010). Moreover, a higher BIW was related to higher overall ADGW during the study period. Therefore, increases in the incidence of VLBIW and LBIW piglets enlarge the DaM period and raise production costs. Consequently, as the number of extra days needed to reach target market weight increases, the production costs relative to heavier BIW pigs also increases. In a lean genotype, Beaulieu et al. (2010) found a 10-day difference in reaching DaM between the lightest and heaviest BIW groups. In our study, the difference was larger in fatty pigs, being 15 days for males and 43 for females.
Effect of offspring birthweight and sex on back fat deposition and metabolism
The differential effects of BIW and sex on postnatal development were also observed in backfat deposition. Pigs from the VLBIW group showed lower backfat depth than pigs in the other BIW groups in the earlier life-periods; however, both VLBIW and LBIW pigs showed greater backfat depth at older ages (except for VLBIW females). This increase was particularly evident in the inner fat layer, which has more metabolic activity (Hausman and Thomas 1984). These results coincide with data from commercial breeds (Gondret et al. 2005a; Attig et al. 2008; Schinckel et al. 2010) and support the fact that lighter pigs have a higher tendency to accumulate fat, based on data supporting the hypothesis of prenatal programming.
Differences in glucose metabolism related to BIW were found at the end of the study. The VLBIW pigs showed higher glucose concentrations, which may be linked to an early prodrome of insulin resistance, modulated by sex, because VLBIW females had a low secretion of insulin but VLBIW males had the highest secretion of insulin (concomitant with a higher backfat depth).
Because the liver is the largest visceral organ, its status in individuals might affect physiological processes. Differences observed in backfat accumulation might be associated with the higher MUFA and desaturation indices of the lowest BIW groups, except VLBIW females, compared with the heaviest BIW groups. High SCD-1 activity has been related to metabolic disorders such as obesity and insulin resistance (Hulver et al. 2005; Poudyal and Brown 2011), although the LBIW group showed a regular at-market value of insulin. However, this higher activity of SCD-1, which plays a central role in de novo lipogenesis, might be linked to the beginning of an increase in liver fat storage. This is supported by the higher C16:1 content of LBIW pigs, this FA being considered an adiposity marker (Paillard et al. 2008). Other studies have found that decreased SCD-1 activity promotes lipid oxidation in lipid storage (Dobrzyn et al. 2004, 2005). This lower activity could be related to the lowest ratios of desaturation indices in VLBIW females and the lowest liver fat content. Further experiments are warranted to increase understanding of fat metabolism in LBIW pigs.
Effect of offspring birthweight and sex on carcass and meat quality at slaughter
The results for carcass traits revealed shorter carcass length for VLBIW pigs than for pigs in the other groups, which would be linked to poorer development in the growing–fattening phase. In fact, VLBIW females had the lowest carcass length and weight, which is consistent with their poorer postnatal development. Such an effect is well known in lean breeds, with carcasses from lighter BIW pigs having a lower weight of primary cuts, lower meat content and poorer meat quality (Rekiel et al. 2014).
Despite previous findings, no differences were found in backfat depth between BIW and sex categories at slaughter. This could be associated with the greater adipogenic capacity of Iberian pigs than of lean breeds (Nieto et al. 2012). Conversely, IMF content was higher in the lowest BIW groups and in males than in the heaviest BIW groups and in females, respectively. These data coincide with earlier studies in both lean and fatty breeds (Rehfeldt et al. 2008; Egea et al. 2016; Martinez-Macipe et al. 2016). Possible causes for IMF deposition in LBIW pigs may be related to a process of hyperplasia (increases in adipose cell number) during prenatal stages (Hausman et al. 2014).
Fatty acids in tissues are mostly distributed in PL and NL fractions, predominantly in the NL fraction (Wood et al. 2008). In the IMF, the NL fraction represents 70% of total FA, and is an estimator of total IMF FA (Ayuso et al. 2015b). Moreover, the membrane composition (PL) is more stable than the composition of storage lipids (NL) because of its functional properties (Sampels et al. 2011).
The loin is one of the most important carcass cuts in pork production and has a high economic value in the Iberian pig. Alvarenga et al. (2014) did not detect any effect of BIW on IMF composition when comparing light and heavy BIW pigs, but in our study, there were significant differences in the NL fraction of IMF between these groups. In the IMF NL fraction, the main differences were between MBIW and HBIW pigs. The HBIW group showed higher C18:1 n-9, MUFA, desaturation indices and unsaturated index than the MBIW group. In the Iberian pig, C18:1 n-9 has been used as an indicator of quality meat products, and high levels in backfat or meat pieces are linked to relevant traits of meat product quality, highly valued by consumers because of the association with the production of volatile fatty acids derived from oleic acid (Barea et al. 2013). Moreover, an increase in PUFA and MUFA meat levels is a priority because it enhances consumer acceptance owing to possible health and sensorial benefits (Laitinen et al. 2006; Jakobsen et al. 2009). In the PL fraction of IMF, the VLBIW group showed lower PUFA values and unsaturated index than heavier BIW groups.
A sex-related effect on the FA profile has been reported in many studies (Segura et al. 2015a; Daza et al. 2016; Egea et al. 2016). Differences between males and females were similar in SCF, IMF and the PL fraction of the liver. Overall, males had higher MUFA, but lower SFA and ∑n-6/∑n-3 ratio than females. This is interesting in view of the human dietary recommendations of a ∑n-6/∑n-3 ratio of 1 : 4 and higher MUFA values, as described above (Simopoulos 2002, 2010). From the present study, the FA profile is better in males than in females regarding meat quality.
Conclusions
This study clearly supports, for fatty breeds, that strong increases in LS are related to higher within-litter BIWV and a higher incidence of lighter BIW piglets. Our results also support the adverse effects of low BIW on postnatal growth traits and meat quality in fatty pigs. Pigs with lower BIW, despite catch-up growth processes, have lower ADWG and higher FCR during the growing and fattening phases, which increases daily feeding costs and induces a longer period to reach target weight. VLBIW females and males took 43 and 15 days longer than respective HBIW females and males to reach the market value. This situation, of course, increases production costs and reduces benefits.
Regarding carcass and meat traits, lighter BIW piglets presented poorer quality in carcass measurements, IMF values and FA profiles. Moreover, the HBIW group showed higher meat quality than MBIW pigs in the IMF FA profile. All of these effects were accentuated by sex, with females showing less growth potential than males during the growing phase and, at market, producing meat with lower organoleptic and health quality attributes.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We thank Antonio Palomo for the diet data, the staff of Ibéricos de Arauzo S.L. for their help with the animals and I. Martín de la Torre and M. L. Pérez-Solana for their technical support with FA and metabolic analyses, respectively. We also thank Anayansi Ramírez, César Ocasio and Bosco Vidovik for their help. This research was supported by the Ministry of Economy and competitiveness (project AGL2013-48121-C3-R and AGL2016-79321-C2-2-R), co-funded by FEDER. More funds were obtained from MEDGAN-REDLAB (project S2013/ABI-2913) and CAM, co-funded by FEDER. MVG and CGC are supported by the FPU and FPI National Program of the Spanish Government, respectively (MVG: FPU014/01285; CGC: BES-2014-070464).
References
Alvarenga A, Sousa R, Parreira G, Chiarini-Garcia H, Almeida F (2014) Fatty acid profile, oxidative stability of pork lipids and meat quality indicators are not affected by birth weight. Animal 8, 660–666.| Fatty acid profile, oxidative stability of pork lipids and meat quality indicators are not affected by birth weight.Crossref | GoogleScholarGoogle Scholar | 24636827PubMed |
Andretta I, Pomar C, Kipper M, Hauschild L, Rivest J (2016) Feeding behavior of growing–finishing pigs reared under precision feeding strategies. Journal of Animal Science 94, 3042–3050.
| Feeding behavior of growing–finishing pigs reared under precision feeding strategies.Crossref | GoogleScholarGoogle Scholar | 27482691PubMed |
Arévalo Mozos P, Palomo Yagüe A (2008) Evolución del peso postdestete según el rango de pesos al destete en lechones ibéricos [Evolution of the weight after weaning according to the range of weights at weaning in Iberian piglets]. Avances en tecnología porcina 5, 6–11.
Attig L, Djiane J, Gertler A, Rampin O, Larcher T, Boukthir S, Anton PM, Madec JY, Gourdou I, Abdennebi-Najar L (2008) Study of hypothalamic leptin receptor expression in low-birth-weight piglets and effects of leptin supplementation on neonatal growth and development. American Journal of Physiology. Endocrinology and Metabolism 295, E1117–E1125.
| Study of hypothalamic leptin receptor expression in low-birth-weight piglets and effects of leptin supplementation on neonatal growth and development.Crossref | GoogleScholarGoogle Scholar | 18780769PubMed |
Ayuso M, Fernández A, Núñez Y, Benítez R, Isabel B, Barragán C, Fernández AI, Rey AI, Medrano JF, Cánovas Á, González-Bulnes A, López-Bote C, Ovilo C (2015a) Comparative analysis of muscle transcriptome between pig genotypes identifies genes and regulatory mechanisms associated to growth, fatness and metabolism. PLoS One 10, e0145162
| Comparative analysis of muscle transcriptome between pig genotypes identifies genes and regulatory mechanisms associated to growth, fatness and metabolism.Crossref | GoogleScholarGoogle Scholar | 26695515PubMed |
Ayuso M, Ovilo C, Rodriguez-Bertos A, Rey AI, Daza A, Fenandez A, Gonzalez-Bulnes A, Lopez-Bote CJ, Isabel B (2015b) Dietary vitamin A restriction affects adipocyte differentiation and fatty acid composition of intramuscular fat in Iberian pigs. Meat Science 108, 9–16.
| Dietary vitamin A restriction affects adipocyte differentiation and fatty acid composition of intramuscular fat in Iberian pigs.Crossref | GoogleScholarGoogle Scholar | 26005912PubMed |
Ayuso M, Fernández A, Núñez Y, Benítez R, Isabel B, Fernández AI, Rey AI, González-Bulnes A, Medrano JF, Cánovas Á, López-Bote CJ, Óvilo C (2016) Developmental stage, muscle and genetic type modify muscle transcriptome in pigs: effects on gene expression and regulatory factors involved in growth and metabolism. PLoS One 11, e0167858
| Developmental stage, muscle and genetic type modify muscle transcriptome in pigs: effects on gene expression and regulatory factors involved in growth and metabolism.Crossref | GoogleScholarGoogle Scholar | 27936208PubMed |
Barbero A, Astiz S, Lopez-Bote CJ, Perez-Solana ML, Ayuso M, Garcia-Real I, Gonzalez-Bulnes A (2013) Maternal malnutrition and offspring sex determine juvenile obesity and metabolic disorders in a swine model of leptin resistance. PLoS One 8, e78424
| Maternal malnutrition and offspring sex determine juvenile obesity and metabolic disorders in a swine model of leptin resistance.Crossref | GoogleScholarGoogle Scholar | 24205230PubMed |
Barea R, Isabel B, Nieto R, Lopez-Bote C, Aguilera JF (2013) Evolution of the fatty acid profile of subcutaneous back-fat adipose tissue in growing Iberian and Landrace × Large White pigs. Animal 7, 688–698.
| Evolution of the fatty acid profile of subcutaneous back-fat adipose tissue in growing Iberian and Landrace × Large White pigs.Crossref | GoogleScholarGoogle Scholar | 23031353PubMed |
Beaulieu AD, Aalhus JL, Williams NH, Patience JF (2010) Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork1. Journal of Animal Science 88, 2767–2778.
| Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork1.Crossref | GoogleScholarGoogle Scholar | 20418451PubMed |
Bérard J, Kreuzer M, Bee G (2008) Effect of litter size and birth weight on growth, carcass and pork quality, and their relationship to postmortem proteolysis1. Journal of Animal Science 86, 2357–2368.
| Effect of litter size and birth weight on growth, carcass and pork quality, and their relationship to postmortem proteolysis1.Crossref | GoogleScholarGoogle Scholar | 18469061PubMed |
Blomberg LA, Schreier LL, Guthrie HD, Sample GL, Vallet J, Caperna T, Ramsay T (2010) The effect of intrauterine growth retardation on the expression of developmental factors in porcine placenta subsequent to the initiation of placentation. Placenta 31, 549–552.
| The effect of intrauterine growth retardation on the expression of developmental factors in porcine placenta subsequent to the initiation of placentation.Crossref | GoogleScholarGoogle Scholar | 20381143PubMed |
Calvo L, Toldrá F, Aristoy MC, López-Bote CJ, Rey AI (2016) Effect of dietary organic selenium on muscle proteolytic activity and water-holding capacity in pork. Meat Science 121, 1–11.
| Effect of dietary organic selenium on muscle proteolytic activity and water-holding capacity in pork.Crossref | GoogleScholarGoogle Scholar | 27232379PubMed |
Daza A, Latorre MA, Olivares A, López Bote CJ (2016) The effects of male and female immunocastration on growth performances and carcass and meat quality of pigs intended for dry-cured ham production: A preliminary study. Livestock Science 190, 20–26.
| The effects of male and female immunocastration on growth performances and carcass and meat quality of pigs intended for dry-cured ham production: A preliminary study.Crossref | GoogleScholarGoogle Scholar |
De Blas C, Gasa J, Mateos GG (2013) ‘Necesidades nutricionales para ganado porcino [Nutritional needs for swine livestock].’ (Fundación Española para el Desarrollo de la Nutrición Animal (FEDNA): Madrid)
Dobrzyn P, Dobrzyn A, Miyazaki M, Cohen P, Asilmaz E, Hardie DG, Friedman JM, Ntambi JM (2004) Stearoyl-CoA desaturase 1 deficiency increases fatty acid oxidation by activating AMP-activated protein kinase in liver. Proceedings of the National Academy of Sciences of the United States of America 101, 6409–6414.
| Stearoyl-CoA desaturase 1 deficiency increases fatty acid oxidation by activating AMP-activated protein kinase in liver.Crossref | GoogleScholarGoogle Scholar | 15096593PubMed |
Dobrzyn A, Dobrzyn P, Lee S-H, Miyazaki M, Cohen P, Asilmaz E, Hardie DG, Friedman JM, Ntambi JM (2005) Stearoyl-CoA desaturase-1 deficiency reduces ceramide synthesis by downregulating serine palmitoyltransferase and increasing β-oxidation in skeletal muscle. American Journal of Physiology. Endocrinology and Metabolism 288, E599–E607.
| Stearoyl-CoA desaturase-1 deficiency reduces ceramide synthesis by downregulating serine palmitoyltransferase and increasing β-oxidation in skeletal muscle.Crossref | GoogleScholarGoogle Scholar | 15562249PubMed |
Egea M, Linares MB, Garrido MD, Madrid J, Hernández F (2016) Feeding Iberian × Duroc cross pigs with crude glycerine: effects of diet and gender on carcass and meat quality. Meat Science 111, 78–84.
| Feeding Iberian × Duroc cross pigs with crude glycerine: effects of diet and gender on carcass and meat quality.Crossref | GoogleScholarGoogle Scholar | 26343013PubMed |
Foxcroft GR, Dixon WT, Novak S, Putman CT, Town SC, Vinsky MDA (2006) The biological basis for prenatal programming of postnatal performance in pigs. Journal of Animal Science 84, E105–E112.
| The biological basis for prenatal programming of postnatal performance in pigs.Crossref | GoogleScholarGoogle Scholar | 16582081PubMed |
Gondret F, Lefaucheur L, Louveau I, Lebret B (2005a) The long-term influences of birth weight on muscle characteristics and eating meat quality in pigs individually reared and fed during fattening. Archiv fur Tierzucht 48, 68–73.
Gondret F, Lefaucheur L, Louveau L, Lebret B, Pichodo X, Le Cozler Y (2005b) Influence of piglet birth weight on postnatal growth performance, tissue lipogenic capacity and muscle histological traits at market weight. Livestock Production Science 93, 137–146.
| Influence of piglet birth weight on postnatal growth performance, tissue lipogenic capacity and muscle histological traits at market weight.Crossref | GoogleScholarGoogle Scholar |
Gondret F, Lefaucheur L, Juin H, Louveau I, Lebret B (2006) Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs. Journal of Animal Science 84, 93–103.
| Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs.Crossref | GoogleScholarGoogle Scholar | 16361495PubMed |
Gonzalez-Bulnes A, Ovilo C (2012) Genetic basis, nutritional challenges and adaptive responses in the prenatal origin of obesity and type-2 diabetes. Current Diabetes Reviews 8, 144–154.
| Genetic basis, nutritional challenges and adaptive responses in the prenatal origin of obesity and type-2 diabetes.Crossref | GoogleScholarGoogle Scholar | 22283677PubMed |
Gonzalez-Bulnes A, Torres-Rovira L, Ovilo C, Astiz S, Gomez-Izquierdo E, Gonzalez-Anover P, Pallares P, Perez-Solana ML, Sanchez-Sanchez R (2012) Reproductive, endocrine and metabolic feto-maternal features and placental gene expression in a swine breed with obesity/leptin resistance. General and Comparative Endocrinology 176, 94–101.
| Reproductive, endocrine and metabolic feto-maternal features and placental gene expression in a swine breed with obesity/leptin resistance.Crossref | GoogleScholarGoogle Scholar | 22251656PubMed |
Gonzalez-Bulnes A, Astiz S, Ovilo C, Lopez-Bote CJ, Sanchez-Sanchez R, Perez-Solana ML, Torres-Rovira L, Ayuso M, Gonzalez J (2014) Early-postnatal changes in adiposity and lipids profile by transgenerational developmental programming in swine with obesity/leptin resistance. The Journal of Endocrinology 223, M17–M29.
| Early-postnatal changes in adiposity and lipids profile by transgenerational developmental programming in swine with obesity/leptin resistance.Crossref | GoogleScholarGoogle Scholar | 25107535PubMed |
Gonzalez-Bulnes A, Astiz S, Ovilo C, Lopez-Bote CJ, Torres-Rovira L, Barbero A, Ayuso M, Garcia-Contreras C, Vazquez-Gomez M (2016) Developmental origins of health and disease in swine: implications for animal production and biomedical research. Theriogenology 86, 110–119.
| Developmental origins of health and disease in swine: implications for animal production and biomedical research.Crossref | GoogleScholarGoogle Scholar | 27238437PubMed |
Hausman GJ, Thomas GB (1984) The development of the inner layer of backfat in fetal and young pigs. Journal of Animal Science 58, 1550–1560.
| The development of the inner layer of backfat in fetal and young pigs.Crossref | GoogleScholarGoogle Scholar | 6746444PubMed |
Hausman GJ, Basu U, Du M, Fernyhough-Culver M, Dodson MV (2014) Intermuscular and intramuscular adipose tissues: bad vs. good adipose tissues. Adipocyte 3, 242–255.
| Intermuscular and intramuscular adipose tissues: bad vs. good adipose tissues.Crossref | GoogleScholarGoogle Scholar | 26317048PubMed |
Hulbert AJ, Pamplona R, Buffenstein R, Buttemer WA (2007) Life and death: metabolic rate, membrane composition, and life span of animals. Physiological Reviews 87, 1175–1213.
| Life and death: metabolic rate, membrane composition, and life span of animals.Crossref | GoogleScholarGoogle Scholar | 17928583PubMed |
Hulver MW, Berggren JR, Carper MJ, Miyazaki M, Ntambi JM, Hoffman EP, Thyfault JP, Stevens R, Dohm GL, Houmard JA, Muoio DM (2005) Elevated stearoyl-CoA desaturase-1 expression in skeletal muscle contributes to abnormal fatty acid partitioning in obese humans. Cell Metabolism 2, 251–261.
| Elevated stearoyl-CoA desaturase-1 expression in skeletal muscle contributes to abnormal fatty acid partitioning in obese humans.Crossref | GoogleScholarGoogle Scholar | 16213227PubMed |
Jakobsen MU, O’Reilly EJ, Heitmann BL, Pereira MA, Bälter K, Fraser GE, Goldbourt U, Hallmans G, Knekt P, Liu S, Pietinen P, Spiegelman D, Stevens J, Virtamo J, Willett WC, Ascherio A (2009) Major types of dietary fat and risk of coronary heart disease: a pooled analysis of 11 cohort studies. The American Journal of Clinical Nutrition 89, 1425–1432.
| Major types of dietary fat and risk of coronary heart disease: a pooled analysis of 11 cohort studies.Crossref | GoogleScholarGoogle Scholar | 19211817PubMed |
Ji Y, Wu Z, Dai Z, Wang X, Li J, Wang B, Wu G (2017) Fetal and neonatal programming of postnatal growth and feed efficiency in swine. Journal of Animal Science and Biotechnology 8, 42
| Fetal and neonatal programming of postnatal growth and feed efficiency in swine.Crossref | GoogleScholarGoogle Scholar | 28484595PubMed |
Laitinen MH, Ngandu T, Rovio S, Helkala EL, Uusitalo U, Viitanen M, Nissinen A, Tuomilehto J, Soininen H, Kivipelto M (2006) Fat intake at midlife and risk of dementia and Alzheimer’s disease: a population-based study. Dementia and Geriatric Cognitive Disorders 22, 99–107.
| Fat intake at midlife and risk of dementia and Alzheimer’s disease: a population-based study.Crossref | GoogleScholarGoogle Scholar | 16710090PubMed |
Lopez-Bote CJ (1998) Sustained utilization of the Iberian pig breed. Meat Science 49, S17–S27.
| Sustained utilization of the Iberian pig breed.Crossref | GoogleScholarGoogle Scholar |
Lopez-Bote C, Rey A, Ruiz J, Isabel B, Sanz Arias R (1997) Effect of feeding diets high in monounsaturated fatty acids and α-tocopheryl acetate to rabbits on resulting carcass fatty acid profile and lipid oxidation. Animal Science 64, 177–186.
| Effect of feeding diets high in monounsaturated fatty acids and α-tocopheryl acetate to rabbits on resulting carcass fatty acid profile and lipid oxidation.Crossref | GoogleScholarGoogle Scholar |
Martinez-Macipe M, Rodríguez P, Izquierdo M, Gispert M, Manteca X, Mainau E, Hernández FI, Claret A, Guerrero L, Dalmau A (2016) Comparison of meat quality parameters in surgical castrated versus vaccinated against gonadotrophin-releasing factor male and female Iberian pigs reared in free-ranging conditions. Meat Science 111, 116–121.
| Comparison of meat quality parameters in surgical castrated versus vaccinated against gonadotrophin-releasing factor male and female Iberian pigs reared in free-ranging conditions.Crossref | GoogleScholarGoogle Scholar | 26398005PubMed |
Milligan BN, Fraser D, Kramer DL (2002) Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livestock Production Science 76, 181–191.
| Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights.Crossref | GoogleScholarGoogle Scholar |
Nieto R, Lara L, Barea R, García-Valverde R, Aguinaga MA, Conde-Aguilera JA, Aguilera JF (2012) Response analysis of the Iberian pig growing from birth to 150 kg body weight to changes in protein and energy supply. Journal of Animal Science 90, 3809–3820.
| Response analysis of the Iberian pig growing from birth to 150 kg body weight to changes in protein and energy supply.Crossref | GoogleScholarGoogle Scholar | 22665661PubMed |
Paillard F, Catheline D, Duff FL, Bouriel M, Deugnier Y, Pouchard M, Daubert J-C, Legrand P (2008) Plasma palmitoleic acid, a product of stearoyl-CoA desaturase activity, is an independent marker of triglyceridemia and abdominal adiposity. Nutrition, Metabolism, and Cardiovascular Diseases 18, 436–440.
| Plasma palmitoleic acid, a product of stearoyl-CoA desaturase activity, is an independent marker of triglyceridemia and abdominal adiposity.Crossref | GoogleScholarGoogle Scholar | 18068341PubMed |
Poudyal H, Brown L (2011) Stearoyl-CoA desaturase: a vital checkpoint in the development and progression of obesity. Endocrine, Metabolic & Immune Disorders Drug Targets 11, 217–231.
| Stearoyl-CoA desaturase: a vital checkpoint in the development and progression of obesity.Crossref | GoogleScholarGoogle Scholar |
Quesnel H, Brossard L, Valancogne A, Quiniou N (2008) Influence of some sow characteristics on within-litter variation of piglet birth weight. Animal 2, 1842–1849.
| Influence of some sow characteristics on within-litter variation of piglet birth weight.Crossref | GoogleScholarGoogle Scholar | 22444091PubMed |
Quiniou N, Dagorn J, Gaudré D (2002) Variation of piglets’ birth weight and consequences on subsequent performance. Livestock Production Science 78, 63–70.
| Variation of piglets’ birth weight and consequences on subsequent performance.Crossref | GoogleScholarGoogle Scholar |
Rehfeldt C, Kuhn G (2006) Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. Journal of Animal Science 84, E113–E123.
| Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis.Crossref | GoogleScholarGoogle Scholar | 16582082PubMed |
Rehfeldt C, Tuchscherer A, Hartung M, Kuhn G (2008) A second look at the influence of birth weight on carcass and meat quality in pigs. Meat Science 78, 170–175.
| A second look at the influence of birth weight on carcass and meat quality in pigs.Crossref | GoogleScholarGoogle Scholar | 22062267PubMed |
Rekiel A, Bartosik J, Więcek J, Batorska M, Kuczyńska B, Łojek A (2014) Effect of piglet birth weight on selected characteristics of pork. Annals of Animal Science 14, 967–975.
| Effect of piglet birth weight on selected characteristics of pork.Crossref | GoogleScholarGoogle Scholar |
Ruiz J, Antequera T, Andres AI, Petron M, Muriel E (2004) Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Analytica Chimica Acta 520, 201–205.
| Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods.Crossref | GoogleScholarGoogle Scholar |
Sampels S, Pickova J, Hogberg A, Neil M (2011) Fatty acid transfer from sow to piglet differs for different polyunsaturated fatty acids (PUFA). Physiological Research 60, 113–124.
Schinckel AP, Einstein ME, Jungst S, Booher C, Newman S (2010) Evaluation of the impact of pig birth weight on grow–finish performance, backfat depth, and loin depth. The Professional Animal Scientist 26, 51–69.
| Evaluation of the impact of pig birth weight on grow–finish performance, backfat depth, and loin depth.Crossref | GoogleScholarGoogle Scholar |
Segura J, Lopez-Bote CJ (2014) A laboratory efficient method for intramuscular fat analysis. Food Chemistry 145, 821–825.
| A laboratory efficient method for intramuscular fat analysis.Crossref | GoogleScholarGoogle Scholar | 24128551PubMed |
Segura J, Cambero M, Cámara L, Loriente C, Mateos G, López-Bote C (2015a) Effect of sex, dietary glycerol or dietary fat during late fattening, on fatty acid composition and positional distribution of fatty acids within the triglyceride in pigs. Animal 9, 1904–1911.
| Effect of sex, dietary glycerol or dietary fat during late fattening, on fatty acid composition and positional distribution of fatty acids within the triglyceride in pigs.Crossref | GoogleScholarGoogle Scholar | 26272512PubMed |
Segura J, Escudero R, Romero de Ávila MD, Cambero MI, López-Bote CJ (2015b) Effect of fatty acid composition and positional distribution within the triglyceride on selected physical properties of dry-cured ham subcutaneous fat. Meat Science 103, 90–95.
| Effect of fatty acid composition and positional distribution within the triglyceride on selected physical properties of dry-cured ham subcutaneous fat.Crossref | GoogleScholarGoogle Scholar | 25644667PubMed |
Simopoulos AP (2002) The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine and Pharmacotherapy 56, 365–379.
| The importance of the ratio of omega-6/omega-3 essential fatty acids.Crossref | GoogleScholarGoogle Scholar | 12442909PubMed |
Simopoulos AP (2010) The omega-6/omega-3 fatty acid ratio: health implications. Oléagineux Corps Gras Lipides 17, 267–275.
| The omega-6/omega-3 fatty acid ratio: health implications.Crossref | GoogleScholarGoogle Scholar |
Soto M, García JM, Palomo Yagüe A (2010) Evolución del peso post-destete según el rango de pesos al destete en lechones blancos [Evolution of the weight after weaning according to the range of weights at weaning in white piglets]. Avances en tecnología porcina 7, 56–62.
Sukhija PS, Palmquist DL (1988) Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. Journal of Agricultural and Food Chemistry 36, 1202–1206.
| Rapid method for determination of total fatty acid content and composition of feedstuffs and feces.Crossref | GoogleScholarGoogle Scholar |
Vazquez-Gomez M, Valent D, Garcia-Contreras C, Arroyo L, Ovilo C, Isabel B, Bassols A, Gonzalez-Bulnes A (2016) Sex and intrauterine growth restriction modify brain neurotransmitters profile of newborn piglets. International Journal of Developmental Neuroscience 55, 9–14.
| Sex and intrauterine growth restriction modify brain neurotransmitters profile of newborn piglets.Crossref | GoogleScholarGoogle Scholar | 27616302PubMed |
Wolf J, Žáková E, Groeneveld E (2008) Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livestock Science 115, 195–205.
| Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning.Crossref | GoogleScholarGoogle Scholar |
Wood JD, Enser M, Fisher AV, Nute GR, Sheard PR, Richardson RI, Hughes SI, Whittington FM (2008) Fat deposition, fatty acid composition and meat quality: a review. Meat Science 78, 343–358.
| Fat deposition, fatty acid composition and meat quality: a review.Crossref | GoogleScholarGoogle Scholar | 22062452PubMed |
Wu G, Bazer F, Wallace J, Spencer T (2006) Board-invited review: intrauterine growth retardation: implications for the animal sciences. Journal of Animal Science 84, 2316–2337.
| Board-invited review: intrauterine growth retardation: implications for the animal sciences.Crossref | GoogleScholarGoogle Scholar | 16908634PubMed |