

Effect of diurnal feeding times and sources of energy supplementation to optimise rearing of F1 Angus × Nellore young bulls
Andressa Scholz Berça A * , Abmael da Silva Cardoso B , Natália Vilas Boas Fonseca A , Dennis P. Poppi C , Luís Orlindo Tedeschi D , Isabela Rita Coletti Micheletti A , Willi Rocha Meireles A , Ana Cláudia Ruggieri A and Ricardo Andrade Reis A
A * , Abmael da Silva Cardoso B , Natália Vilas Boas Fonseca A , Dennis P. Poppi C , Luís Orlindo Tedeschi D , Isabela Rita Coletti Micheletti A , Willi Rocha Meireles A , Ana Cláudia Ruggieri A and Ricardo Andrade Reis A
A Department of Animal Sciences, Sao Paulo State University, Jaboticabal, SP 14884-900, Brazil.
B Range Cattle Research and Education Center, University of Florida, Ona, FL 33865, USA.
C School of Agriculture and Food Sciences, The University of Queensland, Gatton, Qld 4343, Australia.
D Department of Animal Science, Texas A&M University, College Station, TX 77843-2471, USA.
Animal Production Science 63(6) 579-595 https://doi.org/10.1071/AN22075
Submitted: 26 February 2022 Accepted: 14 November 2022 Published: 11 January 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Energy supplementation with highly fermentable carbohydrates can be a strategy to maximise the efficiency of nitrogen use (ENU) from high-nutritional value forages.
Aims: The study aimed to investigate the independent or associated effects of two diurnal feeding times (0900 or 1700 hours) with two sources of energy supplementation (corn or citrus pulp) in the growing of F1 Angus × Nellore young bulls on palisade grass pastures, and their implications on the forage chemical composition, nutrient intake and digestibility, ENU, microbial protein synthesis, animal performance, and gain per area.
Methods: There were 36 bulls used as experimental animals, with initial bodyweight (BW) of 290 ± 5 kg, and 32 similar bulls were used in a put-and-take system to maintain sward characteristics. The experiment was conducted in a complete randomised design with four treatments and three replications (paddocks). The treatments consisted of energy supplementation at 0.3% BW with corn or citrus pulp fed at 0900 or 1700 hours for four periods of 28 days. For evaluation of ruminal and blood parameters, ENU, intake, and nutrient digestibility, eight ruminal cannulated 1/2 Aberdeen Angus × 1/2 Nellore young bulls were used, with 280 ± 7 kg of initial BW, distributed in a 4 × 4 double Latin square design, consisting of four periods and four treatments.
Key results: Supplementation at 0.3% of BW with corn or citrus pulp at 0900 or 1700 hours can reach up to 1.06 kg/day of young bulls reared on palisade grass pastures fertilised with 180 kg nitrogen/ha/year and managed at 25 cm of height compared with corn-based supplements. There were no effects on microbial protein synthesis or its efficiency.
Conclusions: Citrus pulp can be used as an alternative energy source to corn, because it has the same potential for animal performance, ENU and microbial protein synthesis in tropical pastures with a high proportion of soluble protein, and structural and non-fibrous carbohydrates with high dry matter digestibility.
Implications: Supplementing grazing beef cattle at 0.3% BW with corn or citrus pulp in the morning or afternoon is an efficient nutritional strategy to improve animal performance.
Keywords: animal performance, beef cattle, citrus pulp, corn, microbial protein, palisade grass, ruminal parameters, supplementation time.
Introduction
In intensive cattle production systems, pastures fertilised with high doses of nitrogen (N) have high amounts of soluble protein, making energy supplementation essential to capture excess N of forage, and ensure the rumen balance between N and energy (Poppi and McLennan 1995). The energy source and the supplementation time combined with the grazing behavior of animals may directly affect the efficiency of nutrients used from the forage and, consequently, the responses in production.
The high degradability of N compounds in intensively managed tropical pastures, which can reach 14–15% of crude protein (CP), and 40–50% of soluble protein (De Oliveira et al. 2016; Delevatti et al. 2019a; Hoffmann et al. 2021; Leite et al. 2021), associated with the high content of slow degradation structural carbohydrates of forage, can compromise the efficiency of nitrogen use (ENU) and microbial protein synthesis (Detmann et al. 2014). However, this condition generates excessive ammonia (NH3) losses in the urine, generating a protein limitation relative to energy for high weight gains (Poppi et al. 2018). This N loss represents economic inefficiency, and can harm the environment through N losses in the form of volatilised NH3, nitrous oxide emission and nitrate leaching (Aboagye et al. 2018; Cardoso et al. 2019).
To minimise this problem, energy supplementation with highly fermentable carbohydrates can be a strategy to maximise the ENU from high nutritional value forages and microbial protein synthesis (Costa et al. 2011, 2019; Barbero et al. 2015; De Oliveira et al. 2016; Ferrari et al. 2021; Fonseca et al. 2022). The supply of metabolisable energy, combined with suitable amounts of rumen degradable protein in the forage, can capture the excess of rumen N by providing fermentable substrate containing starch, pectin, sugars or digestible fibre, depending on the energy source (Poppi and McLennan 1995; Costa et al. 2011; Barbero et al. 2015). Consequently, there will be a better balance between the availability of energy and NH3 in the rumen and, thus, greater animal performance (Poppi and McLennan 1995; Barbero et al. 2015; Ferrari et al. 2021).
In this scenario, citrus pulp has shown promising results in animal nutrition as an alternative to corn due to its positive effects on rumen fermentation (Ariza et al. 2001). As it is a byproduct of the orange juice industry and other citrus fruits, citrus pulp is a less expensive ingredient and does not compete with humans as a food source, unlike corn (Mottet et al. 2018). It has been shown that pectin promotes greater and faster capture of NH3 from degradation, reducing ruminal ammonia nitrogen (N-NH3), in addition to maximising microbial protein synthesis when compared with the addition of starch from corn (Ariza et al. 2001; Costa et al. 2011; De Oliveira et al. 2016). Moreover, ruminal fermentation of pectin leads to an increase in acetic acid production, and does not affect the production of lactic and propionic acids, which is in contrast to starch fermentation that decreases the rumen pH due to lactic and propionic acid formations (Van Soest 1994; Owens and Basalan 2016). However, although citrus pulp and corn are readily available sources, further studies evaluating the associated and/or independent effects of the energy supplement source and times to grazing animals are needed to maximise microbial protein synthesis and performance.
Besides the type of energy substrate (Poppi et al. 1997), the responses to energy supplementation may occur due to the supplementation timing, coinciding with the end of the main grazing events during the day. Studies on ingestive behavior by ruminants and specifically Nellore cattle show that the animals’ longest period of grazing occurs during the afternoon, more specifically between 1400 and 1700 hours (Thomsom et al. 1985; Páscoa 2009; Casagrande et al. 2011). Thus, the supply of energy supplements in the late afternoon after the intense intake of forage with a high amount of soluble N can be an efficient strategy to optimise animal performance, as the excess of N might be partly assimilated by the energy substrate for MCP.
This research hypothesises that energy supplementation, mainly citrus pulp supplied in the late afternoon, could optimise the use of forage with high CP and soluble N contents by capturing this N in the rumen or by independent effects of the carbohydrate being fermented. This could result in greater efficiency of MCP and, consequently, greater animal performance. Therefore, the objectives of this study were to investigate the independent or associated effects of two times of supplementation (0900 or 1700 hours) with two sources of energy supplementation (corn or citrus pulp) during the rearing of F1 Angus × Nellore young bulls on palisade grass pastures, and their implications on nutrient intake and digestibility, ENU and MCP, animal performance, and gain per area and stocking rate.
Materials and methods
The experiment was approved by the UNESP Council of Animal Experimentation and Animal Use, Campus Jaboticabal, under protocol 015234/19. The experiment was conducted at the Forage and Grasslands sector of the Sao Paulo State University ‘Julio de Mesquita Filho’ (UNESP), in Jaboticabal, SP, Brazil (21°15′22″S latitude, 48°18′58″W longitude and 595 m elevation). The soil of the experimental area is classified as Ferralsol (Embrapa 2013). According to the Köeppen system, the climate is Aw type (tropical, characterised by dry winters).
The experiment was conducted in the rainy season of 2019/2020, between December 2019 and April 2020, during the rearing phase of young bulls. Animals underwent a 15-day adaptation period to the experimental conditions and the diet, and subsequent 112 days of experimental evaluation.
The meteorological data were measured daily through the UNESP/FCAV Agro-Meteorological Weather Station. Over the whole experimental period, the average temperature was 24.2°C, with a minimum of 20.4°C and a maximum of 30.9°C, the average precipitation was 156.2 mm, and the insolation time was 203.0 h.
Experimental design
The experiment was conducted using a complete randomised design in a 2 × 2 factorial arrangement, with four treatments and three replications (paddocks) per treatment. Treatments consisted of energy supplementation at 0.3% dry matter (DM) of bodyweight (BW) with corn or citrus pulp at 0900 or 1700 hours. The evaluated supplements are described in Table 1 with respect to ingredients and chemical composition.
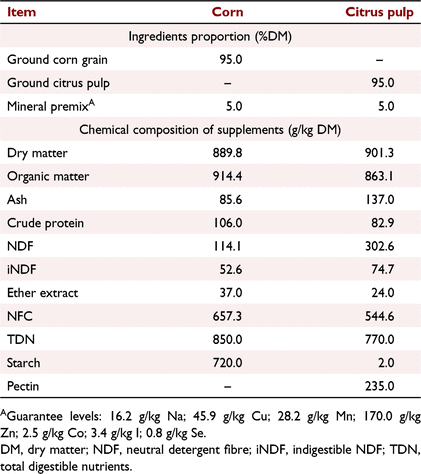
|
Animals and pasture management
We used 36 1/2 Aberdeen Angus × 1/2 Nellore young bulls (aged 14 ± 2-months) as experimental animals, with mean initial BW (IBW) of 290 ± 5 kg, and eight 1/2 Aberdeen Angus × 1/2 Nellore young bulls (aged 14 ± 2-months), cannulated in the rumen, with an IBW of 280 ± 7 kg, distributed in a double 4 × 4 Latin square design. In addition, 32 young bulls were also used as put-and-take animals to adjust the stocking rate and maintain 25 cm of pasture height, with the same genetic pattern and IBW of 256 ± 8 kg.
Before the beginning of the experimental period, all animals were weighed, identified with numbered ear tags, and endo- and ectoparasites controlled using albendazole sulfoxide (Agebendazol 15%, 7.5 mL/head, Agener Uniao®, Sao Paulo, SP, Brazil) as a vermifuge, and fluazuron pour-on (Acatak, 30 mL/head, Novartis®, Campo Grande, MS, Brazil) for cattle tick control. Experimental animals were distributed into the treatments based on average BW, and the adjustment of stocking rate per paddock was based on the canopy height. The experiment with experimental animals had a total duration of 112 days, divided into four periods of 28 days.
Cannulated animals were kept in the same paddocks as experimental animals during the experiment, and received the same type and amount of supplement for each treatment (0.3% BW). First, these animals underwent a 15-day adaptation period to the pastures and the supplement. The total experimental period for cannulated animals was 84 days, divided into four periods of 21 days. The first 15 days were destined for diet adaptation, and the last 6 days were for collections of ruminal fluid, blood, urine and faeces. The eight cannulated animals were randomly divided into four pairs (two cannulated/treatment), rotating in all treatments throughout the four experimental periods. These two animals were allocated in separate paddocks of the same treatment; therefore, within the three paddocks of each treatment, only two received cannulated bulls.
Experimental animals were used to evaluate animal performance, whereas cannulated animals were used to assess nutrient intake and digestibility, urinary, ruminal, and blood parameters.
The experimental pastures were palisade grass (Urochloa brizantha R. D. Webster cv. Marandu). The total experimental area was 13 ha, of which 1 ha was reserve area, and 12 ha for the evaluations of treatments and replications. The site was divided into 12 paddocks (three paddocks/treatment), provided with an open trough with 40 linear cm per animal and a water trough, with fresh, clear water. Each paddock allocated three experimental animals (n = 9 animals/treatment). The management corral is round, and equipped with a cattle chute and a digital scale.
Maintenance fertilisation of pastures was performed before the animals’ adaptation period and determined according to the soil analysis results, following the recommendation of Technical Bulletin 100 (Van Raij et al. 1997). Nitrogen fertilisation of all paddocks was provided by ammonium nitrate (32% N), fractionated in three applications of 60 kg N/ha/year, totalling 180 kg N/ha/year (11 December 2019; 31 January 2020; 28 February 2020).
Pastures were managed under continuous stocking with a variable stocking rate, following the put-and-take technique (Mott and Lucas 1952) to maintain 25 cm of height and at a grazing utilisation efficiency of 50%. The stocking rate was adjusted using put-and-take animals. For this assay, the height/forage mass ratio (bulk density) was evaluated weekly to estimate the forage mass, and adjust the stocking rate using the put-and-take animals by measuring 80 random points of each paddock with a graduated ruler (Barthram 1985).
Previous studies showed that under these experimental conditions, 25 cm of height promotes 95% light interception, which allows achievement of maximum net forage accumulation, with high leaf production and low senescence, resulting in high-quality forage allowance that may increase animal performance (Barbero et al. 2015; de Araújo et al. 2021; Hoffmann et al. 2021).
Forage mass and morphological composition of pasture
To determine forage mass, forage allowance and morphological composition, every 28 days, 80 points/ha of palisade grass height were randomly measured with a graduated ruler (Barthram 1985). From the average height, three representative samples of grass were collected per paddock by cutting at 5-cm height from the soil, all forage contained in a 0.25-m2 metallic frame. Each forage sample was subdivided into two subsamples, one for determination of DM concentration and the other for determination of the morphological composition, fractionating them into leaf, stem + sheath and dead material. After drying in an oven with forced air circulation at 55°C for 72 h, total DM and morphological components were obtained to estimate the total forage mass. The forage allowance (kg DM/kg BW) was calculated as forage mass divided by the animal stocking rate, and multiplied by the percentage of leaves to determine the leaf allowance.
Forage mass, forage and leaf allowances, and morphological composition of pastures are presented in Table 2.

|
Chemical composition of the forage
To evaluate the chemical composition of forage, hand-plucked samples of each paddock were taken every 28 days by collecting the forage at average canopy height after closely observing the animal grazing behaviour, avoiding causing the animals stress, to represent all forage consumed during the day (Sollenberger and Cherney 1995). After drying in an oven with forced air circulation at 55°C for 72 h to determine DM, samples were ground in a Willey mill with a 1-mm sieve and then submitted to chemical analysis, following the methodologies of AOAC (2012). Chemical components analysed were as follows: total N (CP) by the Kjeldahl method; acid detergent fiber (ADF) and neutral detergent fiber (NDF) corrected for ash and protein (apNDF) by the ANKOM fiber analyser method (ANKOM Technology, USA); lignin by the acid hydrolysis method; ash; and ether extract (EE) by the Goldfish method. The organic matter (OM), hemicellulose and cellulose contents were calculated by the difference between previously determined fibrous components, following the Eqns 1–3:



The fractionation of N was determined following the Cornell system (Sniffen et al. 1992). Fraction A was obtained by analysing the hand-plucked samples in trichloroacetic acid to extract soluble N (Detmann et al. 2012), and later calculated by the difference between the total N content and the non-protein nitrogen (NPN). The B3 fraction was calculated by the difference between the neutral detergent-insoluble nitrogen (NDIN) and the acid detergent-insoluble nitrogen (ADIN) contents. Fraction C, in turn, was considered ADIN, described by Sniffen et al. (1992) and thoroughly discussed by Tedeschi and Fox (2020). Finally, Fraction B1 + B2 (true protein) was calculated as the difference between the total N and the other fractions (B1 + B2 = total N − (A + B3 + C)).
Forage carbohydrate fractions were obtained from Eqns 4–6 proposed by Sniffen et al. (1992), respectively, for total carbohydrates (TC), non-fibrous carbohydrates (NFC) and total digestible nutrients (TDN). The chemical composition, carbohydrate and protein fractions of forage are presented in Table 3.

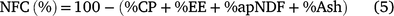

where ‘ad’ means apparently digestible fractions and ‘d’ means digestible fractions.

|
Nutrients intake and digestibility
Intake by animals was determined based on faecal production of cannulated bulls, using indigestible neutral detergent fibre (iNDF) as an internal marker and chromium oxide (Cr2O3) as an external marker, which is the standard used for investigative purposes in Brazil, and approved by the local and national Ethics Committees on the Use of Animals.
For this assay, 10 g of Cr2O3/animal/day were wrapped in paper and allocated directly in the rumen of each cannulated animal for 10 days at the same time, of which the first 7 days for adaptation and the last 3 days for faeces collection (Hopper et al. 1978). The supply of Cr2O3 began on the eighth day of each cannulated experimental period, and collections started on the 15th day. Faecal collections were performed twice a day, according to the schedule: first day – 0700 and 1900 hours; second day – 1100 and 2300 hours; 3rd day – 1500 and 0300 hours.
After collection, faeces samples were dried in an oven with forced air circulation at 55°C for 72 h, weighed, and constituted a composited sample of 3 days of collection per animal and period.
Faecal recovery of Cr2O3 was determined following the methodology of Williams et al. (1962), and, from these data, faecal excretion (FE) was calculated through the equation proposed by Detmann et al. (2001; Eqn 7). Forage intake was calculated based on iNDF concentrations of forage and faeces (Eqn 8).


To estimate supplement intake of cannulated animals, titanium dioxide (TiO2) was used as an external marker (Titgemeyer et al. 2001). Approximately 10 g TiO2/animal/day were provided for 10 days, being properly homogenised to the supplement immediately before feeding the animals from the group supplement trough. Each paddock’s total amount of TiO2 was provided according to the number of grazing animals. The indicator concentration in individual animal faeces was analysed by atomic absorption spectrophotometry, and a standard curve was established with concentrations 0, 2, 4, 6, 8 and 10 (Myers et al. 2004). Individual animal supplement intake was calculated following Eqn 9.
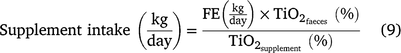
Nutrient digestibility was estimated through the quantification of the iNDF content. Samples of forage, faeces and supplements were individually placed in filter bags model F-57 from ANKOM brand, and allocated in the rumen of cannulated animals for in situ incubation for 288 h (Norris et al. 2019). After removing the bags from the rumen, they were washed until completely clear, dried in a forced circulation oven at 55°C for 72 h and then in a non-ventilated oven at 105°C for 45 min. In sequence, bags were subjected to extraction with neutral detergent in an ANKOM fibre analyser, and the entire drying procedure was repeated to quantify iNDF, according to Detmann et al. (2001). Nutrient intake and digestibility by cannulated animals were calculated from these data, following Eqns 10 and 11, respectively, considering the total intake of forage and supplement (diet).


where TNI refers to total nutrient intake, FE to fecal excretion and NC is the nutrient concentration of diet (forage + supplement).
Efficiency of N use and microbial protein synthesis
Spot samples of urine were collected from each cannulated animal simultaneously with faecal collections: first day at 0700 and 1900 hours, second day at 1100 and 2300 hours, and third day at 1500 and 0300 hours (Chizzotti et al. 2008; Silva Júnior et al. 2018). Composite samples were obtained from the six collections for each animal per period.
Urine was immediately filtered, and 10 mL aliquots were diluted with 40 mL of 0.072 M sulfuric acid (H2SO4) to prevent bacterial degradation of purine derivatives (PD) and precipitation of uric acid. Samples were stored at −15°C until analysis.
Concentrations PD, such as uric acid (UA) and allantoin (ALLA), were determined using the commercial Analisa kits by enzymatic-colorimetric methodology (Cat. 451/MS80022230065), and Young and Conway (1942), respectively.
The total excretion of PD was calculated by summing the amounts of ALLA and UA excreted in the urine, expressed in mmol/day. Absorbed purines (Pabs) were calculated from the excretion of PD, following Eqn 12. Then, the MCP (g/day) was calculated as a function of Pabs, using Eqn 13 (Detmann et al. 2014).
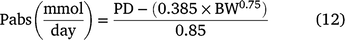

The ENU was calculated following Eqn 14, described by Detmann et al. (2014).
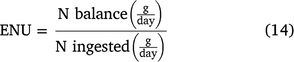
where 70 corresponds to the N concentration in the purines (mg N/mol); 0.137 to the ratio N purine:total N in the bacteria (Barbosa et al. 2006); and 0.85 to the digestibility of microbial purines. The N balance was calculated from the sum of the N excretions in the faeces and the urine, subtracted from the N ingestion.
Another aliquot of concentrated urine was used to estimate the total N concentration by the Kjeldahl methodology (Fenner 1965), and the urea concentration through commercial Analisa kits by the colorimetric-enzymatic method (Cat. 427/MS80022230063). Creatinine concentration in the spot sample was also determined through commercial Analisa kits using the colorimetric method with alkaline picrate (Cat. 435/MS80022230066) and subsequently used to estimate the urinary volume. The daily creatinine excretion (CE) was calculated by the following Eqn 15, proposed by Chizzotti et al. (2008):

The nitrogen excreted via urine was obtained by multiplying the urine volume by urinary N concentration (Chizzotti et al. 2008).
Ruminal and blood parameters
To evaluate the pH, N-NH3 and short-chain fatty acids (SCFA), ruminal fluid samples were collected from the cannulated animals every three alternate hours in the last 2 days of each experimental period. The ruminal fluid was collected at the solid-liquid interface and immediately filtered in a triple layer of gauze for pH measurement with a digital pH analyser, calibrated with pH 7.0 and 4.0 buffers.
One millilitre of sulfuric acid (1:1) was added to a 50-mL sample of ruminal fluid and stored at −20°C for subsequent analysis of N-NH3 using the Kjeldahl methodology (Fenner 1965). Analysis of SCFA was performed using concentrated ruminal fluid samples, following Serafim et al. (2021) methodology on a Shimadzu high-performance liquid chromatography system (model Prominence) equipped with an ultraviolet detector model SPD-20 and programmed to operate at a wavelength of 210 nm. The internal standard for the high-performance liquid chromatography was the crotonic acid. The ruminal fluid samples were centrifuged at 6026g for 5 min and filtered through a 13-mm diameter nylon filter (0.22-μm pore size). The injection volume was 20 μL. The quantified SCFAs were formic, lactic, acetic, propionic, butyric, isovaleric and valeric acids.
Total gas production and in vitro dry matter degradability
The in vitro dry matter degradability (IVDMD) and total gas production trials were conducted at Premix Company Research Center Laboratory, located in Patrocinio Paulista, SP, Brazil.
The analysis was performed on samples of hand-plucked forage, supplement and composite samples of forage + supplement in the real proportion of animal DM intake (88% and 12%, respectively), following the methodology of Bueno et al. (2005, 2008). For this assay, 1 g of the substrate (forage and/or supplement samples) was weighed and incubated in previously weighed and identified glass bottles. The composite sample substrate was formulated by mixing 0.88 g (DM) of forage and 0.12 g (DM) of supplement (corn or citrus pulp).
In addition to the substrate, 90 mL of aqueous culture media were added to each bottle, consisting of a solution of microminerals, macrominerals, buffer solution, resazurin solution, medium B and distilled water. After inserting the culture medium, 10 mL of ruminal fluid was added to each bottle, which was collected on the day of incubation from two cannulated bulls, immediately capped, homogenised and incubated in a forced air circulation oven at 39°C with constant movement and shaker type. Those animals were kept in Marandu grass pastures and adapted to a 0.3% BW energy supplement (810 g/kg TDN, 95 g/kg CP), and the average BW was 310 ± 7 kg.
The IVDMD was determined through the recovery of three bottles of the non-degraded fraction of each treatment, using filtration of the residue in a glass crucible previously prepared with glass wool (Bueno et al. 2005, 2008).
For forage samples incubation, the IVDMD was measured at seven different timepoints: 0, 3, 6, 12, 24, 48 and 72 h after incubation. The treatments were Marandu grass pastures where animals were supplement with corn at 0900 hours, citrus pulp at 0900 hours, corn at 1700 hours and citrus pulp at 1700 hours (n = 4). We used two forage samples/treatment, and the IVDMD was evaluated by recovering two bottles of the non-degraded fraction of each treatment at each timepoint, totalling 14 bottles/treatment.
For supplement and forage + supplement incubations, the IVDMD was measured at six different timepoints: 0, 3, 6, 12, 24 and 48 h after incubation. For supplements only, the treatments were corn or citrus pulp (n = 2), and we used three forage samples/treatment, so the IVDMD was evaluated through the recovery of three bottles of the non-degraded fraction of each treatment at each timepoint totalling 18 bottles/treatment.
For the forage + supplement incubation, the treatments were Marandu grass + corn or Marandu grass + citrus pulp (n = 2), and four samples/treatments were used, so the IVDMD was evaluated through the recovery of four bottles of the non-degraded fraction of each treatment in each timepoint, totalling 24 bottles/treatment.
The potential degradability was calculated following the equation of Ørskov and McDonald (1979) (Eqns 16, 17).

where potential degradability is the potential degradability (%) in incubation time T; A is the fraction of feed that degrades instantly; B is the fraction insoluble in water, but potentially degradable in the rumen; C is a constant rate of degradation of fraction B.
The effective degradability (DE) of feed, allowing for rate of passage, was calculated by the equation below.
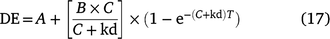
where DE is the effective degradability (%); kd is the degradation rate, estimated by regression analysis.
Total gas production was measured on all remaining bottles throughout the timepoints before recovering the bottles to determine IVDMD, using a transducer and a data logger, the total gas volume was determined in psi and later converted to mL. For forage samples, measurements were taken on all bottles at times 0, 6, 12, 18, 24, 32, 40, 48, 60 and 72 h after incubation. For supplement and forage + supplement samples, gas production was measured at times 0, 3, 6, 12, 24, 32, 40 and 48 h after incubation.
Animal performance
To determine the ADG of experimental animals, weighing was performed at the beginning (IBW), after the adaptation period and the end (final BW (FBW)) of the total experimental period, after a 14-h feed and water fasting. The ADG was calculated by dividing the difference between FBW and IBW by the number of days (112 days). In addition, every 28 days after the adaptation period, we conducted an intermediate weighing, with no previous fasting, to adjust the stocking rate, grazing height and supplement quantity.
The days of the occupation of the put-and-take animals in each paddock were recorded for further calculation of stocking rate and gain per area. The gain per area was calculated according to the experimental ADG, and the number of put-and-take animals and experimental animals of each paddock during the evaluation period (Mott and Lucas 1952). The stocking rate in animal units (AU) per hectare (450 kg BW/ha), forage and leaf allowances were determined from the total BW of the animals in each paddock in each period.
Statistical analysis
Data were analysed for the homoscedasticity and normality of the residues, using the Box–Cox and Cramer–von Mises tests, respectively, using the Nortest package of the R program (R Core Team 2018). For animal performance evaluation, the paddock was assumed as the experimental unit (three paddocks/treatment), considering the mean composed of three animals in each paddock fed the same supplement and forage. The IBW was added as a covariate on the FBW and ADG, and the interactions between the main factors and the IBW were removed if not significant. The ANOVA was conducted using a complete randomised design procedure, in a 2 × 2 factorial arrange, using the Nortest package of the R program (A × B; Factor A – two energy supplementation sources: corn and citrus pulp; Factor B – two supplementation times: 0900 and 1700 hours). Treatments were considered fixed effects, whereas paddocks and periods were random effects.
For variables including nutrient intake and digestibility, ENU, urine, ruminal and blood parameters, we considered cannulated animals and periods as experimental units (two cannulated animals/treatment). ANOVA was conducted in a double 4 × 4 Latin square design, with repeated measures over time (lme), considering four experimental periods, to assess collection times (h), using the Agricolae package of the R program (R Core Team 2018). Latin square, the animal within a square and collection times were considered fixed effects. The period was considered random effects. The best covariance structure used for repeated-measures analyses was chosen as the one that achieved the lowest corrected Akaike and Bayesian information, significant effects for treatment were declared at P < 0.05. When a significant effect was found, the means of the four treatments were compared by the Tukey honest significance test test at 5% probability. When the period was significant, orthogonal polynomial contrasts were used to identify the effect of periods.
Results
Total intake and digestibility of nutrients
The sources and times of energy supplementation did not interfere with the intake and digestibility of nutrients from forage and supplement (P > 0.05) (Table 4). However, the ratio g CP/kg DOM was lower in animals that received corn compared with citrus pulp both times (P = 0.002), with no difference among times for the same source (P > 0.05), both for corn and citrus pulp, neither between the sources at 0900 hours (P > 0.05).

|
Efficiency of N use and microbial protein synthesis
The intake, excretion, retention of N, as well as the excretion of urea by cannulated animals were similar among treatments (P > 0.05; Table 5). In addition, the microbial protein synthesis, the g MCP/kg DOM ratio and the ENU by the animals were not affected by the sources and/or time of energy supplementation (P > 0.05).

|
Ruminal parameters
The N-NH3 of the animals supplemented with corn was higher than citrus pulp (P = 0.005), with means of 14.92 and 13.03 mg/dL, respectively, and the supplementation at 0900 hours promoted lower N-NH3 compared with 1700 hours (P = 0.052). During collection times, N-NH3 values differed both between sources and between supplementation times (P < 0.001); however, there were no interactions between sources, supplementation times and collection times (P = 0.712; Table 6). The peak of N-NH3 occurred at approximately 1800 hours in animals supplemented with corn at 0900 and 1700 hours, and with citrus pulp at 0900 hours.

|
The pH of animals supplemented with citrus pulp was, on average, higher than those supplemented with corn (P = 0.021), but similar between animals supplemented at 0900 and 1700 hours (P = 0.969; Table 6). There was a difference between collection times (P < 0.001); however, there was no interaction between sources and/or supplementation times and collection times (P > 0.05).
In general, animals supplemented with corn in the morning kept the pH consistently lower than those receiving citrus pulp at 0900 hours. In addition, young bulls supplemented with citrus pulp at 1700 hours had a pH range higher than those that received corn at 1700 hours. The lowest pH value occurred around 2100 hours in all supplementation strategies, and animals supplemented in the afternoon had a ruminal pH peak at midday.
Among the SCFAs evaluated, only formic and valeric acids showed differences between sources of energy supplementation (P = 0.005 and P = 0.042, respectively), being lower when the animals were supplemented with corn in relation to the citrus pulp (Table 6). Furthermore, there was an interaction between energy sources and collection times in the concentration of formic acid (P = 0.042). The concentrations of lactic, propionic, butyric and isovaleric acids differed between collection times (P < 0.05). There was no difference in SCFAs concentrations between supplementation times (P > 0.05).
Dry matter in vitro degradability
Fraction A of citrus pulp isolated or associated with forage was higher than that of corn, isolated or associated, unlike Fraction B, which was lower in citrus pulp in both cases (Fig. 1). In contrast, the indigestible and unavailable Fraction (C) of citrus pulp isolated was higher than corn, but when associated with forage, both ingredients presented the same amount of Fraction C. The degradation rate (kd) of citrus pulp separated or associated with forage was higher than corn in both situations. The potential degradability of isolated corn was higher than citrus pulp; however, when associated with forage, the forage + corn mixture had lower potential degradability than the forage + citrus pulp mixture. Estimates of degradability at 2, 5 and 8 h revealed that both citrus pulp and the combination forage + citrus pulp had greater potential for DM degradation than corn separated or combined with pasture.

|
When comparing the in vitro degradability profile of corn and citrus pulp isolated, we observed that up to 24 h after incubation, the citrus pulp was greater than corn (P < 0.0001) and, consequently, presented higher gas production (P = 0.0002; Fig. 2). However, between 48 and 72 h of incubation, DM degradation of corn was greater than citrus pulp (P = 0.0001), as well as its gas production (P < 0.0001).

|
By associating forage with the supplement in the actual proportions of animal intake (88% of forage and 12% of supplement), we found that in the first 3 h of incubation, the DM degradation was the same between forage + corn and forage + citrus pulp (P = 0.499), whereas between 6 and 24 h, the association of pasture with citrus pulp showed greater degradation (P = 0.001; Fig. 2). At 48 h after incubation, however, both associations showed the same degradation (P = 0.785). At the same time, gas production remained similar in the first 3 h (P = 0.227), being higher in the forage + citrus pulp association between 6 and 12 h (P = 0.014), and between 24 and 48 h it was similar between forage + corn and forage + citrus pulp (P = 0.244).
Animal performance and gain per area
The F1 Angus × Nellore young bulls supplemented with corn or citrus pulp at 0900 or 1700 hours had similar IBW, FBW and ADG (P > 0.05; Table 7). The gain per area and stocking rate were also similar between sources and times of energy supplementation (P > 0.05).

|
Discussion
Animal performance and gain per area
The average gain per area in this study, 743.3 kg BW/ha, is higher than the average reported by Ruggieri et al. (2020), 525 kg BW/ha, when evaluating cattle reared in palisade grass pastures fertilised with N and managed at 25 cm of height, and supplemented with mineral salt ad libitum. Furthermore, the results of this study are above those recorded by Casagrande et al. (2011), of 621 kg BW/ha, for Nellore cattle reared in palisade grass pastures managed at 25 cm and supplemented at 0.3% with protein-energy based on citrus pulp and cottonseed meal. Petty et al. (1998) recorded values even higher (1570–2110 kg BW/ha), with a pangola grass–leucaena-based pasture with or without supplements of corn.
It was expected that 0.3% BW supplementation would not interfere with the stocking rate and, consequently, in the forage mass, because the effect of replacing forage with supplement (substitution) and the consequent increase in the carrying capacity of pastures under supplementation level occurs from 0.8% BW (Euclides et al. 2001). Additionally, under the adopted management criteria, even if the animals stopped consuming forage to consume the supplement, other animals would be added into the system to maintain the proposed pasture height and, thus, the canopy structure. The average stocking rate in this study, 4.6 AU/ha, was 4.3 times higher than the national livestock average of 1.06 AU/ha (ABIEC 2020), emphasising that fertilisation with high N doses and better management of sward structure when using the put-and-take technique can increase the stocking rate.
The ADG of bulls ranged from 0.86 to 1.06 kg/day, and did not differ among treatments. As the palisade grass pastures were equally fertilised with 180 kg N/ha/year, and managed at 25 cm of height in a continuous stocking system and variable stocking rate, the pastures presented similar chemical and morphological composition. Additionally, even though supplements had different chemical compositions, the total nutrient intake by animals was similar among treatments. Consequently, there was no effect of treatments on ADG.
Hoffmann et al. (2021) showed that low levels (0.3% BW) of a corn-based supplement increases ADG by approximately 33% when compared with mineral salt ad libitum, the means were 0.83 kg/day and 1.10 kg/day, respectively. The ADG values recorded here are higher than these values, and demonstrate what high-quality pastures and supplements can achieve, and citrus pulp appears to be a suitable replacement for corn as a supplement on these pasture types. Compared with other studies, the ADG obtained in this experiment is close to that recorded by Hoffmann et al. (2021), at 1.1 kg/day, by Nellore bulls reared on palisade grass pastures managed at 25 cm and supplemented at 0.3% BW with concentrate based on cottonseed meal and/or DDG. Furthermore, in the same experimental area, Koscheck et al. (2020) found an average of 0.94 kg/day by Nellore bulls supplemented at 0.3% BW with energy and protein based on citrus pulp, close to that study.
Considering the protein and energy requirements determined by BR-Corte (2016), under the experimental conditions of pasture and supplements provided, which generated an average intake of 0.98 kg CP/day and 4.10 kg TDN/day by forage, added to 0.10 kg CP/day and 0.87 kg TDN/day by the supplement, the predicted ADG for the animals would be 0.98 kg/day, which corresponds to the actual ADG of animals in the experiment, of 0.97 kg/day.
Our ADG results also corroborate with the NASEM (2016), which predicts an average ADG of 0.96 kg/day, considering an intake of 2.2% BW of a diet composed of 88% forage and 12% of energy supplement (corn or citrus pulp), with 61.6% of TDN that provided metabolisable energy approximately 9.33 MJ/kg of DM.
Total intake and digestibility of nutrients
Despite differences in the chemical composition of the supplements, the total intake of nutrients from forage and supplements was similar between the animals that received corn and citrus pulp. However, the g CP/kg DOM ratio was higher in the treatment of citrus pulp compared with corn in both supplementation times.
Poppi and McLennan (1995) suggested that the maximum efficiency in the microbial protein synthesis and transfer of ingested protein to the intestine is reached when amounts <160 g CP/kg DOM are observed, whereas values >210 g CP/kg DOM result in losses of net CP transfer from ingested CP to the passage of CP to the small intestine. In our study, however, it was expected that the g CP/kg DOM ratio was high due to the high soluble protein fraction from pastures fertilised with high doses of N, which justified the adoption of energy supplementation as a way to capture excess N from the forage in the rumen by providing fermentable substrate containing starch or pectin, depending on the source.
Thus, considering forage and supplement intakes, the g CP/kg DOM ratio is above the proposed limit of 160 g CP/kg DOM in all treatments to obtain maximum use of forage nutrients and microbial protein synthesis. Supplementation with corn generated the lowest quotients compared with citrus pulp (197.8 and 183.4 g CP/kg DOM, respectively, to corn at 0900 hours and corn at 1700 hours).
As digestibility is determined as a function of the difference between what was consumed through forage and supplement, and excreted in the faeces of animals using iNDF as an internal marker (Detmann et al. 2001), in this study, nutrient digestibility was similar in all treatments, mainly due to similarity in the intake of forage and supplement of grazing animals and iNDF content.
From a nutritional point of view, DMD exerts an important influence on the animals’ forage intake, so the higher the DMD, the greater the amount of DM that can be ingested, especially if the CP contents are >7% (Van Soest 1994) and if the ADF values are <40%, as it is the component with the greatest influence on digestibility (Figueira et al. 2015). Thus, the digestibility values found in this study can be considered within the expected and adequate range not to decrease the animals’ intake.
Digestibility and CP content was high for these pastures, and total tract digestibility was also high for a tropical pasture supplemented at these low levels with fermentable energy sources. This is a consequence of pasture management, and reinforces why pasture forage mass and structure, and fertilisation are important agronomic tools to present a high-quality pasture to grazing animals. Both energy supplement sources are high in digestibility and fermentable sources of starch or pectin. These values result in the high ADG recorded for a tropical pasture system.
Efficiency of N use and microbial protein synthesis
As there were no differences in the total intake of DM and CP in the four supplementation strategies, the amount of N ingested by the animals was also similar between treatments, as well as the amount of N excreted, which was determined from the multiplication of concentration of N in the urine by the urinary volume of each animal. Consequently, the retention of N by the animals was similar between treatments, as it is primarily due to the excretion of N in the urine (Atkinson et al. 2013).
The ENU by ruminants indicates how much available N is being used by microorganisms and whole-body metabolism, calculated by the difference between ingested and excreted N in faeces (N balance), divided by ingested N (Detmann et al. 2014). As there was no difference in N excreted and N ingested, ENU was similar between treatments. ENU is also affected by the efficiency of absorbed amino acids, which is the function of the metabolisable protein/metabolisable energy ratio (Poppi 1990).
The efficiency of MCP depends on the availability of fermentable carbohydrates and N in the rumen. The microbial growth is maximised when there is synchronism between the degradation of carbohydrates and protein in the rumen environment (NASEM 2016). The absence of differences between sources and times of energy supplementation in MCP suggests that both corn and citrus pulp supplied in the morning or the afternoon have the same potential to increase the uptake of N in the rumen as a microbial protein to reduce the excretion of N in faeces and urine. It is worth emphasising the importance of well-managed tropical pastures on the efficiency of microbial synthesis due to the high content of soluble N of forage and high-quality fibre, in addition to the low content of iNDF.
In general, citrus pulp can be an alternative energy source to corn without causing an increase in N excretions by the animal and, consequently, an increase in nitrous oxide production, nitrate leaching and NH3 emission volatilised, which can be harmful to the environment (Cardoso et al. 2016). In addition, citrus pulp is not directly consumed by humans, as it is a byproduct of the agroindustry and is less expensive than corn, which makes its use advantageous.
Ruminal parameters
As expected, supplementation with citrus pulp showed lower concentrations of N-NH3 during the day compared with supplementation with corn. These results corroborate other studies, which reported that citrus pulp could promote greater and faster capture of NH3 for its degradation, reducing ruminal N-NH3 and protein degradation (Costa et al. 2011; Alvarez Almora et al. 2012; De Oliveira et al. 2016). The results of our study and Alvarez Almora et al. (2012) support the hypothesis on the effects of synchronisation of fermentable energy and N in the rumen in decreasing post-feeding concentrations of N-NH3.
According to Detmann et al. (2014), minimum concentrations of 8 mg/dL of N-NH3 in rumen fluid are ideal for ruminal microorganisms to degrade fibre from forage efficiently. In our study, all treatments presented concentrations of N-NH3 above the minimum value for adequate rumen function and animal intake throughout the day.
The concentration of N-NH3 in the rumen varies depending on the amount and rate of degradation of the protein source and uptake by microbes, and absorption across the rumen wall (Poirier et al. 2017). In this sense, the N-NH3 peak obtained in the period between 1200 and 1800 hours coincides with the peak of animals grazing, observed by Páscoa (2009) and Casagrande et al. (2011). Thomsom et al. (1985) showed that the highest proportion of daily intake occurred in this period in grazing sheep. That is, the animals spent the afternoon consuming forage with a high amount of soluble N, the excess N of which was in part captured by the energy substrate from the supplement for microbial protein synthesis, mainly when citrus pulp was supplied, as it presented the lower N-NH3 peaks when compared with corn.
Thus, supplementation with citrus pulp in the morning or the afternoon can maximise the use of N from high-nutritive value forage, as it reduces N-NH3 concentrations in the rumen by promoting a greater capture of the forage N, which is usually in excess due to the fertilisation.
The replacement of starch sources in the diet with sugars increases the ruminal pH (Chamberlain et al. 1993; Penner et al. 2009), and in the current study, supplement with a pectin source (citrus pulp) similarly maintained a higher rumen pH than corn.
The critical pH level of 6.2 was established as the minimum value to avoid the inhibition of cellulolytic bacteria and, thus, the reduction in forage cell wall degradation (Hoover 1986). In our study, however, citrus pulp promoted an average pH of 6.2 throughout the day, whereas corn maintained an average pH of 6.05. The animals were not at risk of acidosis due to the low inclusion of starch in the diet, and were above the critical range to cause animal disorders; that is, <5.6 for >3 h (Gozho et al. 2005). The pH values found in this study were close to the means reported by Ferrari et al. (2021) and Delevatti et al. (2019b), of 6.3, by beef cattle reared on palisade grass pastures managed at 25 cm height and fed with corn-based supplement at 0.3% BW.
The intense forage intake in the afternoon contributed to the reduction in the ruminal pH, especially between 1800 and 2100 hours, when the lowest pH peak occurred, most likely due to an extensive fermentation and SCFA production in the rumen, which intensifies up to 6 h after ingestion (Owens and Goetsch 1986). However, the pH values were not low enough to reduce NDF digestibility and degradability (Delevatti et al. 2019b).
The molar proportion of SCFA in the rumen may differ due to microbial species, microbial growth rates, substrate type and availability, level of DM intake, and ruminal pH (Schären et al. 2016). In this study, however, there was no effect of sources (starch or pectin) and times on the concentration of the main SCFA, with the average acetate:propionate:butyrate ratio being 63:21:16, close to that stipulated by Wanapat et al. (2014) for forage-based diets, from 65:25:10. The low level of supplement would not be expected to affect SCFA concentration (Van Soest et al. 1991).
Pectin fermentation increases acetate production, and generally does not increase lactic and propionic acid production during fermentation, in addition to maintaining a higher rumen pH than starch fermentation, which increases propionate production (Owens and Basalan 2016). There was no difference in the concentration of acetic and lactic acid between sources or supplementation timing. However, it was found that citrus pulp promoted a higher pH than corn throughout the day at both times. As Yang et al. (2004) reported, the lower pH of corn-supplemented bulls was due to the production of lactate after starch digestion.
The formic acid concentration was higher in animals that received citrus pulp than in corn. However, contrary to what is generally observed, the pH remained higher in this source throughout the day, and there was no influence of formic acid in the production of lactic acid, as reported by Ítavo et al. (2000). Citrus pulp provided a greater concentration of valeric acid compared with corn. Nevertheless, as it makes up a small proportion of the total SCFA produced, it usually does not generate critical information for studies, as it is often not evaluated in routine diagnoses, which makes it of secondary importance in evaluating ruminal fermentation (Filípek and Dvořák 2009).
Dry matter in vitro degradability
Experimental pastures had similar DM degradability profiles among treatments, considering their similarity in chemical and structural compositions. When evaluating corn and citrus pulp separately, however, it was noted that ruminal parameters associated with the ingredients were different. The potential degradability, which does not consider the passage rate, was higher in corn compared with citrus pulp when analysed separately. This suggests that corn has a more degradable carbohydrate fraction in the rumen, but may not affect microbial efficiency (Santos and Mendonça 2006).
Pectin is approximately 95% degraded in the rumen, whereas ruminal starch degradation varies from 50 to 85%, depending on the starch source and degree of processing (Santos and Mendonça 2006). In our study, similarly to the forage + citrus pulp, citrus pulp alone had greater effective degradability than corn, especially in the first 24 h after incubation. Such behaviour was probably due to the readily fermentable status of pectin, as reported by Moreira et al. (2009), characterised as a structural carbohydrate with high and rapid rumen degradation (Van Soest et al. 1991).
In general, the total gas production (mL) of citrus pulp, isolated or associated with pasture, was greater than that of corn in both circumstances. As reported by Miron et al. (2001) and Tedeschi et al. (2009), gas production is directly related to the rate of ruminal fermentation, probably due to the higher NFC concentration that ferments faster than fibre. In this sense, despite being a fibrous carbohydrate, pectin has a fermentative behaviour similar to NFC (Homem Junior et al. 2017).
Supplementation at 0.3% BW with corn or citrus pulp at 0900 or 1700 hours can optimise the performance of F1 Angus × Nellore young bulls reared in palisade grass pastures fertilised with 180 kg N/ha/year and managed at 25 cm of height. Citrus pulp can be used as an alternative energy source to corn, and it has the same potential for animal performance, ENU and microbial protein synthesis in tropical pastures with a high proportion of soluble protein. Because citrus pulp is a byproduct of agroindustry and inedible for humans, it is less expensive than corn and does not show large price fluctuations throughout the year. The management of palisade grass pastures at 25 cm of height, with fertilisation of 180 kg N/ha/year, grazing efficiency of 50% and energy supplementation at 0.3% BW during the rainy season, promotes high production and allowance of high-quality forage, and allows a high stocking rate and production per animal and per hectare, in addition to avoiding long-term degradation of pastures.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This work was supported by the São Paulo Research Foundation (FAPESP, grants nos. 2019/25997-4; 2021/06521-9; 2020/08154-0; 2020/06679-9); and the National Council for Scientific and Technological Development (CNPq, grant 130983/2019-5).
References
ABIEC (2020) Beef report: Perfil da Pecuária no Brasil. ABIEC.Aboagye IA, Oba M, Castillo AR, Koenig KM, Iwaasa AD, Beauchemin KA (2018) Effects of hydrolyzable tannin with or without condensed tannin on methane emissions, nitrogen use, and performance of beef cattle fed a high-forage diet. Journal of Animal Science 96, 5276–5286.
| Effects of hydrolyzable tannin with or without condensed tannin on methane emissions, nitrogen use, and performance of beef cattle fed a high-forage diet.Crossref | GoogleScholarGoogle Scholar |
Alvarez Almora EG, Huntington GB, Burns JC (2012) Effects of supplemental urea sources and feeding frequency on ruminal fermentation, fiber digestion, and nitrogen balance in beef steers. Animal Feed Science and Technology 171, 136–145.
| Effects of supplemental urea sources and feeding frequency on ruminal fermentation, fiber digestion, and nitrogen balance in beef steers.Crossref | GoogleScholarGoogle Scholar |
AOAC (2012) ‘AOAC official methods of analysis.’ 18th edn. (AOAC International: Gaithersburg, MD)
Ariza P, Bach A, Stern MD, Hall MB (2001) Effects of carbohydrates from citrus pulp and hominy feed on microbial fermentation in continuous culture. Journal of Animal Science 79, 2713–2718.
| Effects of carbohydrates from citrus pulp and hominy feed on microbial fermentation in continuous culture.Crossref | GoogleScholarGoogle Scholar |
Atkinson RL, Toone CD, Robinson TJ, Harmon DL, Ludden PA (2013) Effects of ruminal protein degradability and frequency of supplementation on nitrogen retention, apparent digestibility, and nutrient flux across visceral tissues in lambs fed low-quality forage. Journal of Animal Science 88, 727–736.
| Effects of ruminal protein degradability and frequency of supplementation on nitrogen retention, apparent digestibility, and nutrient flux across visceral tissues in lambs fed low-quality forage.Crossref | GoogleScholarGoogle Scholar |
Barbero RP, Malheiros EB, Araújo TLR, Nave RLG, Mulliniks JT, Berchielli TT, Ruggieri AC, Reis RA (2015) Combining Marandu grass grazing height and supplementation level to optimize growth and productivity of yearling bulls. Animal Feed Science and Technology 209, 110–118.
| Combining Marandu grass grazing height and supplementation level to optimize growth and productivity of yearling bulls.Crossref | GoogleScholarGoogle Scholar |
Barbosa AM, Valadares RFD, Valadares Filho SdC, Véras RML, Leão MI, Detmann E, Paulino MF, Marcondes MI, Souza MAd (2006) Efeito do período de coleta de urina, dos níveis de concentrado e de fontes protéicas sobre a excreção de creatinina, de ureia e de derivados de purina e a produção microbiana em bovinos Nelore. Revista Brasileira de Zootecnia 35, 870–877.
| Efeito do período de coleta de urina, dos níveis de concentrado e de fontes protéicas sobre a excreção de creatinina, de ureia e de derivados de purina e a produção microbiana em bovinos Nelore.Crossref | GoogleScholarGoogle Scholar |
Barthram GT (1985) Experimental techniques: the HFRO sward stick. Biennial report, HFRO, Midlothian. pp. 29–30.
BR-Corte (2016) Exigências nutricionais de zebuínos puros e cruzados. In 3rd edn. (Eds Valadares Filho SC, Silva LFC, Gionbelli MP, Rotta PP, Marcondes MI, Chizzotti ML, Prados LF) p. 327. (UFV, DZO: Viçosa, Brazil)
Bueno ICS, Cabral Filho SLS, Gobbo SP, Louvandini H, Vitti DMSS, Abdalla AL (2005) Influence of inoculum source in a gas production method. Animal Feed Science and Technology 123–124, 95–105.
| Influence of inoculum source in a gas production method.Crossref | GoogleScholarGoogle Scholar |
Bueno ICS, Vitti DMSS, Louvandini H, Abdalla AL (2008) A new approach for in vitro bioassay to measure tannin biological effects based on a gas production technique. Animal Feed Science and Technology 141, 153–170.
| A new approach for in vitro bioassay to measure tannin biological effects based on a gas production technique.Crossref | GoogleScholarGoogle Scholar |
Cardoso AS, Berndt A, Leytem A, Alves BJR, de Carvalho IdNO, de Barros Soares LH, Urquiaga S, Boddey RM (2016) Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use. Agricultural Systems 143, 86–96.
| Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use.Crossref | GoogleScholarGoogle Scholar |
Cardoso AS, José Neto A, Azenha MV, Morgado ES, Brito LDF, Janusckiewicz ER, Berchielli TT, Reis RA, Ruggieri AC (2019) Mineral salt intake effects on faecal-N concentration and the volume and composition of beef cattle urine. Tropical Animal Health and Production 51, 171–177.
| Mineral salt intake effects on faecal-N concentration and the volume and composition of beef cattle urine.Crossref | GoogleScholarGoogle Scholar |
Casagrande DR, Ruggieri AC, Moretti MH, Berchielli TT, Vieira BR, Toledo AP, Roth P, Reis RA (2011) Sward canopy structure and performance of beef heifers under supplementation in Brachiaria brizantha cv. Marandu pastures maintained with three grazing intensities in a continuous stocking system. Revista Brasileira de Zootecnia 40, 2074–2082.
| Sward canopy structure and performance of beef heifers under supplementation in Brachiaria brizantha cv. Marandu pastures maintained with three grazing intensities in a continuous stocking system.Crossref | GoogleScholarGoogle Scholar |
Chamberlain DG, Robertson S, Choung J-J (1993) Sugars versus starch as supplements to grass silage: effects on ruminal fermentation and the supply of microbial protein to the small intestine, estimated from the urinary excretion of purine derivatives, in sheep. Journal of the Science of Food and Agriculture 63, 189–194.
| Sugars versus starch as supplements to grass silage: effects on ruminal fermentation and the supply of microbial protein to the small intestine, estimated from the urinary excretion of purine derivatives, in sheep.Crossref | GoogleScholarGoogle Scholar |
Chizzotti ML, Valadares Filho SC, Valadares RFD, Chizzotti FHM, Tedeschi LO (2008) Determination of creatinine excretion and evaluation of spot urine sampling in Holstein cattle. Livestock Science 113, 218–225.
| Determination of creatinine excretion and evaluation of spot urine sampling in Holstein cattle.Crossref | GoogleScholarGoogle Scholar |
Costa VAC, Detmann E, Fonseca Paulino M, Valadares Filho SdC, Carvalho IPCd, Monteiro LP (2011) Consumo e digestibilidade em bovinos em pastejo durante o período das águas sob suplementação com fontes de compostos nitrogenados e de carboidratos. Revista Brasileira de Zootecnia 40, 1788–1798.
| Consumo e digestibilidade em bovinos em pastejo durante o período das águas sob suplementação com fontes de compostos nitrogenados e de carboidratos.Crossref | GoogleScholarGoogle Scholar |
Costa DFA, Da Silva SC, Bittar CM, Takiya CS, Dórea JRR, Del Valle TA, Malafaia P, Santos FAP (2019) Citrus pulp-based supplement reduces the detrimental effects of high grazing pressure on the performance of beef cattle under a rotational system of Urochloa brizantha. Revista Brasileira de Saúde e Produção Animal 20, e0362019
| Citrus pulp-based supplement reduces the detrimental effects of high grazing pressure on the performance of beef cattle under a rotational system of Urochloa brizantha.Crossref | GoogleScholarGoogle Scholar |
de Araújo TLDR, da Silva WL, Berça AS, Cardoso AdS, Barbero RP, Romanzini EP, Reis RA (2021) Effects of replacing cottonseed meal with corn dried distillers’ grain on ruminal parameters, performance, and enteric methane emissions in young nellore bulls reared in tropical pastures. Animals 11, 2959
| Effects of replacing cottonseed meal with corn dried distillers’ grain on ruminal parameters, performance, and enteric methane emissions in young nellore bulls reared in tropical pastures.Crossref | GoogleScholarGoogle Scholar |
De Oliveira AP, Casagrande DR, Bertipaglia LMA, Barbero RP, Berchielli TT, Ruggieri AC, Reis RA (2016) Supplementation for beef cattle on Marandu grass pastures with different herbage allowances. Animal Production Science 56, 123–129.
| Supplementation for beef cattle on Marandu grass pastures with different herbage allowances.Crossref | GoogleScholarGoogle Scholar |
Delevatti LM, Cardoso AS, Barbero RP, Leite RG, Romanzini EP, Ruggieri AC, Reis RA (2019a) Effect of nitrogen application rate on yield, forage quality, and animal performance in a tropical pasture. Scientific Reports 9, 7596
| Effect of nitrogen application rate on yield, forage quality, and animal performance in a tropical pasture.Crossref | GoogleScholarGoogle Scholar |
Delevatti LM, Romanzini EP, Koscheck JFW, da Ross de Araujo TL, Renesto DM, Ferrari AC, Barbero RP, Mullinikis JT, Reis RA (2019b) Forage management intensification and supplementation strategy: intake and metabolic parameters on beef cattle production. Animal Feed Science and Technology 247, 74–82.
| Forage management intensification and supplementation strategy: intake and metabolic parameters on beef cattle production.Crossref | GoogleScholarGoogle Scholar |
Detmann E, Paulino MF, Zervoudakis JT, Valadares Filho SdC, Lana RdP, Queiroz DSd (2001) Suplementação de novilhos mestiços durante a época das águas: parâmetros ingestivos e digestivos. Revista Brasileira de Zootecnia 30, 1340–1349.
| Suplementação de novilhos mestiços durante a época das águas: parâmetros ingestivos e digestivos.Crossref | GoogleScholarGoogle Scholar |
Detmann E, Souza MA, Valadares Filho SC, Queiroz AC, Berchielli TT, Saliba EOS, Cabral LS, Pina DS, Ladeira MM, Azevedo JAG (2012) ‘Métodos para análise de alimentos – Instituto Nacional de Ciência e Tecnologia de Ciência Animal (INCT).’ (Suprema: Visconde do Rio Branco, Brazil)
Detmann E, Valente EEL, Batista ED, Huhtanen P (2014) An evaluation of the performance and efficiency of nitrogen utilization in cattle fed tropical grass pastures with supplementation. Livestock Science 162, 141–153.
| An evaluation of the performance and efficiency of nitrogen utilization in cattle fed tropical grass pastures with supplementation.Crossref | GoogleScholarGoogle Scholar |
Embrapa (2013) ‘Sistema Brasileiro de classificação de solos.’ 3rd edn. (Embrapa: Brasília, Brazil)
Euclides VPB, Euclides Filho K, Paim Costa F, Figueiredo GR (2001) Desempenho de novilhos F1s Angus-Nelore em pastagens de Brachiaria decumbens submetidos a diferentes regimes alimentares. Revista Brasileira de Zootecnia 30, 470–481.
| Desempenho de novilhos F1s Angus-Nelore em pastagens de Brachiaria decumbens submetidos a diferentes regimes alimentares.Crossref | GoogleScholarGoogle Scholar |
Fenner H (1965) Method for determining total volatile bases in rumen fluid by steam distillation. Journal of Dairy Science 48, 249–251.
| Method for determining total volatile bases in rumen fluid by steam distillation.Crossref | GoogleScholarGoogle Scholar |
Ferrari AC, Leite RG, Fonseca NVB, Romanzini EP, Cardoso AS, Barbero RP, Costa DFA, Ruggieri AC, Reis RA (2021) Performance, nutrient use, and methanogenesis of Nellore cattle on a continuous grazing system of Urochloa brizantha and fed supplement types varying on protein and energy sources. Livestock Science 253, 104716
| Performance, nutrient use, and methanogenesis of Nellore cattle on a continuous grazing system of Urochloa brizantha and fed supplement types varying on protein and energy sources.Crossref | GoogleScholarGoogle Scholar |
Figueira DN, Neumann M, Ueno RK, Muller MML, Faria MV (2015) Produção e composição química do capim elefante cv. pioneiro em diferentes alturas de resíduo - revisão de literatura. Revista Brasileira de Tecnologia Aplicada nas Ciências Agrárias 8, 103–110.
| Produção e composição química do capim elefante cv. pioneiro em diferentes alturas de resíduo - revisão de literatura.Crossref | GoogleScholarGoogle Scholar |
Filípek J, Dvořák R (2009) Determination of the volatile fatty acid content in the rumen liquid: comparison of gas chromatography and capillary isotachophoresis. Acta Veterinaria Brno 78, 627–633.
| Determination of the volatile fatty acid content in the rumen liquid: comparison of gas chromatography and capillary isotachophoresis.Crossref | GoogleScholarGoogle Scholar |
Fonseca NVB, Cardoso AdS, Berça AS, Dornellas IA, Ongaratto F, Silva MLC, Ruggieri AC, Reis RA (2022) Effect of different nitrogen fertilizers on nitrogen efficiency use in Nellore bulls grazing on Marandu palisade grass. Livestock Science 263, 105012
| Effect of different nitrogen fertilizers on nitrogen efficiency use in Nellore bulls grazing on Marandu palisade grass.Crossref | GoogleScholarGoogle Scholar |
Gozho GN, Plaizier JC, Krause DO, Kennedy AD, Wittenberg KM (2005) Subacute ruminal acidosis induces ruminal lipopolysaccharide endotoxin release and triggers an inflammatory response. Journal of Dairy Science 88, 1399–1403.
| Subacute ruminal acidosis induces ruminal lipopolysaccharide endotoxin release and triggers an inflammatory response.Crossref | GoogleScholarGoogle Scholar |
Hoffmann A, Cardoso AS, Fonseca NVB, Romanzini EP, Siniscalchi D, Berndt A, Ruggieri AC, Reis RA (2021) Effects of supplementation with corn distillers’ dried grains on animal performance, nitrogen balance, and enteric CH4 emissions of young Nellore bulls fed a high-tropical forage diet. Animal 15, 100155
| Effects of supplementation with corn distillers’ dried grains on animal performance, nitrogen balance, and enteric CH4 emissions of young Nellore bulls fed a high-tropical forage diet.Crossref | GoogleScholarGoogle Scholar |
Homem Junior AC, Ezequiel JMB, Fávaro VR, Almeida MTC, Paschoaloto JR, D’Áurea AP, Carvalho VBd, Nocera BF, Cremasco LF (2017) Methane production by in vitro ruminal fermentation of feed ingredients. Semina: Ciências Agrárias 38, 877–884.
| Methane production by in vitro ruminal fermentation of feed ingredients.Crossref | GoogleScholarGoogle Scholar |
Hoover WH (1986) Chemical factors involved in ruminal fiber digestion. Journal of Dairy Science 69, 2755–2766.
Hopper JT, Holloway JW, Butts WT (1978) Animal variation in chromium sesquioxide excretion patterns of grazing cows. Journal of Animal Science 46, 1096–1102.
| Animal variation in chromium sesquioxide excretion patterns of grazing cows.Crossref | GoogleScholarGoogle Scholar |
Ítavo LCV, Santos GTd, Jobim CC, Voltolini TV, Bortolassi JR, Ferreira CCB (2000) Aditivos na conservação do bagaço de laranja in natura na forma de silagem. Revista Brasileira de Zootecnia 29, 1474–1484.
| Aditivos na conservação do bagaço de laranja in natura na forma de silagem.Crossref | GoogleScholarGoogle Scholar |
Koscheck JFW, Romanzini EP, Barbero RP, Delevatti LM, Ferrari AC, Mulliniks JT, Mousquer CJ, Berchielli TT, Reis RA (2020) How do animal performance and methane emissions vary with forage management intensification and supplementation? Animal Production Science 60, 1201–1209.
| How do animal performance and methane emissions vary with forage management intensification and supplementation?Crossref | GoogleScholarGoogle Scholar |
Leite RG, Cardoso AS, Fonseca NVB, Silva MLC, Tedeschi LO, Delevatti LM, Ruggieri AC, Reis RA (2021) Effects of nitrogen fertilization on protein and carbohydrate fractions of Marandu palisadegrass. Scientific Reports 11, 1–8.
Miron J, Yosef E, Ben-Ghedalia D (2001) Composition and in vitro digestibility of monosaccharide constituents of selected byproduct feeds. Journal of Agricultural and Food Chemistry 49, 2322–2326.
| Composition and in vitro digestibility of monosaccharide constituents of selected byproduct feeds.Crossref | GoogleScholarGoogle Scholar |
Moreira PC, Reis RB, Wascheck RdC, Rezende PLdP, Mendonça AC, Rassi LF (2009) Degradabilidade in situ das rações de vacas da raça holandês em lactação com substituição do milho por polpa cítrica. Ciencia Animal Brasileira 10, 406–412.
Mott GO, Lucas HL (1952) The design conduct and interpretation of grazing trials on cultivated and improved pastures. In ‘Proceedings of the international grassland congress’. pp. 1380–1395. (State College)
Mottet A, Teillard F, Boettcher P, De’ Besi G, Besbes B (2018) Review: Domestic herbivores and food security: current contribution, trends and challenges for a sustainable development. Animal 12, s188–s198.
| Review: Domestic herbivores and food security: current contribution, trends and challenges for a sustainable development.Crossref | GoogleScholarGoogle Scholar |
Myers WD, Ludden PA, Nayigihugu V, Hess BW (2004) Technical note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide. Journal of Animal Science 82, 179–183.
| Technical note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide.Crossref | GoogleScholarGoogle Scholar |
NASEM (2016) ‘Nutrient requirements of beef cattle.’ 8th edn. (The National Academies Press: Washington, DC)
Norris AB, Tedeschi LO, Muir JP (2019) Assessment of in situ techniques to determine indigestible components in the feed and feces of cattle receiving supplemental condensed tannins. Journal of Animal Science 97, 5016–5026.
| Assessment of in situ techniques to determine indigestible components in the feed and feces of cattle receiving supplemental condensed tannins.Crossref | GoogleScholarGoogle Scholar |
Ørskov ER, McDonald I (1979) The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. The Journal of Agricultural Science 92, 499–503.
| The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage.Crossref | GoogleScholarGoogle Scholar |
Owens FN, Basalan M (2016) Ruminal fermentation. In: ‘Rumenology’. (Eds D Millen, MDB Arrigoni, RDL Pacheco) pp. 63–102. (Springer Cham: New York City)
Owens FN, Goetsch AL (1986) Digesta passage and microbial protein synthesis. In ‘Control of digestion and metabolism in ruminants’. (Eds LP Milligan, WL Grovum, A Dobson) pp. 196–226. (Prentice Hall: London)
Páscoa AG (2009) Comportamento de bovinos de corte em resposta à disposição espacial de condicionadores de pastejo. Tese (Doutorado em Zootecnia). Faculdade de Ciências Agrárias e Veterinária – UNESP, Jaboticabal. p. 118.
Penner GB, Taniguchi M, Guan LL, Beauchemin KA, Oba M (2009) Effect of dietary forage to concentrate ratio on volatile fatty acid absorption and the expression of genes related to volatile fatty acid absorption and metabolism in ruminal tissue. Journal of Dairy Science 92, 2767–2781.
| Effect of dietary forage to concentrate ratio on volatile fatty acid absorption and the expression of genes related to volatile fatty acid absorption and metabolism in ruminal tissue.Crossref | GoogleScholarGoogle Scholar |
Petty SR, Poppi DP, Triglone T (1998) Effect of maize supplementation, seasonal temperature and humidity on the liveweight gain of steers grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia. The Journal of Agricultural Science 130, 95–105.
| Effect of maize supplementation, seasonal temperature and humidity on the liveweight gain of steers grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia.Crossref | GoogleScholarGoogle Scholar |
Poirier S, Madigou C, Bouchez T, Chapleur O (2017) Improving anaerobic digestion with support media: mitigation of ammonia inhibition and effect on microbial communities. Bioresource Technology 235, 229–239.
| Improving anaerobic digestion with support media: mitigation of ammonia inhibition and effect on microbial communities.Crossref | GoogleScholarGoogle Scholar |
Poppi DP (1990) Manipulation of nutrient supply to animals at pasture: opportunities and consequences. In ‘Proceedings of the 5th Asian-Australasian Association of Animal Production (AAAP) animal science congress, Vol. 1’. pp. 40–79. (AAAP)
Poppi DP, McLennan SR (1995) Protein and energy utilization by ruminants at pasture. Journal of Animal Science 73, 278–290.
| Protein and energy utilization by ruminants at pasture.Crossref | GoogleScholarGoogle Scholar |
Poppi DP, Mclennan RS, Bediye S, De Vega A, Zorrila-Rios J (1997) Forage quality: strategies for increasing nutritive value of forages. In ‘International grassland congress’. pp. 307–322. (Association Management Cen.)
Poppi DP, Quigley SP, Silva TACCd, McLennan SR (2018) Challenges of beef cattle production from tropical pastures. Revista Brasileira de Zootecnia 47, e20160419
| Challenges of beef cattle production from tropical pastures.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2018) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing) Available at https://www.R-project.org/ [Accessed 20 March 2021]
Ruggieri AC, Cardoso AdS, Ongaratto F, Casagrande DR, Barbero RP, Brito LdF, Azenha MV, Oliveira AA, Koscheck JFW, Reis RA (2020) Grazing intensity impacts on herbage mass, sward structure, greenhouse gas emissions, and animal performance: analysis of brachiaria pastureland. Agronomy 10, 1750
| Grazing intensity impacts on herbage mass, sward structure, greenhouse gas emissions, and animal performance: analysis of brachiaria pastureland.Crossref | GoogleScholarGoogle Scholar |
Santos FA, Mendonça A (2006) Metabolismo de proteínas. In ‘Nutrição de ruminantes’. (Eds TT Berchielli, AV Pires, SG Oliveira) p. 583. (FUNEP: Jaboticabal, Brazil)
Schären M, Seyfang GM, Steingass H, Dieho K, Dijkstra J, Hüther L, Frahm J, Beineke A, von Soosten D, Meyer U, Breves G, Dänicke S (2016) The effects of a ration change from a total mixed ration to pasture on rumen fermentation, volatile fatty acid absorption characteristics, and morphology of dairy cows. Journal of Dairy Science 99, 3549–3565.
| The effects of a ration change from a total mixed ration to pasture on rumen fermentation, volatile fatty acid absorption characteristics, and morphology of dairy cows.Crossref | GoogleScholarGoogle Scholar |
Serafim JA, Silveira RF, Vicente EF (2021) Fast determination of short-chain fatty acids and glucose simultaneously by ultraviolet/visible and refraction index detectors via high-performance liquid chromatography. Food Analytical Methods 14, 1387–1393.
| Fast determination of short-chain fatty acids and glucose simultaneously by ultraviolet/visible and refraction index detectors via high-performance liquid chromatography.Crossref | GoogleScholarGoogle Scholar |
Silva Júnior JM, Rennó LN, Valadares Filho SC, Paulino MF, Detmann E, Menezes GCC, Martins TS, Paula RM, Rodrigues JPP, Marcondes MI (2018) Evaluation of collection days and times to estimate urinary excretion of purine derivatives and nitrogen compounds in grazing Nellore cattle. Livestock Science 217, 85–91.
| Evaluation of collection days and times to estimate urinary excretion of purine derivatives and nitrogen compounds in grazing Nellore cattle.Crossref | GoogleScholarGoogle Scholar |
Sniffen CJ, O’Connor JD, Van Soest PJ, Fox DG, Russell JB (1992) A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. Journal of Animal Science 70, 3562–3577.
| A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability.Crossref | GoogleScholarGoogle Scholar |
Sollenberger LE, Cherney DJR (1995) Evaluating forage production and quality. In ‘Forages: the science of grassland agriculture’. (Eds RF Barnes, DA Miller, CJ Nelson) pp. 97–110. (Iowa State University Press: Ames, IA)
Tedeschi LO, Fox DG (2020) ‘The ruminant nutrition system: an applied model for predicting nutrient requirements and feed utilization in ruminants. Vol. I.’ 3rd edn. (XanEdu: Ann Arbor, MI, USA)
Tedeschi LO, Kononoff PJ, Karges K, Gibson ML (2009) Effects of chemical composition variation on the dynamics of ruminal fermentation and biological value of corn milling (co)products. Journal of Dairy Science 92, 401–413.
| Effects of chemical composition variation on the dynamics of ruminal fermentation and biological value of corn milling (co)products.Crossref | GoogleScholarGoogle Scholar |
Thomsom DF, Beever DE, Haines MJ, Cammell SB, Evans RT, Dhanoa MS, Austin AR (1985) Yield and composition of milk from Friesian cows grazing either perennial ryegrass or white clover in early lactation. Journal of Dairy Research 52, 17–31.
Titgemeyer EC, Armendariz CK, Bindel DJ, Greenwood RH, Löest CA (2001) Evaluation of titanium dioxide as a digestibility marker for cattle. Revista Brasileira de Zootecnia 79, 1059–1063.
| Evaluation of titanium dioxide as a digestibility marker for cattle.Crossref | GoogleScholarGoogle Scholar |
Van Raij B, Cantarella H, Quaggio JA, Furlani AM (1997) ‘Recomendações de adubação e calagem para o estado de São Paulo.’ 2. edn. rev. atual. p. 285. (Instituto Agronômico/Fundação IAC: Campinas, Brazil)
Van Soest PJ (1994) ‘Nutritional ecology of the ruminant.’ (Cornell University Press: Ithaca, NY, USA)
Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science 74, 3583–3597.
| Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition.Crossref | GoogleScholarGoogle Scholar |
Wanapat M, Gunun P, Anantasook N, Kang S (2014) Changes of rumen pH, fermentation and microbial population as influenced by different ratios of roughage (rice straw) to concentrate in dairy steers. The Journal of Agricultural Science 152, 675–685.
| Changes of rumen pH, fermentation and microbial population as influenced by different ratios of roughage (rice straw) to concentrate in dairy steers.Crossref | GoogleScholarGoogle Scholar |
Williams CH, David DJ, Iismaa O (1962) The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. Journal of Agricultural Science 59, 381–385.
| The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry.Crossref | GoogleScholarGoogle Scholar |
Yang WZ, Beauchemin KA, Vedres DD, Ghorbani GR, Colombatto D, Morgavi DP (2004) Effects of direct-fed microbial supplementation on ruminal acidosis, digestibility, and bacterial protein synthesis in continuous culture. Animal Feed Science and Technology 114, 179–193.
| Effects of direct-fed microbial supplementation on ruminal acidosis, digestibility, and bacterial protein synthesis in continuous culture.Crossref | GoogleScholarGoogle Scholar |
Young EG, Conway CF (1942) On the estimation of allantoin by the Rimini-Schryver reaction. Journal of Biological Chemistry 142, 839–853.
| On the estimation of allantoin by the Rimini-Schryver reaction.Crossref | GoogleScholarGoogle Scholar |