

Ecology and movement of urban koalas adjacent to linear infrastructure in coastal south-east Queensland
S. M. de Oliveira A , P. J. Murray B , D. L. de Villiers C and G. S. Baxter A DA University of Queensland, School of Geography, Planning and Environmental Management, St Lucia, Qld 4072, Australia.
B University of Queensland, School of Agriculture and Food Sciences, Gatton Campus, Qld 4343, Australia.
C Koala Conservation Unit, Department of Environment and Heritage Protection, Daisy Hill Road, Daisy Hill, Qld 4127, Australia.
D Corresponding author. Email: gbaxter@uqg.uq.edu.au
Australian Mammalogy 36(1) 45-54 https://doi.org/10.1071/AM12046
Submitted: 31 October 2012 Accepted: 31 August 2013 Published: 8 November 2013
Abstract
In Redland City, koalas (Phascolarctos cinereus) are in rapid decline as they are exposed to anthropogenic threats such as habitat clearing, dog attacks, vehicle collisions and disease. This study investigated the influence of linear infrastructure on the movement and habitat use of urban koalas. Seven koalas were tracked for up to 28 weeks during the breeding season. Home ranges were calculated for 95% Minimum Convex Polygon (MCP95%) and 95% fixed Kernel Density (FK95%). Koalas responded to the landscape in different ways. Linear infrastructure did not restrict the movements of most koalas. Home ranges varied from 1.1 to 31.5 ha MCP95% and from 5 to 55 ha for FK95%. Koalas mainly used Eucalyptus tereticornis throughout the study site. A variety of non-regionally endemic and regionally endemic trees in urban and remnant vegetation areas were used, suggesting that all trees are potentially koala habitat. At the completion of the study, four koalas remained alive, two were killed by trains and one died from a dog attack. Despite the small sample size and short duration, our results suggest that koalas are able to navigate linear infrastructure; however, the high rates of mortality associated with these movements puts the long-term viability of urban koala populations in doubt.
Additional keywords: dog attack, home range, movement, rail line, tree use, urban koalas, vehicle collision.
Introduction
Home ranges and habitat use of koalas (Phascolarctos cinereus) have been widely studied throughout their distribution (Mitchell 1989; Hasegawa 1995; White 1999; Cork et al. 2000; Ellis et al. 2002; Thompson 2006; Kavanagh et al. 2007; Matthews et al. 2007). Home ranges are generally 1–50 ha (Cork et al. 2000) although they are larger in some localities (Ellis et al. 2002). Tree use also varies throughout the koala’s range and includes both eucalypt and non-eucalypt species (Martin and Handasyde 1999; Phillips 1990; White 1999).
In south-east Queensland koalas use trees from different regional ecosystems (Sattler and Williams 1999). For example, the vegetation at Mutdapilly, west of Brisbane, consists of Eucalyptus tereticornis, E. crebra, E. melanophloia, Corymbia tessellaris and C. intermedia, in contrast to the Daisy Hill area, south of Brisbane, where E. propinqua dominates (Parish and Cox 2007). Soil nutrient status also plays a role in tree selection by koalas (Phillips and Callaghan 2000).
Koala populations in south-east Queensland are experiencing rapid reductions in range (Dique et al. 2003a, 2004). This decline has mostly been attributed to habitat fragmentation (Martin and Handasyde 1999) and loss through clearing for agriculture and urban development (Cogger et al. 2003; Sullivan et al. 2004; Gordon et al. 2006; Lane 2008; DERM 2009). Recently, an important role has been suggested for disease in the decline of koala populations, with chlamydial infection being implicated as the primary cause of disease (Timms 2005; Markey et al. 2007; Higgins 2008). Exacerbation of the disease may be attributed to the koala retrovirus (Tarlinton et al. 2005) and stress from habitat clearing and other anthropogenic impacts (Weigler et al. 1988; Lee et al. 2010; Brearley et al. 2013). In addition to these threats, urban koalas face increased mortality from dog attacks and vehicle collisions (Smith and Smith 1990; Melzer et al. 2000; Stratford et al. 2000; Dique et al. 2003b; McAlpine et al. 2006).
Alterations to habitat or the addition of perceived threats can cause animals to exhibit a range of responses such as flight, hide, increased vigilance or stress (Elgar 1989). These behavioural responses can trigger negative physiological changes in an animal, especially when habitat alterations lead to increased predation, competition for resources and mates, and habitat fragmentation (Brearley et al. 2013; Dowle et al. 2012), which in turn can lead to changes in movements (Strevens 2007; Asari et al. 2010). Yet koalas still persist in disturbed urban landscapes where there are resources they can use.
Wildlife displays different behavioural responses when encountering habitat fragmented by linear infrastructure, compared with intact habitat (Goosem and Marsh 1997; St. Clair and Forrest 2009; Asari et al. 2010). For example, pronghorn (Antilocapra americana) exhibit different levels of vigilance and foraging times around roads with changes in traffic volume (Gavin and Komers 2006). Differences in movement patterns have been observed in the bush rat (Rattus fuscipes) and the brown antechinus (Antechinus stuartii) underneath powerlines depending on the presence and characteristics of logs, shrub densities and leaf matter cover (Strevens 2007). In Canada there are differences in mating and sparring frequencies in elk (Cervus elaphus) along roadsides depending on road opening and closure times, and traffic volumes (St Clair and Forrest 2009). Wild mammals can successfully cross roads and power lines (e.g. mahogany gliders, Petaurus gracilis: Asari et al. 2010), but such movements often end in trauma and death (e.g. koalas: Canfield 1991; Dique et al. 2003a). Vehicle collisions have been reported as a key threat to the viability of koala populations in parts of Victoria and New South Wales (Lee and Martin 1988; Canfield 1991; Lunney et al. 1996).
In south-east Queensland the urban and rural areas of Redland City, the southern section of Brisbane City, and eastern portion of Logan City form a region known as the ‘Koala Coast’ because it has historically supported a large koala population that is genetically distinct from other koala populations in south-east Queensland (Lee et al. 2010). The Koala Coast is experiencing rapid human population growth and development (Queensland Government 2005), which has created concern for the long-term viability of the koala population. In this region estimates of koala numbers were over 6000 in 1999 (Dique et al. 2004) and ~2000 in 2010, with the population declining by ~6% per annum over that time (DERM 2010). Previous research in this region has focussed on the ecology (Thompson 2001; Dique et al. 2004), mortality (Dique et al. 2003a), and general movements (Dique et al. 2003b) of bushland koalas. However, little is known about the ecology of koalas inhabiting urban and remnant vegetation patches in Redland City.
The aim of this study was to investigate the ecology and movement of urban koalas occupying habitat within urban environments containing major linear infrastructure (specifically roads and rail lines) and residential zones during the koala breeding season.
Methods
Study site
The suburb of Ormiston in Redland City was selected for this research because it contains a high density of wild koalas, is undergoing development, and contains a State-owned rail line that bisects potential habitat in the western part of the suburb. Four kilometres of rail line traverses the suburb from Wellington Point station to Cleveland station. Ormiston is located ~23 km south-east of the Brisbane CBD. The area around the rail line contains remnant vegetation, parks, suburban yards and infrastructure.
Native vegetation was mostly open eucalypt woodland on mainly Cainozoic alluvial plains belonging to Regional Ecosystems 12.1.1, 12.3.6, 12.5.2 and 12.5.3 (Sattler and Williams 1999). This consisted mainly of Eucalyptus tereticornis, which was often surrounded by Casuarina glauca, Melaleuca quinquenervia and Allocasuarina littoralis. Other common species within the eucalypt woodlands were E. crebra and C. intermedia. E. microcorys was occasionally planted in backyards and along road easements.
Municipal parks were found throughout the study site. These areas include bushland reserves ranging from ~3 to 18 ha. Collectively, the bushland reserves and parks contain a wide variety of native trees (e.g. E. racemosa, C. intermedia, E. tereticornis, E. crebra, C. citriodora, Lophostemon suaveolens, M. quinquenervia and C. glauca). There were also smaller remnant pockets of native vegetation, generally less than 1 ha and mainly dominated by E. tereticornis with a few M. quinquenervia.
Other potential habitat consisted of non-regionally endemic and regionally endemic trees found in urban areas. These trees were classified as ‘urban trees’ where they were found as: (a) footpath trees located along linear infrastructure that were planted or naturally occurring, or (b) trees that grew in front yards or backyards of residential properties.
Capture and collaring of koalas
Seven adult koalas were caught and radio-collared within 400 m of the rail line between Wellington Point and Cleveland railway stations during four capture events between 31 July and 8 October 2009. Arterial (Sturgeon Street and Northern Arterial Road) and suburban roads were adjacent to most of this section of railway. Six koalas were initially chosen for this study but a seventh animal was captured and collared as a replacement for one that was killed three days after capture.
Before capturing the koalas each individual was visually assessed with binoculars from the ground to determine whether it was an adult in good physical condition, without back-young, and was free of overt signs of chlamydiosis such as cystitis and conjunctivitis. Suitable animals were captured by flagging the animal to the ground in a manner similar to that described by Thompson (2001) and Radford et al. (2006). Captured animals were placed in a cloth bag, measured, weighed, visually assessed for disease, given a body condition score (Allen et al. 2010) and, if suitable, collared with a single-stage VHF radio-collar with automatic drop-off feature (Sirtrack, Havelock North, New Zealand) and then released at the site of capture. All collars were programmed to automatically fall off on 31 January 2010. We timed this period to coincide with the koala breeding season (from July to January), when koalas are most active.
At the completion of the project, the surviving four koalas were admitted to the Australia Zoo Wildlife Hospital for treatment or health checks. A fifth koala was taken to Moggill Koala hospital after death. Age was estimated for these koalas by veterinarians as part of their routine health assessment. Only one koala’s collar had fallen off automatically in the field, hence this koala was not admitted for health checks after completion of the study. A necropsy was performed on one koala that died during the project to determine cause of death. Another one was not recovered from the field but was photographed; the photographs were shown to a veterinarian to confirm train strike as a likely cause of death.
Tracking of koalas
All koalas were tracked within 24 h of their release and then within seven days after release. Thereafter they were tracked twice within a 24-h period (during the day and at night) and every second day. Koalas were located on foot with an Australis 26k radio receiver (Titley Electronics, Brisbane, Australia) and a hand-held yagi aerial (Sirtrack, Havelock North, New Zealand). Once found, a GPS location, tree diameter at breast height (DBH), tree species, tree height and koala’s height in the tree were all recorded. One male (M1) koala made regular movements across the rail line and to investigate these further overnight tracking, designed to follow that koala, was conducted from Saturday night to Sunday morning from 24 October 2009 until 10 January 2010 (10 times).
Data analyses
ArcGIS software ver. 9.3.1 was used to estimate movements (in metres) and home ranges for Minimum Convex Polygon (MCP) and fixed Kernel Density Estimations (FK) with use of external extensions Abode, Hawth’s Tools and GT Geowizard. Home-range perimeters and home ranges were estimated for MCP100% and MCP95%; however, only home ranges for 95% MCP and FK are illustrated for a comparison. FK home ranges were estimated only for their core at 50% and total range at 95%. FK ranges were illustrated and measured by a single-parameter smoothing factor. During kernel analysis, we found that FK suited the data better than adaptive kernel estimations. Adaptive kernel did not fit our data and produced scattered home ranges. Single-parameter smoothing factors better defined the koalas’ home ranges than the double-parameter smoothing factors. The latter resulted in koalas having home ranges made up of scattered and disjunct areas.
Results
Three adult male (M1, M2 and M3) and four adult female (F1, F2, F3 and F4) koalas were captured in both remnant and urban areas and tracked for 3–197 days (Table 1). This resulted in three (koala F2) to 94 (koala M1) fixed locations for each animal (Table 1). Koala F2 is not considered further due to the lack of data as this animal was killed by a train just three days after its capture.

|
Home range and movements
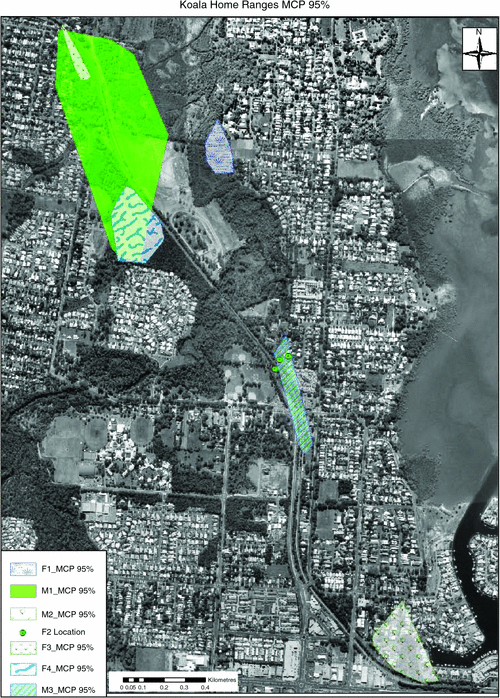
The MCP95% home ranges of the koalas varied from 1.1 to 31.5 ha (Table 2). There was overlap among some of the koalas’ home ranges (Figs 1, 2), particularly between male and female koalas (i.e. that of M1 overlapped with that of F3 and later with that of F4, and that of M3 overlapped with that of F2). Home ranges calculated at MCP95% included linear infrastructure, houses, remnant vegetation and urban trees (Fig. 1).

|
|
|
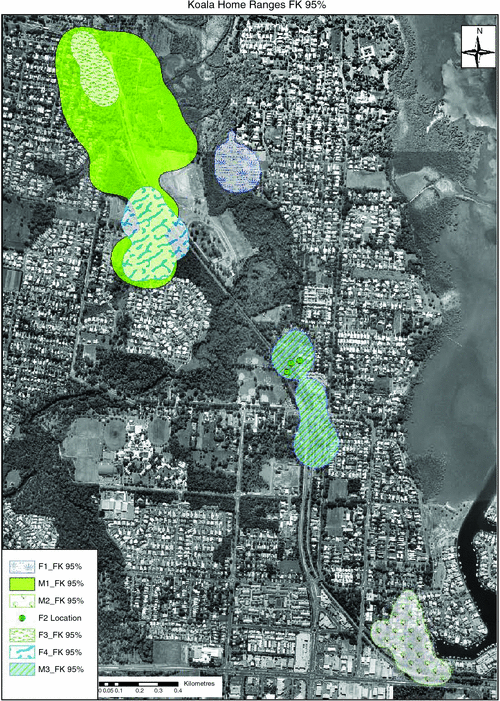
Home ranges estimated at FK95% with a single smoothing parameter (Fig. 2, Table 2) were larger than the MCP95% estimates for all koalas. Also, in some cases with the FK estimates some koalas’ home ranges included roads and the rail line; however these koalas did not have a recorded crossing of any linear infrastructure throughout the monitoring period. The MCP analysis better reflected the tracking results (Fig. 1). Signs of possible exploratory movements by M1, M3 and F1 were observed, although these movements did not exceed 100 m from their home-range boundaries. Accordingly, these fixed locations were not recorded in any of the animals’ ranges for FK50%, FK95% or MCP95%.
Most koalas stayed in any one general location for 1–4 days before moving to another area within their home range. These movements were mainly recorded at night for all koalas (Tables 1, 3). Koala movements differed greatly from a minimum movement of 7 m per night for males and 4 m for females, to a maximum of 634 m per night for males and 234 m for females (Table 3). M1 moved the greatest distance in one night between two fixed locations when he travelled over 634 m in 4 h (Table 3).

|
Five koalas had roads within their ranges and were observed to cross them regularly (Table 3, Fig. 1). Four koalas (M1, M2, M3 and F3) crossed a road at least once, with one individual (M1) crossing roads 78 times (Table 3). Koalas crossed roads at any time of the night. Direct observations showed that some animals ran quickly across the road while others meandered along it. No koalas were killed or injured on roads during this study. Road traffic was lighter at night than at other times of the day. Peak times varied widely: from 0600 until 0900 hours on weekdays and from 1015 until 1230 hours on weekends. Roads with most traffic movement were the two arterial roads (790–1488 vehicles per day) while the remaining roads, comprising suburban streets, had 60–233 vehicles per day. Road traffic between 2330 hours and 0730 hours was significantly lower, being 8–16% that of peak times (Jessie Lee, Administration Officer, Infrastructure Planning, Redland City Council 2009, pers. comm.).
Three koalas (M1, M3 and F2) crossed the rail line at least once during the study (Table 3). All recorded rail line crossings were between 2330 hours and 0500 hours. At these times rail traffic was less than 44% of the frequency of peak times (Translink 2009). Trains to Ormiston station decreased from one train every 6–16 min during the day to one every 10–21 min between 1900 hours and 0146 hours (Translink 2009). F2 successfully crossed the rail line on her first attempt but failed to cross during her second attempt (Fig. 1). These two crossing attempts occurred in one night. One male (M1) included the rail line as part of his home range, using remnant vegetation on either side of the rail line. The animal crossed the rail line at night on any day of the week, with records showing rail line crossings at various areas within the animal’s home range. During overnight tracking sessions of this animal, it was noted that before crossing the rail line the animal would climb one of a few trees along the rail fenceline and spend an average of 7 min in the tree. The animal then moved towards the rail line. On three occasions (30%) the descent of the tree coincided with the passing of a train, and subsequent crossing of the rail line was within 3 min of the train passing. The other sightings were when the animal crossed while the train was not in service (20%), the animal did not cross (40%) or on one occasion while the train was still in service when a crossing was missed (10%). It took M1 an average of 4 min to descend the tree, navigate fences and cross the rail line and enter Station Street refuge. This koala was also recorded to cross between 0200 and 0230 hours on a weekday (21 October 2009). At this time, trains were already out of service.
Other koalas (M2, F3 and F4) moved between areas within their home range but the rail line provided a home range boundary. One male koala (M3) was recorded to not only cross the rail line but also to use the trees along the narrow rail line as a corridor to move between Beckwith Street refuge and an urban area. This animal spent approximately half of its time in this urban habitat, which consisted of footpath trees flanked by Ormiston station and Sturgeon Street. Both these areas also comprised the koala’s core areas according to the FK50% during the breeding season (Fig. 2).
Habitat use
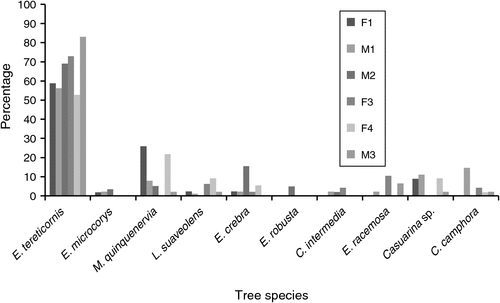
Different patterns of koala use of the vegetation were observed throughout the study site depending on the individual, the degree of fragmentation of the habitat and the variety of tree species found in each koala’s home range. F2 was not included in these results; however, before she was killed she was found only in urban environments, in E. tereticornis trees in the front or back yards of two residences (Fig. 1). All koalas used E. tereticornis with a high frequency (Fig. 3). This was also the case when we analysed tree usage of koalas in urban trees only. Koalas in general used E. tereticornis ~66% of the time in urban areas throughout the breeding season. Other eucalypt species in urban areas were used 17% of the time. Endemic species were used 10% and non-endemic species were used 7% of the time in urban areas. While these trees were being monitored, the koalas were found resting, browsing or moving from branch to branch on the same tree or the tree next to it. Koalas were also found in E. microcorys (~1–3% of fixed locations throughout the tracking period), a eucalypt species that is not part of the local Regional Ecosystems (i.e. REs 12.1.1, 12.3.6, 12.5.2 and 12.5.3). Other tree species that koalas also used included the exotic camphor laurel (Cinnamomum camphora). Koalas M1 and F3 used this species for shelter, with most of the canopies intertwined with other fodder or shelter trees (commonly E. tereticornis and M. quinquenervia), where the koalas were later found. One male (M1) and two female koalas (F1 and F4) were often located in M. quinquenervia trees. Koalas F1, M1, F4 and M3 were also often located in Casuarina species, such as C. glauca and Allocasuarina littoralis (Fig. 3). Both these tree species were found frequently in the local areas as well as within koala home ranges, and they were often the most common species within the home range.
|
Koalas spent ~20% of their time in urban trees over the entire tracking period according to the number of fixes; however, each koala spent different amounts of time in these landscapes. One koala, M3, spent 64% of its time in urban areas throughout the breeding season, while another male, M2, spent about half (i.e. 51%) of the time in urban areas throughout the breeding season. The remaining 80% of koala locations were in trees within remnant vegetation, fragmented by roads and the rail line (Figs 1, 2). Some of the remnant vegetation patches, and therefore koala locations, were within the boundaries of properties adjacent to the rail line.
Fate of tracked koalas
Four of the seven radio-collared koalas remained alive after 28 weeks of tracking. Two koalas were killed after the tracking concluded as a result of health checks. The bodies of two koalas (M3 and F2), located close to rail tracks within the rail corridor, were found showing severe trauma consistent with train strike. Two other females (F1 and F4), despite being free of overt signs of disease at the time of radio-collaring, were captured for treatment of chlamydiosis at the end of the project. On veterinary examination they were killed due to untreatable disease. The other koala (M2) death was from a dog attack, confirmed on necropsy by a veterinarian at the Australia Zoo Wildlife Hospital.
Discussion
This study investigated aspects of the ecology of urban koalas using habitat fragmented by linear infrastructure. For some koalas, rail infrastructure appeared to be a barrier to movement, defining the boundaries of their home ranges, while it did not impede the movements of others. Roads appeared to pose no barrier to the movement of koalas, with multiple crossings recorded for animals using habitat adjacent to roads. Due to the small sample size and short monitoring period, generalisations from this study are limited. However, this study has shown that within our study area some koalas moved more freely than others. This could be influenced by a variety of factors, such as the area and quality of vegetation that each individual inhabits, stage of the breeding cycle, age or sex. This may also be an artefact of the short monitoring period. These animals may or may not have been detected crossing road or rail lines if they were monitored over a longer period. Koalas were observed to cross barriers such as 1.8-m-high barbed wire–topped chain-link fences and large tracts of open tree-less landscapes such as rail easements and cleared areas. In this study, males moved greater distances and covered more area than females, particularly at night. While the limited sample size does not allow any definitive sex-based conclusions to be drawn, these results are consistent with other studies that report sex-based differences in koala movement patterns (Phillips 1990; Thompson 2006).
The rail line acted as a home range boundary for some urban koalas, yet was permeable for others. One of the koalas (M1) that survived the duration of the study was the animal that crossed the rail line most often. This might suggest some degree of awareness of rail traffic; however, the limited data and the mortality of other koalas from train strikes on this section of rail line would suggest other reasons for these successful crossings.
Sampling intensity was greatest in the remnant vegetation areas so it is not unexpected that six of the seven animals were caught in these areas and one was caught in a resident’s backyard. Four of six koalas’ home ranges were within the 2–20-ha estimates for south-east Queensland coastal koalas (Hasegawa 1995; Thompson 2006). The other two animals’ home ranges fell outside these estimates: F3 had a home range of ~1.1 ha for MCP95% and M1 had a home range of 31.5 ha for MCP95%. Due to the limited tracking period, we cannot conclude with any certainty that koala F3 had a home range that was outside the south-east Queensland home-range estimates of Hasegawa (1995) and Thompson (2006), had she been monitored for a year or more. However, analysis does suggest that koala M1’s home range was significantly larger than has been recorded for koalas in this region, with further monitoring only enhancing, rather than decreasing, his ranging area. This may be a result of this animal being forced to travel through a fragmented landscape where suitable resources such as food and mating opportunities are dispersed over a greater area (Melzer and Houston 2001).
Home ranges in this study were generally influenced by the shape of the habitat patch and the degree to which linear infrastructure and other urban landscapes created a barrier. The fencing associated with linear infrastructure is assumed to have provided more of a barrier to koalas than roads or the rail line themselves. Unlike other species of animals that rely on closed canopies, understorey or ground cover to move between habitat (e.g. lemuroid ringtail possum, Hemibelideus lemuroides: Wilson et al. 2007; the bush rat and the brown antechinus: Strevens 2007), this study is consistent with other radio-tracking studies that monitored koalas that were able to navigate across roads, cleared areas and other less permeable natural and artificial landscape features (Dique et al. 2003c; Thompson 2006). Again, a longer study with more detailed monitoring of the behaviour of collared koalas may highlight the reason for the perceived barrier that structures such as koala-climbable fencing pose to some animals and not others.
In the Redland City, we found that koalas used trees within remnant vegetation more frequently than urban trees, despite one koala (M2) spending about half his time in urban trees and another koala (M3) spending just over 60% of his time in urban trees. This is most likely a function of the relative availability of different types of trees but it does emphasise the importance of maintaining urban trees and remnant vegetation in urban environments to maintain a koala population. As koalas were using non-regionally endemic and regionally endemic trees within both urban and remnant habitats, every tree in an urban area may be utilised for food or shelter and is potential habitat for koalas. Koalas used trees in backyards, frontyards, along linear infrastructure corridors and in remnant vegetation as a resource or as a link to travel from one vegetation patch to another. This sort of movement is similar to that recorded by White (1999) in rural properties where koalas used isolated trees in paddocks to move from one habitat patch to the next. Also, koalas used trees for shelter, with most trees having branches that crossed over potential fodder or shelter trees, commonly E. tereticornis and M. quinquenervia where they have been found browsing (Martin and Handasyde 1999; Jackson 2007; Woodward et al. 2008). The fact that three of the seven animals were killed while moving on the ground also highlights the importance of the spatial arrangement of suitable habitat to assist in koala movements and help further reduce fatal accidents and dog attacks.
The key to the survival of urban koalas may be their ability to exploit patchy resources and find a mate in a fragmented landscape, but this may also increase the chance that they will suffer trauma from vehicles and dogs (Backhouse and Crouch 1990; Cork et al. 2000; Phillips 2000; Dique et al. 2004). Our findings are consistent with other studies that have shown a high probability of trauma for koalas moving in and through fragmented and heavily urbanised areas in south-east Queensland (e.g. Dique et al. 2003a, 2003c). One of the koalas that was killed by a train (F2) had crossed the rail line by climbing over chain-wire fencing while a safe underpass under the rail line for pedestrians was within metres of the animal’s crossing point. M3 was killed by a train in a separate incident just metres from Ormiston station. Koala mortality records from this area show that over 30 koalas have been killed in the past decade from train collisions (Redland City Council, unpub. data). Two train-related deaths from this small sample of tracked koalas demonstrates that this is a real threat to koalas living in habitat around rail lines. Coupled with the even greater vehicle-related mortality of koalas on the section of arterial road adjacent to the train line, linear-infrastructure-related mortality in this area has serious implications for the viability of an already declining population of koalas.
The importance of revegetation of land within council ownership such as creek corridors, parks and streetscapes within urban areas for future koala conservation has been highlighted in Redland City’s Redland Koala Policy and Implementation Strategy 2008 and there is the potential for more food trees to be planted in urban properties and remnant patches of habitat in order to enhance the connectivity between habitat patches. The monitoring of a larger sample of koalas over a longer period could allow the prioritisation of areas for habitat revegetation based on high-koala-use areas or mortality hot spots, for example. If new tree plantings encourage koalas to move through hostile environments or to undertake additional risky exploratory movements across linear infrastructure to new habitat areas, their value as a conservation tool is questionable.
Acknowledgements
All capture and radio-tracking of these koalas was done under permit from the University of Queensland Animal Ethics Committee (permit # SIS/078/09) and Queensland Environmental Protection Agency Scientific Purposes Permit (WISP05950609). Safe capture of the koalas for collaring and health checks was facilitated by professional tree climbers, Graeme Lloyd and Ben Colquhoun. Robyn Steiner, Tamsin Barnes and Mark Flint provided veterinary assistance and advice, and Sonya Fardell assisted with capture. The Rescue Unit from Australia Zoo and Ecological Services Unit provided tree-climbing assistance to catch two collared koalas. Ron Worth provided ideas to improve the paper and Natalie Waller assisted with home-range analyses. This research was funded by the (then) Redland Shire Council.
References
Allen, C. D., de Villiers, D. L., Manning, B. D., Dique, D. S., Burridge, M., Chafer, M. L., Nicolson, V. N., Jago, S. C., McKinnon, A. J., Booth, R. J., McKee, J. J., Pyne, M. J., Peng Zee, Y., Lundie-Jenkins, G., Theilemann, P., Wilson, R. J., Carrick, F. N., and Johnston, S. D. (2010). Seasonal reproduction in wild and captive male koala (Phascolarctos cinereus) populations in south-east Queensland. Reproduction, Fertility and Development 22, 695–709.| Seasonal reproduction in wild and captive male koala (Phascolarctos cinereus) populations in south-east Queensland.Crossref | GoogleScholarGoogle Scholar |
Asari, Y., Johnson, C. N., Parsons, M., and Larson, J. (2010). Gap-crossing in fragmented habitats by mahogany gliders (Petaurus gracilis). Do they really cross roads and powerlines? Australian Mammalogy 32, 10–15.
| Gap-crossing in fragmented habitats by mahogany gliders (Petaurus gracilis). Do they really cross roads and powerlines?Crossref | GoogleScholarGoogle Scholar |
Backhouse, G., and Crouch, A. (1990). Koala management in the Western Port region Victoria. In ‘Biology of the Koala’. (Eds A. K. Lee, K. A. Handasyde, and G. D. Sanson.) pp. 313–317. (Surrey Beatty: Sydney.)
Brearley, G., Rhodes, J., Bradley, A., Baxter, G., Seabrook, L., Lunney, D., Liu, Y., and McAlpine, C. (2013). Wildlife disease prevalence in human-modified landscapes: a review. Biological Reviews of the Cambridge Philosophical Society 88, 427–442.
| Wildlife disease prevalence in human-modified landscapes: a review.Crossref | GoogleScholarGoogle Scholar | 23279314PubMed |
Canfield, P. J. (1991). A survey of koala road kills in New South Wales. Journal of Wildlife Diseases 27, 657–660.
| 1:STN:280:DyaK38%2FpvFerug%3D%3D&md5=715592dd414c4034c6d237ce79e406f3CAS | 1758032PubMed |
Cogger, H., Ford, H., Johnson, C., Holman, J. and Butler, D. (2003). Impacts of land-clearing on Australian wildlife in Queensland. WWF Australia Report. WWF Australia, Brisbane.
Cork, S. J., Clark, T. W., and Mazur, N. (2000). An interdisciplinary effort for koala conservation. Conservation Biology 14, 606–9.
| An interdisciplinary effort for koala conservation.Crossref | GoogleScholarGoogle Scholar |
Department of Environment and Resource Management (DERM) (2009). Decline of the Koala Coast Koala Population: Population Status in 2008. Available at: http://www.epa.qld.gov.au/publications/p02966aa.pdf/ Decline_of_the_Koala_Coast_Koala_Population_Population_Status_in_2008.pdf [accessed 25 May 2009].
Department of Environment and Resources Management (DERM) (2010). Koala Coast, Koala Population Report 2010. Available at: http://www.derm.qld.gov.au/wildlife-ecosystems/wildlife/koalas/research/pdf/koala-coast-report2010.pdf [accessed 2nd March 2012].
Dique, D., Thompson, J., Preece, H., Penfold, G., de Villiers, D. L., and Leslie, R. S. (2003a). Koala mortality on roads in southeast Queensland: the koala speed zone trial. Wildlife Research 30, 419–426.
| Koala mortality on roads in southeast Queensland: the koala speed zone trial.Crossref | GoogleScholarGoogle Scholar |
Dique, D., Preece, H., and de Villiers, D. (2003b). Koalas in Pine Rivers Shire: distribution, abundance and management. Queensland Parks and Wildlife Service, Brisbane.
Dique, D. S., Thompson, J., Preece, H. J., de Villiers, D. L., and Carrick, F. N. (2003c). Dispersal patterns in a regional koala population in south-east Queensland. Wildlife Research 30, 281–290.
| Dispersal patterns in a regional koala population in south-east Queensland.Crossref | GoogleScholarGoogle Scholar |
Dique, D. S., Preece, H. J., Thompson, J., and de Villiers, D. (2004). Determining the distribution and abundance of a regional koala population in south-east Queensland for conservation management. Wildlife Research 31, 109–117.
| Determining the distribution and abundance of a regional koala population in south-east Queensland for conservation management.Crossref | GoogleScholarGoogle Scholar |
Dowle, M., Webster, K. N., and Deane, E. (2012). Faecal glucocorticoid metabolite concentrations in the free-ranging bandicoots (Perameles nasuta and Isoodon obesulus) of northern Sydney. Australian Mammalogy 35, 1–7.
| Faecal glucocorticoid metabolite concentrations in the free-ranging bandicoots (Perameles nasuta and Isoodon obesulus) of northern Sydney.Crossref | GoogleScholarGoogle Scholar |
Elgar, M. A. (1989). Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence. Biological Reviews of the Cambridge Philosophical Society 64, 13–33.
| Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL1M3kslKltQ%3D%3D&md5=a87bfdeb3e6b84cad6f712b0cdd28989CAS | 2655726PubMed |
Ellis, W. A. H., Melzer, A., Carrick, F. N., and Hasegawa, M. (2002). Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland. Wildlife Research 29, 303–311.
| Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland.Crossref | GoogleScholarGoogle Scholar |
Gavin, S. D., and Komers, P. E. (2006). Do pronghorn (Antilocapra americana) perceive roads as a predation risk? Canadian Journal of Zoology 84, 1775–1780.
| Do pronghorn (Antilocapra americana) perceive roads as a predation risk?Crossref | GoogleScholarGoogle Scholar |
Goosem, M., and Marsh, H. (1997). Fragmentation of a small mammal community by a powerline corridor through tropical rainforest. Wildlife Research 24, 613–629.
| Fragmentation of a small mammal community by a powerline corridor through tropical rainforest.Crossref | GoogleScholarGoogle Scholar |
Gordon, G., Hrdina, F., and Patterson, R. (2006). Decline in the distribution of the koala Phascolarctos cinereus in Queensland. Australian Zoologist 33, 345–358.
Hasegawa, M. (1995). Habitat utilization by koalas (Phascolarctos cinereus) at Point Halloran, Queensland. M.Sc. Thesis, The University of Queensland, Brisbane.
Higgins, D. (2008). Chlamydial Disease in Koalas. In ‘Wildlife Pathology Short Course: Proceedings. Taronga Zoo, Sydney, 21–24 August‘. pp. 247–256. (Australian Registry of Wildlife Health: Sydney.)
Jackson, S. (2007). ‘Koala: Origins of an Icon.’ (Jacana Books and Allen & Unwin: Sydney.)
Kavanagh, R. P., Stanton, M. A., and Brassil, T. E. (2007). Koalas continue to occupy their previous home-ranges after selective logging in Callitris–Eucalyptus forest. Wildlife Research 34, 94–107.
| Koalas continue to occupy their previous home-ranges after selective logging in Callitris–Eucalyptus forest.Crossref | GoogleScholarGoogle Scholar |
Lane, C. (2008). Caboolture, Pine Rivers & Redcliffe Councils Report for Koala Habitat Survey and Mapping: Final Report May 2008. GHD Pty Ltd, Brisbane.
Lee, A., and Martin, R. (1988). ‘The Koala – A Natural History.’ (University of New South Wales Press: Sydney.)
Lee, K. E., Seddon, J. M., Corley, S. W., William, A. H., Ellis, W. A. H., Johnston, S. D., de Villiers, D. L., Preece, H. J., and Carrick, F. (2010). Genetic variation and structuring in the threatened koala populations of southeast Queensland reveals a genetically distinct population in the Koala Coast. Conservation Genetics 11, 2091–2103.
| Genetic variation and structuring in the threatened koala populations of southeast Queensland reveals a genetically distinct population in the Koala Coast.Crossref | GoogleScholarGoogle Scholar |
Lunney, D., Moon, C., and Matthews, A. (1996). A 1990 survey of the koala Phascolarctos cinereus population at Iluka in northern New South Wales. In ‘Koalas – Research for Management – Proceedings of the Brisbane Koala Symposium’. (Ed. G. Gordon.) pp. 102–122. (World Koala Research Incorporated: Brisbane.)
Markey, B., Wan, C., Hanger, J., Phillips, C., and Timms, P. (2007). Use of quantitative real-time PCR to monitor the shedding of chlamydiae in the koala (Phascolarctos cinereus) . Veterinary Microbiology 120, 334–342.
| Use of quantitative real-time PCR to monitor the shedding of chlamydiae in the koala (Phascolarctos cinereus) .Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhs1Sltbg%3D&md5=86c45ad0c5591805d4c7447d4d86a0e8CAS | 17178441PubMed |
Martin, R. and Handasyde, K. (1999). ‘The Koala: Natural History, Conservation and Management.’ (University of New South Wales Press: Sydney.)
Matthews, A., Lunney, D., Gresser, S., and Maitz, W. (2007). Tree use by koalas (Phascolarctos cinereus) after fire. Wildlife Research 34, 84–93.
| Tree use by koalas (Phascolarctos cinereus) after fire.Crossref | GoogleScholarGoogle Scholar |
McAlpine, C. A., Rhodes, J. R., Callaghan, J. G., Bowen, M. E., Lunney, D., Mitchell, D. L., Pullara, D. E., and Possingham, H. P. (2006). The importance of forest area and configuration relative to local habitat factors for conserving forest mammals: a case study of koalas in Queensland, Australia. Biological Conservation 132, 153–165.
| The importance of forest area and configuration relative to local habitat factors for conserving forest mammals: a case study of koalas in Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Melzer, A., and Houston, W. (2001). An overview of the understanding of koala ecology: how much more do we need to know? In ‘The Research and Management of Non-urban Koala Populations’. (Eds K. Lyons, A. Melzer, F. Carrick and D. Lamb.) pp. 6–45. (Koala Research Centre of Central Queensland.)
Melzer, A., Carrick, F., Menkhorst, P., Lunney, D., and St. John, B. (2000). Overview, critical assessment, and conservation implications of koala distribution and abundance. Conservation Biology 14, 619–628.
| Overview, critical assessment, and conservation implications of koala distribution and abundance.Crossref | GoogleScholarGoogle Scholar |
Mitchell, P. (1989). The social organization of the koala. Ph.D. Thesis, Monash University, Melbourne.
Phillips, B. (1990). ‘Koalas: the Little Australians We’d All Hate to Lose.’ (Australian Government Publishing Service: Australia.)
Phillips, S. (2000). Population trends and the koala conservation debate. Conservation Biology 14, 650–659.
| Population trends and the koala conservation debate.Crossref | GoogleScholarGoogle Scholar |
Phillips, S., and Callaghan, J. (2000). Tree species preferences of koalas (Phascolarctos cinereus) in the Campbelltown area south-west of Sydney, New South Wales. Wildlife Research 27, 509–516.
| Tree species preferences of koalas (Phascolarctos cinereus) in the Campbelltown area south-west of Sydney, New South Wales.Crossref | GoogleScholarGoogle Scholar |
Radford, S. L., McKee, R. L., Goldingay, R. L., and Kavanagh, R. P. (2006). The protocols for koala research using radio-collars: a review based on its application in a tall coastal forest in New South Wales and the implications for future research. Australian Mammalogy 28, 187–200.
| The protocols for koala research using radio-collars: a review based on its application in a tall coastal forest in New South Wales and the implications for future research.Crossref | GoogleScholarGoogle Scholar |
Redland City Council (2012). Redlands Koala Policy and Implementation Strategy 2008, Redland City Council. Available at: www.redland.qld.gov.au [accessed 26 April 2012].
Sattler, P., and Williams, R. (1999). The conservation status of Queensland’s bioregional ecosystem. Queensland Environmental Protection Agency, Brisbane.
Smith, P., and Smith, J. (1990). Decline of the urban koala (Phascolarctos cinereus) population in Warringah Shire, Sydney. Australian Zoologist 26, 109–129.
St. Clair, C. C., and Forrest, A. (2009). Impacts of vehicle traffic on the distribution and behaviour of rutting elk, Cervus elaphus. Behaviour 146, 393–413.
| Impacts of vehicle traffic on the distribution and behaviour of rutting elk, Cervus elaphus.Crossref | GoogleScholarGoogle Scholar |
Strevens, T. C. (2007). Powerline easements: ecological impacts and effects on small mammal movement. Ph.D. Thesis, School of Biological Sciences, University of Wollongong.
Stratford, E., Mazur, N., Lunney, D., and Bennett, D. (2000). Managing the koala problem: interdisciplinary perspectives. Conservation Biology 14, 610–618.
| Managing the koala problem: interdisciplinary perspectives.Crossref | GoogleScholarGoogle Scholar |
Sullivan, B. J., Baxter, G. S., Lisle, A. T., Pahl, L., and Norris, W. M. (2004). Low density koala (Phascolarctos cinereus) populations in the mulgalands of south-west Queensland. IV. Abundance and conservation status. Wildlife Research 31, 19–29.
| Low density koala (Phascolarctos cinereus) populations in the mulgalands of south-west Queensland. IV. Abundance and conservation status.Crossref | GoogleScholarGoogle Scholar |
Tarlinton, R., Meers, J., Hanger, J., and Young, P. (2005). Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral and neoplastic disease in koalas. The Journal of General Virology 86, 783–787.
| Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral and neoplastic disease in koalas.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXit12lt7k%3D&md5=43f5c8d6789a920bc571bc157905f73cCAS | 15722540PubMed |
Thompson, J. (2001). The role of research in koala management: case studies from south-east Queensland. In ‘The Research and Management of Non-urban Koala Populations’. (Eds K. Lyons, A. Melzer, F. Carrick, and D. Lamb.) pp. 89–104. (Koala Research Centre of Central Queensland, Central Queensland University: Rockhampton.)
Thompson, J. (2006). The comparative ecology and population dynamics of koalas in the Koala Coast region of south-east Queensland. Ph.D. Thesis, The University of Queensland, Brisbane.
Timms, P. (2005). Chlamydial infection and disease in the koala. Microbiology Australia 26, 65–68.
Translink (2009). Cleveland Line Timetable. Available at: http://www.download.translink.com.au/timetables/090223_cleveland.pdf [accessed 13 November 2009].
Weigler, B. J., Girjes, A. A., White, N. A., Kunst, N. D., Carrick, F. N., and Lavin, M. F. (1988). Aspects of the epidemiology of Chlamydia psittaci infection in a population of koalas (Phascolarctos cinereus) in southeastern Queensland, Australia. Journal of Wildlife Diseases 24, 282–291.
| 1:STN:280:DyaL1c3jtVOitw%3D%3D&md5=4951815d4fb13a8e6a028ec3c08c1b7dCAS | 3373633PubMed |
White, N. A. (1999). Ecology of the koala (Phascolarctos cinereus) in rural south-east Queensland, Australia. Wildlife Research 26, 731–744.
| Ecology of the koala (Phascolarctos cinereus) in rural south-east Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Wilson, R. F., Marsh, H., and Winter, J. (2007). Importance of canopy connectivity for home range and movements of the rainforest arboreal ringtail possum (Hemibelideus lemuroides). Wildlife Research 34, 177–184.
| Importance of canopy connectivity for home range and movements of the rainforest arboreal ringtail possum (Hemibelideus lemuroides).Crossref | GoogleScholarGoogle Scholar |
Woodward, W., Ellis, W. A., Carrick, F. N., Tanizaki, M., Bowen, M., and Smith, P. (2008). Koalas on North Stradbroke Island: diet, tree use and reconstructed landscapes. Wildlife Research 35, 606–611.