

Long-term impact of severe wildfire and post-wildfire salvage logging on macroinvertebrate assemblage structure in Alberta’s Rocky Mountains
Amanda M. Martens A F , Uldis Silins A , Heather C. Proctor B , Chris H. S. Williams A , Michael J. Wagner C , Monica B. Emelko D and Micheal Stone E
A F , Uldis Silins A , Heather C. Proctor B , Chris H. S. Williams A , Michael J. Wagner C , Monica B. Emelko D and Micheal Stone E
A Department of Renewable Resources, University of Alberta, Edmonton, AB T6G 2H1, Canada.
B Department of Biological Sciences, University of Alberta, Edmonton, AB T6G 2R3, Canada.
C Alberta Agriculture and Forestry, Calgary, AB T3L 3M4, Canada.
D Department of Civil and Environmental Engineering, University of Waterloo, Waterloo, ON N2L 3G1, Canada.
E Department of Geography and Environmental Management, University of Waterloo, Waterloo ON, N2L 3G1, Canada.
F Corresponding author. Email: amartens@ualberta.ca
International Journal of Wildland Fire 28(10) 738-749 https://doi.org/10.1071/WF18177
Submitted: 4 October 2018 Accepted: 23 June 2019 Published: 19 September 2019
Journal Compilation © IAWF 2019 Open Access CC BY-NC-ND
Abstract
Wildfire is an important natural disturbance on forested landscapes influencing both physical and biological processes. The Lost Creek wildfire was one of the most severe on Alberta’s eastern slopes and provided a unique opportunity to assess the long-term impacts of wildfire and post-wildfire salvage logging on northern Rocky Mountain catchments. Macroinvertebrate sampling conducted 8 years after the wildfire indicated distinct macroinvertebrate assemblages in reference, burned, and burned and salvage-logged catchments. Reference catchments were characterised by more sensitive taxa (e.g. stoneflies) and had the lowest abundance of macroinvertebrates. Burned catchments had the greatest abundance of macroinvertebrates and were characterised by greater abundance of chironomids and caddisflies. Burned and salvage-logged catchments supported high numbers of riffle beetles and crane flies. The unique assemblage structures between catchments indicate different mechanisms drive the ecological response in wildfire-affected catchments. Resource availability was an important driver of assemblage structure in the more productive burned catchments and habitat quality was a dominant driver in the burned and salvage-logged catchments. Despite the legacy effects observed in this study, fire-affected catchments still supported sensitive taxa and functional macroinvertebrate assemblages.
Additional keywords: natural disturbance, stream ecology.
Introduction
Wildfire is an important natural disturbance that can strongly influence both physical and biological environmental processes (Gresswell 1999; Moody and Martin 2001; Hauer et al. 2007). Since the 1980s, increases in wildfire frequency, severity, duration and season length in the western United States have been correlated with warming atmospheric temperatures (Schoennagel et al. 2004; Westerling et al. 2006). Between 1970 and 2003, 60% of the observed increases in wildfire activity occurred in the Rocky Mountains (Westerling et al. 2006). These ecosystems will be particularly susceptible to climate change-induced shifts in wildfire behaviour and it is expected that by the end of the 21st century, the annual area burned in Canada will increase by 74 to 118% compared with the latter half of the 20th century (1959–97) (Flannigan et al. 2005). Such increases in area burned will have strong effects on both terrestrial and freshwater ecosystems (Romme et al. 2011). Post-fire landscape management such as salvage logging can also influence or compound the effects of fire on aquatic ecosystems by increasing ground disturbance and adding linear features (e.g. skid trails, haul roads and stream crossings) (Beschta et al. 1995; Wagenbrenner et al. 2016).
Forested mountain catchments provide critical habitat for many species, including macroinvertebrates and salmonid fishes (Hauer et al. 2007; Isaak et al. 2016). These streams typically have higher water quality than lowland streams (Hauer et al. 2007), and are sensitive to the changes in vegetation and soils caused by wildfire (Hynes 1975; Hauer et al. 2016). Wildfire alters rainfall-runoff dynamics, which can change sediment and nutrient regimes, channel morphology and water temperature (Gresswell 1999). Elevated sediment loading, export of nutrients (nitrogen, phosphorus and dissolved organic carbon), export of heavy metals (e.g. mercury and methyl mercury) and stream temperature are commonly reported after wildfire in mountain catchments (Hauer and Spencer 1998; Kelly et al. 2006; Shakesby and Doerr 2006; Writer et al. 2012; Wagner et al. 2014). Increased nutrients stimulate microbial activity and autochthonous (in-stream) primary productivity (Spencer et al. 2003; Mihuc and Minshall 2005) while the influx of fine sediment can affect the availability and quality of in-stream habitat and food resources (Bjornn et al. 1977; Jones et al. 2012). Macroinvertebrate assemblages invariably respond to the water-quality changes associated with wildfire (Barbour et al. 1999; Minshall 2003). Assemblage composition often shifts to favour disturbance-adapted taxa, while more sensitive taxa can decline in abundance or be excluded entirely (Mihuc et al. 1996; Minshall et al. 2001a, 2001b; Malison and Baxter 2010b). Changes in biomass and density are less predictable but are often related to factors such as primary productivity, water chemistry and the occurrence of drought or flooding in the months or years following wildfire (Verkaik et al. 2015). Rinne (1996) documented large declines (70–90%) in invertebrate density following high-flow events with heavy sediment loads after a wildfire in Arizona. In contrast, Malison and Baxter (2010b) reported greater invertebrate productivity in severely burned reaches following the Diamond Peak wildfire in Idaho and associated this with increases in food resources. The extent of wildfire effects on macroinvertebrate assemblages depends on fire severity, local catchment characteristics including the hydroclimatic setting, vegetation type, topography, soils and geology, as well as the influences of climate change, including warming and changes in precipitation patterns (Moody and Martin 2001; Shakesby and Doerr 2006; Rugenski and Minshall 2014).
Macroinvertebrates are an important component of Rocky Mountain stream ecosystems. They contribute to nutrient cycling and provide a critical food source for fish and riparian insectivores (Mihuc et al. 1996; Rinne 1996; Minshall 2003; Malison and Baxter 2010a). Despite their importance in the trophic structure of stream ecosystems, very little is known about the long-term effects of wildfire on macroinvertebrate assemblages. Previous post-fire invertebrate studies report on shorter-term (1–5-year) impacts (Rinne 1996; Minshall et al. 1997; Spencer et al. 2003; Malison and Baxter 2010a; Malison and Baxter 2010b). However, fire can produce longer-term changes in the physical stream environment and water-quality conditions regulating stream ecology (Silins et al. 2014; Emelko et al. 2016). Although several studies in the United States have reported wildfire impacts on macroinvertebrate assemblages lasting as long as 5–10 years (Minshall et al. 1989; Roby 1989; Mihuc et al. 1996; Vieira et al. 2004; Malison and Baxter 2010b), both the longevity of fire effects on macroinvertebrates and factors regulating longer-term patterns of ecosystem recovery are presently not well understood across more northern mountain environments (Bixby et al. 2015).
The Lost Creek wildfire was one of the more severe fires in recent decades on the eastern slopes of Alberta’s Rocky Mountains and provided an opportunity to assess the short-, mid- and long-term effects of wildfire on northern Rocky Mountain catchments. The Southern Rockies Watershed Project (SRWP) was established to document the effects of the wildfire and post-wildfire salvage logging on hydrology, biochemistry and aquatic ecology (Silins et al. 2016). As reported elsewhere, 11 years of continuous monitoring have demonstrated that wildfire effects on water quality and biogeochemistry are persistent in the study catchments, with many parameters including sediment production, nutrient (phosphorus and dissolved organic carbon) export and primary productivity showing no sign of return to reference conditions over a decade post wildfire (Silins et al. 2009; Emelko et al. 2011; Silins et al. 2014; Wagner et al. 2014; Emelko et al. 2016; Silins et al. 2016). Given these persistent impacts on hydrology and water quality, disturbance-associated effects on macroinvertebrate abundance and assemblage structure between reference and wildfire-affected catchments also were expected. We expected greater abundance in the more productive wildfire-affected catchments, and greater diversity in the unburned systems. Thus, the objectives of the present study were to assess the long-term (8 years post fire) impacts of the Lost Creek wildfire on macroinvertebrates by:
-
quantifying differences in macroinvertebrate abundance and assemblage structure in streams draining three types of catchments: reference (unburned), burned, and burned and salvage-logged,
-
determining the environmental parameters (water quality, streamflow and periphyton productivity) that best explain variation in macroinvertebrate assemblage structure, and
-
analysing a selection of ecological traits of more abundant taxa along a disturbance gradient (reference, burned, and burned and salvage-logged) to infer mechanisms that influence the response of macroinvertebrate assemblages after wildfire.
Methods
Site description
The Lost Creek wildfire burned 211.6 km2 in the forested region of the Crowsnest and Castle River catchments in southern Alberta, Canada, from July to September 2003. The high-severity fire effectively consumed all of the forest cover (closed canopy, mature or old growth), understorey vegetation and forest floor organic material. The upper portions of the study catchments that did not burn (particularly South York and Lynx Creeks) were treeless alpines zones will little available fuel (Fig. 1, Table 1). Seven research catchments were established, including three burned catchments (South York, Lynx and Drum Creeks) and two reference (unburned) catchments (Star and North York Creeks) (Fig. 1, Table 1). Two catchments (Lyons East and Lyons West Creeks) were salvage-logged and instrumented in 2005. Clear-cut salvage harvest occurred over the winters of 2004 and 2005; 2.6 and 2.4 km2 (19.9 and 33.6% of the catchment area) were harvested in Lyons East and Lyons West respectively (Fig. 1, Table 1). Forest regeneration was slow in wildfire-affected catchments; canopy cover did not recover in the 8 years before sampling and no subsequent fires occurred in research catchments during the study period.

|

|
The hydrologic regime of these catchments is snowmelt-dominated. The highest flows occur in late May to early June during the peak of spring snowmelt. Mean annual precipitation in the reference, burned, and burned and salvage-logged catchments was 1087 (898–1398), 1146 (950–1431) and 775 mm (582–971 mm) respectively (2004–2014), most of which fell as snow or as rain during summer storm events. Mean annual area-weighted discharge in the reference, burned, and burned and salvage-logged catchments over the same period was 744 (483–1080), 871 (571–1091) and 592 mm (373–887 mm) respectively. Forest cover before the fire was similar between catchments and consisted predominantly of lodgepole pine (Pinus contorta var. latifolia) at lower elevations (upper Montane ecozones), Englemann spruce (Picea engelmanii) and subalpine fir (Abies lasiocarpa) at mid elevations (subalpine ecozones), and treeless alpine meadow vegetation and bare rock at higher elevations, characteristic of the upper alpine ecozones. Although no predisturbance hydrologic, water quality or aquatic ecological data existed from the study area (as is typical for most wildfire studies), modest replication (2–3) of undisturbed (reference) and disturbed (burned, and burned and salvage-logged) catchments with similar physical and environmental characteristics have been previously used to support broader inferences on wildfire and salvage-logging impacts (Spencer et al. 2003; Reid et al. 2010). This assumes that disturbed catchments would behave similarly to reference catchments had the disturbance not occurred.
Environmental parameters
All parameters used in this analysis were calculated from data collected 8 years after the fire, during the ice-free period from April to October 2011 (Table 2). Water samples were collected every 2 weeks and analysed according to standard methods (Rice 2012; US Environmental Protection Agency (EPA) 2012) for ammonium (NH4+), nitrate + nitrite (NO3− + NO2−), soluble reactive phosphorous (SRP), dissolved organic carbon (DOC), dissolved inorganic carbon (DIC), alkalinity (as CaCO3), total suspended solids (TSS) and total dissolved solids (TDS). Total inorganic nitrogen (TIN) was determined by the sum of ammonium, nitrate and nitrite.

|
Stream discharge (m3 s−1) was measured every 2 weeks using a Sontek acoustic Doppler velocity meter (Flow Tracker ADV, Sontek/YSI). Stage was recorded every 10 min using HOBO water level loggers suspended in PVC stilling wells (U20–001–04; Onset Computer Corporation) or with a dry gas bubbler and pressure transducer (Waterlog Model H-350 Lite and H-355, Design Analysis Associates Inc.) connected to a measurement and control data logger (CR1000, Campbell Scientific). Discharge and manual stage measurements were used to develop rating curves for each stream and calculate mean daily discharge. Stream temperature was recorded either every 60 min using HOBO temperature data loggers or every 10 min using HOBO temperature and water level loggers (H08–001–02 or U20–001–04; Onset Computer Corporation). Mean daily stream temperature (AVDT) was calculated and used for this analysis.
Periphyton samples were collected using three replicate unglazed ceramic tiles (155 cm2) anchored to the streambed in riffle sections of all seven catchments in late April–early May after ice-out. Tiles were scrubbed clean in running stream water before deployment and periphyton was allowed to colonise naturally. Samples were collected monthly for 5 months (June to October) by scraping and rinsing periphyton from the tile surface into acid-washed (10% HCl) Nalgene high-density polyethylene (HDPE) sample bottles. Ash-free dry mass (AFDM) was used as a surrogate for primary productivity (Aloi 1990); samples were frozen before analysis.
Macroinvertebrate sampling
Macroinvertebrate sampling was conducted at the end of the growing season in October 2011. A Surber sampler (500-μm mesh, 0.096 m2) was used to collect invertebrates from eight randomly chosen riffle sections along a 100-m transect upstream of each hydrometric gauging station. The sampler was placed on the streambed facing upstream. Large rocks within the sampling area were rubbed clean and removed, then the bed material was vigorously disturbed for 2 min. Sampler contents were placed in a white plastic basin and large debris (e.g. whole leaves, twigs and rocks) were rinsed and removed. Macroinvertebrates, associated plant material and detritus were transferred to Nalgene HDPE sample bottles and preserved in 95% ethanol. The 500-μm mesh of the Surber sampler permitted the collection of larger-bodied water mites and later insect instars. Macroinvertebrates were sorted, enumerated and identified to the lowest practical taxonomic unit (usually family) using a dissecting microscope and the following keys: Clifford (1991) and Thorp and Covich (2009). Identification of freshwater macroinvertebrates to the level of family has been shown to be sufficient for multivariate analyses comparing assemblage structure between disturbed and undisturbed systems (Bowman and Bailey 1997; Bailey et al. 2001).
Roundworms (Nematoda) and the water mite family Feltriidae were excluded from the analysis as the large mesh size (500 μm) prevented consistent collection of a representative sample. Specimens that could not be identified to class were also excluded. Individuals from the stonefly families Capniidae and Leuctriidae were too young to be distinguished from each other and were grouped for this analysis as Capniidae + Leuctridae. Excluded specimens represented 2.61% of the total specimens collected.
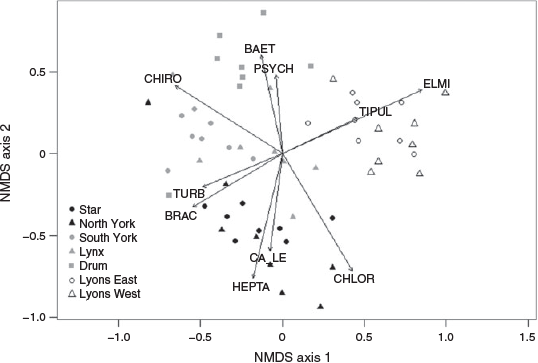
Parameters describing community composition (taxonomic richness, diversity and evenness) were calculated using formulae from Morris et al. (2014). Taxonomic richness (S) is the total number of taxa observed in each catchment (Eqn 1). Diversity (D) was calculated using Simpsons Index of Diversity where pi is the proportion of individuals of taxon i (Eqn 2). Evenness (E) was calculated using Simpson’s Evenness (Eqn 3):
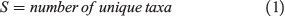


Statistical analysis
Statistical analyses were conducted using R (R Core Team 2018). Community ordination and analysis were conducted using the vegan package (Oksanen et al. 2019). One-way analysis of variance (ANOVA) was used to test for an effect of disturbance category on univariate environmental predictors (streamflow, temperature, water chemistry and primary productivity). Parameters that did not meet the assumptions of parametric tests (normal distribution and equal variance) were either log-transformed or analysed using a non-parametric Kruskal–Wallis rank sum test. Post-hoc comparisons between treatments were conducted following one-way ANOVA using Tukey’s honest significant difference (Tukey’s HSD) test or following Kruskal–Wallis rank sum test using Fisher’s least significant difference test. Permutational multivariate analysis of group dispersions (PERMDISP2) was used to test for multivariate homogeneity of group dispersions. Tukey’s HSD test was used to conduct post-hoc pairwise comparisons of dispersion between treatments. A multivariate analysis of variance (mANOVA) was conducted on the Bray–Curtis dissimilarity matrix to test for significant differences in taxonomic composition between catchments and treatments.
Non-metric multidimensional scaling (NMDS) using Bray–Curtis dissimilarity matrices was used to visualise the variability of invertebrate taxa in multidimensional space and evaluate the similarity of invertebrate assemblages among catchments within disturbance categories. NMDS ordinations were conducted using both raw counts and log (x + 1) transformed data. As invertebrate abundance was considerably higher in the burned catchments, ordinations using counts were heavily influenced by burned catchments. To reduce this effect, ordinations were also conducted using relative abundances instead of counts to compute the dissimilarity matrix.
Redundancy analysis (RDA) was used to determine the strongest environmental correlates of macroinvertebrate assemblage structure among disturbance categories. Environmental variables used in the RDA are summarised in Table 2. Detrended correspondence analysis (DCA) indicated RDA was the appropriate model, as axis lengths were less than 3 standard deviations (between 1.75 and 2.34) (Borcard et al. 2011). Variables with high multicollinearity (variance inflation factor >10) were removed before the selection of the most important environmental parameters using a forward stepwise selection procedure (ordiR2step). A Hellinger transformation was applied to the species counts and environmental parameters were standardised to mean zero and unit variance before fitting the RDA model (Legendre and Gallagher 2001).
Results
Environmental parameters
Stream discharge (Q) in 2011 was similar between reference, burned, and burned and salvage-logged catchments (Qdaily = 0.36 ± 0.02, 0.30 ± 0.02 and 0.35 ± 0.03 m3 s−1 respectively; χ2 = 7.8498, P = 0.0197). The annual flow regime (range of flows) varied more between treatments than did mean discharge (Fig. 2). Reference catchments were the least variable, with the least pronounced snowmelt and stormflow peaks and the lowest peak discharge (2.03 m3 s−1). The burned catchments were intermediate, with peak discharge of 2.11 m3 s−1. Burned and salvage-logged catchments were the most variable, with steeper snowmelt and stormflow limbs and the highest peak discharge (5.18 m3 s−1). Mean daily stream temperature was significantly different between treatments (χ2 = 38.492, P < 0.001). Stream temperature was highest and most variable in the burned and salvage-logged catchments (6.51 ± 0.24°C compared with 4.64 ± 0.11 and 3.58 ± 0.11°C in the reference and burned catchments respectively).

|
Periphyton productivity (AFDM) varied throughout the growing season in all catchments; mean seasonal AFDM was lowest in the reference catchments and greatest in the burned and burned and salvage-logged catchments (0.09 ± 0.03, 0.21 ± 0.05 and 0.28 ± 0.07 mg cm−2 in the reference, burned, and burned and salvage-logged catchments respectively; F1,61 = 7.504, P < 0.01). AFDM was significantly lower in the reference catchments compared with the burned and burned and salvage-logged catchments.
Water chemistry values are summarised in Table 2. TIN, TDS, DIC and alkalinity were greatest in the reference and lowest in the burned and salvage-logged catchments, showing a generally monotonic pattern across disturbance categories. In contrast, SRP, TSS and DOC were greatest in the burned and salvage-logged catchments, following a pattern of burned and salvage-logged > burned > reference.
Macroinvertebrate assemblage structure
A total of 21 409 individuals from 29 taxonomic groups were identified, enumerated and included in this analysis (Table 3). Insects (Ephemeroptera, Plecoptera, Trichoptera, Diptera and Coleoptera) made up the majority of the macroinvertebrate assemblages (94.0, 94.3 and 97.7% in the reference, burned, and burned and salvage-logged catchments respectively). Flatworms (Platyhelminthes: Turbellaria), aquatic mites (Acariformes: Prostigmata: Parasitengonina: Hydrachnidiae) and ostracods (Crustacea) comprised the remainder of the assemblages.

|
Macroinvertebrates were most abundant in burned streams, with a mean density of 5959.6 ± 577.1 individuals m−2 (Fig. 3). Abundance was similar between reference and burned and salvage-logged streams with densities of 2529.5 ± 405.5 and 2731.1 ± 421.5 individuals collected per sample respectively. Taxonomic richness (S) was similar across catchments (S = 25.0 ± 3.0, 26 ± 0.0 and 23.5 ± 0.5 in the reference, burned, and burned and salvage-logged catchments respectively). All taxa sampled were represented at least once in each of the reference, burned, and burned and salvage-logged catchments with the exception of two aquatic mite families. The family Hygrobatidae was absent from the burned and salvage-logged catchments and the family Torrenticolidae was absent from the reference and burned catchments; both families were relatively rare across catchments, accounting for 0.03 and 0.02% of the total individuals sampled respectively. Diversity (D) and evenness (E) were moderately higher in reference catchments and similar across the wildfire-affected catchments (D = 0.85 ± 0.01, 0.80 ± 0.05 and 0.79 ± 0.01 and E = 0.28 ± 0.06, 0.22 ± 0.05 and 0.21 ± 0.01 in the reference, burned, and burned and salvage-logged catchments respectively). Counts and relative abundances of all taxonomic groups are summarised by disturbance category in Table 3.

|
Reference catchments were characterised by high relative abundances of stoneflies (Plecoptera) (41.9% compared with 13.1 and 24.2% in the burned and burned and salvage-logged catchments respectively). In particular, the relative abundance of Capniidae + Leuctridae was high (4.15% compared with 0.72 and 0.47% in the burned and burned and salvage-logged catchments). Burned catchments showed high relative abundances of both true flies (Diptera) and mayflies (Ephemeroptera) (22.0 and 45.1% compared with 11.8 and 31.5%, and 11.1 and 29.44% in the reference and burned and salvage-logged catchments respectively). In particular, the true fly families Chironomidae and Psychodidae and the mayfly family Baetidae were abundant in burned catchments. The burned and salvage-logged catchments were characterised by particularly high abundances of riffle beetles (Coleoptera: Elmidae) (28.63% compared with 0.76 and 4.01% in the reference and burned catchments respectively) as well as crane flies (Diptera: Tipulidae) (1.59% compared with 0.13 and 0.30% in the reference and burned catchments respectively). The burned and salvage-logged catchments had particularly low numbers of flatworms; only two individuals were collected, representing 0.05% of the macroinvertebrate assemblage compared with 1.84 and 3.56% in the reference and burned catchments respectively.
Multivariate analysis of macroinvertebrate assemblage structure
Invertebrate assemblages formed distinct groups according to catchment and disturbance category in NMDS ordinations (Fig. 4). Multivariate ANOVA of the Bray–Curtis dissimilarity matrices confirmed significant differences in assemblage structure between catchments (F = 7.845, P < 0.01) and treatments (F = 11.403, P < 0.01). The test of homogeneity of multivariate dispersions (PERMDIPS2) indicated that β diversity was homogeneous between disturbance category (number of permutations = 999, F = 2.821, P = 0.07) as confirmed with Tukey’s HSD test (P > 0.12).
|
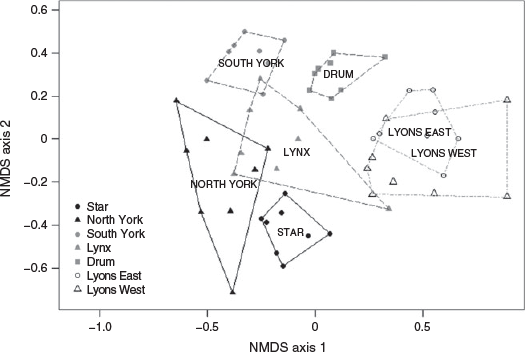
Several taxonomic groups were strongly associated (P < 0.01) with study catchments across disturbance categories (Fig. 5). Heptageniidae and Capniidae + Leuctridae were strongly associated with reference catchments. Burned catchments were positively associated with the families Baetidae, Psychodidae and Chironomidae whereas Tipulidae and Elmidae were strongly associated with burned and salvage-logged sites. Several taxa were also associated with catchments in two disturbance categories including Peltoperlidae, Brachycentridae and Turbellaria associated with both reference and burned catchments and Chloroperlidae, which was characteristic of both reference and burned and salvage-logged catchments.
|
The RDA model explained 34.4% of the variance in invertebrate assemblage structure (P < 0.01, Fig. 6). The first two RDA axes explained 20.3 and 7.8% of the variation respectively. The variables selected by the model in decreasing order of strength were: DOC, stream discharge (Q), periphyton productivity (AFDM) and TDS. Reference reaches were characterised by higher discharge and lower periphyton productivity. The burned catchments were highly variable; Drum Creek had particularly high TDS, South York Creek was characterised by low discharge and DOC, and Lynx Creek was more similar to the reference sites, having high discharge and low DOC. Burned and salvage-logged sites were characterised by high DOC and elevated periphyton productivity. Environmental predictors are summarised in Table 2.

|
Discussion
Persistent wildfire effects on macroinvertebrate assemblages were observed in both burned and burned and salvage-logged catchments 8 years after the Lost Creek wildfire (Fig. 4). This observation is consistent with the slow post-wildfire recovery of biogeochemical and biological factors (including increased sediment, phosphorus and carbon export and increased periphyton productivity) observed in these wildfire-affected watersheds (Bladon et al. 2008; Silins et al. 2009; Emelko et al. 2011; Silins et al. 2014; Wagner et al. 2014; Emelko et al. 2016; Silins et al. 2016). Previous reports on the duration of wildfire impacts to water quality and stream biology have focused on shorter timescales (2–3 years) than those described here (Moody and Martin 2001; Robson et al. 2018). Results of the present study demonstrate that the effects of both wildfire and post-fire salvage logging on macroinvertebrate assemblages (the latter effects have not been previously described) can be long-lasting (Fig. 6) and these effects are strongly associated with the persistence of fire effects on the chemical and biological stream environments. In particular, DOC and periphyton productivity (AFDM) were closely associated with variation in invertebrate assemblage structure among disturbance categories and continue to be influenced by wildfire-related changes to runoff and water quality (Emelko et al. 2016; Silins et al. 2016). It is important to recognise that multiple stream water quality and physical variables after wildfire were strongly correlated, thus although redundancy analysis identified DOC and AFDM as the dominant fire-associated variables, sediment, phosphorus, carbon and periphyton productivity are all likely key drivers regulating the persistence of wildfire effects. The glacial history of these research catchments contributes to long-lasting wildfire effects compared with mountain ecosystems in the western United States (e.g. Yellowstone; Minshall et al.1997) owing to the presence of highly erodible glacial-fluvial deposits that provide a source of bioavailable phosphorus (Emelko et al. 2016).
The differences in ecological response (Fig. 5) of the macroinvertebrate assemblages in the burned and burned and salvage-logged catchments were unexpected (Table 2). Sediment export, phosphorus and DOC concentrations (biogeochemical) and periphyton productivity (biological) consistently follow the pattern burned and salvage-logged > burned > reference. Accordingly, it would be expected that the response of macroinvertebrate assemblages in burned and burned and salvage-logged catchments would follow the same pattern. Notably, assemblage structure differed between burned and burned and salvage-logged watersheds (Figs 4, 5) and macroinvertebrate abundance was 2× greater in burned catchments whereas abundance in burned and salvage-logged catchments did not differ significantly from reference assemblages (Fig. 3). Chironomidae and Baetidae were predominant in burned catchments whereas Tipulidae and Elmidae were indicative of burned and salvage-logged systems (Fig. 5; Table 3). These unique assemblage structures and macroinvertebrate abundances between disturbance categories suggest that different mechanisms are driving the ecological responses in wildfire-affected catchments.
Wildfire can affect macroinvertebrate assemblages through several biophysical mechanisms; in this study, we consider the influence of resource limitation and habitat quality. Macroinvertebrate responses to wildfire can also be influenced by other factors including life stage and physiology, which were not accounted for in this study. The resource-limitation mechanism occurs when macroinvertebrates respond to changes in the availability (quantity and quality) of both allochthonous (terrestrial) and autochthonous (in-stream) food resources (Minshall et al. 1989; Spencer et al. 2003; Mihuc and Minshall 2005). Food webs in undisturbed forested headwater streams will be supported primarily by allochthonous sources of energy as low nutrient concentrations and shading from the riparian canopy prevent significant in-stream primary productivity (Vannote et al. 1980). Unburned research catchments consistently fit this pattern, having low primary productivity compared with nearby disturbed, both burned (Emelko et al. 2016) and harvested (Hawthorn 2014) catchments. The habitat quality mechanism is related to physical habitat characteristics, where increases in runoff, erosion, sediment and nutrient loading alter both habitat availability and suitability (Hauer and Spencer 1998; Shakesby and Doerr 2006). Considering the reference and burned disturbance categories, both periphyton productivity and macroinvertebrate abundance were greater in the burned catchments, suggesting that resource availability was likely the primary mechanism regulating macroinvertebrate assemblage response. However, the burned and salvage-logged catchments (influenced by two consecutive disturbances) consistently showed disturbance metrics (suspended sediment, DOC, phosphorus and periphyton productivity) higher than catchments affected by the wildfire only (Table 2). The unique invertebrate assemblage structure in the burned and salvage-logged catchments, and the pattern of high primary productivity but low macroinvertebrate abundance suggest that habitat quality had a greater influence on the ecological response in these systems. The increased sediment loading and more variable flow regime in the burned and salvage-logged catchments likely reduced habitat quality to the point that few taxa were able to take advantage of the increased productivity. These observations are consistent with a previous study that reported differential effects of wildfire on invertebrate assemblages in low- and high-severity burns (Malison and Baxter 2010b); these differences were not interpreted in the context of broader ecological limitations proposed here.
The macroinvertebrate assemblage of the reference catchments was consistent with streams with high water quality and a more stable flow regime. Taxa known to be sensitive to environmental perturbation including stoneflies (Plecoptera) and flatheaded mayflies (Ephemeroptera: Heptageniidae) (Barbour et al. 1999; Voshell 2002) were relatively abundant in the reference catchments (Fig. 5, Table 3). Stoneflies are particularly sensitive to sedimentation and excess periphyton (Zwick 1992; Barbour et al. 1999; Voshell 2002). Diversity and evenness were moderately higher in reference catchments, likely owing to greater stability of the flow regime and high-quality habitat compared with burned and burned and salvage-logged catchments.
Macroinvertebrate assemblages in the burned catchments included greater abundance of disturbance-adapted taxa as well as taxa adapted to capitalise on increased food availability. Invertebrate families in burned catchments included the true flies Chironomidae and Psychodidae and the mayfly family Baetidae (Fig. 5). Many species within Chironomidae and Baetidae are widely considered to be disturbance-adapted taxa and increase in abundance post fire as they have short generation times and can reproduce quickly (Richards and Minshall 1992; Malison and Baxter 2010b). The family Baetidae contains several genera specialised for consuming periphyton (Minshall et al. 2001b); their high abundance suggests that individuals were able to capitalise on increased periphyton productivity in the burned catchments. The abundance of a given taxon will be greatest in the area that provides the most suitable habitat (Brittain and Eikeland 1988); the wildfire alone did not compromise habitat quality to the point that invertebrate taxa were unable to take advantage of the excess periphyton resources.
In contrast, salvage logging contributed to greater deterioration in water quality and stream habitat compared with catchments affected by the fire alone, preventing invertebrates from utilising the additional periphyton productivity. Streamflow was very responsive to rainfall and snowmelt events, resulting in higher peak streamflows than in reference catchments. Suspended sediment and turbidity were consistently higher during these periods of high flows and considerable amounts of fine sediment were deposited on and intruded into gravel interstices in the streambed. The reduced invertebrate abundance observed in these catchments is consistent with the observations of Wood and Armitage (1997) and Bjornn et al. (1977), who showed that increased fine sediment reduced invertebrate density and abundance. Although sediment is an important natural component of stream ecosystems, excess fine sediment can fill pore spaces and reduce oxygen availability in gravel bed streams, compromising habitat and food resources and altering macroinvertebrate behaviour including foraging, respiration and drift (Bjornn et al. 1977; Brittain and Eikeland 1988; Wood and Armitage 1997; Jones et al. 2012). Crane flies (Diptera: Tipulidae) and riffle beetles (Coleoptera: Elmidae) were strongly associated with burned and salvage-logged catchments (Fig. 5). Crane flies have a burrowing lifestyle and are well adapted to high-sediment environments (Giller and Malmqvist 1998; Voshell 2002). As well, riffle beetle larvae can grip substrate during high flows and protect themselves from sediment abrasion by withdrawing their gills (Bjornn et al. 1977; Brown 1987; Voshell 2002; Yee and Kehl 2015). Fine sediment deposition likely contributed to the low abundance of caddisflies (Trichoptera) as some taxa require clean substrate to attach their cases and retreats (Bjornn et al. 1977; Voshell 2002). The low abundance of baetid mayflies in the burned and salvage-logged catchments is further evidence of the habitat-limitation mechanism overriding the resource mechanism. Many genera of Baetidae are disturbance-adapted scrapers (consume periphyton) and do well in post-fire environments with high periphyton productivity (Vieira et al. 2004; Malison and Baxter 2010b). High suspended sediment loads, however, can be detrimental as many genera are also clingers (Bjornn et al. 1977); impact from moving sediment dislodges individuals from rocks, causing injury and reducing abundance (Naman et al. 2016).
Conclusion
The clear differences in macroinvertebrate assemblage structure between reference, burned, and burned and salvage-logged catchments 8 years after the Lost Creek wildfire indicate that the effects of wildfire on aquatic communities in this northern Rocky Mountain ecosystem are persistent. However, despite clear differences in the relative abundance of taxa between disturbance categories, taxonomic richness and diversity did not vary strongly. Only 2 of 29 taxa were missing from one or more disturbance categories; both these families (Hygrobatidae and Torrenticolidae) were locally rare across disturbance categories. It is clear that despite the legacy effects observed in this study, fire-affected catchments still support sensitive taxa and functional macroinvertebrate assemblages 8 years after severe wildfire. The cumulative impacts of wildfire and forest harvest should be considered when making post-wildfire management decisions related to salvage logging. Macroinvertebrates provide many important ecosystems services including sediment mixing, breakdown of organic matter, and nutrient and energy cycling (Covich et al. 1999) and are an important energy source for salmonid fishes (Rinne 1996; Malison and Baxter 2010b). Salvage logging had an additive effect on the severity of the wildfire disturbance, the burned and salvage-logged catchments had poorer water quality and higher sediment loads than burned catchments. If the decision is made to apply management treatments following wildfire, best practices should be followed to minimise habitat degradation and sediment inputs, particularly in sensitive catchments (Wagenbrenner et al. 2016).
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
Funding for the Southern Rockies Watershed Project was provided by Alberta Agriculture and Forestry – Forest Management Division, Alberta Innovates, and the Natural Sciences and Engineering Research Council of Canada (NSERC). We are especially grateful to Kevin Bladon for technical assistance, Samantha Karpyshin and Ashley Peter-Rennich for field and laboratory assistance, Evan Luchkow for invertebrate identification and Zhuoyan Song for data analysis and statistical assistance. We thank the anonymous reviewers whose insights greatly improved this manuscript.
References
Aloi J (1990) A critical review of recent freshwater periphyton field methods. Canadian Journal of Fisheries and Aquatic Sciences 47, 656–670.| A critical review of recent freshwater periphyton field methods.Crossref | GoogleScholarGoogle Scholar |
Bailey RC, Norris RH, Reynoldson TB (2001) Taxonomic resolution of benthic macroinvertebrate communities in bioassessments. Journal of the North American Benthological Society 20, 280–286.
| Taxonomic resolution of benthic macroinvertebrate communities in bioassessments.Crossref | GoogleScholarGoogle Scholar |
Barbour MT, Gerritsen J, Snyder BD, Stribling JB (1999) Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish, second edn. US Environmental Protection Agency, Office of Water, Report EPA 841-B-99–002. (Washington, DC, USA)
Beschta RL, Frissell CA, Gresswell RE, Hauer FR, Karr JR, Minshall GW, Perry DA, Rhodes JJ (1995) ‘Wildfire and salvage logging: recommendations for ecologically sound post-fire salvage logging and other post-fire treatments on Federal lands in the west.’ (Oregon State University: Corvallis, OR, USA)
Bixby R, Cooper S, Gresswell R, Brown L, Dahm C, Dwire K (2015) Fire effects on aquatic ecosystems: an assessment of the current state of the science. Freshwater Science 34, 1340–1350.
| Fire effects on aquatic ecosystems: an assessment of the current state of the science.Crossref | GoogleScholarGoogle Scholar |
Bjornn T, Brusven M, Molnau M, Milligan J, Klamt R, Cacho E, Schaye C (1977) Transport of granitic sediment in streams and its effects on insects and fish. Idaho Forest Wildfire and Range Experiment Station Bulletin 17, 11–20.
Bladon K, Silins U, Wagner M, Stone M, Emelko M, Mendoza C, Devito K, Boon S (2008) Wildfire impacts on nitrogen concentration and production from headwater streams in southern Alberta’s Rocky Mountains. Canadian Journal of Forest Research 38, 2359–2371.
| Wildfire impacts on nitrogen concentration and production from headwater streams in southern Alberta’s Rocky Mountains.Crossref | GoogleScholarGoogle Scholar |
Borcard D, Gillet F, Legendre P (2011) ‘Numerical ecology with R.’ (Springer: New York, NY, USA)
Bowman M, Bailey R (1997) Does taxonomic resolution affect the multivariate description of the structure of freshwater benthic macroinvertebrate communities? Canadian Journal of Fisheries and Aquatic Sciences 54, 1802–1807.
| Does taxonomic resolution affect the multivariate description of the structure of freshwater benthic macroinvertebrate communities?Crossref | GoogleScholarGoogle Scholar |
Brittain J, Eikeland T (1988) Invertebrate drift – a review. Hydrobiologia 166, 77–93.
| Invertebrate drift – a review.Crossref | GoogleScholarGoogle Scholar |
Brown H (1987) Biology of riffle beetles. Annual Review of Entomology 32, 253–273.
| Biology of riffle beetles.Crossref | GoogleScholarGoogle Scholar |
Clifford HF (1991) ‘Aquatic invertebrates of Alberta: an illustrated guide.’ (University of Alberta Press: Edmonton, AB, Canada)
Covich A, Palmer M, Crowl T (1999) The role of benthic invertebrate species in freshwater ecosystems – zoobenthic species influence energy flows and nutrient cycling. Bioscience 49, 119–127.
| The role of benthic invertebrate species in freshwater ecosystems – zoobenthic species influence energy flows and nutrient cycling.Crossref | GoogleScholarGoogle Scholar |
Emelko M, Silins U, Bladon K, Stone M (2011) Implications of land disturbance on drinking water treatability in a changing climate: demonstrating the need for ‘source water supply and protection’ strategies. Water Research 45, 461–472.
| Implications of land disturbance on drinking water treatability in a changing climate: demonstrating the need for ‘source water supply and protection’ strategies.Crossref | GoogleScholarGoogle Scholar | 20951401PubMed |
Emelko MB, Stone M, Silins U, Allin D, Collins AL, Williams CHS, Martens AM, Bladon KD (2016) Sediment-phosphorus dynamics can shift aquatic ecology and cause downstream legacy effects after wildfire in large river systems. Global Change Biology 22, 1168–1184.
| Sediment-phosphorus dynamics can shift aquatic ecology and cause downstream legacy effects after wildfire in large river systems.Crossref | GoogleScholarGoogle Scholar | 26313547PubMed |
Flannigan M, Logan K, Amiro B, Skinner W, Stocks B (2005) Future area burned in Canada. Climatic Change 72, 1–16.
| Future area burned in Canada.Crossref | GoogleScholarGoogle Scholar |
Giller P, Malmqvist B (1998) ‘The biology of streams and rivers.’ (Oxford University Press: New York, NY, USA)
Gresswell R (1999) Fire and aquatic ecosystems in forested biomes of North America. Transactions of the American Fisheries Society 128, 193–221.
| Fire and aquatic ecosystems in forested biomes of North America.Crossref | GoogleScholarGoogle Scholar |
Hauer F, Locke H, Dreitz V, Hebblewhite M, Lowe W, Muhlfeld C, Nelson C, Proctor M, Rood S (2016) Gravel-bed river floodplains are the ecological nexus of glaciated mountain landscapes. Science Advances 2, e1600026
| Gravel-bed river floodplains are the ecological nexus of glaciated mountain landscapes.Crossref | GoogleScholarGoogle Scholar | 27386570PubMed |
Hauer FR, Spencer CN (1998) Phosphorus and nitrogen dynamics in streams associated with wildfire: a study of immediate and long-term effects. International Journal of Wildland Fire 8, 183–198.
| Phosphorus and nitrogen dynamics in streams associated with wildfire: a study of immediate and long-term effects.Crossref | GoogleScholarGoogle Scholar |
Hauer FR, Stanford JA, Lorang MS (2007) Pattern and process in northern Rocky Mountain headwaters: ecological linkages in the headwaters of the Crown of the Continent. Journal of the American Water Resources Association 43, 104–117.
| Pattern and process in northern Rocky Mountain headwaters: ecological linkages in the headwaters of the Crown of the Continent.Crossref | GoogleScholarGoogle Scholar |
Hawthorn K (2014) The role of fine sediment in phosphorus dynamics and stream productivity in Rocky Mountain headwater streams: possible long-term effects of extensive logging. MSc thesis, University of Alberta, Canada.
Hynes HBN (1975) The stream and its valley. Verhandlungen - Internationale Vereinigung für Theoretische und Angewandte Limnologie 19, 1–15.
| The stream and its valley.Crossref | GoogleScholarGoogle Scholar |
Isaak D, Young M, Luce C, Hostetler S, Wenger S, Peterson E, Hoef J, Groce M, Horan D, Nagel D (2016) Slow climate velocities of mountain streams portend their role as refugia for cold-water biodiversity. Proceedings of the National Academy of Sciences of the United States of America 113, 4374–4379.
| Slow climate velocities of mountain streams portend their role as refugia for cold-water biodiversity.Crossref | GoogleScholarGoogle Scholar | 27044091PubMed |
Jones J, Murphy J, Collins A, Sear D, Naden P, Armitage P (2012) The impact of fine sediment on macro-invertebrates. River Research and Applications 28, 1055–1071.
| The impact of fine sediment on macro-invertebrates.Crossref | GoogleScholarGoogle Scholar |
Kelly E, Schindler D, St Louis V, Donald D, Vlaclicka K (2006) Forest fire increases mercury accumulation by fishes via food web restructuring and increased mercury inputs. Proceedings of the National Academy of Sciences of the United States of America 103, 19380–19385.
| Forest fire increases mercury accumulation by fishes via food web restructuring and increased mercury inputs.Crossref | GoogleScholarGoogle Scholar | 17158215PubMed |
Legendre P, Gallagher E (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129, 271–280.
| Ecologically meaningful transformations for ordination of species data.Crossref | GoogleScholarGoogle Scholar | 28547606PubMed |
Malison R, Baxter C (2010a) The fire pulse: wildfire stimulates flux of aquatic prey to terrestrial habitats driving increases in riparian consumers. Canadian Journal of Fisheries and Aquatic Sciences 67, 570–579.
| The fire pulse: wildfire stimulates flux of aquatic prey to terrestrial habitats driving increases in riparian consumers.Crossref | GoogleScholarGoogle Scholar |
Malison RL, Baxter CV (2010b) Effects of wildfire of varying severity on benthic stream insect assemblages and emergence. Journal of the North American Benthological Society 29, 1324–1338.
| Effects of wildfire of varying severity on benthic stream insect assemblages and emergence.Crossref | GoogleScholarGoogle Scholar |
Mihuc TB, Minshall GW (2005) The trophic basis of reference and post-fire stream food webs 10 years after wildfire in Yellowstone National Park. Aquatic Sciences 67, 541–548.
Mihuc TB, Minshall GW, Robinson CT (1996) Response of benthic macroinvertebrate populations in Cache Creek, Yellowstone National Park to the 1988 wildfires. In ‘The ecological implications of fire in Greater Yellowstone: proceedings on the second biennial Conference on the Greater Yellowstone Ecosystem’, 19–21 September 1993, Yellowstone National Park, WY, USA. (Ed JM Greenlee) International Association of Wildland Fire, pp. 83–94. (Fairfield, WA, USA)
Minshall GW (2003) Responses of stream benthic macroinvertebrates to fire. Forest Ecology and Management 178, 155–161.
| Responses of stream benthic macroinvertebrates to fire.Crossref | GoogleScholarGoogle Scholar |
Minshall GW, Brock JT, Varley JD (1989) Wildfires and Yellowstone stream ecosystems. Bioscience 39, 707–715.
| Wildfires and Yellowstone stream ecosystems.Crossref | GoogleScholarGoogle Scholar |
Minshall GW, Robinson CT, Lawrence DE (1997) Post-fire responses of lotic ecosystems in Yellowstone National Park, USA. Canadian Journal of Fisheries and Aquatic Sciences 54, 2509–2525.
| Post-fire responses of lotic ecosystems in Yellowstone National Park, USA.Crossref | GoogleScholarGoogle Scholar |
Minshall GF, Brock JT, Andrews DA, Robinson CT (2001a) Water quality, substratum and biotic responses of five central Idaho (USA) streams during the first year following the Mortar Creek fire. International Journal of Wildland Fire 10, 185–199.
| Water quality, substratum and biotic responses of five central Idaho (USA) streams during the first year following the Mortar Creek fire.Crossref | GoogleScholarGoogle Scholar |
Minshall GW, Robinson CT, Lawrence DE, Andrews DA, Brock JT (2001b) Benthic macroinvertebrate assemblages in five central Idaho (USA) streams over a 10-year period following disturbance by wildfire. International Journal of Wildland Fire 10, 201–213.
| Benthic macroinvertebrate assemblages in five central Idaho (USA) streams over a 10-year period following disturbance by wildfire.Crossref | GoogleScholarGoogle Scholar |
Moody J, Martin D (2001) Initial hydrologic and geomorphic response following a wildfire in the Colorado Front Range. Earth Surface Processes and Landforms 26, 1049–1070.
| Initial hydrologic and geomorphic response following a wildfire in the Colorado Front Range.Crossref | GoogleScholarGoogle Scholar |
Morris E, Caruso T, Buscot F, Fischer M, Hancock C, Maier T, Meiners T, Mulle C, Obermaier E, Prati D, Socher S, Sonnemann I, Waschke N, Wubet T, Wurst S, Rillig M (2014) Choosing and using diversity indices: insights for ecological applications from the German Biodiversity Exploratories. Ecology and Evolution 4, 3514–3524.
| Choosing and using diversity indices: insights for ecological applications from the German Biodiversity Exploratories.Crossref | GoogleScholarGoogle Scholar | 25478144PubMed |
Naman S, Rosenfeld J, Richardson J (2016) Causes and consequences of invertebrate drift in running waters: from individuals to populations and trophic fluxes. Canadian Journal of Fisheries and Aquatic Sciences 73, 1292–1305.
| Causes and consequences of invertebrate drift in running waters: from individuals to populations and trophic fluxes.Crossref | GoogleScholarGoogle Scholar |
Oksanen JF, Blanchet G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Henry M, Stevens H, Szoecs E, Wagner H (2019) vegan: community ecology package. R package version 2.5–4. R Foundation for Statistical Computing. (Vienna, Austria) Available at https://CRAN.R-project.org/package=vegan [Verified 30 July 2019]
R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing. (Vienna, Austria) Available at https://www.R-project.org/ [Verified 30 July 2019]
Reid D, Quinn J, Wright-Stow A (2010) Responses of stream macroinvertebrate communities to progressive forest harvesting: influences of harvest intensity, stream size and riparian buffers. Forest Ecology and Management 260, 1804–1815.
| Responses of stream macroinvertebrate communities to progressive forest harvesting: influences of harvest intensity, stream size and riparian buffers.Crossref | GoogleScholarGoogle Scholar |
Rice EW (2012) ‘Standard methods for the examination of water and wastewater.’ (American Public Health Association: Washington, DC, USA)
Richards C, Minshall GW (1992) Spatial and temporal trends in stream macroinvertebrate communities – the influence of catchment disturbance. Hydrobiologia 241, 173–184.
| Spatial and temporal trends in stream macroinvertebrate communities – the influence of catchment disturbance.Crossref | GoogleScholarGoogle Scholar |
Rinne JN (1996) Management briefs: short-term effects of wildfire on fishes and aquatic macroinvertebrates in the south-western United States. North American Journal of Fisheries Management 16, 653–658.
| Management briefs: short-term effects of wildfire on fishes and aquatic macroinvertebrates in the south-western United States.Crossref | GoogleScholarGoogle Scholar |
Robson B, Chester E, Matthews T, Johnston K (2018) Post-wildfire recovery of invertebrate diversity in drought-affected headwater streams. Aquatic Sciences 80, 21
| Post-wildfire recovery of invertebrate diversity in drought-affected headwater streams.Crossref | GoogleScholarGoogle Scholar |
Roby KB (1989) Watershed response and recovery from the Will Fire: ten years of observation. USDA Forest Service, Pacific Southwest Forest and Range Experiment Station, General Technical Report PSW-109, pp. 131–136. (Berkeley, CA, USA)
Romme WH, Boyce MS, Gresswell R, Merrill EH, Minshall GW, Whitlock C, Turner MG (2011) Twenty years after the 1988 Yellowstone fires: lessons about disturbance and ecosystems. Ecosystems 14, 1196–1215.
| Twenty years after the 1988 Yellowstone fires: lessons about disturbance and ecosystems.Crossref | GoogleScholarGoogle Scholar |
Rugenski AT, Minshall GW (2014) Climate-moderated responses to wildfire by macroinvertebrates and basal food resources in montane wilderness streams. Ecosphere 5, art25
| Climate-moderated responses to wildfire by macroinvertebrates and basal food resources in montane wilderness streams.Crossref | GoogleScholarGoogle Scholar |
Schoennagel T, Veblen T, Romme W (2004) The interaction of fire, fuels, and climate across Rocky Mountain forests. Bioscience 54, 661–676.
| The interaction of fire, fuels, and climate across Rocky Mountain forests.Crossref | GoogleScholarGoogle Scholar |
Shakesby R, Doerr S (2006) Wildfire as a hydrological and geomorphological agent. Earth-Science Reviews 74, 269–307.
| Wildfire as a hydrological and geomorphological agent.Crossref | GoogleScholarGoogle Scholar |
Silins U, Stone M, Emelko MB, Bladon KD (2009) Sediment production following severe wildfire and post-fire salvage logging in the Rocky Mountain headwaters of the Oldman River Basin, Alberta. Catena 79, 189–197.
| Sediment production following severe wildfire and post-fire salvage logging in the Rocky Mountain headwaters of the Oldman River Basin, Alberta.Crossref | GoogleScholarGoogle Scholar |
Silins U, Bladon K, Kelly E, Esch E, Spence J, Stone M, Emelko M, Boon S, Wagner M, Williams C, Tichkowsky I (2014) Five-year legacy of wildfire and salvage logging impacts on nutrient runoff and aquatic plant, invertebrate, and fish productivity. Ecohydrology 7, 1508–1523.
| Five-year legacy of wildfire and salvage logging impacts on nutrient runoff and aquatic plant, invertebrate, and fish productivity.Crossref | GoogleScholarGoogle Scholar |
Silins U, Anderson A, Bladon KD, Emelko MB, Stone M, Spencer SA, Williams CHS, Wagner MJ, Martens AM, Hawthorn K (2016) Southern Rockies Watershed Project. Forestry Chronicle 92, 39–42.
| Southern Rockies Watershed Project.Crossref | GoogleScholarGoogle Scholar |
Spencer C, Gabel K, Hauer F (2003) Wildfire effects on stream food webs and nutrient dynamics in Glacier National Park, USA. Forest Ecology and Management 178, 141–153.
| Wildfire effects on stream food webs and nutrient dynamics in Glacier National Park, USA.Crossref | GoogleScholarGoogle Scholar |
Thorp J, Covich A (2009) ‘Ecology and classification of North American freshwater invertebrates, 3rd edn.’ (Academic Press: London, UK)
US EPA (2012) Selected analytical methods for environmental remediation and recovery (SAM) 2012. US EPA EPA/600/R-12/555. (Washington, DC, USA)
Vannote R, Minshall G, Cummins K, Sedell J, Cushing C (1980) River continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37, 130–137.
| River continuum concept.Crossref | GoogleScholarGoogle Scholar |
Verkaik I, Vila-Escale M, Rieradevall M, Baxter CV, Lake PS, Minshall GW, Reich P, Prat N (2015) Stream macroinvertebrate community responses to fire: are they the same in different fire-prone biogeographic regions? Freshwater Science 34, 1527–1541.
| Stream macroinvertebrate community responses to fire: are they the same in different fire-prone biogeographic regions?Crossref | GoogleScholarGoogle Scholar |
Vieira N, Clements W, Guevara L, Jacobs B (2004) Resistance and resilience of stream insect communities to repeated hydrologic disturbances after a wildfire. Freshwater Biology 49, 1243–1259.
| Resistance and resilience of stream insect communities to repeated hydrologic disturbances after a wildfire.Crossref | GoogleScholarGoogle Scholar |
Voshell JR (2002) ‘A guide to common freshwater invertebrates of North America.’ (The McDonald and Woodward Publishing Company: Blacksburg, VA, USA)
Wagenbrenner J, Robichaud P, Brown R (2016) Rill erosion in burned and salvage logged western montane forests: effects of logging equipment type, traffic level, and slash treatment. Journal of Hydrology 541, 889–901.
| Rill erosion in burned and salvage logged western montane forests: effects of logging equipment type, traffic level, and slash treatment.Crossref | GoogleScholarGoogle Scholar |
Wagner MJ, Bladon KD, Silins U, Williams CHS, Martens AM, Boon S, MacDonald RJ, Stone M, Emelko MB, Anderson A (2014) Catchment-scale stream temperature response to land disturbance by wildfire governed by surface–subsurface energy exchange and atmospheric controls. Journal of Hydrology 517, 328–338.
| Catchment-scale stream temperature response to land disturbance by wildfire governed by surface–subsurface energy exchange and atmospheric controls.Crossref | GoogleScholarGoogle Scholar |
Westerling A, Hidalgo H, Cayan D, Swetnam T (2006) Warming and earlier spring increase western US forest wildfire activity. Science 313, 940–943.
| Warming and earlier spring increase western US forest wildfire activity.Crossref | GoogleScholarGoogle Scholar | 16825536PubMed |
Wood P, Armitage P (1997) Biological effects of fine sediment in the lotic environment. Environmental Management 21, 203–217.
| Biological effects of fine sediment in the lotic environment.Crossref | GoogleScholarGoogle Scholar | 9008071PubMed |
Writer J, McCleskey R, Murphy S, Stone M, Collins A, Thoms M (2012) Effects of wildfire on source-water quality and aquatic ecosystems, Colorado Front Range. Wildfire and Water Quality: Processes, Impacts and Challenges 354, 117–122.
Yee D, Kehl S (2015) Order Coleoptera. In ‘Thorp and Covich’s freshwater invertebrates. 4th edn.’ (Eds J Thorp, D Rogers) pp. 1003–1042. (Academic Press: Cambridge, MA, USA)
Zwick P (1992) Stream habitat fragmentation – a threat to biodiversity. Biodiversity and Conservation 1, 80–97.
| Stream habitat fragmentation – a threat to biodiversity.Crossref | GoogleScholarGoogle Scholar |