

Antibacterial properties of oil extracts of black soldier fly larvae reared on bread waste
Mung Kwan Shu A , Cheuk Ming Li B , William Eduardo Furtado A , Qianjun Huang A , Sophie St-Hilaire B and Ákos Kenéz B *
B *
A
B
Abstract
Farming black soldier fly larvae (BSFL) has become an emerging agricultural sector for upcycling food waste into high-quality protein and oil biomass. Depending on the chemical composition of the food waste, the oil extracted from BSFL can reach high levels of lauric acid, a natural antimicrobial compound.
We aimed to evaluate whether feeding a bakery waste-based substrate can enhance the lauric acid synthesis of BSFL. In addition, we aimed to test the antibacterial activity of these BSFL oil extracts against E. coli in vitro.
In a 12-day-long feeding trial, we reared BSFL either on a ground corn-based control substrate (n = 6) or a substrate based on bread waste sourced from a local bakery in Hong Kong (n = 6). We studied the growth performance, crude fat and lauric acid content, and agar well-diffusion assay-based antibacterial activity of oil samples extracted from the larvae.
The BSFL reared on the bread waste had a higher average weight (P < 0.001) and average daily gain (P < 0.001), similar crude fat content (P = 0.17), and higher lauric acid content (P < 0.001) than did the control larvae. Both oil samples moderately inhibited the growth of E. coli in vitro; the inhibition zones were 1.92 ± 1.0 mm and 1.25 ± 0.75 mm (means ± s.d.) big in the bread waste-based and the control oil samples respectively (P = 0.28).
Our results indicated that locally sourced bread waste is a suitable substrate for farming BSFL, providing a sustainable alternative for waste management.
The inclusion of locally produced BSFL oil as a feed ingredient for the local livestock farming sector might be an avenue not only to achieve circular economy, but also to reduce antimicrobial drug use. However, the effects on animal health and productive performance warrant further in vivo experiments.
Keywords: black soldier fly, bread waste, circular economy, E. coli, food-waste recycling, insect farming, lauric acid, sustainable agriculture.
Introduction
Insect farming is an emerging field for the valorisation of organic waste (Singh and Kumari 2019; Jing et al. 2020). Hermetia illucens (common name: black soldier fly; BSF) is one of the insect species that is highly efficient in digesting organic materials such as food waste, without having a significant risk of disease transmission (Jing et al. 2020). Black soldier fly larvae (BSFL) can upcycle various low-value wastes into high-quality protein and fat, which offers a sustainable way of food-waste disposal as opposed to landfilling (Cheng and Lo 2016; Cheng et al. 2017; Jing et al. 2020). The larvae and their by-products have diverse applications such as fertilisers, biodiesel (Singh and Kumari 2019), production animal feed (Makkar et al. 2014) and pet feed (Kotob et al. 2022), which contributes to the formation of a circular economy and carbon neutrality. Because BSFL farming is characterised by low land-use requirements and low environmental impact (Smetana et al. 2023), this can potentially alleviate the food-waste challenge Hong Kong and other highly urbanised areas are facing. Daily, 3300 t of food waste were generated in Hong Kong in 2022 (EPD 2022); however, investigating the quality of BSFL grown on locally available waste streams and exploring practicable applications of BSFL-based products are needed to stimulate the growth of the local BSFL-farming sector (Jing et al. 2020).
Larvae with varying lipid content and fatty acid profiles were obtained when substrates with different carbohydrate, lipid and protein compositions were used to rear them (Makkar et al. 2014). Substrates containing higher concentrations of non-fibre carbohydrates yielded larvae with greater crude fat content (Ewald et al. 2020). The acetyl-CoA from the breakdown of carbohydrates can feed into the fatty acid-synthesis pathways, resulting in de novo synthesis of fatty acids (Hoc et al. 2020). Substrates such as wheat, barley and bread waste have high concentrations of non-fibre carbohydrates and energy densities; hence, they are desirable for increasing the yield of fatty acids from the larvae (Danieli et al. 2019; Cohn et al. 2022).
BSFL are richer in saturated fatty acids than unsaturated fatty acids. Evaluating 11 diets, Ewald et al. (2020) found that lauric acid (C12:0) was consistently the most abundant one, even when only minimal amounts were present in the diet. Lauric acid has the most potent antibacterial activity among medium-chained fatty acids (Borrelli et al. 2021). The exact mechanisms have not been fully elucidated; however, its amphipathic nature causes destabilisation of bacterial cell membranes (Mohamed et al. 2021). At a higher concentration, it has a broad-spectrum activity with multiple molecular targets such as the membrane protein glucosyltransferase (Kumar et al. 2020). In vitro studies have shown that lauric acid has a broad-spectrum antibacterial activity against Staphylococcus aureus, Mycobacterium tuberculosis, Listeria monocytogenes, Clostridium perfringens, Enterococcus faecalis and others (Nitbani et al. 2022). Thus, the lauric acid content of BSFL is a determining factor in their antibacterial properties.
In vivo studies have shown that BSFL meal can offer a sustainable dietary alternative in fish nutrition (Franco et al. 2021; Hua 2021). Dietary lauric acid supplementation enhances the gut microbiota and intestinal health of black sea bream (Ullah et al. 2022), and pathogenic bacteria such as Lactococcus and Vagococcus species are reduced by feeding BSFL to rainbow trout (Biasato et al. 2022). Fatty acids are unlikely to impose selective pressure on bacteria; hence, the supplementation of BSFL oil could help slow down the development of antibiotic resistance in common fish pathogens such as Aeromonas spp. (Yang et al. 2020; Mohamed et al. 2021). Whereas development of sustainable waste management and circular economy has a high priority in Hong Kong (Environment Bureau, The Government of the Hong Kong Special Administrative Region of the People’s Republic of China 2014; Wong 2022), there has been limited research on the lauric acid content of BSFL reared on locally available food-waste streams and the resulting antibacterial properties.
The present study aimed to explore the in vitro antibacterial activity of extracts of BSFL reared on bread waste from Hong Kong bakeries against Escherichia coli. We hypothesised that feeding bread waste can provide adequate carbohydrate substrate to enhance lauric acid synthesis of BSFL, thereby providing antibacterial activity. We tested the in vitro antibacterial activity of the larval extracts against E. coli, in preparation for further evaluation against pathogenic strains and in vivo feeding trials. Our objectives were to (1) compare the lauric acid yield of BSFL reared on bread waste and control standard diet and (2) evaluate the in vitro antibacterial properties of these BSFL oil extracts.
Materials and methods
Feeding trial
The larvae were reared at E-Farm Biotech Inc. (Hong Kong SAR, China), a local insect farm specialised in BSF rearing (http://efb.com.hk/en). Larvae hatched from the farm’s own colony were raised under a controlled environment at 26°C, 70% relative humidity and 12:12 light:dark period on a wheat-bran substrate until they reached Day 7 of age. Then they were transferred to grower trays and assigned to one of the two dietary treatments. The ingredients of the diets are listed in Table 1. The control diet consisted of ground corn and soy pulp (also known as okara; a by-product of tofu and soy milk production), and the experimental diet consisted of unsold bread (white bread loaf) sourced from a local bakery. The feed ingredients were ground to a particle size of ~2 mm and water was added to achieve the desired wet pasty texture of a BSFL-rearing substrate with ~65% moisture content. Both diets were divided into trays of six replicates with 7 kg feed each. The number of replicates was oriented to previously published studies with a comparable design (Meneguz et al. 2018; Truzzi et al. 2020). An estimated number of 10,000 larvae were added to each replicate. The number of larvae was estimated by counting and weighing a subset of the larvae. The trays used in this study measured 38 cm by 56 cm with a depth of 12 cm; hence, the larval density was 4.7 larvae/cm2. The larvae were kept at standard rearing conditions at 26°C, 70% relative humidity and constant darkness.
| Item | Control diet | Experimental diet | |
|---|---|---|---|
| Ground corn (kg) | 17 | 0 | |
| Soy pulp (kg) | 9 | 4 | |
| Bread (kg) | 0 | 22 | |
| Water (kg) | 14 | 14 |
Two 500-gram samples of both diets were submitted to ALS Technichem (HK) Pty Ltd (Hong Kong SAR, China) for proximate composition analysis based on the Association of Official Analytical Chemists (AOAC) methods. The moisture content was analysed on the basis of the AOAC Official Method 950.46B (AOAC 2016a) by oven drying at 103°C; the crude protein was analysed on the basis of the AOAC Official Method 2001.11 (AOAC 2016b); the crude fat was analysed on the basis of the AOAC Official Method 2003.05 (AOAC 2016c); the total dietary fibre was analysed on the basis of the AOAC Official Method 985.29 (AOAC 2016d); the total ash was analysed on the basis of the AOAC Official Method 923.03 (AOAC 2016e) by ashing at 550°C; and the total carbohydrate was determined by calculation (total carbohydrates = dry matter − crude protein − crude fat − total ash).
Larvae oil extraction
The larval weights were measured at the beginning of the trial and on Day 8 and Day 11 of the feeding trial. The larvae were monitored for cuticle colour change and they were harvested when ~10% of them entered the prepupal stage (Liu et al. 2017; Tippayadara et al. 2021). This occurred at 18 days of age. They were separated from the remaining substrate and frass via a rotary vibrating sieve machine. The isolated larvae were desiccated in the microwave at 1000 W (approximately 200°C) for 5 min. A screw oil press with a vacuum oil filter was used to extract the oil from the dried BSFL. The extracts were centrifuged at 1500g for 5 min at room temperature to remove the debris.
Fifty grams of the whole larvae from each replicate were sampled and submitted to ALS Technichem (HK) Pty Ltd for crude fat analysis on the basis of the standard method AOAC Official Method 2003.05 (AOAC 2016c). A volume of 10 mL of the oil sample was taken from each replicate and submitted to the Qingdao Sci-Tech Innovation Quality Testing Co. Ltd (Qingdao, China) for fatty acid analysis. The samples were hydrolysed with pyrogallol, ethanol and hydrochloric acid. Ethanol and a mixture of diethyl ether and petroleum ether were added to the hydrolysed sample in a separatory funnel. The ether layer was further extracted with the same steps for three times and then dried. Saponification and methyl esterification of fatty acids was performed using sodium hydroxide–methanol and boron trifluoride–methanol mixtures. Hexane was used to filter the upper layer with membranes. The filtered sample was analysed using column chromatography (TG-FAME 50 m × 0.25 mm × 0.20 μm) with flame ionisation detection (FID). The relative lauric acid content was expressed as the percentage of the total fatty acids detected in the oil samples.
Evaluation of antibacterial activity
The bacterial strain used was Escherichia coli ATCC 25922 obtained from The Hong Kong Polytechnic University (Hong Kong). A volume of 200 μL of the frozen bacterial stock was inoculated to a tryptic soy agar (TSA) plate and was incubated for 28 h at 37°C. A sterilised inoculation loop was used to pick one colony from the TSA. To culture, the colony was incubated overnight in 15 mL Mueller Hinton (MH) broth at 37°C in a thermomixer with continuous gentle shaking. The absorbance of the overnight culture was adjusted to 0.09–0.1, which was ~400 million colony-forming units (CFU)/mL. All experiments were undertaken under aseptic conditions.
The agar well-diffusion method was used to determine the antibacterial activity of BSFL oil extracts. The techniques were adapted from the study of Marusich et al. (2020). In brief, MH agar plates in 90 mm Petri dishes were inoculated with bacteria. Five wells of 9-mm diameter were bored on the plates with even spacing. All samples of larval oil were warmed up in a 35°C water bath and dissolved in dimethyl sulfoxide (DMSO) in a ratio of 1:1. The solutions were thoroughly vortexed. One positive control was a 30 μg/mL oxytetracycline solution and another positive control was a 0.25 M lauric acid solution (in DMSO). The negative control was DMSO. A volume of 100 μL of each solution was added into the five wells of the plate, namely, DMSO control, oxytetracycline control, lauric acid control, one of the six BSF oil samples of the control group, and one of the six BSF oil samples of the bread waste-based group. The plates were incubated at 37°C for 24 h under aerobic conditions, in duplicate. After incubation, the size of the inhibition zone formed around each well was measured.
Statistical analyses
Shapiro–Wilk test was used to evaluate normal distribution of the data. The weight and composition data had a normal distribution, whereas the agar well-diffusion inhibition-zone data did not pass the normality test. The weight data were evaluated by repeated-measures two-way ANOVA for the treatment and time effects and their interactions. Sidak’s post-test was used to test multiple comparisons between treatments on the individual days. The average daily gain was calculated for the entire 11-day treatment period and evaluated by unpaired Student’s t-test. The crude fat content and lauric acid percentage of the larvae were also evaluated by unpaired Student’s t-test. The inhibition zone data were evaluated by Kruskal–Wallis test with Dunn’s post-test. The level of significance was considered at P < 0.05. All statistical analyses were conducted in GraphPad Prism 10 for Windows (Prism, La Jolla, CA, USA). Data are expressed as means ± standard deviation (s.d.).
Results
The bread waste-based diet had higher protein and ash content and lower crude fat, total carbohydrate and total dietary fibre content than did the control (Table 2). Both diets had a similar dry-matter content.
| Item | Control diet | Experimental diet | |
|---|---|---|---|
| Dry matter (g/100 g) | 35.5 | 35.3 | |
| Crude protein (g/100 g) | 9.63 | 16.24 | |
| Crude fat (g/100 g) | 3.70 | 2.58 | |
| Total carbohydrate (g/100 g) | 72.59 | 69.59 | |
| Total dietary fibre (g/100 g) | 12.35 | 9.28 | |
| Total ash (g/100 g) | 1.73 | 2.32 |
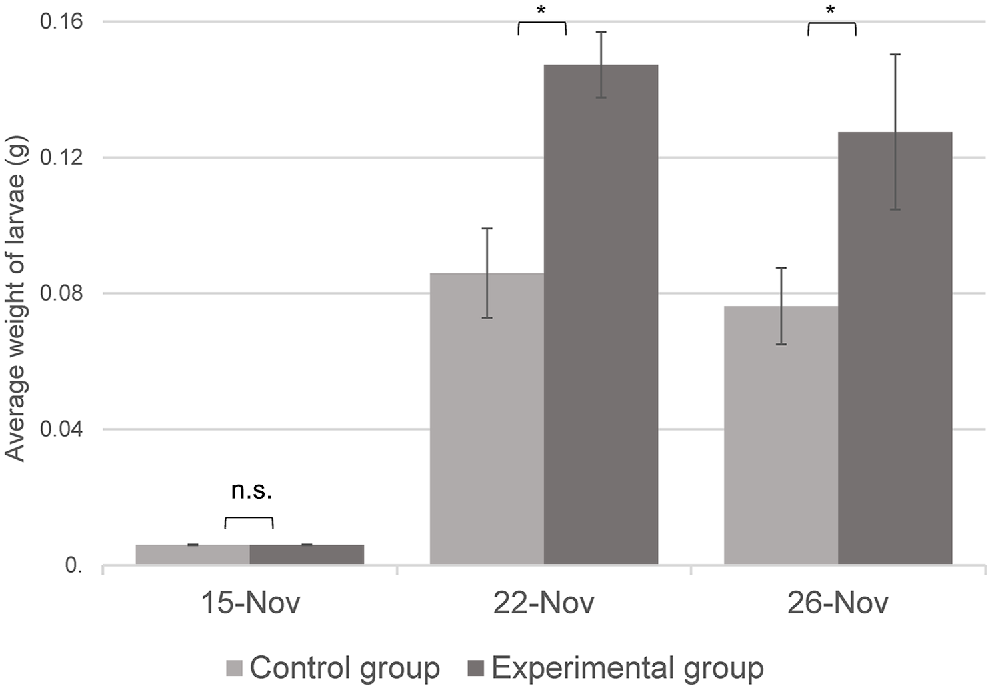
The average weight of the larvae was 0.006 g in both groups on Day 1 of the feeding trial. The average weight of the control group increased by 1266% to 0.076 g, whereas the average weight of the bread waste-based group increased by 2233% to 0.128 g by the end of the trial. Accordingly, the average daily gain considering the entire 11-day-long treatment period was 0.006 g/day and 0.011 g/day in the control and bread waste-based groups respectively (P < 0.001). The ANOVA showed a significant treatment (P < 0.001), time (P < 0.001), and interaction effect (P < 0.001) because the larvae reared on the bread waste-based diet were significantly heavier than those on the control diet on both Day 8 and Day 11 of the trial (both days, P < 0.001 by post hoc test), as shown in Fig. 1.
Bars (mean ± s.d.) indicate the average weight of larvae on 14 November, 21 November and 25 November 2022 in the control group and the bread waste-based (experimental) group. n = 6 (Sidak’s post hoc test: n.s., non-significant; *P < 0.001).
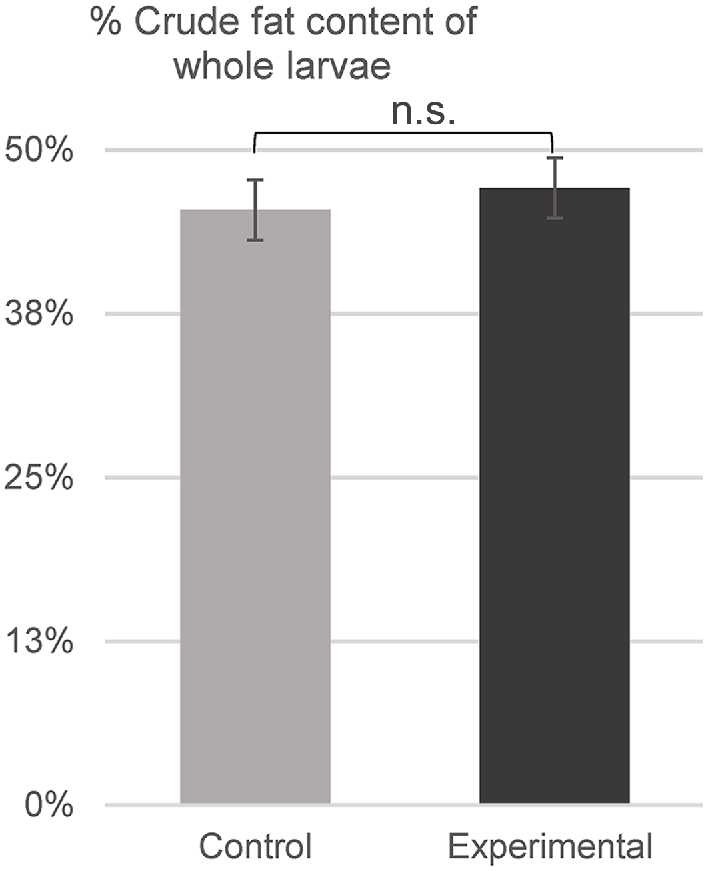
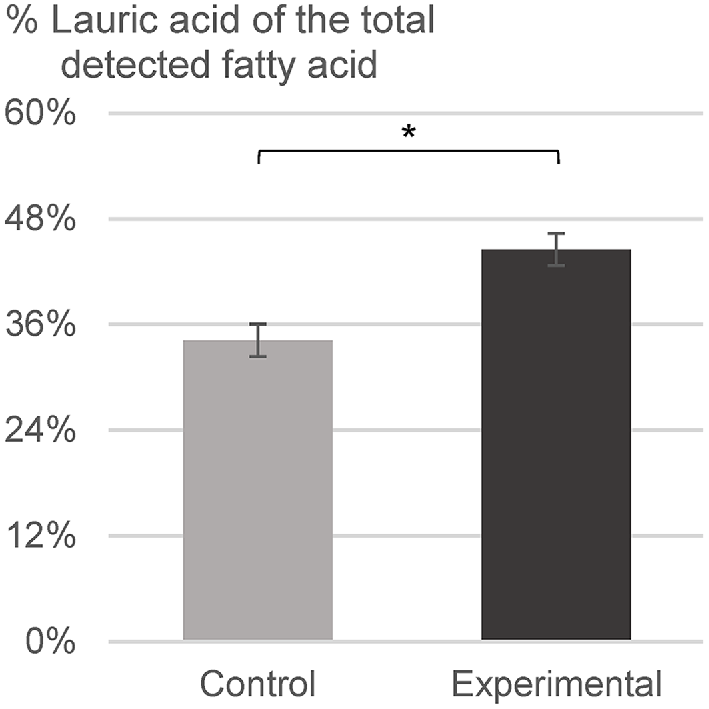
The crude fat content of the whole larvae reared on the control and bread waste-based diets was 45% and 47% respectively, on a dry-matter basis (P = 0.17) (Fig. 2). The relative lauric acid content of the oil samples extracted from the larvae was 34% and 45% in the control and bread waste-based group (P < 0.001) respectively (Fig. 3).
Bars (mean ± s.d.) indicate the percentage of the crude fat content of the whole larvae in the control group and the bread waste-based (experimental) group. n = 6 (t-test: n.s., non-significant).
Bars (mean ± s.d.) indicate the percentage of lauric acid of the total detected fatty acids in the control group and the bread waste-based (experimental) group. n = 6 (*: P < 0.001).
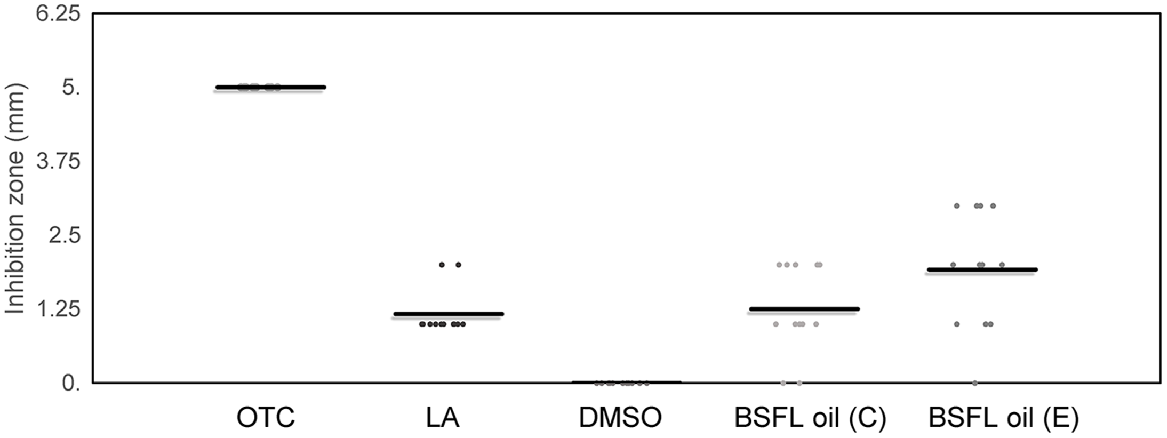
The size of the inhibition zones varied significantly (P < 0.001) between the tested compounds. The positive control (OTC) showed maximum inhibition, causing bigger inhibition than did all other tested compounds (P < 0.01). The average size of the inhibition zones was numerically bigger in the oil of the bread waste-based group (1.92 ± 1.0 mm) than in that of the control group (1.25 ± 0.75 mm), and the lauric acid control (1.17 ± 0.39 mm), as shown in Fig. 4. However, these differences was statistically not significant (P > 0.1). Nevertheless, both tested oils caused greater inhibition than did the negative control DMSO (P < 0.01). A representative image of agar well-diffusion tests is shown in Fig. 5.
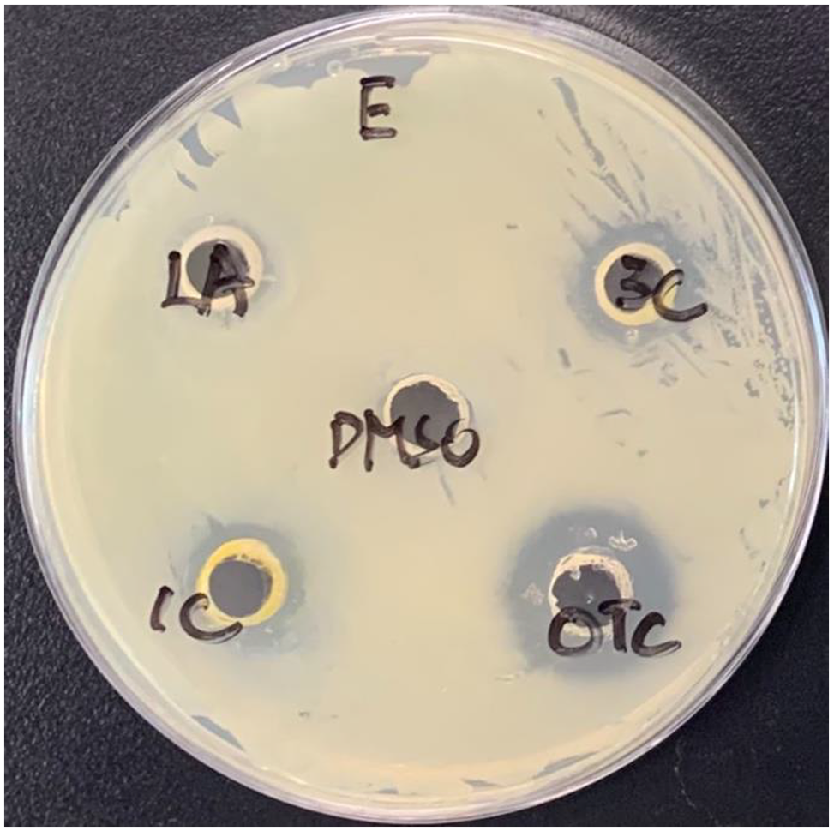
Discussion
The average crude fat content of the larvae reared on the bread waste-based diet and that reared on the control diet were similar, i.e. 47% and 45% respectively. These concentrations are relatively higher than the crude fat content yielded from BSFL reared on vegetable and agro-industrial by-products in previous studies (Meneguz et al. 2018), where the crude fat content ranged from 26.3% to 40.7%. The concentrations observed herein were also higher than the findings of Spranghers et al. (2017), who reported 37.1% and 38.6% fat content in prepupae when reared on vegetable and restaurant wastes respectively. However, the crude fat content in our study was not as high as in BSFL reared on a pure bread diet (57.8% fat content) in the study of Ewald et al. (2020). Out of all fatty acids, 45% was lauric acid in our larvae reared on the bread waste-based diet, which was significantly higher than the 34% lauric acid measured in the control group. Because the crude fat content was similar across the two groups, the absolute lauric acid yield was higher in the bread waste-based group than in the control. Nevertheless, the lauric acid concentrations herein were lower than in the study of Ewald et al. (2020), which reported a lauric acid concentration of 51.8% from the larvae reared on the pure bread substrate.
On the basis of the composition analysis of the two diets in this study, the slightly lower carbohydrate content in the bread waste-based diet (69.59%) than in the control diet (72.59%) was not intentional. Although Meneguz et al. (2018) confirmed the ability of BSFL to bio-convert wastes with a high fibre content because of their intestinal bacteria that can digest cellulose, the lower energy density of the control diet owing to the higher dietary fibre may be the reason for a lower lauric acid ratio (Spranghers et al. 2017). The fact that the lauric acid yield in the present study was lower than the in the bread substrate used in a previous study (Ewald et al. 2020) suggests that the proportion of soy pulp mixed into the bread waste in our bread waste-based diet could be reduced in future studies. Because of a greater soluble carbohydrate content, this could provide more substrate for the citric acid cycle and the fatty acid-synthesis pathways, ultimately leading to an increased lauric acid yield (Hoc et al. 2020). However, it should be noted that different types of bread may have different nutritional compositions, and their effects on the lauric acid synthesis of BSFL require further research.
The age of harvest in the present study was 18 days when the majority of the larvae were close to the prepupal stage, which was comparable to that in similar feeding trials that harvested at the prepupal stage by Cheng et al. (2017), Ewald et al. (2020) and Spranghers et al. (2017). The growth rate and the final biomass yield show variation across studies not only because of differences in substrate compositions and genetic strains but also because of physico-chemical properties of different organic waste streams (Bosch et al. 2020). Moreover, the larval weight reaches a peak as the larvae grow, then decreases as they stop feeding and empty their digestive tract to prepare for pupation (Liu et al. 2017). We have observed this pattern in our weight data and similar trends in the larval weight were displayed in the feeding trials by Cheng et al. (2017) and Meneguz et al. (2018) too. The average final biomass in our bread waste-based group (0.128 g) was slightly lower than in the pure bread substrate-fed larvae (0.137 g) by Ewald et al. (2020), but it was still significantly higher than the average final biomass in our control group (0.076 g). The primary reason for the weight difference observed herein is likely to be the difference in the substrate protein content (9.63 vs 16.24 g/100 g in the control and the bread-waste based diets respectively). Dietary protein content was shown to determine growth rate in BSFL in previous studies comparing several waste streams, whereas carbohydrate content was less correlated with larval weights (Beniers and Graham 2019; Broeckx et al. 2021). Our main protein source was soybean protein and plant-based proteins were discussed to support high growth rate in BSFL (Cammack and Tomberlin 2017). Another reason for the lower growth rate in our control larvae was likely to be the greater dietary fibre content because this was shown to decrease growth rate on the basis of evaluating vegetable and fruit waste-based substrates (Surendra et al. 2020).
The inhibition zones formed in the agar well-diffusion assay were numerically bigger around the oil extracted from the larvae reared on the bread waste-based diet, than in the control diet, and also bigger than the inhibition zone of the lauric acid control. This can be explained by the higher lauric acid content in the larval oil extracted from the bread waste-based group. The weaker antibacterial activity from the lauric acid control may be due to the low concentration (0.25 M), or the presence of other bioactive substances in the oil, in addition to lauric acid. Cases in point are palmitic acid (C16:0) (Kumar et al. 2020), and other unidentified chemical compounds exerting a synergistic effect with the fatty acids in the larval extracts (Park et al. 2014). The in vitro antibacterial efficacy against E. coli is consistent with studies by Auza et al. (2020) and Harlystiarini et al. (2019) in spite of the different BSFL extraction techniques used. The antibacterial activity detected in the present study demonstrated that adequate bioactive substances can be obtained even by simple mechanical pressing of the larvae, which may be a technically easier alternative to multi-step chemical extractions methods used in other studies (Marusich et al. 2020; Mohamed et al. 2021).
Conclusions
The bread waste available in Hong Kong is suitable for BSFL rearing, to enhance the lauric acid content and provide antibacterial efficacy against E. coli in vitro. The collaboration between local bakeries and insect farms could be a promising avenue for the valorisation of food waste. The potential for large-scale food recycling and the application of insect products still warrants further research.
Declaration of funding
This research was supported by internal funding from City University of Hong Kong (grant number 7005878).
Acknowledgements
The authors thank Ka Ming Chan and Zorro Cheng at E-Farm Biotech (Hong Kong, https://efb.com.hk/efb) for providing the larvae and the resources at the farm for the purpose of this research.
References
AOAC (2016b) AOAC Official Method 2001.11. Protein (crude) in animal feed, forage (plant tissue), grain, and oilseeds block digestion method using copper catalyst and steam distillation into boric acid, in ‘Official methods of analysis of AOAC International,’ 20th ed. pp. 24–35. (AOAC International: Gaithersburg, MD, USA)
Auza FA, Purwanti S, Syamsu JA, Natsir A (2020) Antibacterial activities of black soldier flies (Hermetia illucens. l) extract towards the growth of Salmonella typhimurium, E.coli and Pseudomonas aeruginosa. IOP Conference Series: Earth and Environmental Science 492, 012024.
| Crossref | Google Scholar |
Beniers JJA, Graham RI (2019) Effect of protein and carbohydrate feed concentrations on the growth and composition of black soldier fly (Hermetia illucens) larvae. Journal of Insects as Food and Feed 5, 193-199.
| Crossref | Google Scholar |
Biasato I, Chemello G, Oddon SB, Ferrocino I, Corvaglia MR, Caimi C, Resconi A, Paul A, van Spankeren M, Capucchio MT, Colombino E, Cocolin L, Gai F, Schiavone A, Gasco L (2022) Hermetia illucens meal inclusion in low-fishmeal diets for rainbow trout (Oncorhynchus mykiss): effects on the growth performance, nutrient digestibility coefficients, selected gut health traits, and health status indices. Animal Feed Science and Technology 290, 115341.
| Crossref | Google Scholar |
Borrelli L, Varriale L, Dipineto L, Pace A, Menna LF, Fioretti A (2021) Insect derived lauric acid as promising alternative strategy to antibiotics in the antimicrobial resistance scenario. Frontiers in Microbiology 12, 620798.
| Crossref | Google Scholar |
Bosch G, Oonincx DGAB, Jordan HR, Zhang J, van Loon JJA, van Huis A, Tomberlin JK (2020) Standardisation of quantitative resource conversion studies with black soldier fly larvae. Journal of Insects as Food and Feed 6, 95-109.
| Crossref | Google Scholar |
Broeckx L, Frooninckx L, Slegers L, Berrens S, Noyens I, Goossens S, Verheyen G, Wuyts A, Van Miert S (2021) Growth of black soldier fly larvae reared on organic side-streams. Sustainability 13, 12953.
| Crossref | Google Scholar |
Cammack J, Tomberlin J (2017) The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 8, 56.
| Crossref | Google Scholar |
Cheng JYK, Lo IMC (2016) Investigation of the available technologies and their feasibility for the conversion of food waste into fish feed in Hong Kong. Environmental Science and Pollution Research 23, 7169-7177.
| Crossref | Google Scholar |
Cheng JYK, Chiu SLH, Lo IMC (2017) Effects of moisture content of food waste on residue separation, larval growth and larval survival in black soldier fly bioconversion. Waste Management 67, 315-323.
| Crossref | Google Scholar |
Cohn Z, Latty T, Abbas A (2022) Understanding dietary carbohydrates in black soldier fly larvae treatment of organic waste in the circular economy. Waste Management 137, 9-19.
| Crossref | Google Scholar |
Danieli PP, Lussiana C, Gasco L, Amici A, Ronchi B (2019) The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 9, 178.
| Crossref | Google Scholar |
Environment Bureau, The Government of the Hong Kong Special Administrative Region of the People’s Republic of China (2014) A food waste and yard waste plan for Hong Kong 2014–2022. Available at https://www.eeb.gov.hk/sites/default/files/pdf/FoodWastePolicyEng.pdf [accessed 01 March 2023]
EPD (2022) Monitoring of solid waste in Hong Kong – waste statistics for 2021. Available at https://www.wastereduction.gov.hk/sites/default/files/resources_centre/waste_statistics/mms2021_eng.pdf [Accessed 1 May 2023]
Ewald N, Vidakovic A, Langeland M, Kiessling A, Sampels S, Lalander C (2020) Fatty acid composition of black soldier fly larvae (Hermetia illucens) – possibilities and limitations for modification through diet. Waste Management 102, 40-47.
| Crossref | Google Scholar |
Franco A, Scieuzo C, Salvia R, Petrone AM, Tafi E, Moretta A, Schmitt E, Falabella P (2021) Lipids from Hermetia illucens, an innovative and sustainable source. Sustainability 13, 10198.
| Crossref | Google Scholar |
Harlystiarini H, Mutia R, Wibawan IWT, Astuti DA (2019) In vitro antibacterial activity of black soldier fly (Hermetia illucens) larva extracts against Gram-negative bacteria. Buletin Peternakan 43, 125-129.
| Crossref | Google Scholar |
Hoc B, Genva M, Fauconnier M-L, Lognay G, Francis F, Caparros Megido R (2020) About lipid metabolism in Hermetia illucens (L. 1758): on the origin of fatty acids in prepupae. Scientific Reports 10, 11916.
| Crossref | Google Scholar |
Hua K (2021) A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 530, 735732.
| Crossref | Google Scholar |
Jing R, Liu T, Tian X, Rezaei H, Yuan C, Qian J, Zhang Z (2020) Sustainable strategy for municipal solid waste disposal in Hong Kong: current practices and future perspectives. Environmental Science and Pollution Research 27, 28670-28678.
| Crossref | Google Scholar |
Kotob G, Sluczanowski N, Anusha Siddiqui S, Martin Tome N, Dalim M, van der Raad P, Aarts K, Paul A (2022) Potential application of black soldier fly fats in canine and feline diet formulations: a review of literature. Journal of Asia-Pacific Entomology 25, 101994.
| Crossref | Google Scholar |
Kumar P, Lee J-H, Beyenal H, Lee J (2020) Fatty acids as antibiofilm and antivirulence agents. Trends in Microbiology 28, 753-768.
| Crossref | Google Scholar |
Liu X, Chen X, Wang H, Yang Q, ur Rehman K, Li W, Cai M, Li Q, Mazza L, Zhang J, Yu Z, Zheng L (2017) Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 12, e0182601.
| Crossref | Google Scholar |
Makkar HPS, Tran G, Heuzé V, Ankers P (2014) State-of-the-art on use of insects as animal feed. Animal Feed Science and Technology 197, 1-33.
| Crossref | Google Scholar |
Marusich E, Mohamed H, Afanasev Y, Leonov S (2020) Fatty acids from Hermetia illucens larvae fat inhibit the proliferation and growth of actual phytopathogens. Microorganisms 8, 1423.
| Crossref | Google Scholar |
Meneguz M, Schiavone A, Gai F, Dama A, Lussiana C, Renna M, Gasco L (2018) Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae: rearing substrate effects on performance and nutritional composition of black soldier fly. Journal of the Science of Food and Agriculture 98, 5776-5784.
| Crossref | Google Scholar |
Mohamed H, Marusich E, Afanasev Y, Leonov S (2021) Fatty acids-enriched fractions of Hermetia illucens (Black Soldier Fly) larvae fat can combat MDR pathogenic fish bacteria Aeromonas spp. International Journal of Molecular Sciences 22, 8829.
| Crossref | Google Scholar |
Nitbani FO, Tjitda PJP, Nitti F, Jumina J, Detha AIR (2022) Antimicrobial properties of lauric acid and monolaurin in virgin coconut oil: a review. ChemBioEng Reviews 9, 442-461.
| Crossref | Google Scholar |
Park S-I, Chang BS, Yoe SM (2014) Detection of antimicrobial substances from larvae of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Entomological Research 44, 58-64.
| Crossref | Google Scholar |
Singh A, Kumari K (2019) An inclusive approach for organic waste treatment and valorisation using black soldier fly larvae: a review. Journal of Environmental Management 251, 109569.
| Crossref | Google Scholar |
Smetana S, Bhatia A, Batta U, Mouhrim N, Tonda A (2023) Environmental impact potential of insect production chains for food and feed in Europe. Animal Frontiers 13, 112-120.
| Crossref | Google Scholar |
Spranghers T, Ottoboni M, Klootwijk C, Ovyn A, Deboosere S, De Meulenaer B, Michiels J, Eeckhout M, De Clercq P, De Smet S (2017) Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. Journal of the Science of Food and Agriculture 97, 2594-2600.
| Crossref | Google Scholar |
Surendra KC, Tomberlin JK, van Huis A, Cammack JA, Heckmann L-HL, Khanal SK (2020) Rethinking organic wastes bioconversion: evaluating the potential of the black soldier fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae) (BSF). Waste Management 117, 58-80.
| Crossref | Google Scholar |
Tippayadara N, Dawood MAO, Krutmuang P, Hoseinifar SH, Doan HV, Paolucci M (2021) Replacement of fish meal by black soldier fly (Hermetia illucens) larvae meal: effects on growth, haematology, and skin mucus immunity of Nile Tilapia, Oreochromis niloticus. Animals 11, 193.
| Crossref | Google Scholar |
Truzzi C, Giorgini E, Annibaldi A, Antonucci M, Illuminati S, Scarponi G, Riolo P, Isidoro N, Conti C, Zarantoniello M, Cipriani R, Olivotto I (2020) Fatty acids profile of black soldier fly (Hermetia illucens): influence of feeding substrate based on coffee-waste silverskin enriched with microalgae. Animal Feed Science and Technology 259, 114309.
| Crossref | Google Scholar |
Ullah S, Zhang J, Xu B, Tegomo AF, Sagada G, Zheng L, Wang L, Shao Q (2022) Effect of dietary supplementation of lauric acid on growth performance, antioxidative capacity, intestinal development and gut microbiota on black sea bream (Acanthopagrus schlegelii). PLoS ONE 17, e0262427.
| Crossref | Google Scholar |
Wong MH (2022) Integrated sustainable waste management in densely populated cities: the case of Hong Kong. Sustainable Horizons 2, 100014.
| Crossref | Google Scholar |
Yang C, Song G, Lim W (2020) A review of the toxicity in fish exposed to antibiotics. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 237, 108840.
| Crossref | Google Scholar |