

Non-invasive monitoring and reintroduction biology of the brush-tailed rock-wallaby (Petrogale penicillata) in the Grampians National Park, Australia
S. Kleemann A * , D. Sandow B , M. Stevens C , D. J. Schultz D , D. A. Taggart E F and A. Croxford F
A * , D. Sandow B , M. Stevens C , D. J. Schultz D , D. A. Taggart E F and A. Croxford F
A School of Biological Science, University of Adelaide (Waite Campus), Urrbrae, SA 5064, Australia.
B Landscape South Australia - Northern and Yorke, Clare, SA 5435, Australia.
C Landscape South Australia - Limestone Coast, Mount Gambier, SA 5290, Australia.
D Schultz Foundation, 38 Wood Street, Millswood, SA 5034, Australia.
E School of Animal and Veterinary Science, University of Adelaide (Waite Campus), Urrbrae, SA 5064, Australia.
F Fauna Research Alliance, PO Box 98, Callaghan, NSW 2308, Australia.
Australian Journal of Zoology 69(2) 41-54 https://doi.org/10.1071/ZO21009
Submitted: 24 July 2020 Accepted: 13 December 2021 Published: 8 February 2022
© 2021 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC)
Abstract
Thirty-nine endangered brush-tailed rock-wallabies (Petrogale penicillata) were reintroduced to Grampians National Park, western Victoria, between 2008 and 2012. Subsequent high mortality, low breeding, and no recruitment were linked to fox predation and physical disturbance during monitoring. From 2014 to 2017, the colony was left undisturbed and monitored only by remote camera. Five adult animals were identified across this period (1 ♂ and 3 ♀s – all tagged; and one untagged female), and an average of 0.7 pouch young were birthed per tagged female per year. In 2019, camera-monitoring and non-invasive genetic monitoring (faecal) were used to identify colony members, genetic diversity, and breeding. Camera monitoring in 2019 identified the same five individuals, whereas genetic monitoring using 12 microsatellites identified eight individuals (two male and six female genotypes). Genetic diversity within the colony was moderate (expected heterozygosity (He) = 0.655, observed heterozygosity (Ho) = 0.854). Leaving the colony undisturbed after 2013 correlated with improved adult survival, increased breeding, and successful recruitment of young to the population. Recommendations for the Grampians colony include continuation of regular camera- and scat monitoring to improve our understanding of the reintroduction biology of P. penicillata and other marsupials in open, unfenced landscapes.
Keywords: camera trap, conservation genetics, genetic monitoring, macropod, marsupial, reintroduction biology, Petrogale, threatened species.
Introduction
Reintroduction is a common conservation tool to help threatened species and prevent wildlife extinctions. However, the success rates of reintroductions are highly variable. The factors that influence success or failure are numerous and vary with species and release site (Bubac et al. 2019; Berger-Tal et al. 2020). Within Australia, the main factor identified in the failure of many reintroductions is predation by exotic carnivores (e.g. Moseby et al. 2011; Sheean et al. 2012; Hardman et al. 2016; Short 2016; Watkins et al. 2018; Robinson et al. 2020).
While post-release monitoring of reintroduction attempts is now frequently conducted, it is often conducted poorly, and difficulties with monitoring are commonly reported (Bubac et al. 2019; Berger-Tal et al. 2020; Gant et al. 2020). Survival, abundance, population growth, breeding, and recruitment are all parameters that determine reintroduction success and can be assessed only through post-release monitoring (Richards and Short 2003; White et al. 2003; Parlato and Armstrong 2013; Johnson et al. 2018). Although genetic diversity is highly valued when selecting groups of animals for release (e.g. Lapbenjakul et al. 2017), without post-release monitoring, it cannot be known whether the genetics of the reintroduced population remain viable for long-term success (Zeng et al. 2007; Scott et al. 2020). Post-release monitoring data are vital to guide management decisions such as interventions to ensure population viability, understanding why a reintroduction succeeded or failed, to assess effectiveness of predator control, and to optimise future reintroduction attempts (Richards and Short 2003).
Monitoring reintroduced animals through observation, trapping, or mark–recapture can be difficult in steep or hard-to-access terrain, or when animals are cryptic or shy, and can have occupational safety risks to researchers and animal welfare risks for the study species (Piggott et al. 2006a; Berger-Tal et al. 2020). Trap-shy animals can lead to the detection of only a subset of a population (Hoyle et al. 1995; Cutler and Swann 1999; Wegge et al. 2004).
Alternatively, non-invasive wildlife monitoring requires no contact with animals and can often be preferable owing to its efficacy in labour, cost, and animal and staff welfare (De Bondi et al. 2010). Camera traps can be used to obtain information on population abundance, behaviour, habitat occupancy, and population structure (Silveira et al. 2003; Claridge et al. 2010). Non-invasive genetic monitoring, including the use of faecal samples, can provide information on population abundance, population structure, sex, diet, habitat occupancy, and population genetic diversity (Piggott et al. 2006a; Lachish et al. 2011; Tende et al. 2014; Moßbrucker et al. 2015; Morales-Contreras et al. 2016; Camp et al. 2020; Bannister et al. 2020).
The brush-tailed rock-wallaby (BTRW; Petrogale penicillata) is one of many currently declining, threatened mammal species in Australia (Woinarski et al. 2014). The small to medium-sized, cryptic macropod species historically occurred in abundance throughout much of the Great Dividing Range and has a strong dependence on complex rocky habitat (Wakefield 1961; Kaufmann 1974). In the Grampians, Victoria, the species became locally extinct in 1999 and, in response, a translocation strategy was devised to reintroduce captive-bred animals into the Moora Moora Creek area within the Grampians National Park (Delaney et al. 2005; Bramwell et al. 2008; Taggart et al. 2008, 2015).
Reintroduction of 39 animals began in November 2008 and concluded in November 2012, with only seven animals surviving by December 2013 (Taggart et al. 2015). Predation by exotic predators was recognised as a key cause of mortality, despite the presence of an intensive fox control program around the site (Robley et al. 2014; Taggart et al. 2015). The frequency of colony disruption from supplementations, physical monitoring (e.g. cage-trapping and handling) and release group size were hypothesised to result in poor levels of social cohesion during the establishment phase, leading to increased predation vulnerability (Taggart et al. 2015).
Few pouch young were identified within the colony between 2008 and 2013, no juveniles or subadults were observed or trapped, and no animals were recruited into the adult population (M. Stevens, Parks Victoria, pers. comm.). From 2014 onward, the colony was left undisturbed by physical monitoring and supplementation and was monitored only by remote camera.
Non-invasive faecal genetics was pioneered in this species by Piggott and colleagues to test its potential for monitoring elusive or endangered species, especially in rough terrain (e.g. Browning et al. 2001; Piggott and Taylor 2003; Eldridge et al. 2004; Hazlitt et al. 2004, 2006a, 2006b, 2014; Piggott 2004; Piggott et al. 2006a, 2006b, 2018).
The aim of this study was to examine the efficacy of remote cameras and faecal genotyping as non-invasive post-release monitoring methods to better understand the population structure, current distribution, recruitment, genetic diversity, and parentage of the remaining reintroduced colony. This enabled us to assess the difference in the population growth following a period of 6 years with no animal supplementations or other human interference, which can provide useful insights to help optimise reintroduction of BTRW and similar species.
Materials and methods
Study site
This study was located at Moora Moora Creek, within the Grampians National Park in western Victoria, Australia (Fig. 1). The reintroduced BTRW colony is located centrally within the national park, approximately 10 km southwest of the township of Halls Gap.

|
Camera-monitoring data
Three motion triggered remote cameras (BuckEye Orion XIR; BuckEye Cam, Australia) were installed at the Moora Moora Creek release site by Parks Victoria staff in 2014. Images recorded by these cameras were used to determine successful matings and breeding of the BTRW in the colony from 2014 to mid-2017 and to monitor the ongoing survival of tagged animals. Cameras were not operational between mid-2017 and December 2018.
Previously published BTRW growth data were used to calculate the approximate dates of birth of the pouch young recorded by remote cameras in the colony. Calculations were based on estimates of gestation length (∼30 days), first pouch exit (∼180 days), permanent pouch exit (190–210 days) and weaning (220–240 days; Taggart et al. 2005).
In January 2019, six new motion-triggered remote cameras (BuckEye X80; BuckEye Cam Australia) were installed at Moora Moora Creek. The new cameras were set up ∼250 m apart, along a 1.2 km length of the Moora Moora Creek escarpment above Moora Moora Creek where the BTRWs were released, and which now forms the main colony site. Cameras uploaded photos, via a cellular base station, directly to an office computer on weekdays.
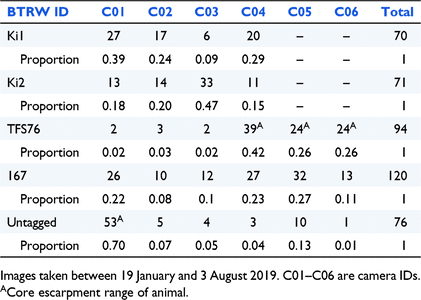
Recorded sightings of BTRWs on the cameras from the 19 January 2019 to the 3 August 2019 were obtained from Parks Victoria staff. The identity of the BTRW based on clear visualisation of an identifying ear tag or definite absence of an ear tag was provided with the date and camera location of the sighting.
Camera locations and BTRW ear-tag identity were used to estimate the escarpment length occupied by each individual BTRW (referred to as their ‘escarpment range’). The escarpment range of each animal is defined as the number of remote cameras on which an individual was captured, signifying how much of the escarpment each individual was using. Home-range studies of rock-wallaby species have typically found that core ranges were best estimated using 50–75% of radio tracking/GPS/spotlight locations (Horsup 1994; Laws and Goldizen 2003; Telfer and Griffiths 2006; Sharp 2009; Hayward et al. 2011). For this study, a cumulative frequency across multiple cameras where >60% of sightings were recorded were considered an animal’s ‘core escarpment range’.
For visualisation purposes, a buffer zone of 100 m was created around the camera location where each animal was recorded to represent an area in which an animal was thought to occur using ArcGIS ArcMap 10.8.1 (Esri, USA). Additionally, the escarpment range of an individual was considered identical to its core escarpment range if the proportion of images of an individual recorded at each camera was equal, because it was assumed that the animal spent similar amounts of time around each camera location along the escarpment.
Genetic sample collection
BTRWs are highly dependent on their rocky habitat, and so their scats are easy to locate if present. Rock-wallaby scats can be readily distinguished from other macropod scats in the region (Macropus fuliginosus, M. giganteus, M. rufogriseus and Wallabia bicolor; Aussavy et al. 2011) by their size, shape (typically cylindrical with a single pointed end), and texture (mix of small and larger plant fragments; Jarman and Capararo 1996). Fresh scats (<2 weeks old) can be identified by their glossy, black colour, and strong odour (Piggott 2004; David Taggart and Lauren Werner, pers. comm.).
In this study, fresh BTRW scats were collected opportunistically from across the length of the rocky escarpment area in and around the known colony site (the Moora Moora Creek Escarpment) in September 2018 and February to April 2019. Only one scat, selected at random, was taken from locations where multiple scats were present in close proximity to one another.
The scat search and collection began at the western end of the escarpment, approximately 0.3 km south-west of Camera C01 and concluded 1.13 km past the eastern end of the main colony, including the entire colony (approximately 1.2 km in length; Fig. 1). A 0.08 km length of escarpment on the north-western side of the creek under a landmark known as ‘flat rock’, directly opposite the western end of the main colony, was also searched (Fig. 1). Additionally, an adjacent 2.3 km of escarpment on both the northern and southern sides of Mount Rosea Creek, located north of the known Moora Moora Creek BTRW colony site, was searched in autumn 2019 (Fig. 1). The area searched represented the closest escarpments to the main colony, with potentially suitable habitat that dispersing BTRWs might have moved to.
Scats were collected in individual zip-lock bags to avoid DNA contamination among scats. Scat locations were recorded using a hand-held GPS unit. The scat samples were then stored in a portable cooler while travelling and on site. Scat samples were kept frozen at −20°C in the laboratory before DNA extraction (e.g. Walker et al. 2009; Kolodzieg et al. 2013; Sabino-Marques et al. 2018). For comparison of genetic diversity, DNA extracted from stored ear biopsies of 10 BTRWs (progeny of three males and four females) of known sex were included in the study from the animal facility at the Waite Campus of University of Adelaide, Adelaide, Australia. The translocated animals and the captive Waite Campus animals were founded from the same set of individuals. These founder individuals came from two distinct evolutionarily significant units (ESU), the southern and the central ESU (Browning et al. 2001; Paplinska et al. 2011; David Taggart, pers. comm.).
Genomic DNA extraction
Faecal DNA was extracted from scats by using the Bioline Isolate II Genomic DNA Kit protocol (Bioline, Australia), with modifications that included using sterile cotton swabs soaked in 200 μL of nanopure water (Sigma-Aldrich, Australia) to remove epithelial cells from all surfaces of the scats (e.g. Akomo-Okoue et al. 2015; Ramón-Laca et al. 2015; Dimsoski 2017; Bourgeois et al. 2019) and using an additional centrifugation step to remove excess debris before binding onto the silica membrane.
Genomic DNA from the ear biopsies was extracted using the Bioline Isolate II Genomic DNA Kit (Bioline, Australia), using approximately 20 mg of each ear biopsy and extending the proteinase digestion at 56°C to 24 h.
Total DNA extracted from faecal and ear biopsy samples were quantified and assessed for quality by using a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific Inc., USA) and stored at −20°C until analysis.
Genotyping
A panel of 12 nuclear microsatellite markers, seven di-nucleotide (PI22 (Luikart et al. 1997), BT_1, BT_3, BT_6, BT_11, BT_17, BT_25; Accession numbers: OK070787, OK070788, OK070789, OK070790, OK070791, OK070792) and five tri-nucleotide (SK8, SK62, SK65; accession numbers: OK031000, OK031001, OK031002; ET32, ET35; unpubl. data, Adam Croxford, pers. comm., Accession numbers: XM_036734328.1, XM_020966173.1; mean He = 0.7) were used to genotype the BTRW faecal DNA (Supplementary Table S1). A subset of six microsatellite markers (PI22, SK8, SK62, SK65, ET32, ET35) (mean He = 0.74) were used to genotype captive ear-tissue DNA due to budget constraints. Two Y-chromosome linked markers (SRY; Watson et al. 1998, SRY; Katsura et al. 2018) were checked for amplification in all samples (tissue and faecal DNA) for sex determination. Flanking primers of unpublished microsatellite markers were designed using Primer3 software (Koressaar and Remm 2007; Untergasser et al. 2012; Koressaar et al. 2018) with a length of between 80 and 300 bp. Markers were selected on the basis of reliability of amplification ease of scoring (low stuttering), and polymorphism. The markers were modified with truncated Illumina linkers to allow the amplicons to be converted into Illumina libraries (sequences: forward: TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG; reverse: GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG). PCRs of the ear-biopsy DNA were conducted in 10 μL volumes containing 1× Biomix (Bioline, Australia), 0.2 nM of each forward and reverse primers, and 10 ng of DNA. The same protocol was used for faecal DNA analysis following a 1 in 10 dilution of the DNA.
Polymerase chain reactions were conducted using a BioRad T100 Thermal Cycler (Bio-Rad, USA), using the following PCR protocol: an initial denaturation at 95°C for 5 min, followed by 35 cycles of 95°C for 15 s, 55°C for 30 s, 72°C for 45 s and a final extension at 72°C for 5 min. The annealing temperature for the Y-linked SRY marker (Watson et al. 1998) was set to 64°C instead of 55°C and the cycle was repeated 40 times for both Y-linked markers. The sexing PCR was performed with ET32 and ET33 to ensure that there were no false negatives. PCR products were visualised after electrophoresis on a 1% agarose gel stained with SybrSafe (Invitrogen, USA).
Polymerase chain reaction products for each individual (including both ear-tissue samples and scat samples) were pooled and purified using Ampure XP beads (Agencourt Bioscience Corporation, USA) at a ratio of 1:1. The two-step Illumina indexing approach described by Wilkinson et al. (2017) was used to develop sequencing libraries for each sample and all samples were pair-end sequenced using a 300-cycle MiSeq version 2 kit (Illumina, USA) at Australian Genome Research Facility (AGRF), Melbourne.
The MiSeq sequences were demultiplexed using the MiSeq Reporter software (Illumina) and paired reads were then merged using the Geneious Prime 2019.2 software (Biomatters, New Zealand). Genotypes were determined from the amplicon length of the microsatellite. To test the fidelity of the genotyping, three replicate amplifications from 10 different samples (two ear-tissue and eight scat DNA samples) were performed and run on the MiSeq, including the single scat genotyped as F6.
Individual identification
Hardy–Weinberg equilibrium and linkage disequilibrium tests were conducted across all loci for all samples, by using the Markov-chain algorithm of Raymond and Rousset (1995) in Genepop version 4.7.5 (Rousset 2008). The presence of null alleles and allele dropout were assessed using the MICRO-CHECKER 2.2.3 program (Van Oosterhout et al. 2004). Owing to budget constraints, samples with non-amplifying loci could not be re-analysed in this study, and were excluded.
The probability of identity (PID), meaning the probability of two individuals having the same genotype in a randomly mating population, was calculated among full siblings (P(ID)sib) for the Moora Moora Creek individuals (12 loci; 1.0 × 10−4) by using GenAlEx 6.5 (Peakall and Smouse 2006, 2012). GenAlEx 6.5 was also used to match genotypes, with zero mismatches allowed. Samples with 100% allelic sharing at all 12 loci were considered to be the same individual because of the low PID values and exclusion of genotypes with non-amplifying loci.
Basic genetic-diversity parameters, including mean number of alleles per locus (Na), observed (Ho) and expected heterozygosity (He), and an inbreeding parameter (Fis) were obtained using GenAlEx 6.5 (Peakall and Smouse 2006, 2012), by using the unique scat genotypes and the 10 tissue samples. Because only six loci were amplified in tissue DNA, genetic diversity parameters were calculated using the six-locus subset for both the Waite (tissue) and Moora Moora Creek (scat) samples and all 12 loci for Moora Moora Creek (scat) samples only.
Parentage analysis
Parentage of scat genotypes (conducted with 12 loci) was assigned using the program Cervus 3.0.7 (Kalinowski et al. 2007). Because the identity of all BTRW genotypes was unknown compared with release records, all genotypes were considered as both offspring and candidate parents with sexes known. Parentage was determined using parent-pair analysis where the simulation was conducted with the following parameters: 10 000 offspring, three candidate mothers with a proportion of 0.9 sampled, two candidate fathers with a proportion of 0.9 sampled, a proportion of 0.9 loci typed and 0.1 loci mistyped, and a minimum typed number of loci of 12. The parent-pair analysis was then conducted with a strict confidence level of 95% and a relaxed confidence level of 80%. Of all parentage analyses, there was one locus pair mismatch between individuals F6 and F3. However, because of the low number of individuals and lack of known relationships, the power to detect error was low. GenAlEx 6.5 was used to calculate the probability of exclusion with no parents known.
Results
Camera-monitoring along the escarpment (2019)
Brush-tailed rock-wallaby was the only macropod species recorded by the remote cameras in 2019. Images of BTRWs in the colony showed that the animals were in good body condition with clean glossy pelts and no obvious signs of disease.
Camera data suggested that at least five BTRWs persisted at the Moora Moora Creek site in early 2019, including four tagged founder animals (1 ♂ and 3 ♀) and an untagged animal. The identity, age, and sex of the known Moora Moora Creek BTRWs are listed in Table 1. During 2019, 76 images of an untagged BTRW were recorded (Table 2). All sightings of the untagged animal suggested that it was a young adult female, including a visible pouch. Tagged male 167 was twice recorded mating an untagged animal at Camera C03 in April and May of 2019. The total escarpment and core escarpment ranges of known BTRWs overlapped (Fig. 2a–e; Table 2).

|
|

|
Camera records of breeding events (2014–2017, 2019)
Breeding events in the colony deduced from camera images of females with pouch young were recorded between 2014 and 2017 (Supplementary Table S2). All tagged adult females (Ki1, Ki2, and TFS76) were detected with pouch young, indicating that all tagged adult females and the tagged male in colony were fertile and breeding. Camera data from 2014 to 2017 and in 2019 indicated that the majority of mating events in the colony occurred in late summer and autumn (69%), with pouch young emergence occurring in late spring and early summer (Supplementary Table S2). Less frequent mating events were also detected on camera in late spring (15%) and winter (15%).
Remote camera images indicated that nine pouch young were born between 2014 and mid-2017. A further five attempted matings were recorded between January and August 2019, including four of unknown outcome and one failed attempt. All three tagged females (Ki1, Ki2 and TFS76) were observed mating with male 167 in 2019. An untagged female was also observed with a pouch young (Supplementary Table S2). No other images of pouch young or young at foot were recorded in 2019.
From the camera observations made between 2014 and 2017, the three tagged adult females produced, on average, 0.7 pouch young per female per year (Supplementary Table S3). No female weaned a young every year between 2014 and 2017. Three pouch young were detected only as a pouch ‘bulge’ late within the observation period and so their status remains unknown. Of six pouch young detected on camera with heads out of pouch, five (83%) reached at least the young-at-foot stage. Two (33%) young were later observed as subadults, three young (50%) were not observed past the young-at-foot stage and the sixth young had just reached pouch emergence when the cameras were removed in mid-2017 (Supplementary Table S2).
Scat DNA genotypes identified
In total, 218 BTRW scats were collected from the colony site (44 collected in September 2018 and 174 collected across February and April 2019). No scats were found more than 250 m away from the known colony area (Fig. 1). No BTRW scats were found in the adjacent Mount Rosea Creek search area, despite the presence of potentially suitable habitat (Fig. 1). In addition to BTRW scats, swamp wallaby (Wallabia bicolor) scats were frequently found at the base and top of the rocky escarpments in and around the known colony site at Moora Moora Creek and in the Mount Rosea Creek area. All Waite BTRWs tested were of known sex. Genotyping confirmed their sex, and thus that the technique was working. DNA extracted from 4 of the 218 scats collected at Moora Moora Creek failed to amplify and it was deemed that the scats were too old to produce viable DNA. Genotypes including loci that did not amplify were excluded from further analysis. This reduced the sample size from 218 to 169. The PID for the Moora Moora Creek BTRWs is within the recommended values of 1 × 10−2 to 1 × 10−4 for accurate population estimates and individual identification (Mills et al. 2000; Waits et al. 2001). The Hardy–Weinberg equilibrium test showed no significant deviation from equilibrium after Bonferroni correction and no linkage disequilibrium was present for the loci used in the study. The analysis in MICRO-CHECKER revealed no evidence of null alleles being present and no allelic dropout occurring across all the microsatellite loci. The results from the replicated samples showed 100% genotyping match and all three replicates were scored with no failure of PCR. This supports the high-fidelity of the genotyping system employed in this study.
Analysis of faecal DNA from the Moora Moora Creek colony identified eight unique BTRW genotypes (Supplementary Table S4). Of the genotypes identified, two were male and six were female. This increased the total number of BTRWs known to be alive at Moora Moora Creek from five to eight and established a sex ratio in the colony of 1:3 male to female (Supplementary Table S5). The female genotypes were designated F1 (34 scats), F2 (54 scats), F3 (37 scats), F4 (13 scats), F5 (18 scats) and F6 (1 scat). The male genotypes were designated M1 (48 scats) and M2 (9 scats) (Supplementary Table S5).
The approximate range of each individual BTRW along the Moora Moora Creek Escarpment was mapped on the basis of the GPS locations of the scats collected and genotyped (Fig. 3a–h). Although habitat sharing occurred among all individuals, genotyped BTRW females F4, F5 and F6 and male M2 had far more localised escarpment ranges than did females F1, F2, F3 and male M1.

|
Genetic diversity
An assessment of the genetic diversity of the BTRWs at the Moora Moora Creek colony showed that the average number of alleles per locus (Na) was similar in both the Waite (6 loci; 3.3 ± 0.7) and the Moora Moora Creek (6 loci; 3.3 ± 0.3, 12 loci; 3.8 ± 0.4) BTRW colonies (Supplementary Table S6). The same was true for both expected heterozygosity (He) (Waite, 6 loci; 0.598 ± 0.052; Moora Moora Creek, 6 loci; 0.620 ± 0.051, 12 loci; 0.655 ± 0.029) and observed heterozygosity (Ho) (Waite, 6 loci; 0.865 ± 0.095; Moora Moora Creek, 6 loci; 0.813 ± 0.070, 12 loci; 0.854 ± 0.053). Negative Fis values (Waite, 6 loci; −0.476 ± 0.196; Moora Moora Creek, 6 loci; −0.315 ± 0.052, 12 loci; −0.310 ± 0.067) were calculated for both groups.
Parentage analysis
The probability of exclusion with no parents known was calculated as 1.000. The male M1 was assigned as the most likely sire of two BTRWs (F4 and M2) with at least 95% confidence and of another two BTRWs (F5 and F6) with less than 80% confidence (Table 3). The female F2 was assigned as the most likely mother of F4, F5, and M2 (at least 80% confidence) (Table 3). Female F3 was the most likely mother of female F6 (95% confidence; Table 3), while female F1 was designated as neither mother nor offspring.

|
Discussion
The current study used images gathered from non-invasive camera monitoring and genetic monitoring via scats to assess the status of the reintroduced Moora Moora Creek BTRW colony, and the impact of leaving the colony undisturbed (no supplementation or trapping) for 6 years.
Breeding and recruitment
Camera monitoring showed that all tagged females were breeding. The Moora Moora Creek pouch young appeared to have a higher rate of survival to pouch emergence at 83% than the 73% recorded previously for this species by Wynd et al. (2006). During the 6-year (2008–2013) BTRW colony-establishment phase at Moora Moora Creek, while physical animal/colony interference was occurring, a total of seven pouch young was recorded from 6 of 23 released females, two young were recorded out of pouch and no recruitment into the adult population was observed (Taggart et al. 2015). An increase in the number of pouch young produced per female (0.7/year), in pouch emergence (83%), and in survival of young to subadulthood (n = 2) was thus confirmed during 2014–2017 via camera-monitoring.
In this study, BTRW births in the Moora Moora Creek colony were found to peak in late summer and autumn, with pouch emergence occurring in spring (Taggart et al. 2005; Wynd et al. 2006). This contrasts with earlier reports from Wynd et al. (2006) that found that female BTRWs bred in synchrony in autumn. Tight breeding synchrony, such as that described by Wynd et al. (2006), was observed in female BTRWs at Moora Moora Creek only during 2015, with all young emerging from the pouch within a month of each other.
Previous research suggests that one birth per year is common for BTRWs (Taggart et al. 2005; Wynd et al. 2006); however, the Moora Moora Creek females birthed on average 0.7 pouch young per year. Although the body condition of all individuals recorded on camera at Moora Moora Creek appeared very good, the tagged females were of older ages (between 5 and 8 years old in 2019), which might account for the lower breeding rate as age is known to decrease breeding success (Cassinello and Aldos 1996; Tettamanti et al. 2015). Another explanation may be poor mate choice, with potentially only one dominant adult male available to females at this site.
As the Moora Moora Creek BTRW colony is only small, further monitoring of breeding, which is planned for the colony, will provide more robust estimates of reproductive rates, survival of young and recruitment to the adult population.
Genetic monitoring and diversity
The microsatellite markers used in the current study are different from those used in previous genetic studies on BTRWs, and, so, direct comparisons of genetic diversity cannot be made between the Moora Moora Creek colony and previously published works. Both the Moora Moora Creek and the Waite individuals had a higher Ho than He and negative Fis values are typically representitive of an excess of heterozygotes. As the founder animals of the Moora Moora Creek colony and the Waite individuals come from both the southern and the central ESU (David Taggart, pers. comm.), this genetic mixing may account for a heterozygosity excess. However, another likely explanation is that the small sample size and related individuals of both groups is skewing the diversity statistics (e.g. Hale et al. 2012). The mixed source of the colonies (central and southern ESU) may also explain deviations from Hardy–Weinberg equilibrium because of the Wahlund effect.
The genetic diversity of the Moora Moora Creek BTRW colony is highly comparable to the genetic diversity of the captive Waite individuals, regardless of whether it is calculated at 6 or 12 loci. As the colony is very small and includes only two or possibly three generations, continued breeding without intervention will lead to reduced genetic diversity because of the small number of successful founders, which included a single male and four females. This can have serious consequences, including the reduced ability of a population to adapt to change, loss of fitness through inbreeding depression (Willoughby et al. 2019) and the potential to reduce long-term population growth rates, which are crucial to establishing a reintroduced population (Bozzuto et al. 2019).
Only one male was assigned as a father (M1), and, so, is likely to be the dominant male in the colony. This sire and the dams were predominantly assigned with a high confidence (80–95%); however, M1 was assigned as the father of F5 and F6 with a lower confidence (<80%). This low confidence may be due to the suitability of the microsatellite markers used, because the assessment of markers was limited by the small sample size, as well as because M2 is the full or half sibling of all offspring and so shares many alleles with these individuals. Because M1 shares half its alleles with both F5 and F6, and none of the founders was a full sibling, it is highly likely that M1 is also the father of F5.
Three of the four surviving offspring were assigned to a single dam, female (F2). This bias may be random, may be due to scat sampling effort, with other offspring yet to be discovered, or may indicate that F2 is acting as a dominant breeding female within the colony. Observations of dominant female BTRWs supressing the breeding and/or limiting the successful reproduction of subordinate females has previously been reported (Jarman 1991). This uneven genetic founder representation could increase the rate of inbreeding and loss of genetic fitness within this colony and, ultimately, reduce its population viability if left unchecked (Jamieson 2011).
Genetic monitoring and camera-monitoring
As previously reported in BTRWs and other macropod species, both monitoring techniques identified significant habitat sharing among colony members (Jarman 1991; Laws and Goldizen 2003; Hazlitt et al. 2006b; Molyneux et al. 2011).
Monitoring through faecal genetics and remote cameras produced different population estimates in this study. Camera-monitoring showed that four of the original tagged animals (1 male and 3 females) still survived and one untagged BTRW was present. Faecal genetic monitoring identified eight genotypes within the colony. The results of previous comparisons between these techniques in wildlife species have varied from being highly comparable (Rodgers et al. 2014) to detecting different subsets of the same population (Bluff et al. 2011; Velli et al. 2015).
From the parentage analysis, it is likely that M1, F1, F2, and F3 are the tagged animals. However, the corresponding genotype of the untagged animals identified on camera cannot be determined. As a result of the difficulty in distinguishing untagged BTRWs on camera, we could not be certain how many untagged BTRWs were actually recorded. It is possible, therefore, that the two techniques are detecting different subsets of the colony here as well. These results support previous studies suggesting that non-invasive genetic monitoring and camera-monitoring should be used in combination to obtain the most accurate monitoring results (Bluff et al. 2011; Velli et al. 2015). Such systems can also support each other, because scat searches of sites such as Moora Moora Creek could provide guidance on where to place additional cameras for further monitoring, and both techniques offer unique benefits, such as behavioural observations or genetic analyses.
Both monitoring techniques also had limitations. The information gathered from remote cameras was determined by the number and placement of cameras across the colony. A large limitation of non-invasive faecal genotyping was that the genotypes identified is impacted by the location of the scat searches and the impact of terrain on access across the site.
In this study, less than half the number of scats was found of animals identified as offspring of that of older, parent animals, and all scats of offspring were highly localised within the colony area compared with those of older, parent animals. In particular, only nine scats, over a small area, were identified as belonging to male M2, and DNA from one scat was genotyped as belonging to female F6. This may indicate that older BTRWs at Moora Moora Creek are defending their established home ranges, as has been observed in the allied rock-wallaby (Bleistein et al. 1994; Horsup 1994), thus restricting the location and movement of subordinate animals and potentially forcing them into much smaller areas, or areas less accessible for monitoring. As such, it is possible that BTRWs additional to those currently detected may be present at Moora Moora Creek (Bluff et al. 2011), although, because considerable lengths of escarpment within, and adjacent to, the known colony were searched for scats, it is likely that the majority of individuals in the colony has been detected. Considering the rugged terrain used by BTRWs at the study site, the use of non-invasive genetic methods for population estimates appears appropriate and advantageous.
Insights into the reintroduction success of BTRWs at Moora Moora Creek
Taggart et al. (2015) hypothesised that either regular supplementation of small numbers of naïve captive-bred BTRWs into the establishing Moora Moora Creek colony, or a single larger release of these animals, disrupted colony social cohesion and increased the animal visibility and vulnerability to predation, negatively affecting colony establishment. This hypothesis correlates with the results and observations obtained in this, and other, studies (Moehrenschlager and MacDonald 2003; Shier and Swaisgood 2012). In this study, leaving the BTRW colony undisturbed between 2014 and 2019 through non-invasive monitoring correlated with a decline in BTRW predation, increased breeding, and the recruitment of at least four animals into the adult population. It would appear now that the social structure within the colony has settled without continual disturbances, and along with successful fox baiting and the elimination of resident predators, has promoted breeding and recruitment. This encouraging result justifies continual monitoring of this colony because there is still much to be learnt to inform and improve future reintroduction efforts and recovery of this species.
Despite the benefits of non-invasive monitoring observed in this study, management interventions, such as supplementation, are to be expected if the population is established only on a small number of animals. Non-invasive monitoring can provide information on when such intervention is required, with minimal disturbance to the target population while ensuring that issues, such as inbreeding, do not reach a stage where they severely compromise the population.
Management intervention in the Moora Moora Creek colony took place post-study to address the need for increased genetic diversity. Male 167 was replaced with two new unrelated males. This one-off event of adding new genetics into the colony was deemed worth the risk, despite the colony interference. This lower-level physical interference stands in contrast to that which occurred between 2008 and 2013, when the colony was being regularly interfered with associated trapping for monitoring purposes and the frequent addition of new animals across the colony establishment period. However, the following 8 years (2014–2021), which include the current study, have seen minimal interference owing to camera monitoring and scat genotyping, allowing the animals within the colony to settle and establish.
The results of this study should be considered when planning future reintroductions of highly territorial and social species such as the BTRW (Hazlitt et al. 2004), where limited information is available on breeding rates, survival, recruitment, site carrying capacity, and animal home range in open, unfenced, wild populations.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
The authors acknowledge the support of World Wildlife Fund Australia, The FAUNA Research Alliance Inc. and the Schultz Foundation.
Acknowledgements
We thank Parks Victoria staff who assisted with maintenance of cameras and collection of camera images.
References
Akomo-Okoue, EF, Inoue, E, Atteke, C, Nakashima, Y, Hongo, S, Inoue-Murayama, M, and Yamagiwa, J (2015). Noninvasive genetic analysis for assessing the abundance of duiker species among habitats in the tropical forest of Moukalaba, Gabon. Mammal Research 60, 375–384.| Noninvasive genetic analysis for assessing the abundance of duiker species among habitats in the tropical forest of Moukalaba, Gabon.Crossref | GoogleScholarGoogle Scholar |
Aussavy, M, Bernardin, E, Corrigan, A, Hufschmid, J, and Beveridge, I (2011). Helminth parasite communities in four species of sympatric macropodids in western Victoria. Australian Mammalogy 33, 13–20.
| Helminth parasite communities in four species of sympatric macropodids in western Victoria.Crossref | GoogleScholarGoogle Scholar |
Bannister, H, Croxford, A, Brandle, R, Paton, DC, and Moseby, K (2020). Time to adjust: changes in the diet of a reintroduced marsupial after release. Oryx 55, 755–764.
| Time to adjust: changes in the diet of a reintroduced marsupial after release.Crossref | GoogleScholarGoogle Scholar |
Berger-Tal, O, Blumstein, DT, and Swaisgood, RR (2020). Conservation translocations: a review of common difficulties and promising directions. Animal Conservation 23, 121–131.
Bleistein, S, Horsup, A, and Gansloßer, U (1994). Border conflicts among territorial rock wallabies (Petrogale assimilis). Aggressive Behaviour 20, 325–333.
| Border conflicts among territorial rock wallabies (Petrogale assimilis).Crossref | GoogleScholarGoogle Scholar |
Bluff, LA, Clausen, L, Hill, A, and Bramwell, MD (2011). A decade of monitoring the remnant victorian population of the brush-tailed rock-wallaby (Petrogale penicillata). Australian Mammalogy 33, 195–201.
| A decade of monitoring the remnant victorian population of the brush-tailed rock-wallaby (Petrogale penicillata).Crossref | GoogleScholarGoogle Scholar |
Bourgeois, S, Kaden, J, Senn, H, Bunnefeld, N, Jeffery, KJ, Akomo-Okoue, EF, Ogden, R, and McEwing, R (2019). Improving cost-efficiency of faecal genotyping: new tools for elephant species. PLoS ONE 14, e0210811.
| Improving cost-efficiency of faecal genotyping: new tools for elephant species.Crossref | GoogleScholarGoogle Scholar | 30699177PubMed |
Bozzuto, C, Biebach, I, Muff, S, Ives, AR, and Keller, LF (2019). Inbreeding reduces long-term growth of Alpine ibex populations. Nature Ecology & Evolution 3, 1359–1364.
| Inbreeding reduces long-term growth of Alpine ibex populations.Crossref | GoogleScholarGoogle Scholar |
Bramwell M, Andrews L, Brook L, Corrigan T, Stevens M, Schultz D, Taggart D (2008) Reintroduction plan for the release of brush-tailed rock-wallabies (Petrogale penicillata) into Moora Moora Creek, Grampians National Park, Victoria. Department of Sustainability and Environment, Bairnsdale, Vic., Australia.
Browning, TL, Taggart, DA, Rummery, C, Close, RL, and Eldridge, MDB (2001). Multifaceted genetic analysis of the “critically endangered” brush-tailed rock-wallaby Petrogale penicillata in Victoria, Australia: implications for management. Conservation Genetics 2, 145–156.
| Multifaceted genetic analysis of the “critically endangered” brush-tailed rock-wallaby Petrogale penicillata in Victoria, Australia: implications for management.Crossref | GoogleScholarGoogle Scholar |
Bubac, CM, Johnson, AC, Fox, JA, and Cullingham, CI (2019). Conservation translocations and post-release monitoring: identifying trends in failures, biases, and challenges from around the world. Biological Conservation , 108239.
| Conservation translocations and post-release monitoring: identifying trends in failures, biases, and challenges from around the world.Crossref | GoogleScholarGoogle Scholar |
Camp, A, Croxford, AE, Ford, CS, Baumann, U, Clements, PR, Hiendleder, S, Woolford, L, Netzel, G, Boardman, WSJ, Fletcher, MT, and Wilkinson, MJ (2020). Dual-locus DNA metabarcoding reveals southern hairy-nosed wombats (Lasiorhinus latifrons Owen) have a summer diet dominated by toxic invasive plants. PLoS ONE , e0229390.
| Dual-locus DNA metabarcoding reveals southern hairy-nosed wombats (Lasiorhinus latifrons Owen) have a summer diet dominated by toxic invasive plants.Crossref | GoogleScholarGoogle Scholar | 32142513PubMed |
Cassinello, J, and Aldos, CL (1996). Female reproductive success in Ammontragus lervia (Bovidae, Artiodactyla). Study of its components and effects of heriarchy and inbreeding. Journal of Zoology 239, 141–153.
| Female reproductive success in Ammontragus lervia (Bovidae, Artiodactyla). Study of its components and effects of heriarchy and inbreeding.Crossref | GoogleScholarGoogle Scholar |
Claridge, AW, Paull, DJ, and Barry, SC (2010). Detection of medium-sized ground-dwelling mammals using infrared digital cameras: an alternative way forward? Australian Mammalogy 32, 165–171.
| Detection of medium-sized ground-dwelling mammals using infrared digital cameras: an alternative way forward?Crossref | GoogleScholarGoogle Scholar |
Cutler, TL, and Swann, DE (1999). Using remote photography in wildlife ecology: a review. Wildlife Society Bulletin 27, 571–581.
De Bondi, N, White, JG, Stevens, M, and Cooke, R (2010). A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities. Wildlife Research 37, 456–465.
| A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities.Crossref | GoogleScholarGoogle Scholar |
Delaney R, Temple-Smith P, McLean N (2005) Strategy for the translocation of the brush-tailed rock-wallaby in Victoria. Report for the Victorian Brush-tailed Rock-wallaby Recovery Team, Department of Sustainability and Environment and Parks Victoria, Vic., Australia.
Dimsoski, P (2017). Genotyping horse epithelial cells from fecal matter by isolation of polymerase chain reaction products. Croatian Medical Journal 58, 239–249.
| Genotyping horse epithelial cells from fecal matter by isolation of polymerase chain reaction products.Crossref | GoogleScholarGoogle Scholar | 28613041PubMed |
Eldridge, MDB, Rummery, C, Bray, C, Zenger, KR, Browning, T, and Close, RL (2004). Genetic analysis of a population crash in brush-tailed rock-tailed rock-wallabies (Petrogale penicillata) from Jenolan Caves, South-Eastern Australia. Wildlife Research 31, 229–240.
| Genetic analysis of a population crash in brush-tailed rock-tailed rock-wallabies (Petrogale penicillata) from Jenolan Caves, South-Eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Gant, JR, Mair, L, and McGowan, PJK (2020). Fragmented evidence for the contribution of ex situ management to species conservation indicates the need for better reporting. Oryx 55, 573–580.
| Fragmented evidence for the contribution of ex situ management to species conservation indicates the need for better reporting.Crossref | GoogleScholarGoogle Scholar |
Hale, ML, Burg, TM, and Steeves, TE (2012). Sampling for microsatellite-based population genetic studies: 25 to 30 individuals per population is enough to accurately estimate allele frequencies. PLoS ONE 7, e45170.
| Sampling for microsatellite-based population genetic studies: 25 to 30 individuals per population is enough to accurately estimate allele frequencies.Crossref | GoogleScholarGoogle Scholar | 22984627PubMed |
Hardman, B, Moro, D, and Calver, M (2016). Direct evidence implicates feral cat predation as the primary cause of failure of a mammal reintroduction programme. Ecological Management & Restoration 17, 152–158.
| Direct evidence implicates feral cat predation as the primary cause of failure of a mammal reintroduction programme.Crossref | GoogleScholarGoogle Scholar |
Hayward, MW, Bellchambers, K, Herman, K, Bentley, J, and Legge, S (2011). Spatial behaviour of yellow-footed rock-wallabies, Petrogale xanthopus, changes in response to active conservation management. Australian Journal of Zoology 59, 1–8.
| Spatial behaviour of yellow-footed rock-wallabies, Petrogale xanthopus, changes in response to active conservation management.Crossref | GoogleScholarGoogle Scholar |
Hazlitt, SL, Eldridge, MDB, and Goldizen, AW (2004). Fine-scale spatial genetic correlation analyses reveal strong female philopatry within a brush-tailed rock-wallaby colony in southeast Queensland. Molecular Ecology 13, 3621–3632.
| Fine-scale spatial genetic correlation analyses reveal strong female philopatry within a brush-tailed rock-wallaby colony in southeast Queensland.Crossref | GoogleScholarGoogle Scholar | 15548278PubMed |
Hazlitt, SL, Goldizen, AW, and Eldridge, MDB (2006a). Significant patterns of population genetic structure and limited gene flow in a threatened macropodid marsupial despite continuous habitat in Southeast Queensland, Australia. Conservation Genetics 7, 675–689.
| Significant patterns of population genetic structure and limited gene flow in a threatened macropodid marsupial despite continuous habitat in Southeast Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Hazlitt, SL, Goldizen, AW, Nicholls, JA, and Eldridge, MDE (2014). Three divergent lineages within an Australian Marsupial (Petrogale penicillata) suggest multiple major refugia for mesic taxa in Southeast Australia. Ecology and Evolution 4, 1102–1116.
| Three divergent lineages within an Australian Marsupial (Petrogale penicillata) suggest multiple major refugia for mesic taxa in Southeast Australia.Crossref | GoogleScholarGoogle Scholar | 24772286PubMed |
Hazlitt, SL, Sigg, DP, Eldridge, MDB, and Goldizen, AW (2006b). Restricted mating dispersal and strong breeding group structure in a mid-sized marsupial mammal (Petrogale penicillata). Molecular Ecology 15, 2997–3007.
| Restricted mating dispersal and strong breeding group structure in a mid-sized marsupial mammal (Petrogale penicillata).Crossref | GoogleScholarGoogle Scholar | 16911216PubMed |
Horsup, A (1994). Home range of the allied rock-wallaby, Petrogale assimilis. Wildlife Research 21, 65–83.
| Home range of the allied rock-wallaby, Petrogale assimilis.Crossref | GoogleScholarGoogle Scholar |
Hoyle, SD, Horsup, AB, Johnson, CN, Crossman, DG, and McCallum, H (1995). Live-trapping of the northern hairy-nosed wombat (Lasiorhinus krefftii): population-size estimates and effects on individuals. Wildlife Research 22, 741–755.
| Live-trapping of the northern hairy-nosed wombat (Lasiorhinus krefftii): population-size estimates and effects on individuals.Crossref | GoogleScholarGoogle Scholar |
Jamieson, IG (2011). Founder effects, inbreeding, and loss of genetic diversity in four avian reintroduction programs. Conservation Biology 25, 115–123.
| Founder effects, inbreeding, and loss of genetic diversity in four avian reintroduction programs.Crossref | GoogleScholarGoogle Scholar | 20825445PubMed |
Jarman, PJ (1991). Social behaviour and organization in the macropodoidea. Advances in the Study of Behaviour 20, 1–50.
| Social behaviour and organization in the macropodoidea.Crossref | GoogleScholarGoogle Scholar |
Jarman, PJ, and Capararo, SM (1996). Use of rock-wallaby faecal pellets for detecting and monitoring populations and examining habitat use. Australian Mammalogy 19, 257–264.
| Use of rock-wallaby faecal pellets for detecting and monitoring populations and examining habitat use.Crossref | GoogleScholarGoogle Scholar |
Johnson, TF, Brown, TJ, Richardson, DS, and Dugdale, HL (2018). The importance of post–translocation monitoring of habitat use and population growth: insights from a Seychelles warbler (Acrocephalus sechellensis) translocation. Journal of Ornithology 159, 439–446.
| The importance of post–translocation monitoring of habitat use and population growth: insights from a Seychelles warbler (Acrocephalus sechellensis) translocation.Crossref | GoogleScholarGoogle Scholar |
Kalinowski, ST, Taper, ML, and Marshall, TC (2007). Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Molecular Ecology 16, 1099–1106.
| Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment.Crossref | GoogleScholarGoogle Scholar | 17305863PubMed |
Katsura, Y, Kondo, HX, Ryan, J, Harley, V, and Satta, Y (2018). The evolutionary process of mammalian sex determination genes focusing on marsupial SRYs. BMC Evolutionary Biology 18, 3.
| The evolutionary process of mammalian sex determination genes focusing on marsupial SRYs.Crossref | GoogleScholarGoogle Scholar | 29338681PubMed |
Kaufmann, JH (1974). Habitat use and social organization of nine sympatric species of macropodid marsupials. Journal of Mammalogy 55, 66–80.
| Habitat use and social organization of nine sympatric species of macropodid marsupials.Crossref | GoogleScholarGoogle Scholar |
Kolodzieg, K, Nikolov, I, Schulz, HK, Theissinger, K, and Schulz, R (2013). Evaluation of fecal storage and DNA extraction methods in wild boar (Sus scrofa). Biochemical Genetics 51, 406–412.
| Evaluation of fecal storage and DNA extraction methods in wild boar (Sus scrofa).Crossref | GoogleScholarGoogle Scholar |
Koressaar, T, Lepamets, M, Kaplinski, L, Raime, K, Andreson, R, and Remm, M (2018). Primer3_masker: integrating masking of template sequence with primer design software. Bioinformatics 34, 1937–1938.
| Primer3_masker: integrating masking of template sequence with primer design software.Crossref | GoogleScholarGoogle Scholar | 29360956PubMed |
Koressaar, T, and Remm, M (2007). Enhancements and modifications of primer design program Primer3. Bioinformatics 23, 1289–1291.
| Enhancements and modifications of primer design program Primer3.Crossref | GoogleScholarGoogle Scholar | 17379693PubMed |
Lachish, S, Passmore, A, and Jones, M (2011). A new PCR assay for reliable molecular sexing of endangered tasmanian devils (Sarcophilus harrisii) from non-invasive genetic samples. Conservation Genetics Resources 3, 279–281.
| A new PCR assay for reliable molecular sexing of endangered tasmanian devils (Sarcophilus harrisii) from non-invasive genetic samples.Crossref | GoogleScholarGoogle Scholar |
Lapbenjakul, S, Thapana, W, Twilprawat, P, Muangmai, N, Kanchanaketu, T, Temsiripong, Y, Unajak, S, Peyachoknagul, S, and Srikulnath, K (2017). High genetic diversity and demographic history of captive Siamese and saltwater crocodiles suggest the first step toward the establishment of a breeding and reintroduction program in Thailand. PLoS ONE 12, e0184526.
| High genetic diversity and demographic history of captive Siamese and saltwater crocodiles suggest the first step toward the establishment of a breeding and reintroduction program in Thailand.Crossref | GoogleScholarGoogle Scholar | 28953895PubMed |
Laws, RJ, and Goldizen, AW (2003). Nocturnal home ranges and social interactions of the brushtailed rock-wallaby Petrogale penicillata at Hurdle Creek, Queensland. Australian Mammalogy 25, 169–176.
| Nocturnal home ranges and social interactions of the brushtailed rock-wallaby Petrogale penicillata at Hurdle Creek, Queensland.Crossref | GoogleScholarGoogle Scholar |
Luikart, G, Painter, J, Crozier, RH, Westerman, M, and Sherwin, WB (1997). Characterization of microsatellite loci in the endangered long-footed potoroo Potorous longipes. Molecular Ecology 6, 497–498.
| Characterization of microsatellite loci in the endangered long-footed potoroo Potorous longipes.Crossref | GoogleScholarGoogle Scholar | 9161018PubMed |
Mills, LS, Citta, JJ, Lair, KP, Schwartz, MK, and Tallmon, DA (2000). Estimating animal abundance using noninvasive DNA sampling: promise and pitfalls. Ecological Applications 10, 283–294.
| Estimating animal abundance using noninvasive DNA sampling: promise and pitfalls.Crossref | GoogleScholarGoogle Scholar |
Moehrenschlager, A, and MacDonald, DW (2003). Movement and survival parameters of translocated and resident swift foxes Vulpes velox. Animal Conservation 6, 199–206.
| Movement and survival parameters of translocated and resident swift foxes Vulpes velox.Crossref | GoogleScholarGoogle Scholar |
Molyneux, J, Taggart, DA, Corrigan, A, and Frey, S (2011). Home-range studies in a reintroduced brush-tailed rock-wallaby (Petrogale penicillata) population in the Grampians National Park, Victoria. Australian Mammalogy 33, 128–134.
| Home-range studies in a reintroduced brush-tailed rock-wallaby (Petrogale penicillata) population in the Grampians National Park, Victoria.Crossref | GoogleScholarGoogle Scholar |
Morales-Contreras, J, Escalante, P, Martínez-Ruíz, M, and Matías-Ferrer, N (2016). Black hawk-eagle (Spizaetus tyrannus) idenitfied by DNA from a feather recovered in the rain-forest region of Veracruz. The Southwestern Naturalist 61, 137–142.
| Black hawk-eagle (Spizaetus tyrannus) idenitfied by DNA from a feather recovered in the rain-forest region of Veracruz.Crossref | GoogleScholarGoogle Scholar |
Moseby, KE, Read, JL, Paton, DC, Copley, P, Hill, BM, and Crisp, HA (2011). Predation determines the outcome of 10 reintroduction attempts in arid South Australia. Biological Conservation 144, 2863–2872.
| Predation determines the outcome of 10 reintroduction attempts in arid South Australia.Crossref | GoogleScholarGoogle Scholar |
Moßbrucker, AM, Apriyana, I, Fickel, J, Imron, MA, Pudyatmoko, S, and Sumardi, SH (2015). Non-invasive genotyping of sumatran elephants: implications for conservation. Tropical Conservation Science 8, 745–759.
| Non-invasive genotyping of sumatran elephants: implications for conservation.Crossref | GoogleScholarGoogle Scholar |
Paplinska, JZ, Taggart, DA, Corrigan, T, Eldridge, MDB, and Austin, JJ (2011). Using DNA from museum specimens to preserve the integrity of evolutionarily significant unit boundaries in threatened species. Biological Conservation 144, 290–297.
| Using DNA from museum specimens to preserve the integrity of evolutionarily significant unit boundaries in threatened species.Crossref | GoogleScholarGoogle Scholar |
Parlato, EH, and Armstrong, DP (2013). Predicting post-release establishment using data from multiple reintroductions. Biological Conservation 160, 97–104.
| Predicting post-release establishment using data from multiple reintroductions.Crossref | GoogleScholarGoogle Scholar |
Peakall, R, and Smouse, PE (2006). GenAlEx 6: genetic analysis in Excel. Population genetic software for teaching and research: an update. Molecular Ecology Notes 6, 288–295.
| GenAlEx 6: genetic analysis in Excel. Population genetic software for teaching and research: an update.Crossref | GoogleScholarGoogle Scholar |
Peakall, R, and Smouse, PE (2012). GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research: an update. Bioinformatics 28, 2537–2539.
| GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research: an update.Crossref | GoogleScholarGoogle Scholar | 22820204PubMed |
Piggott, MP (2004). Effect of sample age and season of collection on the reliability of microsatellite genotyping of faecal DNA. Wildlife Research 31, 485–493.
| Effect of sample age and season of collection on the reliability of microsatellite genotyping of faecal DNA.Crossref | GoogleScholarGoogle Scholar |
Piggott, MP, Banks, SC, Stone, N, Banffy, C, and Taylor, AC (2006a). Estimating population size of endangered brush-tailed rock-wallaby (Petrogale penicillata) colonies using faecal DNA. Molecular Ecology 15, 81–91.
| Estimating population size of endangered brush-tailed rock-wallaby (Petrogale penicillata) colonies using faecal DNA.Crossref | GoogleScholarGoogle Scholar | 16367832PubMed |
Piggott, MP, Banks, SC, and Taylor, AC (2006b). Population structure of brush-tailed rock-wallaby (Petrogale penicillata) colonies inferred from analysis of faecal DNA. Molecular Ecology 15, 93–105.
| Population structure of brush-tailed rock-wallaby (Petrogale penicillata) colonies inferred from analysis of faecal DNA.Crossref | GoogleScholarGoogle Scholar | 16367833PubMed |
Piggott, MP, Hansen, B, Soderquist, T, Eldridge, MDB, and Taylor, AC (2018). Population monitoring of small and declining brush-tailed rock-wallaby (Petrogale penicillata) colonies at the extreme of their range using faecal DNA sampling. Australian Mammalogy 40, 58–66.
| Population monitoring of small and declining brush-tailed rock-wallaby (Petrogale penicillata) colonies at the extreme of their range using faecal DNA sampling.Crossref | GoogleScholarGoogle Scholar |
Piggott, MP, and Taylor, AC (2003). Remote collection of animal DNA and its applications in conservation management and understanding the population biology of rare and cryptic species. Wildlife Research 30, 1–13.
| Remote collection of animal DNA and its applications in conservation management and understanding the population biology of rare and cryptic species.Crossref | GoogleScholarGoogle Scholar |
Ramón-Laca, A, Soriano, L, Gleeson, D, and Godoy, JA (2015). A simple and effective method for obtaining mammal DNA from faeces. Wildlife Biology 21, 195–203.
| A simple and effective method for obtaining mammal DNA from faeces.Crossref | GoogleScholarGoogle Scholar |
Raymond, M, and Rousset, F (1995). An exact test for population differentiation. Evolution , 1280–1283.
| An exact test for population differentiation.Crossref | GoogleScholarGoogle Scholar | 28568523PubMed |
Richards, JD, and Short, J (2003). Reintroduction and establishment of the western barred bandicoot Perameles bougainville (Marsupialia: Peramelidae) at Shark Bay, Western Australia. Biological Conservation 109, 181–195.
| Reintroduction and establishment of the western barred bandicoot Perameles bougainville (Marsupialia: Peramelidae) at Shark Bay, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Robinson, NM, Dexter, N, Brewster, R, Maple, D, MacGregor, C, Rose, C, Hall, J, and Lindenmayer, DB (2020). Be nimble with threat mitigation: lessons learned from the reintroduction of an endangered species. Restoration Ecology 28, 29–38.
| Be nimble with threat mitigation: lessons learned from the reintroduction of an endangered species.Crossref | GoogleScholarGoogle Scholar |
Robley A, Ramsey D, Holmes B (2014) Grampians arc fox monitoring research partnership project 2013–2014. Arthur Rylah Institute for Environmental Research, unpublished client report for Parks Victoria, Department of Environment and Primary Industries, Melbourne, Vic., Australia.
Rodgers, TW, Giacalone, J, Heske, EJ, Janecka, JE, Phillips, CA, and Schooley, RL (2014). Comparison of noninvasive genetics and camera trapping for estimating population density of ocelots (Leopardus pardalis) on Barro Colorado Island, Panama. Tropical Conservation Science 7, 690–705.
| Comparison of noninvasive genetics and camera trapping for estimating population density of ocelots (Leopardus pardalis) on Barro Colorado Island, Panama.Crossref | GoogleScholarGoogle Scholar |
Rousset, F (2008). Genepop’007: a complete re-implementation of the genepop software for Windows and Linux. Molecular Ecology Resources 8, 103–106.
| Genepop’007: a complete re-implementation of the genepop software for Windows and Linux.Crossref | GoogleScholarGoogle Scholar | 21585727PubMed |
Sabino-Marques, H, Ferreira, CM, Paupério, J, Costa, P, Barbosa, S, Encarnação, C, Alpizar-Jara, R, Alves, PC, Searle, JB, Mira, A, Beja, P, and Pita, R (2018). Combining genetic non-invasive sampling with spatially explicit capture–recapture models for density estimation of a patchily distributed small mammal. European Journal of Wildlife Research 64, 44.
| Combining genetic non-invasive sampling with spatially explicit capture–recapture models for density estimation of a patchily distributed small mammal.Crossref | GoogleScholarGoogle Scholar |
Scott, PA, Allison, LJ, Field, KJ, Averill-Murray, RC, and Shaffer, HB (2020). Individual heterozygosity predicts translocation success in threatened desert tortoises. Science 370, 1086–1089.
| Individual heterozygosity predicts translocation success in threatened desert tortoises.Crossref | GoogleScholarGoogle Scholar | 33243888PubMed |
Sharp, A (2009). Home range dynamics of the yellow-footed rock-wallaby (Petrogale xanthopus celeris) in central-western Queensland. Austral Ecology 34, 55–68.
| Home range dynamics of the yellow-footed rock-wallaby (Petrogale xanthopus celeris) in central-western Queensland.Crossref | GoogleScholarGoogle Scholar |
Sheean, VA, Manning, AD, and Lindenmayer, DB (2012). An assessment of scientific approaches towards species relocations in Australia. Austral Ecology 37, 204–215.
| An assessment of scientific approaches towards species relocations in Australia.Crossref | GoogleScholarGoogle Scholar |
Shier, DM, and Swaisgood, RR (2012). Fitness costs of neighbourhood disruption in translocations of a solitary mammal. Conservation Biology 26, 116–123.
| Fitness costs of neighbourhood disruption in translocations of a solitary mammal.Crossref | GoogleScholarGoogle Scholar | 21978094PubMed |
Short, J (2016). Predation by feral cats key to the failure of a long-term reintroduction of the western barred bandicoot (Perameles bougainville). Wildlife Research 43, 38–50.
| Predation by feral cats key to the failure of a long-term reintroduction of the western barred bandicoot (Perameles bougainville).Crossref | GoogleScholarGoogle Scholar |
Silveira, L, Jácomo, ATA, and Diniz-Filho, JAF (2003). Camera trap, line transect census and track surveys: a comparative evaluation. Biological Conservation 114, 351–355.
| Camera trap, line transect census and track surveys: a comparative evaluation.Crossref | GoogleScholarGoogle Scholar |
Taggart, DA, Schultz, D, White, C, Whitehead, P, Underwood, G, and Phillips, K (2005). Cross-fostering, growth and reproductive studies in the brush-tailed rock-wallaby, Petrogale penicillata (Marsupialia: Macropodidae): efforts to accelerate breeding in a threatened marsupial species. Australian Journal of Zoology 53, 313–323.
| Cross-fostering, growth and reproductive studies in the brush-tailed rock-wallaby, Petrogale penicillata (Marsupialia: Macropodidae): efforts to accelerate breeding in a threatened marsupial species.Crossref | GoogleScholarGoogle Scholar |
Taggart, DA, Schultz, DJ, Corrigan, TJ, Schultz, TJ, Stevens, M, Panther, D, and White, CR (2015). Reintroduction methods and a review of mortality in the brush-tailed rock-wallaby, Grampians National Park, Australia. Australian Journal of Zoology 63, 383–397.
| Reintroduction methods and a review of mortality in the brush-tailed rock-wallaby, Grampians National Park, Australia.Crossref | GoogleScholarGoogle Scholar |
Taggart D, Clausen L, Stevens M, Bramwell M, Corrigan A (2008) Brush-tailed rock-wallaby reintroduction site assessment guide for the Grampians National Park, Victoria. Report for the Victorian Brush-tailed Rock Wallaby Recovery Team, Victorian Department of Sustainability and Environment, Melbounrne, Vic., Australia.
Telfer, WR, and Griffiths, AD (2006). Dry-season use of space, habitats and shelters by the short-eared rock-wallaby (Petrogale brachyotis) in the monsoon tropics. Wildlife Research 33, 207–214.
| Dry-season use of space, habitats and shelters by the short-eared rock-wallaby (Petrogale brachyotis) in the monsoon tropics.Crossref | GoogleScholarGoogle Scholar |
Tende, T, Hansson, B, Ottosson, U, Åkesson, M, and Bensch, S (2014). Individual identification and genetic variation of lions (Panthera leo) from two protected areas in Nigeria. PLoS ONE 9, e84288.
| Individual identification and genetic variation of lions (Panthera leo) from two protected areas in Nigeria.Crossref | GoogleScholarGoogle Scholar | 24427283PubMed |
Tettamanti, F, Grignolio, S, Filli, F, Apollonio, M, and Bize, P (2015). Senescence in breeding success of female Alpine chamois (Rupicapra rupicapra): the role of female quality and age. Oecologia 178, 187–195.
| Senescence in breeding success of female Alpine chamois (Rupicapra rupicapra): the role of female quality and age.Crossref | GoogleScholarGoogle Scholar | 25556530PubMed |
Untergasser, A, Cutcutache, I, Koressaar, T, Ye, J, Faircloth, BC, Remm, M, and Rozen, SG (2012). Primer3: new capabilities and interfaces. Nucleic Acids Research 40, e115.
| Primer3: new capabilities and interfaces.Crossref | GoogleScholarGoogle Scholar | 22730293PubMed |
Van Oosterhout, C, Hutchinson, WF, Wills, DP, and Shipley, P (2004). MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes 4, 535–538.
| MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data.Crossref | GoogleScholarGoogle Scholar |
Velli, E, Bologna, MA, Silvia, C, Ragni, B, and Randi, E (2015). Non-invasive monitoring of the european wildcat (Felis silvestris silvestris Schreber, 1777): comparative analysis of three different monitoring techniques and evaluation of their integration. European Journal of Wildlife Research 61, 657–668.
| Non-invasive monitoring of the european wildcat (Felis silvestris silvestris Schreber, 1777): comparative analysis of three different monitoring techniques and evaluation of their integration.Crossref | GoogleScholarGoogle Scholar |
Waits, LP, Luikart, G, and Taberlet, P (2001). Estimating the probability of identity among genotypes in natural populations: cautions and guidelines. Molecular Ecology 10, 249–256.
| Estimating the probability of identity among genotypes in natural populations: cautions and guidelines.Crossref | GoogleScholarGoogle Scholar | 11251803PubMed |
Wakefield, N (1961). Victoria’s rock-wallabies. Victorian Naturalist 77, 322–332.
Walker, FM, Horsup, A, and Taylor, AC (2009). Leader of the pack: faecal pellet deposition order impacts PCR amplification in wombats. Molecular Ecology Resources 9, 720–724.
| Leader of the pack: faecal pellet deposition order impacts PCR amplification in wombats.Crossref | GoogleScholarGoogle Scholar | 21564730PubMed |
Watkins, GE, Willers, N, Raudino, H, Kinloch, J, and van Dongen, R (2018). Success criteria not met, but valuable information gained: monitoring a reintroduction of the tammar wallaby. Wildlife Research 45, 421–435.
| Success criteria not met, but valuable information gained: monitoring a reintroduction of the tammar wallaby.Crossref | GoogleScholarGoogle Scholar |
Watson, CM, Margan, SH, and Johnston, PG (1998). Sex-chromosome elimination in the bandicoot Isoodon macrourus using Y-linked markers. Cytogenetics and Cell Genetics 81, 54–59.
| Sex-chromosome elimination in the bandicoot Isoodon macrourus using Y-linked markers.Crossref | GoogleScholarGoogle Scholar | 9691176PubMed |
Wegge, P, Pokheral, CP, and Jnawali, SR (2004). Effects of trapping effort and trap shyness on estimates of tiger abundance from camera trap studies. Animal Conservation 7, 251–256.
| Effects of trapping effort and trap shyness on estimates of tiger abundance from camera trap studies.Crossref | GoogleScholarGoogle Scholar |
White, PCL, McClean, CJ, and Woodroffe, GL (2003). Factors affecting the success of an otter (Lutra lutra) reinforcement programme, as identified by post-translocation monitoring. Biological Conservation 112, 363–371.
| Factors affecting the success of an otter (Lutra lutra) reinforcement programme, as identified by post-translocation monitoring.Crossref | GoogleScholarGoogle Scholar |
Wilkinson, MJ, Szabo, C, Ford, CS, Yarom, Y, Croxford, AE, Camp, A, and Gooding, P (2017). Replacing Sanger with next generation sequencing to improve coverage and quality of reference DNA barcodes for plants. Scientific Reports 7, 46040.
| Replacing Sanger with next generation sequencing to improve coverage and quality of reference DNA barcodes for plants.Crossref | GoogleScholarGoogle Scholar | 28401958PubMed |
Willoughby, JR, Waser, PM, Brüniche-Olsen, A, and Christie, MR (2019). Inbreeding load and inbreeding depression estimated from lifetime reproductive success in a small, dispersal-limited population. Heredity 123, 192–201.
| Inbreeding load and inbreeding depression estimated from lifetime reproductive success in a small, dispersal-limited population.Crossref | GoogleScholarGoogle Scholar | 30809076PubMed |
Woinarski JCZ, Burbidge AA, Harrison PL (2014) Taxon profiles. In ‘The Action Plan for Australian Mammals 2012’. pp. 51–857. (CSIRO Publishing: Melbourne, Vic., Australia)
Wynd, NM, Sigg, DP, Pople, AR, Hazlitt, SL, and Goldizen, AW (2006). Factors affecting female reproductive success and the survival of pouch young in the threatened brush-tailed rock-wallaby, Petrogale penicillata. Australian Journal of Zoology 54, 61–70.
| Factors affecting female reproductive success and the survival of pouch young in the threatened brush-tailed rock-wallaby, Petrogale penicillata.Crossref | GoogleScholarGoogle Scholar |
Zeng, Y, Jiang, Z, and Li, C (2007). Genetic variability in relocated Père David’s deer (Elaphurus davidianus) populations: iImplications to reintroduction program. Conservation Genetics 8, 1051–1059.
| Genetic variability in relocated Père David’s deer (Elaphurus davidianus) populations: iImplications to reintroduction program.Crossref | GoogleScholarGoogle Scholar |