

Nest remains are insufficient to identify predators of waterfowl nests
Kaylan M. Kemink A * , Kyle J. Kuechle A , Mason L. Sieges A , Sam Krohn A B , Cailey D. Isaacson A B , John Palarski A B , Nick Conrad A B , Allicyn Nelson A B , Boyan Liu A , Thomas K. Buhl C and Susan N. Ellis-Felege B
A * , Kyle J. Kuechle A , Mason L. Sieges A , Sam Krohn A B , Cailey D. Isaacson A B , John Palarski A B , Nick Conrad A B , Allicyn Nelson A B , Boyan Liu A , Thomas K. Buhl C and Susan N. Ellis-Felege B
A Great Plains Regional Office, Ducks Unlimited Inc., 2525 River Road, Bismarck, ND, USA.
B Department of Biology, University of North Dakota, 10 Cornell Street, Grand Forks, ND, USA.
C U.S. Geological Survey, Northern Prairie Wildlife Research Center, Jamestown, ND, USA.
Wildlife Research 50(3) 182-189 https://doi.org/10.1071/WR22042
Submitted: 4 March 2022 Accepted: 23 July 2022 Published: 19 September 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution 4.0 International License (CC BY)
Abstract
Context: Nest predation is a leading cause of nest failure for most ground-nesting birds. Methods that allow for accurate classification of fate and identification of predators are important for understanding productivity and conservation strategies. Past studies have used a visual inspection of nest remains to determine nest fate and predict predator identity. Most formal assessments of these methods have addressed small-bodied birds nesting in trees or shrubs, and have revealed that use of evidence at nests can be relatively accurate for determining nest fate but may lead to incorrect conclusions regarding predator identity. However, few have tested the latter hypothesis for larger ground-nesting birds with precocial young.
Aim: We aimed to evaluate a classification system developed for determining nest fate and identifying predators of waterfowl nests, at both the scale of individual nests and across the study area.
Methods: From 2016 to 2020, we located 989 blue-winged teal (Spatula discors), mallard (Anas platyrhynchos) and gadwall (Mareca strepera) nests in central North Dakota. We placed cameras at a subset of 249 nests and recorded evidence of nest remains at depredated nests.
Key results: The most common predators were American badgers (Taxidea taxus), followed by striped skunks (Mephitis mephitis), raccoons (Procyon lotor) and red foxes (Vulpes vulpes). Using evidence of nest remains, we determined nest fates with high accuracy (98.0%). However, evidence of nest remains was only sufficient for identifying predators at 50% of nests, and the classification system was correct only 69.7% of the time. The predicted proportion of predators across the study area differed between the classification system and our video evidence as well.
Conclusions: The accuracy of predator identifications based upon the classification system that we evaluated was not supported at any scale.
Implications: Our results suggest that evidence of nest remains can be used to determine nest fate for large-bodied precocial, ground-nesting birds, but accurate identification of nest predators will require alternative methods such as nest cameras.
Keywords: badger, conservation, grassland, nest camera, Prairie Pothole Region, predation, waterfowl, wetland.
Introduction
Nest cameras revolutionised the study of nesting behaviour and success in birds (Thompson et al. 1999; Pietz and Granfors 2000; Ellis-Felege and Carroll 2012; Ribic et al. 2012), allowing more accurate descriptions of nesting phenology (Burnam 2008), success (Ellis et al. 2018), and causes of failure (Burr et al. 2017). However, the cost (both financial and temporal) of using nest cameras may be prohibitive for many investigators (Cox et al. 2012; Prinz et al. 2016) and may also bias nest fate (Ellis-Felege and Carroll 2012; Ribic et al. 2012). Thus, alternative methods for assessing the causes of nest failures could be useful, especially for studies focusing on identifying nest predators and for mitigating factors influencing the risk of predation (Weidinger 2008).
One possible way to identify nest predators is to use physical signs left behind (Williams and Wood 2002; Anthony et al. 2004; Staller et al. 2005). Methods for categorising sign at nests for ground-nesting birds like waterfowl were based on extensive research of predators (Sargeant et al. 1998), but assignment of fates at other nests was less empirical and based largely on assumptions (Pietz et al. 2012). As a result, use of physical sign to identify predators at failed nests has been criticised, with noted weaknesses such as the potential for secondary predation, parental activity or disturbance of nest remains, and observer experience [reviewed by Lariviere (1999); also see Staller et al. 2005; Andes et al. 2019].
Despite critiques of more traditional predator identification techniques, assessments of their efficacy are sparse and have focused primarily on small birds with altricial young such as passerines (Thompson et al. 1999; Pietz and Granfors 2000; Williams and Wood 2002). Some investigators concluded after their assessment that traditional methods were sufficient for discerning nest success from failure, but not for identifying predators (Williams and Wood 2002; Andes et al. 2019). However, others have recommended against using physical sign for any level of nest fate determination. For example, Thompson et al. (1999) identified predation events at apparently undisturbed nests, so advised against using physical sign at nests to identify predators. Similarly, Pietz and Granfors (2000) concluded that evidence at nests was an unreliable method for identifying predators of passerine nests because the same predators left sign at both successful and unsuccessful nests.
Although deemed unreliable for passerines and recommended against for predator determination in all bird species, physical sign is still used due to logistical and financial benefits over predator identification devices like cameras (Lariviere 1999). Further, the use of physical sign to determine nest fates and identify nest predators has not been tested for accuracy for larger ground-nesting birds with precocial young, where nest remains might be easier to detect due to their larger egg and clutch sizes. Sargeant et al. (1998) developed a hierarchical process of classifying physical sign at depredated waterfowl nests that could potentially be used to identify nest predators at the landscape scale. Lariviere (1999) subsequently published a commentary that included a list of factors that could preclude the accurate identification of nest predators using Sargeant et al.’s (1998) method, including overlapping predator behaviours, multi-predator visits, and parental activity at failed nests. However, Lariviere (1999) did not conduct any empirical assessment of the method, and the Sargeant et al. (1998) classification system is still referenced by some investigators studying waterfowl nest success and predation (e.g. Opermanis et al. 2001; Sherfy et al. 2018; Blythe and Boyce 2020).
We tested the accuracy of the Sargeant et al. (1998) hierarchical classification system using evidence collected from video-monitored waterfowl nests. We focused our study on three species of ducks, including mallards (Anas platyrhynchos), blue-winged teal (Spatula discors), and gadwall (Mareca strepera). Using data from successful and failed nests, our objectives were to determine if the hierarchical system accurately identified nest fate and predators at the nest and study area scales. Here, we define study area as the plots specifically searched for nests during our 5-year monitoring period.
Methods
Study area
We located and monitored mallard, blue-winged teal, and gadwall nests at two field sites in the Prairie Pothole Region of North Dakota, USA, from 2016 to 2020. Ducks Unlimited’s Coteau Ranch (1214 ha), and The Nature Conservancy’s Davis Ranch (2832 ha) are adjacent to each other (47.383336°N, 100.278731°W) near Denoff in Sheridan County, North Dakota (Fig. 1). Both sites are composed of mixed grass prairie with numerous wetlands. Rotational cattle grazing occurs at both study areas, and the latter site incorporates prescribed burning in its management. We selected four plots for nest searching on each study site. Individual plots were ∼65 ha in size. Because the study sites were being actively managed, we selected plots that would facilitate continuity in our nest searching from year to year (Fig. 1).

|
Nest monitoring
We searched for nests between 0800 and 1400 hours using the standard chain drag technique (Klett et al. 1986). Plots were searched at least every other week, and nesting females were flushed with a 35-m long chain advancing over the vegetation between two all-terrain vehicles at a speed of 6–11 kph. Once found, we recorded the species of female flushed, number of eggs, vegetation heights or visual obstruction readings (VOR) at all four cardinal directions using a Robel pole (Robel et al. 1970), and clutch age using the candling technique (Weller 1956). We estimated egg hatch dates using the clutch age dates determined by candling eggs and rechecked nests at least every other week until eggs hatched or nests failed. At every subsequent nest check, we recorded the number of eggs, stage of incubation, and state of the nest (normal, missing eggs, partially destroyed, or completely destroyed).
At a subset of nests, we installed continuously recording video surveillance cameras (Model YCC-520TW-24DS; Jet Security USA, Buena Park, CA, USA, and Advanced Security, Bellevue, IL, USA). We deployed cameras at nests where females were incubating eggs, focusing on blue-winged teal in 2016 and 2020, mallards in 2017 and 2018, and gadwall in 2019 and 2020. Cameras recorded colour during the day, black and white at night, and were equipped with 24 light-emitting diodes (LEDs; 950 nm). We attached cameras to a 1.27-m stake placed 0.25–0.5 m from nests and connected to a 12-V, 35-amp battery along with a camouflaged camera box (SEAHORSE Protective Equipment Cases, Fuertes Cases, La Mesa, CA, USA) by a 25-m cable. Inside the camera boxes were a digital video recorder (DVR) which recorded video onto a 32-GB SD card and a remote control for the DVR. We used a tote vision (i.e. small LCD monitor) to view nests during camera setup to be sure the camera was functioning, recording had started and the nest was centred in the field of view. Batteries and camera boxes were placed ∼25 m from nests to minimise possible effects on females and predators caused by equipment, battery, and SD card changes. This placement also allowed us to check nests more frequently via tote vision and exchange batteries and SD cards without disturbing females. We removed cameras after eggs hatched or nests failed. We then deployed cameras at other nests if available.
Field evaluation of nest fate
Waterfowl nests with hatched eggs have eggshell membranes separated from shells. Nest failure is assumed when evidence of these membranes is absent in the nest bowl (Girard 1939; Klett et al. 1986). When this occurred, observers filled out a predation card (Sargeant et al. 1998), recording information within a 3-m radius around the nest about the following variables:
-
The amount (percentage categories of: 0%, <1% (trace), 1–5%, 6–10%, 11–25%, 26–50%, 51–75%, and 76–100%) and displacement of nest materials >5 cm from the edge of nest bowl;
-
Number of cached or buried eggs, size and number of dug areas (holes or areas where soil has been disturbed and may have been piled or refilled with soil and/or debris);
-
Number and location of whole eggs;
-
Number of shell fragments (individual or connected pieces of ≤1/2 egg);
-
Size (small hole = >3/4 of shape intact, large hole = >1/2–3/4 of shape intact, fractured = connected pieces of >1/2 egg but only 1/4–1/2 shape intact, trampled = connected pieces of >1/2 egg on flattened membrane and <1/4 shape intact, and crushed = >1/2 egg wadded in sphere and <1/4 shape intact) and position (side, end, or side and end) of holes on egg remains;
-
Signs of remains of either ducklings or adult females (Fig. 2).

|
We completed predation cards at all suspected depredated nests to facilitate comparison between efficacy of camera and a visual inspection of nest remains in identifying nest predators.
Field-evidence-based predator identification
We assigned predator identities to nests when characteristics of a predation event matched species-specific criteria identified by Sargeant et al. (1998). The original program developed by Sargeant et al. (1998) for determining predators from nest remains was written in SAS code. We rewrote this code in program R. According to Sargeant et al. (1998), a single nest might have signs consistent with multiple predator species, with similar or varying levels of confidence. Thus, this code assigns a predator to a nest ‘with confidence’ if all criteria are met for one species. However, if some but not all criteria are met, the predator is identified as ‘possible’, and this could result in more than one predator being ‘assigned’ to a nest. To address this potential issue, when a predator was assigned ‘with confidence’ to a nest, we removed any ‘possible’ predators from that same nest. When the Sargeant et al. (1998) program assigned a predator as ‘possible’ to a nest, and that nest had no ‘with confidence’ predator assigned, the possible predator was retained as the ‘field-evidence’ predator. In the rare instance (N = 2) that multiple possible predators were assigned, and no predator was determined with confidence, or two predators were assigned with confidence, we removed that nest from the dataset. For example, if a nest was assigned to an American badger (Taxidea taxus; hereafter badger) with confidence and raccoon (Procyon lotor) as possible, we only considered the badger as the nest predator. In contrast, if a nest had no ‘with confidence’ assignment, but raccoon and striped skunk (Mephitis mephitis; hereafter skunk) were each assigned as ‘possible’, we labelled the nest as unknown in the analysis.
Video-evidence-based predator identification
Across all years, when we assumed predation or hatch from nest remains, we corroborated the event with video evidence. We then calculated the proportion of nests that had been correctly and incorrectly identified using the nest remains (Sargeant et al. 1998). We reviewed video recordings after nests failed or eggs hatched from the time cameras were installed to the time they were removed. While watching videos, we recorded nest fate, predator (if applicable), and number of eggs that hatched (if applicable).
Evaluation of field-based predator identification accuracy
The Sargeant et al. (1998) classification system was developed for applications in proportion of predators across the landscape rather than at specific nests. Therefore, we used a Cochran–Mantel–Haenszel (CMH) chi-squared test of independence to compare the proportion of destroyed nests assigned to each predator at the study area scale between predator identification methods, while accounting for sampling across years. Further, for camera-monitored nests and the program based on evidence of nest remains for all nests (which included nests without cameras), we tested for differences within each year using chi-squared tests. We considered P-values ≤0.05 to be significant.
Animal ethics
This research was conducted in compliance with the Guidelines to the Use of Wild Birds in Research. All nest searching and monitoring followed approved methods from the University of North Dakota’s Institutional Animal Care and Use Committee (#A3917-01, Protocols: 1505-10 and 1801-3C), permits from the North Dakota Game and Fish (2016: GNF7701, 2017: GNF8225, 2018: GNF04611027, 2019: GNF04912726, 2020: GNF05182785), and annual approval from The Nature Conservancy.
Results
Evidence of nest predators from nest remains
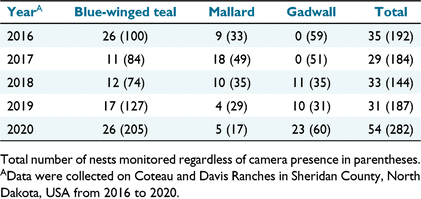
From 2016 to 2020, we located 989 nests, including 590 blue-winged teal nests, 236 gadwall nests, and 163 mallard nests. Of the 989 nests, eggs hatched in 163 (16.5%). We monitored 752 depredated nests and identified predators at 389, based on Sargeant et al. (1998). Criteria assigned 247 predators to nests ‘with confidence’ and assigned ‘possible’ predators to 142 nests. Of the 389 predators identified, the Sargeant et al. (1998) criteria predicted that at least 50% were badgers during each year of our study (range = 50.0% [2020]−81.5% [2017]), with skunks being the second-most common predator (range = 12.7% [2016]−35.0% [2020]). Red foxes (Vulpes vulpes; hereafter fox) and raccoons were classified as the third and fourth most abundant predators, respectively (Fig. 3).

|
Video evidence of nest predators
We deployed cameras at 249 nests. Of camera-monitored nests, we removed cameras at 67 before determining nest fate due to either camera failure or other circumstances (e.g. cattle grazing). For camera nests where we determined fate (N = 182; Table 1), eggs successfully hatched in 45 nests (apparent nest success = 24.7%), five nests were abandoned (2.7%), and the remaining 132 (72.5%) were consumed by predators. We accurately determined nest fate (hatch or failure) at 98% of nests with a camera (N = 129), using evidence at nests. We misclassified nest fate at three nests (2%) because membranes were present, but camera footage revealed that predation by a badger occurred mid-hatch.
|
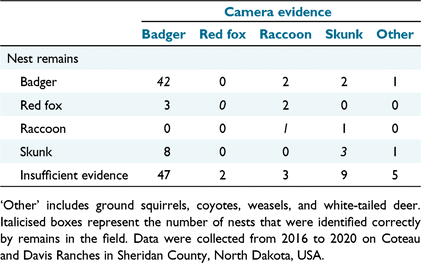
Predators observed on camera included badgers, skunks, raccoons, red foxes, ground squirrels (Urocitellus richardsonii and Poliocitellus franklinii), coyotes (Canis latrans), weasels (Mustela spp.), and white-tailed deer (Odocoileus virginianus). Using cameras to confirm predator identity, we determined that, with sufficient evidence (50% of nests), the Sargeant et al. (1998) criteria correctly predicted the predator at 69.7% of failed nests and incorrectly identified 30.3% of predators. Overall, the Sargeant et al. (1998) criteria allowed us to correctly predict predator identity at 34.8% of nests. Of the four main species we assessed using the Sargeant et al. (1998) criteria, our data suggested that raccoons were most likely to be misclassified (Table 2).
|
Evaluation of field-based predator identification accuracy
Across the study area, proportions of nest predators determined using the Sargeant et al. (1998) classification system were different from those determined using nest cameras (N = 514, χ2 = 16.55, d.f. = 3, P < 0.05). Results of within-year tests suggested that differences between the methods were greatest in 2020 (2016: N = 93, χ2 = 7.11, d.f. = 3, P = 0.07; 2017: N = 103, χ2 = 2.27, d.f. = 3, P = 0.52; 2018: N = 81, χ2 = 7.43, d.f. = 3, P = 0.06; 2019: N = 90, χ2 = 1.90, d.f. = 3, P = 0.59; 2020: N = 147, χ2 = 16.11, d.f. = 3, P < 0.05).
Discussion
We conducted an empirical evaluation of the Sargeant et al. (1998) classification system’s accuracy for fating nests and identifying nest predators of waterfowl. Our field-based estimates of nest fate accuracy (98%) were higher than those reported in previous studies of snowy plovers (Charadrius nivosus: Ellis et al. 2018: 84%), grassland-nesting passerines (Pietz and Granfors 2000: 85%), wood thrushes (Hylocichla mustelina: Williams and Wood 2002: 85%), and boreal-forest songbirds (Ball and Bayne 2012: 85%). Causes of misclassification in these studies included removal of eggshells by adults, size of remaining fragments, terrain surrounding nests, partial nest predation, and removal of eggs by large predators (Pietz and Granfors 2000; Ball and Bayne 2012; Ellis et al. 2018). Although we also encountered some of the challenges mentioned by these other studies, the only errors observed in our study were caused by a predator eating ducklings while they were hatching. Even these misclassifications could have been considered correct because ducklings were in the process of emerging from the eggs at the time of predation. We hypothesise that the larger eggs and clutch sizes of waterfowl compared with passerines and shorebirds might provide more evidence post-predation.
Although highly accurate (98%) for identifying waterfowl nest fates, the Sargeant et al. (1998) classification system was less clear-cut regarding predator identification. When sufficient evidence was available, estimates of accuracy for predator identification at the scale of individual nests were relatively high compared with those in previous studies (e.g. Williams and Wood 2002: 43%, Staller et al. 2005: 30% vs 69.7% in our study). However, based on Sargeant et al. (1998), 50% of the nests we examined via camera (N = 66) had insufficient evidence to assign a predator species. Many of those were removed early in the classification process because they had too few eggs (less than the six eggs) to provide evidence for identifying behavioural patterns of predation (Sargeant et al. 1998). Video footage revealed on several occasions that the low egg count was due to removal of eggs from nests by smaller predators (e.g. ground squirrels) or predation of nests before six eggs were laid. Although useful for eliminating sources that provide poor to no data, removing nests from the classification process due to insufficient evidence could unintentionally eliminate predators in a biased manner, suggesting that caution is needed when using this method for predator identification.
The Sargeant et al. (1998) classification system was developed as an approach to determine study site-level predation rates, suggesting that higher levels of accuracy might be expected at this scale. However, based upon the results of our pooled tests we also recommend that investigators should apply Sargeant et al. (1998) at the study area scale with equal amounts of caution. Although the Sargeant et al. (1998) classification system did identify badgers and skunks as the first and second-most abundant predators, it placed a heavier emphasis on red foxes and striped skunks than the camera data. Incidentally, these two predators were also the ones for which Sargeant et al. (1998) were able to obtain the highest sample size for verification data in their development of the classification system, which might play a role in this observed bias.
Based on our observations, some misclassifications likely occurred because the assumptions about predator behaviour made using the classification system were not fully supported. For example, according to Sargeant et al. (1998), dug areas were supposed to occur at almost 100% of badger predations, but we saw evidence of this behaviour at less than 40% of our nests where we observed badgers on camera (N = 100). Our observations at badger-depredated nests also suggest that other behaviours, like the size or position of holes left in eggs, might be more helpful in accurately identifying badgers as predators. At nests destroyed by badgers, we found large holes in 70% of eggs and those holes were mostly commonly located on the side of the eggs (67% of nests).
Because of the large number of misclassifications, the current Sargeant et al. (1998) criteria are likely to be unreliable for providing researchers with information about the proportion of predators in the study area population. The only reliable use of this system would be for identifying nest fate. However, it may be possible to improve field protocols such as those of Sargeant et al. (1998) by using camera observations at the nest to enhance our understanding of predator behaviours. To do this, cameras will need to be positioned with a field of view to capture predator behaviours both in the nest bowl and in the surrounding areas where behaviours like digging or caching may occur. Alternatively, recent studies have demonstrated that DNA from saliva samples can be used to identify predators of ground-nesting birds (Hopken et al. 2016), although this approach might not be as accessible. We recommend the continued use of camera studies and exploration of DNA-based identification where possible, and suggest that investigators in future studies explore the possibility of improving the Sargeant et al. (1998) criteria using data collected with video cameras.
Data availability
Predation data and R code are available upon request. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Funding for this project was generously provided by Ducks Unlimited, Inc., Enbridge Ecofootprint Grant Program, the Minnesota Association of Resource Conservation and Development Councils, The University of North Dakota’s (UND) College of Arts and Sciences Undergraduate Creative Activity Award, and the UND Department of Biology. We also appreciate The Nature Conservancy for providing us with access to the Davis Ranch, and the U.S. Geological Survey Northern Prairie Wildlife Research Center for use of additional ATVs for nest searching each summer.
Acknowledgements
This research would not have been possible without the efforts of several undergraduate interns and Ducks Unlimited staff members who collected data that contributed to this project. In addition, M. Sovada provided our team with field training and valuable study considerations. We would also like to thank T.W. Arnold for his review and comments, which greatly improved the quality of the manuscript.
References
ndes, ndes, (2019). Accuracy of nest fate classification and predator identification from evidence at nests of least terns and piping plovers. Ibis 161, 286–300.| Accuracy of nest fate classification and predator identification from evidence at nests of least terns and piping plovers.Crossref | GoogleScholarGoogle Scholar |
Anthony, RM, Grand, JB, Fondell, TF, and Manly, BFJ (2004). A quantitative approach to identifying predators from nest remains. Journal of Field Ornithology 75, 40–48.
| A quantitative approach to identifying predators from nest remains.Crossref | GoogleScholarGoogle Scholar |
Ball, JR, and Bayne, EM (2012). Using video monitoring to assess the accuracy of nest fate and nest productivity estimates by field observation. The Auk 129, 438–448.
| Using video monitoring to assess the accuracy of nest fate and nest productivity estimates by field observation.Crossref | GoogleScholarGoogle Scholar |
Blythe, EM, and Boyce, MS (2020). Trappings of success: predator removal for duck nest survival in Alberta parklands. Diversity 12, 119.
| Trappings of success: predator removal for duck nest survival in Alberta parklands.Crossref | GoogleScholarGoogle Scholar |
Burnam JS (2008) Using continuous video to monitor patterns of nest defense and incubation behavior in northern bobwhites Colinus virginianus. MS thesis, University of Georgia, Athens, GA, USA.
Burr, PC, Robinson, AC, Larsen, RT, Newman, RA, and Ellis-Felege, SN (2017). Sharp-tailed grouse nest survival and nest predator habitat use in North Dakota’s Bakken oil field. PLoS One 12, e0170177.
| Sharp-tailed grouse nest survival and nest predator habitat use in North Dakota’s Bakken oil field.Crossref | GoogleScholarGoogle Scholar |
Cox WA, Pruett MS, Benson TJ, Chiavacci SJ, Thompson FR III (2012) Development of camera technology for monitoring nests. In ‘Video Surveillance of Nesting Birds’. (Eds CA Ribic, FR Thompson III, PJ Pietz) pp. 185–210. Studies in Avian Biology No. 43. (University of California Press: Berkeley, CA, USA)
Ellis, KS, Cavitt, JF, Larsen, RT, and Koons, DN (2018). Using remote cameras to validate estimates of nest fate in shorebirds. Ibis 160, 681–687.
| Using remote cameras to validate estimates of nest fate in shorebirds.Crossref | GoogleScholarGoogle Scholar |
Ellis-Felege SN, Carroll JP (2012) Gamebirds and nest cameras: present and future. In ‘Video Surveillance of Nesting Birds’. (Eds CA Ribic, FR Thompson III, PJ Pietz), pp. 35–44. Studies in Avian Biology No. 43. (University of California Press: Berkeley, CA, USA)
Girard GL (1939) Life history of the shoveler. In ‘Transactions of the 14th North American Wildlife Conference’, Vol. 4. pp. 364–371. (Wildlife Management Institute: Washington, DC, USA)
Hopken, MW, Orning, EK, Young, JK, and Piaggio, AJ (2016). Molecular forensics in avian conservation: a DNA-based approach for identifying mammalian predators of ground-nesting birds and eggs. BMC Research Notes 9, 14.
| Molecular forensics in avian conservation: a DNA-based approach for identifying mammalian predators of ground-nesting birds and eggs.Crossref | GoogleScholarGoogle Scholar |
Klett AT, Duebbert HF, Faanes CA, Higgins KF (1986) Techniques for studying nest success of ducks in upland habitats in the Prairie Pothole Region. Resource Publication 158. United States Department of the Interior Fish and Wildlife Service, Washington, DC, USA. Available at https://apps.dtic.mil/sti/pdfs/ADA323206.pdf [Accessed 20 April 2021]
Lariviere, S (1999). Reasons why predators cannot be inferred from nest remains. The Condor 101, 718–721.
| Reasons why predators cannot be inferred from nest remains.Crossref | GoogleScholarGoogle Scholar |
Opermanis, O, Mednis, A, and Bauga, I (2001). Duck nests and predators: interaction, specialisation and possible management. Wildlife Biology 7, 87–96.
| Duck nests and predators: interaction, specialisation and possible management.Crossref | GoogleScholarGoogle Scholar |
Pietz, PJ, and Granfors, DA (2000). Identifying predators and fates of grassland passerine nests using miniature video cameras. The Journal of Wildlife Management 64, 71–87.
| Identifying predators and fates of grassland passerine nests using miniature video cameras.Crossref | GoogleScholarGoogle Scholar |
Pietz PJ, Granfors DA, Ribic CA (2012) Knowledge gained from video-monitoring grassland passerine nests. In ‘Video Surveillance of Nesting Birds’. (Eds CA Ribic, FR Thompson III, PJ Pietz) pp. 3–22. Studies in Avian Biology No. 43. (University of California Press: Berkeley, CA, USA)
Prinz, ACB, Taank, VK, Voegeli, V, and Walters, EL (2016). A novel nest-monitoring camera system using a Raspberry Pi micro-computer. Journal of Field Ornithology 87, 427–435.
| A novel nest-monitoring camera system using a Raspberry Pi micro-computer.Crossref | GoogleScholarGoogle Scholar |
Ribic CA, Thompson FR, Pietz PK (Eds) (2012) ‘Video Surveillance of Nesting Birds.’ Studies in Avian Biology No. 43. (University of California Press: Berkley, CA, USA)
Robel, RJ, Briggs, JN, Dayton, AD, and Hulbert, LC (1970). Relationships between visual obstruction measurements and weight of grassland vegetation. Journal of Range Management 23, 295–297.
| Relationships between visual obstruction measurements and weight of grassland vegetation.Crossref | GoogleScholarGoogle Scholar |
Sargeant AB, Sovada MA, Greenwood RJ (1998) Interpreting evidence of depredation at duck nests in the Prairie Pothole Region. US Geological Survey, Northern Prairie Wildlife Research Center, Jamestown, ND, USA.
Sherfy, MH, Anteau, MJ, Shaffer, TL, Johnson, MA, Reynolds, RE, and Ringelman, JK (2018). Density and success of upland duck nests in native- and tame-seeded conservation fields. Wildlife Society Bulletin 42, 204–212.
| Density and success of upland duck nests in native- and tame-seeded conservation fields.Crossref | GoogleScholarGoogle Scholar |
Staller, EL, Palmer, WE, Carroll, JP, Thornton, RP, and Sisson, DC (2005). Identifying predators at northern bobwhite nests. Journal of Wildlife Management 69, 124–132.
| Identifying predators at northern bobwhite nests.Crossref | GoogleScholarGoogle Scholar |
Thompson, FR, Dijak, W, and Burhans, DE (1999). Video identification of predators at songbird nests in old fields. The Auk 116, 259–264.
| Video identification of predators at songbird nests in old fields.Crossref | GoogleScholarGoogle Scholar |
Weidinger, K (2008). Identification of nest predators: a sampling perspective. Journal of Avian Biology 39, 640–646.
| Identification of nest predators: a sampling perspective.Crossref | GoogleScholarGoogle Scholar |
Weller, MW (1956). A simple field candler for waterfowl eggs. The Journal of Wildlife Management 20, 111–113.
| A simple field candler for waterfowl eggs.Crossref | GoogleScholarGoogle Scholar |
Williams, GE, and Wood, PB (2002). Are traditional methods of determining nest predators and nest fates reliable? An experiment with wood thrushes (Hylocichla mustelina) using miniature video cameras. The Auk 119, 1126–1132.
| Are traditional methods of determining nest predators and nest fates reliable? An experiment with wood thrushes (Hylocichla mustelina) using miniature video cameras.Crossref | GoogleScholarGoogle Scholar |