

The threatened Gouldian finch preferentially forages in prescribed burnt savannas
Sydney J. Collett A * , Ian J. Radford B , Mirjam Kaestli A , Tara L. Crewe A C , Stephen T. Garnett A and Hamish A. Campbell AA Research Institute for the Environment and Livelihoods, Charles Darwin University, Ellengowan Drive, Darwin, NT, Australia. Email: mirjam.kaestli@cdu.edu.au, stephen.garnett@cdu.edu.au, hamish.campbell@cdu.edu.au
B Department of Biodiversity, Conservation and Attractions, Biodiversity and Conservation Science, Kings Park, WA, Australia. Email: ian.radford@dbca.wa.gov.au
C Department of Lands and Forestry, Government of Nova Scotia, Kentville, Nova Scotia, Canada. Email: Tara.Crewe@novascotia.ca
International Journal of Wildland Fire 32(9) 1277-1290 https://doi.org/10.1071/WF22185
Submitted: 17 August 2022 Accepted: 10 June 2023 Published: 3 August 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Background: The granivorous finches of Australia’s tropical savannas heavily rely on a sequence of perennial and annual grass seed production to feed throughout the year. An increase in late dry season wildfires has been suggested to detrimentally effect seed production sequence and has been attributed to poor physiological condition and a reduction in fitness of granivorous finches. Early dry season prescribed burning is an asset protection management strategy often implemented to reduce the incidence of late dry season wildfire, but has also been shown to improve the abundance and nutritional quality of grass seed
Aims: To assess whether the Gouldian finch (Chloebia gouldiae) preferentially used areas that were subject to early dry season prescribed burning over areas that were not burnt.
Methods: The creation of a landscape mosaic across the landscape with varying fire histories. Then the assessment of individual finch movement and site utilisation using VHF-radio telemetry transmitters, detected by an array of static receivers deployed across the landscape.
Key results: Finches significantly preferred to forage in areas burnt in the early season every 2–3 years.
Conclusions: The study demonstrates that early dry season prescribed burning creates preferred foraging habitat patches for Gouldian finches within savanna fire mosaics.
Keywords: automated telemetry, early dry season burning, fire frequency, Gouldian finch, granivore, late dry season burning, prescribed burning, time since last fire.
Introduction
Fire has pervasive effects on many tropical and temperate ecosystems (e.g. Pausas and Ribeiro 2017; Pausas and Parr 2018). Open vegetation in the ‘savanna’ biome is largely maintained through fire (Bond et al. 2005; Sankaran et al. 2008), and many flora and fauna savanna species have adapted to the effects of fire (Andersen et al. 2005; Andersen 2021). However, in the northern Australian savannas, some savanna dwelling flora and fauna have become threatened by unmanaged fire (Andersen et al. 2005; Woinarski and Legge 2013; Ziembicki et al. 2015) as changes in management, invasive weeds, grazing pressure, and climate change alter fire regimes. While a key tool for managing fire in such environments is prescribed burning, there is also a need to understand how savanna species respond. This can be challenging because fire regimes interact with different organisms in variable ways (Williams et al. 2003; Woinarski et al. 2005; Woinarski and Legge 2013; Radford et al. 2020; Andersen 2021). Plants generally respond to fire intensity and interval, which can influence both mortality and reproduction, via fire response traits or adaptations (Clarke and Gorley 2015). Unlike plants, animals can often avoid the direct heat impact of fire by moving or hiding in safe refuges (Sutherland and Dickman 1999; Johnson 2006), but the resources they depend on are influenced by the spatial distribution, intensity and frequency of the fire.
Granivorous finches are found throughout the fire-prone savannas of northern Australia (Tidemann et al. 1992; Franklin 1999; Woinarski and Legge 2013). Declines of some granivorous finches, including the threatened Gouldian finch, have been attributed, at least in part, to an increase in the frequency and extent of late dry season fires (Franklin et al. 2005; Reside et al. 2012; Williams et al. 2020). This is because such fires both burn surface seed and reduce both seed availability and quality the following year (Weier et al. 2017, 2018) while finches captured within areas exposed to late season hot fires had higher levels of stress hormones and lower body condition (Maute et al. 2013; Legge et al. 2015). Early dry season prescribed burning can reduce the frequency and intensity of late dry season fires in tropical savannas, creating refuges from hot late dry season fires where grass seed resources have a greater diversity, density, and quality than in surrounding areas (Weier et al. 2017, 2018, 2019).
However, the fine-scale of free-ranging granivorous finches movement has never been assessed, so it is unknown whether they seek out and preferentially occupy areas that have undergone early dry season prescribed burning over areas that have not. Such information would greatly assist managers in determining the timing and frequency of prescribed burning as a management strategy for threatened species conservation.
A possible reason why the fine-scale tracking of free-ranging granivorous finches has not yet been undertaken is due to their small body size and highly mobile nature. This means that satellite fixing devices are too bulky to be attached to individuals, and active tracking provides too few and infrequent location fixes to adequately model site utilisation over sufficient periods of time. To overcome these limitations, we trialled the application of coded VHF-nano transmitters (<0.23 g) and an array of static listening stations deployed throughout the landscape. As a case study, we chose the Gouldian finch (Chloebia gouldiae). The species has undergone significant decline across Australia’s tropical savannas, and whilst the population has shown recent signs of recovery, it is still listed as threatened under the Australian Environmental Protection and Biodiversity Conservation Act (1999). Out of all the Australian tropical savanna finches the Gouldian finch may be expected to have the greatest sensitivity to the impacts of fire because it solely feeds upon grass seeds. Whereas other sympatric finches will have a broader diet, especially during the late dry season when grass seed becomes scarce (Collett et al. 2022a).
Early season prescribed burning was undertaken within an 100 km2 region of the Kimberly’s over a 3-year period to create a fine-scale mosaic of differing fire histories. Remotely sensed fire scar information was overlaid with individual site utilisation information to test if: (1) finches would show a preference for recently but infrequently burnt savannas; (2) finches would preferentially use patches burnt in the early dry season and avoid areas burnt in the late dry season; and (3) finches would expand site utilisation as the dry season progressed to locate declining seed resources.
Methods
Study area
The study was conducted during 2019 in the hills around Wyndham (15°28′S, 128°07′E; Fig. 1) in the east Kimberley region of Western Australia. Most of the average annual rainfall of ~910 mm falls between December and March (monsoon wet season; Bureau of Meteorology 2020). The annual mean day temperature for 2019 was 37.2°C, with daily maximums ranging from 25.0 (June) to 45.5°C (December). The main topographic features of this area are rocky sandstone ridges and gently sloping savanna hills with a tree canopy dominated by Corymbia dichromophloia and Eucalyptus miniata (Brazill-Boast et al. 2013), a ground layer in which the annual grass Sorghum stipoideum and the perennial grasses Triodia bitextura, Eriachne obtusa and Sorghum plumosum predominated. There is a pronounced fire season throughout the east Kimberley region. In the early dry season (before July), small, low-intensity hazard reduction burns are lit by land management agencies to create fire breaks. Few naturally or accidentally ignited fires occur during this season. In the late dry season (July–November), large high intensity wildfires are ignited annually, either by lightning, particularly during the build-up or storm season (September–November), or by anthropogenic ignition sources (roadworks, campfires, accidents, arson). An average of 60% of the savanna in the immediate Wyndham area was burnt annually from 2016 to 2019.
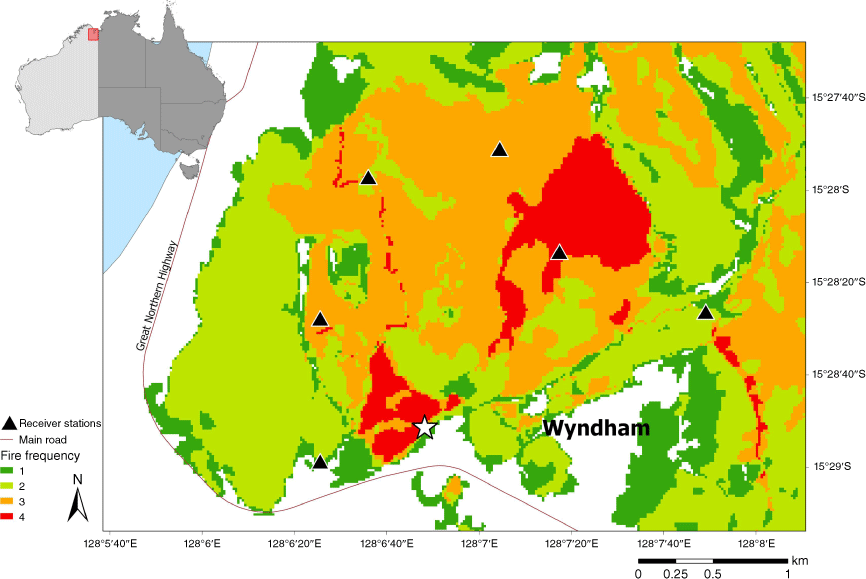
Prescribed burning
Prescribed burning was undertaken both to increase the abundance and quality of grass seeds available for Gouldian finches (Weier et al. 2016, 2017, 2018, 2019), and also to reduce the negative impact of high intensity wildfires on tree nesting hollow density that finches rely on during breeding (Brazill-Boast et al. 2011; Radford et al. 2021). To achieve this, strips of inter-connected savanna vegetation were burnt early in the dry season. Finch habitat in the study area has a history of annual incursions by wildfires from the adjacent township, and one of the functions of networks of burnt habitat was to reduce the incursion of these high-intensity wildfires into finch habitat. Prescribed burning was applied by hand in a series of walked and driven lines using matches and drip torches. Fire lines were planned to be a sufficient distance apart that fire spread rates did not allow adjacent lines to meet prior to self-extinguishing (soon after ignition or overnight). To achieve small patches of prescribed burnt habitat (~<300 m wide or 10–100 ha), ignition was planned for the wet-dry season transition, immediately after annual Sorghum seed had dehisced, under humid, low wind conditions, and in grass fuels which were not fully cured (usually March–April in east Kimberley). The target for prescribed burning in Gouldian finch breeding habitat was ~20–30% of finch habitat annually. This burning regime was designed to provide sufficient newly burnt savanna to benefit finches while breeding and feeding during the dry season (Weier et al. 2016), and also provide habitat with infrequent enough fires (once every 3–4 years) to allow slow maturing perennial grasses such as Triodia spp. to produce seeds for wet season feeding (Dostine et al. 2001; Legge et al. 2015).
Bird sampling
In 2019, we conducted three finch sampling sessions in the early (April/May), mid (July) and late (October) dry season. Mist nets (12 m length, 2.7 m height, 25 mm mesh) were used to catch finches across 25 days at two waterholes approximately 11 km apart. Nets were positioned to capture birds leaving the water source following their morning drink and were continually monitored. After the birds were extracted from the nets, a uniquely numbered size-two aluminium band was placed on the left tarsus of any bird not captured previously. Each bird was weighed using a Pesola scale (±0.5 g), and the wing, head-bill, tail, and tarsus length were measured (±1 mm). The sex and age of the birds was determined from their plumage – adult male Gouldian finches are more brightly coloured than females, particularly the purple colouration of the breast. Juvenile Gouldian finches are a dull grey and cannot be sexed using plumage.
Monitoring movement
Coded VHF transmitters (Avian NanoTag model NTQB2-2, Lotek Wireless Inc.) were attached to Gouldian finches using a figure-8 leg loop harness made from an elastic thread (Rappole and Tipton 1991). The elastic thread degraded and fell off from 30 to 130 days after application, as confirmed by recapturing birds that had previously carried transmitters. Each transmitter and harness weighed 0.31 g, which was 2–3% of the average weight of the individual birds (13.8 g). Several sizes of harness (29, 30, 31, and 32 mm loop sizes) were used because size varied among individuals. Each transmitter was coded with a uniquely identifiable sequence of pulses emitted every 11–13 s at a radio frequency of 151.500 MHz. Tags were active for a 12 h on/off cycle to capture movement during the daylight hours (05:00 hours–17:00 hours) but save the battery at night.
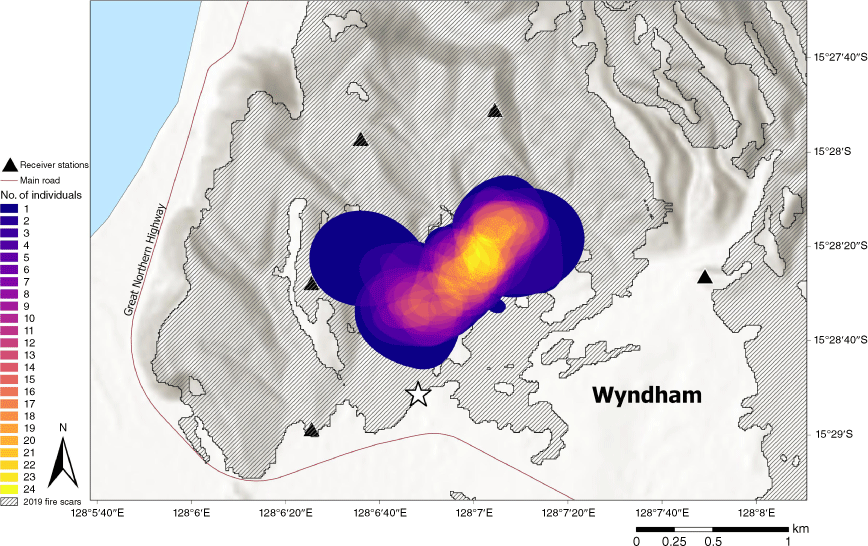
Movements of tagged Gouldian finches were monitored using an array of six receiver stations. Each receiver station listened continually and recorded signal strength, receiver location, and the time and date of each detection of a coded transmission. Receivers were, on average, 1.7 ± 0.78 km apart and provided continual coverage over approximately 21.2 km2 and encompassed all tagging locations. Each receiver contained a power source, a receiver, and an antenna tuned to the 151.500 MHz frequency. Each receiver was connected to a monopole antenna (5.1 dBi), which had a 360° detection radius and a range of up to 4 km (mean = 3 ± 0.963 km). Each antenna was vertically mounted to a 5 m length × 5 cm diameter galvanised steel post. The Sensorgnome device was mounted to the pole approximately 1 m above the ground. Receivers ran the open-source Sensorgnome software on a raspberry-pi 3 device (Compudata Systems, London, Ont., Canada) with Adafruit Ultimate GPS Hat maintaining time synchronisation for each device, and Nooelec NESDR SMArTee RTL-SDR radio dongles receiving the signals from the tags. Each receiver station was powered by solar panels and a 12 V deep-cycle battery. This arrangement provided 5 days of power redundancy and lasted throughout the dry season and well into the wet season, when cloud cover was more prevalent.
When multiple receivers detect the same bird through the unique sequence of beeps and pauses, the position of the bird can be estimated by the variation in signal strength of the detections. Using multilateration, the signal strength from several receiver stations (3+ simultaneous) was used to estimate the location of a tagged individual. Distance to position estimates were ground-truthed using a drone, as described in Collett (2023).
Fire mapping
Sentinel 2 imagery (bands 11 and 12; resolution 20 ×20 m) was used to map fire history and frequency from when it first became available from February 2016 to November 2019. Whilst other imagery was available from before February 2016, it lacked the fine-scale resolution of the Sentinel imagery required for the analysis. At least one image per month with minimal cloud cover (<5%) was chosen from February through to November each year. No fires were recorded in January and December. Images were downloaded as level 1C products from the Sentinel-hub EO browser (https://app.sentinel-hub.com/eo-browser), and the 11 and 12 bands were used as they reflect short-wave infrared reflection (SWIR).
Object-based image analysis (OBIA) in SAGA GIS (ver. 2.3.2; Conrad et al. 2015) was used to classify burnt areas. The two steps involved in OBIA are segmentation and classification. Firstly, the image was segmented into unclassified objects based on spectral properties, as defined and set by the user. Secondly, images were classified as either ‘burnt’ or ‘unburnt’ using a set of predetermined features and criteria. Manual on-screen editing also complemented the classification process and resulted in four burnt-area maps from 2016 to 2019. From these maps, we determined four variables attributed to fire history: (1) area burnt (m2); (2) year of burn; (3) month of burn; and (4) whether the fire was an early dry season (February–June) or late dry season burn (July–November).
The map of each variable was converted to a raster, and the raster calculator in QGIS was used to create fire frequency and time-since-last-fire maps. These maps showed: (1) overall frequency of fire through the area (FF); (2) frequency of early dry season burns (number of years burnt between 1 March and 30 June in the previous 4 years (EDSFF)); (3) frequency of late dry season wildfire (number of years burnt between 1 July and 1 December in the previous 4 years (LDSFF)); and (4) time-since-last-burn (months, TSLF); Table 1, see Supplementary Fig. S1).
| Variable | Abbreviation | Description | Range |
|---|---|---|---|
| Time-since-last-fire (months) | TSLF | The number of months since the last fire occurred | 0–48 |
| Fire frequency | FF | The number of times site burned in 2016–2019 | 1–4 |
| Early dry season (EDS) fire frequency | EDSFF | The number of times site burned in the early dry season (March–June) in 2016–2019 | 1–4 |
| Late dry season (LDS) wildfire frequency | LDSFF | The number of times site burned in the late dry season (July–November) in 2016–2019 | 1–4 |
| Core area month | Month | The calendar month during which the bird was tracked | 5–11 |
Data preparation and analysis
Four levels of filtering were applied to the data. First, tracking data from the first 24 h following tag deployment were excluded from analysis to allow for a post-tagging acclimation. Second, detections with a run length of <3 were excluded to minimise the chances of false positives (Crewe et al. 2018), a run length being defined as the number of consecutive tag transmissions received by any one antenna. Third, detections were filtered to the six receivers that were active over the entire sampling period. Finally, birds were dropped from analysis if they were detected for fewer than 5 days per month in order to ensure that the sample of an individual’s behaviour was robust and representative.
Individual centres of activity (CoA) were estimated using non-linear least squares (NLS), whereby a position during each 7-min time interval was estimated by minimising the difference between estimated and ‘known’ distances from a tower for a minimum of three simultaneous detections. The ‘known’ distance from a tower was estimated using the relationship between signal strength and distance from each tower, collected using drones with nanotags attached (Fig. S2; Crewe, Udyawer, Collett, Radford, Micheli-Campbell, Campbell, unpubl. data). The 7-min interval was used, as this minimised the error in position estimates when comparing the known drone positioning (via onboard GPS) with the estimated position of the drone (with the maximum likelihood estimation method used here; Fig. S3).
We used the fixed kernel method to estimate utilisation densities (KUD) to assess core use areas, where positions were predicted via the non-linear least squares method, using the raw detections. For each individual, the 50% volume contour (in m2) of the KUD was created for each month of the tracking period using the ‘adehabitatHR’ package (Calenge 2006) in the software R. All KUD calculations used the default reference bandwidth as the smoothing parameter.
In order to determine if there was a relationship between core use areas and fire history, we clipped fire variable maps to the extent of the receiver station detection range and calculated the area for each level of the fire variable within the detection range. Each monthly core area was then overlaid with the clipped fire variables. Finally, the proportion of the available habitat that the birds used was calculated by determining the area of each fire level that intersected with the core use area using the ‘sp’ package in R software.
To determine the preferences in site use in response to the different fire variables, we used a permutational multivariate analysis of variance (PERMANOVA) in PRIMER ver. 7 (Anderson 2001; Clarke and Gorley 2015). Fire variables were normalised, and a Euclidean distance matrix calculated that was used as an outcome in the PERMANOVA models. Main and pairwise testing was conducted using 997–999 permutations to assess changes in habitat use according to each level of fire history. The available habitat was randomly sampled to test whether finches actively favoured a particular element of the fire history. To sample the survey area randomly, circular polygons were created that had an area equal to the average core use area of all birds. These polygons were located within the available habitat, where multilateration would be possible (maximum detection range for two or more towers). For each month, the number of these polygons sub-sampled was twice the number of core use areas estimated for that month. Up to half of the random samples were the same between 1 month and the next, with the rest randomly sub-sampled each month. The same random samples were used to assess the four fire variables in four different PERMANOVA models. A binary fixed factor was included in the tests to distinguish between site use (i.e. Kernel utilisation distributions (KUD) of birds and the random samples of available habitat; hereafter called ‘Bird vs Avail.’). Tag ID marking individuals was included as a random effect, nested within the ‘bird vs available’ factor, and both the ‘bird vs available’ variable and month were included as fixed factors.
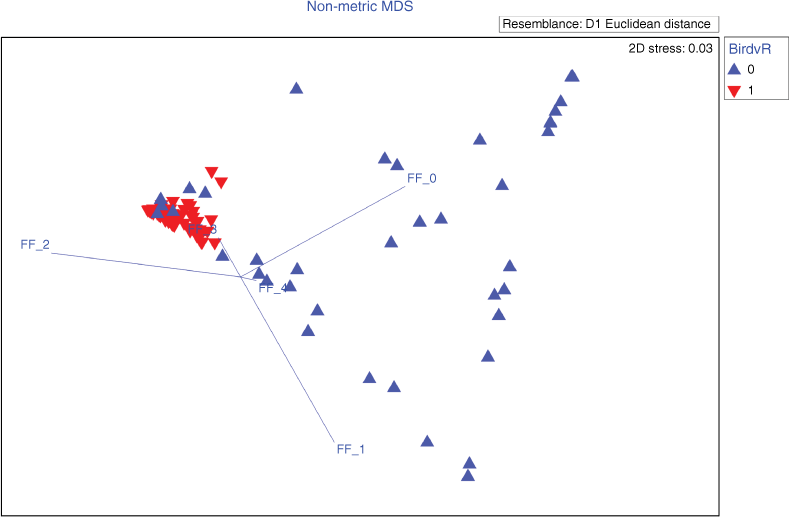
Variables that showed a significant difference between the birds’ KUD and random samples of the available habitat or across months (P < 0.05) were further used for Similarity Percentages Procedure (SIMPER) analysis. SIMPER was used to identify the most influential fire interval or fire frequency for each fire history variable model, which accounted for most of the observed dissimilarity in fire patterns between months and/or between used habitat versus available habitat. A Non-metric Multi-Dimensional Scaling (nMDS) ordination was performed to visualise the multivariate data (i.e. the fire history patterns of each bird’s use area compared to a random sample of the available habitat (Fig. S4)). The nMDS for FF, EDFF and LDSFF had stress levels <0.2, allowing 2D interpretation of birds’ space use. However, the TSLF variable had an nMDS stress of 0.27, which did not allow for a confident interpretation of the results.
The most influential levels of each variable identified in the SIMPER analysis were analysed further to identify which variable had greatest influence on the birds’ space use. A generalised linear mixed model (GLMM) was used to assess the influence of fire history variables and the KUD months on individuals’ space use in response to selected fire variables. The level of each fire variable that contributed the most to the observed dissimilarity between the habitat in core use areas and available habitat (2-years EDFF, 1-year LDFF, 3-years FF and 16-months TSLF) was used as an explanatory variable in the model. As this model examined the proportion of the total area occupied within a bird’s core use area, a beta regression was used. The Tag ID was included as a random effect in the model. Models were run using the ‘glmmTMB’ package (Brooks et al. 2017) in R Software (). Model residuals were checked to ensure a lack of pattern across predictors and fitted factors, as well as to identify influential outliers.
Results
Finch capture
Across three tagging sessions between 30 April and 10 October 2019 (tagging sessions – April, July, October), 68 Gouldian finches were fitted with nano VHF transmitters and their movements tracked. Of these tagged birds, 30 had sufficient data to conduct Kernel Density estimates (KUD; a minimum of five re-locations). These individuals were detected from between 7 and 130 days, with detections occurring at an average of 29.4 ± 2.8 tracking days per month.
Activity space
After the pre-processing, a total of 3 203 979 detections were collected from the 30 birds in this study (Table 2, Table S1). The mean core use area (50% KUD) in July was almost twice the size of the core use areas in other months (433 838 ± 68 842 m2 s.e.). The smallest core area used by tracked birds, in October (176 462 ± 44 169 m2 s.e.), was approximately half that used by birds tracked in July. There was little variation in the location of the core use areas throughout the dry season between individuals or months (Fig. 2, Figs S5, S6, Table S2).
| Month | No. of birds | Mean detections (± s.e.) | Mean days (± s.e.) | 50% KUD area (m2 ± s.e.) |
|---|---|---|---|---|
| May | 5 | 34 026 ± 5277 | 20 ± 1.1 | 292 933 ± 76 167.4 |
| June | 6 | 56 540 ± 16 078 | 25 ± 3.6 | 304 538 ± 82 052.1 |
| July | 18 | 36 804 ± 5804 | 16 ± 1.7 | 433 838 ± 68 842.4 |
| August | 13 | 48 543 ± 6422 | 27 ± 1.5 | 264 879 ± 54 412.8 |
| September | 8 | 29 450 ± 5591 | 21 ± 2.2 | 266 685 ± 48 569.1 |
| October | 11 | 24 341 ± 4048 | 18 ± 2.2 | 176 462 ± 44 168.8 |
| November | 5 | 25 032 ± 4756 | 22 ± 1.9 | 231 658 ± 63 785.7 |
Mean detections, mean days and 50% KUD area is given as the mean across all tagged birds for that month.
Note: KUD, kernel method to estimate utilisation densities.
Fire
Fire history variables had a strong influence on 50% of the birds’ core use area, as shown by a significant difference between the fire-related characteristics of an individual’s core use area and those of the surrounding area (Table 3). The fire patterns differed significantly between the birds’ core use area and available area for all fire pattern models (Permanova P < 0.01, for all).
| Bird vs Avail.A | Month | Tag (nested in month) | Bird × month | Residuals | |
|---|---|---|---|---|---|
| Overall fire frequency (FF) | |||||
| Pseudo-F (d.f.) | 28.5 (1) | 2.4 (6) | 2.4 (100) | 2.7 (6) | – |
| P-value | 0.001*** | 0.007** | 0.001*** | 0.002** | – |
| ECVB | 0.35 | 0.12 | 0.29 | 0.18 | 0.29 |
| Prescribed burn frequency (EDSFF) | |||||
| Pseudo-F (d.f.) | 46.5 (1) | 3.3 (6) | 3.0 (100) | 3.3 (6) | – |
| P-value | 0.001*** | 0.004** | 0.004** | 0.002** | – |
| ECVB | 0.46 | 0.15 | 0.33 | 0.21 | 0.29 |
| Wildfire frequency (LDSFF) | |||||
| Pseudo-F (d.f.) | 15.0 (1) | 0.7 (6) | 2.1 (100) | 0.9 (1) | – |
| P-value | 0.001*** | 0.645 | 0.001*** | 0.532 | – |
| ECVB | 0.21 | −0.05 | 0.23 | −0.04 | 0.27 |
| Time-since-last-fire (TSLF) | |||||
| Pseudo-F (d.f.) | 6.0 (1) | 16.4 (6) | 1.9 (100) | 4.8 (6) | – |
| P-value | 0.001*** | 0.001*** | 0.001*** | 0.001*** | – |
| ECVB | 0.18 | 0.51 | 0.3 | 0.36 | 0.39 |
Bird vs Avail., month tag and bird × month are the variates and co-variates included in the individual PERMANOVAs for the four fire history types. The Residuals ECV shows the unexplained within-group component of variance.
*P < 0.05; **P < 0.01; ***P < 0.001.
A‘Bird vs Avail.’ identification of whether the sample was from a bird’s KUD, or randomly sampled surrounding area.
BECV is the estimates of components of variance and is noted as the square root of this value. It marks the effect size in the unit of the dissimilarity matrix and is independent of the df.
There was a large difference in fire frequency patterns for early dry season fires between the habitat in core use areas and the available habitat (PERMANOVA: P-value = 0.001, the square root of the estimate of component of variation (ECV) = 0.46; Table 3). The SIMPER testing found that 48% of the observed dissimilarity between habitat in core use areas and the available habitat could be attributed to the 2-year (out of 4-year) prescribed burning frequency (Fig. 3). Between 75% and 99% of an individual’s core use area consisted of areas where there had been 2-year early dry season prescribed burning, compared to an average of 52% for the randomly chosen areas (Fig. 4a).
Non-metric MDS ordination plot comparing the prescribed burning fire histories (fire frequency 0–4) chosen by birds (red ‘down’ triangles), with those histories that were available in the habitat (blue ‘up’ triangles). The vectors (FF 0–4) indicate the direction and strength of the Pearson correlation of the fire variables with the nMDS axes (with the centre point of the vectors placed at a random location on the nMDS). FF, fire frequency; MDS, multi-dimensional scaling.
The proportion of an individual bird’s core use area, made up of (a) the varying frequencies by any fire, prescribed burns, wildfires for the bird’s core use areas (dark grey), and that which is available in the habitat (light grey); (b) the varying time-since-last-fire histories, between bird’s core use areas (dark grey), and that which is available elsewhere in the habitat (light grey).
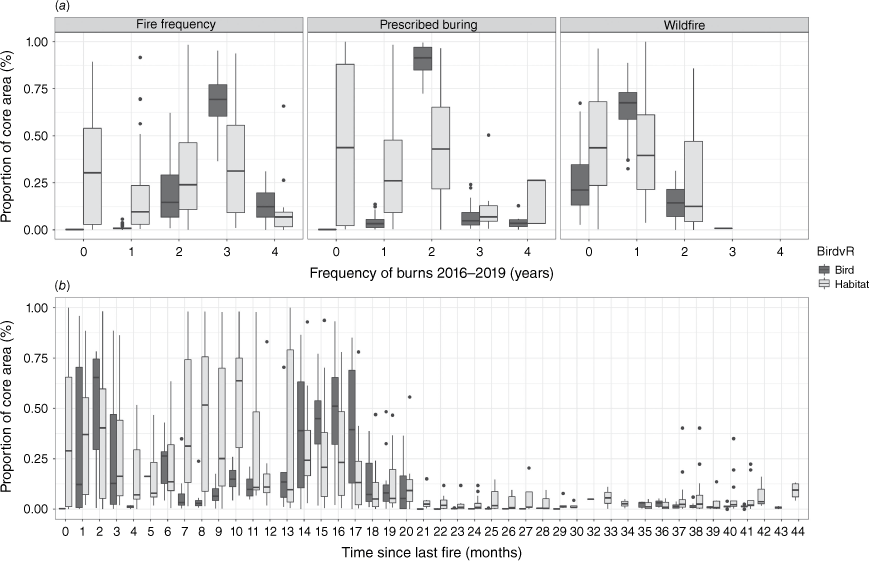
The overall fire frequency also influenced the core use area of birds, albeit slightly less than the prescribed burning patterns (PERMANOVA: P-value = 0.001, ECV = 0.35 (Table 3); beta regression: −1.63; P < 0.001 (Table 4, Fig. 5)). On average, 65% of the birds’ core use areas had experienced fire three times over the past 4 years (Figs 4a, 5) – thereby contributing 74% to the observed fire pattern similarity for birds – compared to 28% for available random samples.
| Estimate | s.e. | z-value | Pr (>|z|) | |
|---|---|---|---|---|
| (Intercept) | 1.96 | 0.20 | 10.03 | <0.001*** |
| 1-year late dry season fire | −1.70 | 0.12 | −14.40 | <0.001*** |
| 16-months since last fire | −2.37 | 0.14 | −16.95 | <0.001*** |
| 3-years overall frequency | −1.64 | 0.12 | −13.87 | <0.001*** |
| June | 0.19 | 0.21 | 0.90 | 0.367 |
| July | 0.44 | 0.19 | 2.34 | 0.019* |
| August | −0.05 | 0.19 | −0.26 | 0.795 |
| September | 0.22 | 0.21 | 1.05 | 0.295 |
| October | 0.49 | 0.21 | 2.30 | 0.022* |
| November | 0.55 | 0.28 | 1.93 | 0.054 |
Explanatory variables included the level of each fire variable that contributed the most to a bird’s core use area (2-years early dry season [intercept], 1-year of late dry season wildfire, 16-months-since-last-burn and 3-years of overall fire frequency) and the month for which the core use area was estimated (May [intercept] to November 2019).
*P < 0.05; **P < 0.01; ***P < 0.001.
Based on the beta regression model, predicted mean ± 95% CI proportion of the bird’s core use area occupied by the most influential level of the four fire variables (1-year wildfire frequency, 16-months-time-since-last-fire, 2-years prescribed burning frequency, and 3-years overall fire frequency).
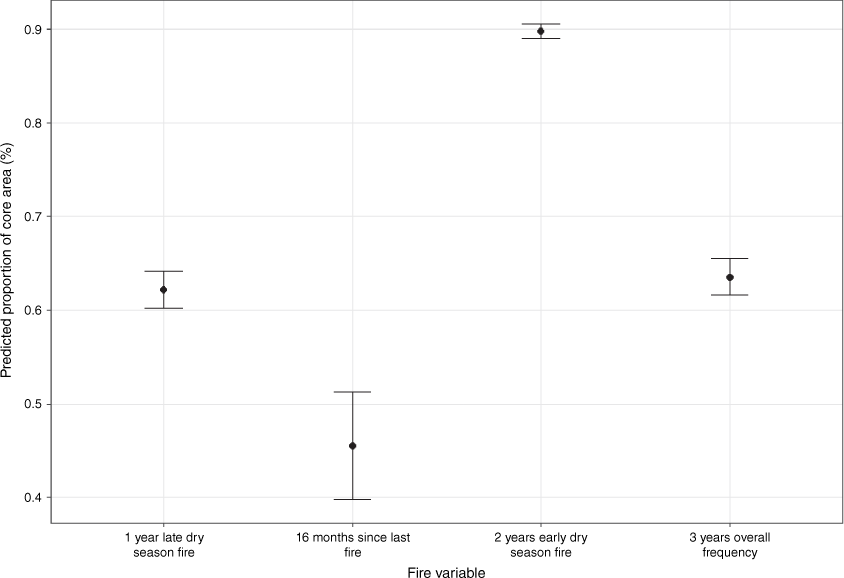
The frequency of late dry season wildfires was the least influential fire frequency type, particularly compared to the prescribed burning patterns (PERMANOVA: P-value = 0.001, ECV = 0.21 (Table 3); beta regression: −1.69, P < 0.001 (Table 4, Fig. 5)). Gouldian finches avoided areas that were burnt more than twice in the past 4 years by late season wildfires with on average, 64% of their core use areas burnt once by late season wildfires (Fig. 4a) – thereby contributing 72% to the observed fire pattern similarity for core use areas, as compared to 53% of the randomly chosen areas.
Recently burnt areas (0–2 months) made up some of a bird’s core use area (contributing 11.5% [0 months], 9.2% [1 month] and 10.6% [2 months] to the observed dissimilarity of fire pattern between the birds’ space use and randomly chosen areas), while 16-months post-fire contributed slightly more (15% to the observed similarity in core use areas). However, time-since-last-fire was the least influential fire variable to explain a bird’s use of the core area (PERMANOVA: P-value = 0.001, ECV = 0.18 (Table 3); beta regression: −2.37; <0.001) (Fig. 4b).
The dissimilarity between the fire history of a bird’s core use area and the fire history of the landscape changed to during the year (Table 3) and between fire history variables. The average dissimilarity between the fire frequency patterns of a bird’s core use area and the surrounding habitat was greatest during November and least in August (Table 5). However, the greatest dissimilarity between the birds’ core use area and the surrounding landscape was in May for EDSFF and November for LDSFF (Table 5). This suggests that birds more strongly preferred high EDSFF habitat early in the dry season and high LDSFF habitat late in the dry season.
| Overall Fire frequency (FF) | Prescribed burn frequency (EDSFF) | Wildfire frequency (LDSFF) | Time-since-last-fire (TSLF) | |||||
|---|---|---|---|---|---|---|---|---|
| P-value | Average distance | P-value | Average distance | P-value | Average distance | P-value | Average distance | |
| May | 0.02* | 0.682 | 0.002** | 1.026 | 0.98 | 0.456 | 0.001*** | 1.05 |
| Jun | 0.011* | 0.652 | 0.013* | 0.709 | 0.101 | 0.519 | 0.004*** | 0.900 |
| July | 0.001*** | 0.691 | 0.001*** | 0.819 | 0.001*** | 0.510 | 0.001*** | 0.864 |
| Aug | 0.002** | 0.588 | 0.001*** | 0.756 | 0.104 | 0.491 | 0.001*** | 0.834 |
| Sept | 0.003** | 0.703 | 0.001*** | 0.811 | 0.037* | 0.495 | 0.009** | 0.591 |
| Oct | 0.001*** | 0.662 | 0.002** | 0.626 | 0.009** | 0.477 | 0.004*** | 0.544 |
| Nov | 0.012* | 0.726 | 0.032* | 0.819 | 0.038* | 0.550 | 0.064** | 0.674 |
Average distance is noted as the average Euclidean distance of the fire frequency or TSLF patterns between the individual birds’ core use areas and that is available in the surrounding habitat.
*P < 0.05; **P < 0.01; ***P < 0.001.
Discussion
The results supported the hypothesis that the granivorous Gouldian finch (Chloebia gouldiae) preferentially foraged within areas that had undergone early dry season prescribed burning. This is because early dry season prescribed burning increases the density and nutritional value of annual Sorghum (Weier et al. 2018) and perennial grass seeds (e.g. Triodia spp.) (Dostine et al. 2001; Legge et al. 2015). During this study period, annual Sorghum seed density was higher in areas preferentially used by finches (41.5 seeds m−2) than elsewhere in the landscape (31.7 m−2), and seeds were more frequently present in randomly placed quadrats in preferred habitat (75% of quadrats compared to 60%). Individual finches will seek to offset the energetic costs of foraging by maximising energy intake, and therefore, we would expect individuals to spend a disproportionate amount of their time foraging within areas with greater seed availability. Prescribed burning has also been shown to improve the retention of old eucalypt trees with nesting hollows used by breeding Gouldians, (Radford et al. 2021), and this study provides further merit for the practice for the conservation and management of Gouldian finches.
A surprising observation from this current study was that areas subject to a single prescribed fire were not preferred by foraging Gouldian finches. Instead they preferred to forage within areas burnt two or three times. An explanation for this may be that this single prescribed fire lit during the wet season of 2019 spread further than anticipated, with older, relatively dry litter fuels sustaining fire over several days because the wet season rainfall had been unusually low in 2019. We propose that this fire killed most growing Sorghum plants before they set seed, reducing seed availability to very low levels compared to areas burnt after seed set in the early dry season fires later the same year. This inadvertent negative impact of wet season burning on Sorghum plant and seed set density is consistent with several previous studies (Williams and Lane 1999; Weier et al. 2018). This evidence provides a cautionary tale for fire managers in known Gouldian finch breeding areas. Prescribed fires in finch breeding habitats should be restricted to small areas or not done during the wet season before Sorghum seed is set in March/April. Conversely, prescribed patchy medium to low frequency, low-intensity burning during the early dry season once Sorghum seed has matured and dropped can benefit foraging finches based on the evidence provided here and elsewhere (Weier et al. 2018, 2019).
Previous research suggested that Gouldian finches would avoid areas frequently burnt in the late dry season because seed density was lower in the following year, but this had remained untested until now (Weier et al. 2016). However, our results showed that late dry season fire did not influence Gouldian finch foraging, with areas burnt in the late dry season used at the same frequency as their availability in the landscape. This suggests that these areas were not actively avoided during foraging in this study, presumably because seeds were available here, albeit possibly at lower densities than in areas of early dry season burning. Late dry season fires are a ubiquitous feature of savanna ecology. While these fires can negatively impact seed density (Weier et al. 2016, 2018) and nest hollow availability (Radford et al. 2021), the greatest negative influences of late dry season burning may be confined to unmanaged landscapes with a high frequency of very extensive late dry season wildfires and an absence of early fire. Whilst the space used by Gouldian finches was not influenced by late dry season fire here (either positively or negatively), it was noted that finches did not occupy areas frequently burnt by late dry season fires. Previous studies highlighted that the probability of Gouldian finch occupancy declined after 3 years of LDS fires (Reside et al. 2012). Fire regimes dominated by extensive, late dry-season wildfires have been associated with poor outcomes for fauna, including some bird species (Reside et al. 2012; Woinarski and Legge 2013) as well as other native savanna fauna (Andersen et al. 2005; Woinarski et al. 2011; Lawes et al. 2015; Radford et al. 2020; Stobo-Wilson et al. 2020). These repeated extensive late dry season wildfires are associated with a loss of resource patchiness (Price et al. 2003, 2012; Radford et al. 2020). A loss of patchy resources, particularly perennial grass seed resources produced only during the wet season when annual Sorghum seeds are not available (Dostine et al. 2001), will not only have detrimental impacts on the ongoing recovery of Gouldian finches but will also affect the distribution of many other savanna granivores.
We also found that Gouldian finches reduced the time spent within the study area towards the end of the dry season when key food resources were at their lowest abundance. This was consistent with our hypothesis that finches increase their foraging movements in the late dry season to search for remaining patches of resources across savanna landscapes (Collett et al. 2022a). Gouldian finches’ resource specialisation requires them to track the remaining Sorghum seed throughout the landscape in the late dry season until the wet season resources become available with the initiation of flowering and seed set by a range of perennial grass species. Morphological adaptations for long-distance flight are thought to enable increased flight activity by the granivore specialist Gouldian finch towards the end of the dry season relative to less specialised finches when there are resource bottlenecks (Franklin et al. 2017; Collett et al. 2022a). This supports findings that Gouldian finches reduce activity in core use areas as the dry season progresses, as they disperse more widely across the savanna to locate the remaining seed resources (Collett et al. 2022a).
Does enhanced seed foraging resources in prescribed burnt areas fully explain the marked concentration of finch activity in a relatively small part of the study landscape? Or are other factors involved? We propose that only minor differences other than seed availability occurred in preferred and non-preferred habitats, which are unlikely to be as important. There was little difference between preferred and non-preferred habitat in the percentage of habitat burnt when finches were foraging (38 and 41% burnt, respectively). Perennial and annual Sorghum grass cover was similar in preferred vs non-preferred habitat (4.1, 2.3% compared to 3.3, 2.1%). In the herbaceous layer, Triodia spp. were more commonly recorded in preferred than in non-preferred habitats. Gouldian finches have a huge daily requirement for water (Collett et al. 2022b). However, there was equal access to off-site watering points among preferred and non-preferred habitats. Tree cover was slightly higher in the preferred habitat (18.1% canopy cover compared to 14.6%) and there was also slightly higher cover of smooth-barked eucalypts favoured by breeding finches (C. dichromophloia, E. miniata) (Radford et al. 2021). Future research must assess the relative contribution of these additional environmental factors compared to prescribed burning enhanced seed availability if a full understanding of Gouldian finch ecology is to be achieved.
Future research would ideally extend the temporal scale over which the relationship between finches and fire was assessed. We could analyse only one savanna area in this study where we had detailed fire regime information due to the limitations of our radio-tracking tower array. This area lacked a range of possible fire mosaics, including long unburnt patches. It would also be helpful to know how fire history affects the availability of food for Gouldian finches during the wet season when they are thought to rely on the sequential seeding of several grass species (Dostine et al. 2001). Such a study would require overcoming the difficulty of catching finches in the wet season when water is plentiful and a more extensive array of receivers to detect more extensive movements.
It would be desirable to study fire and finch movements in different regions of tropical savanna. Our results may not be universally applicable across the entire Gouldian finch distribution, including some areas lacking annual Sorghum. While fire is pervasive in much of northern Australia, scale, timing, intensity, and management vary geographically. For example, fire regimes in areas occupied by Gouldian and other granivorous finches range from relatively infrequent in many parts of north-eastern Australia to very frequent in the northwest (Franklin et al. 1999). Work is still needed to investigate spatial and temporal aspects of different fire regimes on finch movement and space use at broad regional scales before a comprehensive understanding of finch meta-population dynamics with respect to climatic and disturbance (fire and grazing) responses can be gained.
Passive VHF-telemetry is a novel methodology generally used to determine migrating birds’ arrival and departure times. The value of the technique in the current study was its ability to locate multiple finches simultaneously and continuously over a fairly extended period of time, as they moved over a broad area of tropical savanna. This enabled the generation of fine-scale utilisation distribution models from the point data, the spatial extent of which was overlaid with the fire history information. The technology was relatively easy to deploy and required little attendance once the birds were tagged and released. The receivers were fireproofed, enabling us to follow the movements of the finches during and after a wildfire had swept through the area. The technology offers ecologists and managers a new tool for tracking and assessing site utilisation and habitat usage in animals that are too small-bodied to carry satellite devices and too mobile to be accurately located through active tracking techniques and will be invaluable in better understanding how finches respond to fire and other sources of habitat change.
Management Implications
Current land and fire management varies across the Australian savanna. While fire management is largely determined by the proximity of human populations and local production industries, it should not overlook ecological impacts, particularly where savannas are managed for conservation. Ecological benefits of prescribed early dry season burnt savanna has been demonstrated here in terms of preferential finch foraging patterns but have been previously highlighted regarding finch nest box occupancy (Weier et al. 2016), grass seed availability (Weier et al. 2018, 2019) and retention of high-density nesting habitat (Radford et al. 2021). Areas burnt in patchy, low-intensity prescribed fires have greater temporal and spatial availability of annual and perennial grass seed throughout the year than extensively burnt habitats. This is because the patchy application of prescribed burning allows retention of both recently burnt and longer unburnt vegetation (Dostine et al. 2001; Dostine and Franklin 2002; Weier et al. 2018, 2019), which in turn enhances both annual and perennial grass seed availability and is thus associated with increased breeding success and body condition/stress levels for Gouldian finches throughout the year (Legge et al. 2015; Weier et al. 2016). We recommend that land managers use a mosaic of patchy, low-intensity burns in 20–30% of known breeding habitat annually to achieve both protection against extensive wildfires. This will improve grass seed resource availability and heterogeneity (both dry season annuals and wet season perennial grass seeds) across the landscape. Implementation of 20–30% annual prescribed burning will allow individual patches to be burnt no more frequently than once every 3years in line with greater Gouldian breeding success being achieved in recently but infrequently burnt savannas (Weier et al. 2016). We recommend limited strategic use of wet season burning to avoid burning large areas of annual Sorghum before it has set seed in March/April, thereby reducing Sorghum seed availability in the following dry season. A network of early dry season burns would reduce the overall within-site fire frequency whilst still maintaining recently burnt areas and patches of long unburnt vegetation to retain flowering and seeding perennial grasses, including the slow-maturing Triodia spp. (Dostine et al. 2001). Such a fire regime will help ensure the landscape is not dominated by high-frequency, extensive wildfires with all their negative consequences for biodiversity in northern Australia (Franklin et al. 1999; Andersen et al. 2005; Woinarski and Legge 2013; Lawes et al. 2015; Radford et al. 2020).
Data availability
The datasets generated during and/or analysed during the current study are available in the Dryad Data Repository.
Declaration of funding
The research was funded by an Australian Research Council (LP1601101716) grant, with Save the Gouldian Fund, Western Australia Department of Biodiversity, Conservation and Attractions, and WWF-Australia as linkage partners. Charles Darwin University provided additional support and S.J.C. was also supported by the Australian Government Research Training Program Stipend Scholarship. This study was undertaken under scientific permits from the Western Australian Department of Parks and Wildlife (permit no. 08-001715-2), Western Australian Department of Biodiversity Conservation and Attractions (permit no. FO25000021), Parks and Wildlife Commission of the Northern Territory (permit no. 61442), the Australian Bird and Bat Banding Scheme (project no. 545–9), and the Charles Darwin University Animal Ethics Committee (AEC no. A17015).
Author contributions
S.J.C., conceptualisation, formal analysis, data curation, investigation, methodology, writing – original draft, review, editing; I.J.R., conceptualisation, methodology, supervision; writing – review; M.K., formal analysis, writing – review; T.L.C., conceptualisation, formal analysis, data curation, investigation, methodology, writing – review; S.T.G., conceptualisation, supervision, writing – review; H.A.C., conceptualisation, funding acquisition, investigation, methodology, resources, supervision, writing – original draft & review.
Acknowledgements
We thank the Balanggarra Aboriginal Corporation for access to their native title land and other landowners who allowed us access to the study sites so that this research could take place. The Balanggarra Aboriginal Corporation and Balanggarra Rangers undertook prescribed burning. We are also thankful for field assistance from Mark Conboy and Alexander Watson (WWF-Australia), and additional training provided by Jan Lewis and John Rawsthorne. Assistance with construction of radio towers was provided by David Loewensteiner, Matthew Northwood, Tom Rasmussen, Ellie Boyle, Michael Brand, Rebecca Rogers, Brittany Hayward-Brown, Richard Fairman, Vanessa Solano and Balanggarra Rangers. We also thank Jeremy Garnett of Top End Editing for proofreading.
References
Andersen AN (2021) Faunal responses to fire in Australian tropical savannas: insights from field experiments and their lessons for conservation management. Diversity and Distributions 27, 828-843.
| Crossref | Google Scholar |
Andersen AN, Cook GD, Corbett LK, Douglas MM, Eager RW, Russell-Smith J, Setterfield SA, Williams RJ, Woinarski JCZ (2005) Fire frequency and biodiversity conservation in Australian tropical savannas: implications from the Kapalga fire experiment. Austral Ecology 30, 155-167.
| Crossref | Google Scholar |
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecology 26, 32-46.
| Crossref | Google Scholar |
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a world without fire. New Phytologist 165, 525-538.
| Crossref | Google Scholar |
Brazill-Boast J, van Rooij E, Pryke SR, Griffith SC (2011) Interference from Long-tailed Finches constrains reproduction in the endangered Gouldian Finch. Journal of Animal Ecology 80, 39-48.
| Crossref | Google Scholar |
Brazill-Boast J, Pryke SR, Griffith SC (2013) Provisioning habitat with custom-designed nest-boxes increases reproductive success in an endangered finch. Austral Ecology 38, 405-412.
| Crossref | Google Scholar |
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Machler M, Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. The R Journal 9, 378-400.
| Crossref | Google Scholar |
Bureau of Meteorology (2020) Daily rainfall - Wyndham. (Australian Bureau of Meteorology) Available at http://www.bom.gov.au/jsp/ncc/cdio/weatherData/av?p_nccObsCode=136&p_display_type=dailyDataFile&p_startYear=2019&p_c=-264465&p_stn_num=001013
Calenge C (2006) The package ‘adehabitat’ for the R software: A tool for the analysis of space and habitat use by animals. Ecological Modelling 197, 516-519.
| Crossref | Google Scholar |
Clarke KR, Gorley RN (2015) Getting started with PRIMER v7 Plymouth Routines In Multivariate Ecological Research. (PRIMER-E) Available at www.primer-e.com
Collett SJ, Crewe TL, Radford IJ, Garnett ST, Campbell HA (2022a) Sympatric finches increase trophic and spatiotemporal partitioning to enable coexistence through periods of low resource availability. Functional Ecology 36, 2138-2151.
| Crossref | Google Scholar |
Collett SJ, Crewe TL, Radford IJ, Garnett ST, Campbell HA (2022b) Sympatric finches differ in visitation patterns to watering holes: implications for site-focused bird counts. Emu - Austral Ornithology 122, 269-275.
| Crossref | Google Scholar |
Conrad O, Bechtel B, Bock M, Dietrich H, Fischer E, Gerlitz L, Wehberg J, Wichmann V, Böhner J (2015) System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geoscientific Model Development Discussion 8, 2271-2312.
| Crossref | Google Scholar |
Crewe TL, Crysler ZJ, Taylor PD (2018) ‘Motus R Book: a walk through the use of R for Motus Automated Radio-telemetry Data.’ (Bird Studies Canada: Port Rowam, ON) Available at https://raw.githubusercontent.com/MotusWTS/motus/master/inst/extdata/MotusRBook2018-01.pdf
Dostine PL, Franklin DC (2002) A comparison of the diet of three finch species in the Yinberrie Hills area, Northern Territory. Emu - Austral Ornithology 102, 159-164.
| Crossref | Google Scholar |
Dostine PL, Johnson GC, Franklin DC, Zhang Y, Hempel C (2001) Seasonal use of savanna landscapes by the Gouldian finch, Erythrura gouldiae, in the Yinberrie Hills area, Northern Territory. Wildlife Research 28, 445-458.
| Crossref | Google Scholar |
Franklin DC (1999) Evidence of disarray amongst granivorous bird assemblages in the savannas of northern Australia, a region of sparse human settlement. Biological Conservation 90, 53-68.
| Crossref | Google Scholar |
Franklin DC, Burbidge AH, Dostine PL (1999) The harvest of wild birds for aviculture: an historical perspective on finch trapping in the Kimberley with special emphasis on the Gouldian Finch. Australian Zoologist 31, 92-109.
| Crossref | Google Scholar |
Franklin DC, Whitehead PJ, Pardon G, Matthews J, McMahon P, McIntyre D (2005) Geographic patterns and correlates of the decline of granivorous birds in northern Australia. Wildlife Research 32, 399-408.
| Crossref | Google Scholar |
Franklin DC, Legge S, Skroblin A, Heathcote J, Maute K, Schaefer DJ, Garnett ST (2017) Wings of tropical finches: interspecific differences in shape are consistent with levels of mobility, but moult and feather fault patterns are more complex. Emu - Austral Ornithology 117, 370-381.
| Crossref | Google Scholar |
Johnson CN (2006) ‘Australia’s Mammal Extinctions: A 50,000-Year History.’ (Cambridge University Press: Cambridge, UK) Available at https://books.google.com.au/books?hl=en&lr=&id=gcmfAgQcXDMC&oi=fnd&pg=PR7&dq=johnson+2006+fire+refuge&ots=peFWfQzwv1&sig=o3Qk3MEkUpyCUH2vcxoN49TIl2k#v=onepage&q&f=false
Lawes MJ, Murphy BP, Fisher A, Woinarski JCZ, Edwards AC, Russell-Smith J (2015) Small mammals decline with increasing fire extent in northern Australia: evidence from long-term monitoring in Kakadu National Park. International Journal of Wildland Fire 24, 712-722.
| Crossref | Google Scholar |
Legge SM, Garnett ST, Maute KL, Heathcote J, Murphy S, Woinarski JCZ, Astheimer L (2015) A Landscape-Scale, Applied Fire Management Experiment Promotes Recovery of a Population of the Threatened Gouldian Finch, Erythrura gouldiae, in Australia’s Tropical Savannas. PLoS One 10, e0137997.
| Crossref | Google Scholar |
Maute KL, French K, Legge S, Astheimer L (2013) Seasonal stress physiology and body condition differ among co-occurring tropical finch species. Journal of Comparative Physiology B 183, 1023-1037.
| Crossref | Google Scholar |
Pausas JG, Parr CL (2018) Towards an understanding of the evolutionary role of fire in animals. Evolutionary Ecology 32, 113-125.
| Crossref | Google Scholar |
Pausas JG, Ribeiro E (2017) Fire and plant diversity at the global scale. Global Ecology and Biogeography 26, 889-897.
| Crossref | Google Scholar |
Price O, Russell-Smith J, Edwards A (2003) Fine-scale patchiness of different fir intensities in sandstone heath vegetation in northern Australia The fire regimes of New South Wales, Australia, and their ecological effects View project. International Journal of Wildland Fire 12, 227-236.
| Crossref | Google Scholar |
Price OF, Russell-Smith J, Watt F (2012) The influence of prescribed fire on the extent of wildfire in savanna landscapes of western Arnhem Land, Australia. International Journal of Wildland Fire 21, 297-305.
| Crossref | Google Scholar |
Radford IJ, Woolley L-A, Corey B, Vigilante T, Hatherley E, Fairman R, Carnes K, Start AN (2020) Prescribed burning benefits threatened mammals in northern Australia. Biodiversity and Conservation 2020 29(9), 2985-3007.
| Crossref | Google Scholar |
Radford IJ, Oliveira SLJ, Byrne B, Woolley L-A (2021) Tree hollow densities reduced by frequent late dry-season wildfires in threatened Gouldian finch (Erythrura gouldiae) breeding habitat. Wildlife Research 48, 511-520.
| Crossref | Google Scholar |
Rappole JH, Tipton AR (1991) New Harness Design for Attachment of Radio Transmitters to Small Passerines. Journal of Field Ornithology 62, 335-337.
| Google Scholar |
Reside AE, VanDerWal J, Kutt A, Watson I, Williams S (2012) Fire regime shifts affect bird species distributions. Diversity and Distributions 18, 213-225.
| Crossref | Google Scholar |
Sankaran M, Ratnam J, Hanan N (2008) Woody cover in African savannas: the role of resources, fire and herbivory. Global Ecology and Biogeography 17, 236-245.
| Crossref | Google Scholar |
Stobo-Wilson AM, Stokeld D, Einoder LD, Davies HF, Fisher A, Hill BM, Mahney T, Murphy BP, Scroggie MP, Stevens A, Woinarski JCZ, Gillespie GR, Bawinanga Rangers, Warddeken Rangers (2020) Bottom-up and top-down processes influence contemporary patterns of mammal species richness in Australia’s monsoonal tropics. Biological Conservation 247, 108638.
| Crossref | Google Scholar |
Sutherland EF, Dickman CR (1999) Mechanisms of recovery after fire by rodents in the Australian environment: a review. Wildlife Research 26, 405-419.
| Crossref | Google Scholar |
Tidemann SC, Mcorist S, Woinarski JCZ, Freeland WJ (1992) PARASITISM OF WILD GOULDIAN FINCHES (ERYTHRURA GOULDIAE) BY THE AIR-SAC MITE STERNOSTOMA TRACHEACOLUM. Journal of Wildlife Diseases 28, 80-84.
| Crossref | Google Scholar |
Weier A, Radford IJ, Oliveira SLJ, Lawes MJ (2016) Recently but infrequently burnt breeding sites are favoured by threatened Gouldian finches (Erythrura gouldiae. International Journal of Wildland Fire 25, 1281-1290.
| Crossref | Google Scholar |
Weier A, Radford IJ, Manson A, Durrans LJ, Lawes MJ (2017) Frequent fires reduce the nutritional quality of Sorghum stipoideum seed, a keystone food resource for the Gouldian finch (Erythrura gouldiae). The Rangeland Journal 39, 105-112.
| Crossref | Google Scholar |
Weier A, Radford IJ, Woolley L-A, Lawes MJ (2018) Fire regime effects on annual grass seeds as food for threatened grass-finch. Fire Ecology 14, 8.
| Crossref | Google Scholar |
Weier A, Radford IJ, Bellairs SM, Lawes MJ (2019) Seed set in Sorghum stipoideum, and not fire, determines the timing of breeding by Gouldian finches (Erythrura gouldiae). Austral Ecology 44, 151-162.
| Crossref | Google Scholar |
Williams RJR, Lane AM (1999) Wet season burning as a fuel management tool in wet-dry tropical savannas: applications at Ranger Mine, Northern Territory, Australia. In ‘People and rangelands: building the Future. Proceedings of the VI International Rangeland Congress’. (Eds D Eldridge, D Freudenberger) pp. 972–977. (International Rangeland Congress, Inc.) Available at https://publications.csiro.au/rpr/pub?list=BRO&pid=procite:1bb51b9a-2fb3-459e-96f4-625d6c83d81e
Williams RJ, Woinarski JCZ, Andersen AN (2003) Fire experiments in northern Australia: contributions to ecological understanding and biodiversity conservation in tropical savannas. International Journal of Wildland Fire 12, 391-402.
| Crossref | Google Scholar |
Williams PA, McMahon KB, Collins EA, Grice TC, Mack AD, Marshall LE, Owbridge A EF (2020) Black-throated Finch habitat values promoted by patchy fire. North Queensland. Naturalist 50, 38-43.
| Crossref | Google Scholar |
Woinarski JCZ, Legge S (2013) The impacts of fire on birds in Australia’s tropical savannas. Emu - Austral Ornithology 113, 319-352.
| Crossref | Google Scholar |
Woinarski JCZ, Williams RJ, Price O, Rankmore B (2005) Landscapes without boundaries: wildlife and their environments in northern Australia. Wildlife Research 32, 377-388.
| Crossref | Google Scholar |
Woinarski JCZ, Legge S, Fitzsimons JA, Traill BJ, Burbidge AA, Fisher A, Firth RSC, Gordon IJ, Griffiths AD, Johnson CN, McKenzie NL, Palmer C, Radford I, Rankmore B, Ritchie EG, Ward S, Ziembicki M (2011) The disappearing mammal fauna of northern Australia: context, cause and response. Conservation Letters 4, 192-201.
| Crossref | Google Scholar |
Ziembicki MR, Woinarski JCZ, Webb JK, Vanderduys EP, Tuft K, Smith J, Ritchie EG, Reardon TB, Radford IJ, Preece N, Perry JJ, Murphy BP, McGregor HW, Legge SM, Leahy L, Lawes MJ, Kanowski J, Johnson CN, James A, Griffiths AD, Gillespie GR, Frank ASKK, Fisher A, Burbidge AA (2015) Stemming the tide: progress towards resolving the causes of decline and implementing management responses for the disappearing mammal fauna of northern Australia. Therya 6, 169-225 Available at http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-33642015000100169&lng=es&nrm=iso&tlng=en.
| Google Scholar |