

Anthropogenic fire practices only ‘best’ if they promote a large seed buildup: comment on the conservation needs of a fire-killed grevillea
Byron B. Lamont A *
A *
A Ecology Section, School of Molecular and Life Sciences, Curtin University, PO Box U1987, Perth, WA 6845, Australia.
International Journal of Wildland Fire 31(8) 816-820 https://doi.org/10.1071/WF21160
Submitted: 9 November 2021 Accepted: 21 June 2022 Published: 20 July 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF.
Abstract
Fire regimes control the population dynamics of fireprone plant species. Here, I re-analyse the data acquired in a study of the conservation requirements of a rare, fire-killed grevillea. I conclude that specific knowledge is required on the rate at which seeds accumulate in the soil, and the effect of fire frequency on population dynamics for a comprehensive understanding of the conservation biology of this species. Modelling indicates that fires at intervals in the order of multiple decades and covering at least 60% of the ground surface are optimal for this species, independently of the type of fire employed.
Keywords: fire frequency, fire intensity, Grevillea, indigenous fire management, obligate seeder, population modelling, rare plant species, wildfire.
Introduction
McKemey et al. (2021) compared the outcomes on population dynamics of a rare plant, the Backwater grevillea (Grevillea scortechinii subsp. sarmentosa), with different types of fire. The fire (1) undertaken by Indigenous rangers tasked to use traditional fire-management practices, was compared with (2) an unburnt control, (3) a ‘hazard-reduction’ burn by contemporary fire managers, and (4) a ‘wildfire’ that passed over both (1) and (2) 4 years later. From this, the authors concluded that ‘cultural burning was the best fire management for conservation of the Backwater grevillea’ (noted four times). Although I applaud the objectives of this study, I question its ability to give a mechanistic explanation for understanding population dynamics, and to address other relevant management issues, especially fire frequency. I do this by re-analysing their data, noting what population traits need to be monitored to provide informed decisions, and providing a preliminary model of the type of dynamics that can be expected to occur as a guide for future management where species conservation is the goal. The conservation requirements of targeted species, as outlined for this grevillea, highlight the challenges facing managers in terms of the need to understand their particular population demographics for effective conservation management (Enright et al. 2014; Lamont and Witkowski 2021).
Results and discussion
The initial cultural burn consumed ~60% of the ground cover and killed 40% of the pre-existing plants, but resulted in a net 7.6 times increase in population size by the next year, due to the abundance of postfire seedlings (Fig. 1). Two years later, the population had fallen to 5.5 times the prefire level due to seedling death (reasons not given). Inflorescence/fruit production by the postburn plants was 30% of the control plants at this time due to reduced flowering of the surviving plants. The ‘hazard-reduction’ burn cleared ~80% of the groundcover but its impact on the grevillea was not assessed, reducing its value as an instructive comparison among alternative management practices. The ‘wildfire’ 4 years and 3 months after the cultural burn cleared 100% of the ground cover and resulted in incineration of all pre-existing plants, including the controls. The seedling count 6 months after the wildfire was nine times higher among the (reburnt) cultural plots and double that (~18 times higher) among the (burnt) control plots (Fig. 1).

|
Although the population traits measured were invaluable, several key aspects were missed that are required for informed decisions about the effect of the various fires on the conservation status of the species. Postfire numbers were declining steadily with time and, in the absence of further monitoring, were on track to take the population to prefire levels 6 years after the burn (had the wildfire not occurred). Annual monitoring might have shown that most of this decrease was in the first year, as is often the pattern for fire-killed species dependent on post-fire seedling recruitment (Enright and Lamont 1989; Lamont and Groom 1998), so that a negative exponential trend would have reduced the rate of decline. In its absence, the decrease appears to be linear, at least for the first few years, and is accepted here for the tentative extrapolation purposes. Among grevilleas, all seeds are soil-stored and sampling would be needed to assess the rate at which this occurs. Ideally, this includes quantifying annual seed set (viability, granivory – pre- and post-release) and arrival in the soil (to what extent ant dispersers are involved and effective would be of conservation relevance), fate of soil-stored seeds (viability, granivory) and what fraction of the seed store is released from dormancy versus the type of fire (Keith 1997).
However, an idea of seed accumulation can still be gleaned from the data of McKemey et al. (2021) by using 6-month-old seedlings as a surrogate for the seed store where substantial seedling death would not yet have occurred (Fig. 2). A reviewer noted that seedlings may have been differently exposed to drought and herbivores following the August and December fires, thus invalidating this approach. I consulted long-term records for a weather station in New England (bom.gov.au/climate/averages/tables/cw_056056.shtml) and calculated 442 mm in the 6 months after August and 436 mm after December, so there is no obvious rainfall bias there. I checked the recovery plans for three Grevillea species (G. obtusiflora, G. rivularis and G. calliantha), and the only mention of herbivory as a threatening process in any of them was a note that ‘feral deer are present in the area’ so even this was not a direct observation. Herbivory can indeed be fitful across the landscape (Lamont and Groom 1998) but it seems non-critical among grevilleas. Nevertheless, its possible impact is now noted in Fig. 1. Had a comprehensive population dynamic approach as I advocate here been adopted by McKemey et al. (2021), there would have been no need for conjecture on these points.

|
When adjusted for the presence of mature plants as a source of seeds over the period 30 and 34 years since the last fire, these show that the unburnt control for the cultural burn resulted in a small (20%) increase in seed store compared with the cultural burn 4 years before. This was due either to (1) the unburnt plants being more productive (it was noted that flowering of plants that survived the cultural burn was suppressed subsequently for unclear reasons), and/or (2) the much ‘hotter’ wildfire causing more extensive dormancy release (the net effect of higher temperatures at the surface causing viability loss versus beneficial heat penetrating further into the soil profile where seeds resided, Auld and O’Connell 1991), and/or (3) simply a sampling error effect. Although only three data points are available (the first point is no seeds initially), seed accumulation appears to be rising steadily in this species with an apparent doubling in numbers between 17 and 34 years. Population senescence is clearly decades away.
The results here, plus known population dynamic principles, can be used to model the likely scenarios operating in the McKemey et al. (2021) study (Fig. 3). Adult plants are killed in proportion to the area of ground burnt. Seeds germinate in proportion to the area of ground burnt, but the more seedlings, the greater is intraspecific competition such that initial seedling densities reach an asymptote, and may even start to decline via density-dependent thinning (Lamont et al. 2022). Putting these two demographic variables together results in a broad-peaked initial population size. The amplitude of this peak depends on time since the last fire: as long as maximum seed store has yet to be reached, the longer time since the last fire, the greater is the seed store. Cultural and hazard-reduction burns are usually only practicable at moderately short fire intervals, or at longer intervals soon after rain, because of the steady accumulation of fuel that controls fire intensity. Both are labour-intensive and a logistic challenge, and might explain why only 4 ha was undertaken in the cultural burn – whereas the hazard-reduction burn was twice the size as planned. Doubling the fire interval will at least double the seed store, within the 34-year interval relevant here (Fig. 2).
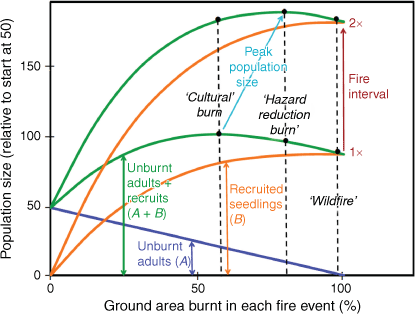
|
Once 50% of the ground flora is burnt the number of seedlings produced varies little with subsequent levels of area burned, and type of patchiness, up to 100% burnt. The key to population recovery then becomes time since the last fire (Fig. 3). Because subsequent attrition of young plants is substantial (Fig. 1), the logical focus is to get that initial number up as high as possible (Keith 1997). The task for fire managers then is to resist the temptation to implement low-intensity cultural or hazard-reduction burns at frequent intervals. It is clear that the grassy savanna model, with frequent (1–5-year intervals) burning of the resprouting vegetation, is not applicable to the forest vegetation under consideration here, instead possessing many fire-killed understorey species (Bradstock et al. 1998). Thus, fire managers should take no action until adult plants have been setting seeds for several decades, and then undertake a fire that burns at least 60% of the ground cover. The challenge is to execute the burn in a way that does not get out of hand. The traditional custodians have shown here that it may be possible, but it would need to be demonstrated on a broader scale than 4 ha. Monitoring the level of flowering as noted is a start, but what matters is the absolute conversion of flowers to stored seeds.
Population stability through minimising death of adult plants is much more likely with cultural burns, but overall population size remains low. The greatest stability is, of course, not to burn at all over at least the 34-year period considered here (and probably for some decades later) but this cannot contribute to increasing the conservation status of the species. With frequent burns seed accumulation is low, and relevant hazard-reduction concerns shift from ‘people and property’ to plant species survival as, increasingly, recruited plants will have insufficient time to contribute substantially to the seed bank. The ‘vital attribute’, time to maturation, would be another population trait worth adding to the list for converting knowledge of population dynamics from the observational to the mechanistic (Enright et al. 1998). The current data indicate that seedling numbers decline back to prefire numbers, effectively zero, over the 6–7 years following the fire, but without self-replacement because 40% of adult plants were killed by the burn. That is, the population will gradually decline to extreme rarity status within a century if the population is routinely burnt at similarly short intervals. There are ample precedents for this danger in ‘fire-sensitive’ (fire-killed) vegetation (Bradstock et al. 1997, 1998; Russell-Smith et al. 1998; Enright et al. 2014).
This is not to assume that the intention would automatically be to adopt a frequent-burning policy now, although it would be tempting to do so, as justified by the conclusions of McKemey et al. (2021) that light burns are the most beneficial. If so, then the easiest way to ensure patchy, low intensity fires is to burn frequently, although associated approaches include burning in winter–spring rather than autumn (as done here), during mild windless weather, soon after rain and/or in the morning. The point is more that the critical variable, fire frequency, was not addressed in this study. However, there would be no urgency to burn, because the population would remain stable for decades in the absence of fire, and seeds would continue to accumulate in the soil, according to Fig. 2.
In contrast, delayed fire leads to a marked increase in population size that declines to the prefire level by 15 years or so on the present (quite inadequate) data, but the seed store will be vast. This gives great buffering capacity for surviving even two wildfires at an interval of only a few years, for many seeds may escape the first fire. Admittedly, the population is now even-aged but those ages varied from X to (X + 15) years over that time, and distribution patchiness, as gauged by the 95% confidence interval in Fig. 1, is much greater after the wildfire than for the cultural burn (50% in the current case); these two properties, time and patchiness, can be viewed as enhancing the conservation status of this species by increasing temporal and spatial variability over the landscape (it is not just about numbers). In particular, management fires, interspersed with the inevitable wildfire, can be out-of-phase across the landscape to create further mosaic effects (pyrodiversity; He et al. 2019) that way. Why should the goal of temporal heterogeneity only be applied to within populations?
Conclusions
What we see above ground is only part of the picture. The key to the conservation management of this species lies with the size of the seed store underground, even if only annual seed production is quantified as a surrogate for that. Such a task lacks the strong tradition of lighting and controlling fires but no management task could be more vital than monitoring the size of the seed store. Speaking proverbially, the answer lies in the soil. Above all, initiating fires should not be seen as an urgent task; time is on your side. For this grevillea, the seed store was still rising steadily from 30 to 34 years so the population is not in danger of senescing, and the traditional custodians have shown here that it is possible to undertake ‘safe’ burns even 30 years since the last fire. Perhaps the fuel load had even reduced somewhat, short-lived shrubs having long since senesced and decomposed (Zylstra 2018), over what would usually be regarded by fire managers as an extremely long period? And hazard-reduction fires may fail their purpose in the face of intense wildfires (Bradstock et al. 1998) as noted in McKemey et al. (2021). Other solutions may be needed to address the challenge of species conservation, where fires that burn 80% of the ground cover at intervals in the order of multiple decades and out-of-phase over the landscape are shown (in the case of this grevillea) to be optimal, versus the need to safeguard human welfare that such a preferred fire regime might imply.
Data availability
All data used here for Figs 1, 2 were abstracted from McKemey et al. (2021), or were of a hypothetical nature only, as retrievable from Fig. 3.
Conflicts of interest
I declare no conflict of interest.
Declaration of funding
This research was not supported by any non-personal funds.
Acknowledgements
Thanks for comments on a draft to Neal Enright, who declined to be a co-author for fear of an ‘unproductive exchange’, and to the reviewers for their helpful comments on the manuscript. The topic of hazard-reduction burning is clearly in need of further examination.
References
Auld TD, O’Connell MA (1991) Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae. Australian Journal of Ecology 16, 53–70.| Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae.Crossref | GoogleScholarGoogle Scholar |
Bradstock RA, Tozer MG, Keith DA (1997) Effects of high frequency fire on floristic composition and abundance in a fire-prone heathland near Sydney. Australian Journal of Botany 45, 641–655.
| Effects of high frequency fire on floristic composition and abundance in a fire-prone heathland near Sydney.Crossref | GoogleScholarGoogle Scholar |
Bradstock RA, Bedward M, Kenny BJ, Scott J (1998) Spatially-explicit simulation of the effect of prescribed burning on fire regimes and plant extinctions in shrublands typical of south-eastern Australia. Biological Conservation 86, 83–95.
| Spatially-explicit simulation of the effect of prescribed burning on fire regimes and plant extinctions in shrublands typical of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Enright NJ, Lamont BB (1989) Seed banks, fire season, safe sites and seedling recruitment in five co-occurring Banksia species. Journal of Ecology 77, 1111–1122.
| Seed banks, fire season, safe sites and seedling recruitment in five co-occurring Banksia species.Crossref | GoogleScholarGoogle Scholar |
Enright NJ, Marsula R, Lamont BB, Wissel C (1998) The ecological significance of canopy seed storage in fire-prone environments: a model for nonsprouting shrubs. Journal of Ecology 86, 946–959.
| The ecological significance of canopy seed storage in fire-prone environments: a model for nonsprouting shrubs.Crossref | GoogleScholarGoogle Scholar |
Enright NJ, Fontaine JB, Lamont BB, Miller BP, Westcott VC (2014) Resistance and resilience to changing climate and fire regime depend on plant functional traits. Journal of Ecology 102, 1572–1581.
| Resistance and resilience to changing climate and fire regime depend on plant functional traits.Crossref | GoogleScholarGoogle Scholar |
He T, Lamont BB, Pausas JG (2019) Fire as a key driver of Earth’s biodiversity. Biological Reviews 94, 1983–2010.
| Fire as a key driver of Earth’s biodiversity.Crossref | GoogleScholarGoogle Scholar |
Keith DA (1997) Combined effects of heat shock, smoke and darkness on germination of Epacris stuartii Stapf, an endangered fire-prone Australian shrub. Oecologia 112, 340–344.
| Combined effects of heat shock, smoke and darkness on germination of Epacris stuartii Stapf, an endangered fire-prone Australian shrub.Crossref | GoogleScholarGoogle Scholar |
Lamont BB, Groom K (1998) Seed and seedling biology of the woody-fruited Proteaceae. Australian Journal of Botany 46, 387–406.
| Seed and seedling biology of the woody-fruited Proteaceae.Crossref | GoogleScholarGoogle Scholar |
Lamont BB, Witkowski ETF (2021) Plant functional types determine how close postfire seedlings are from their parents in a species-rich shrubland. Annals of Botany 127, 381–395.
| Plant functional types determine how close postfire seedlings are from their parents in a species-rich shrubland.Crossref | GoogleScholarGoogle Scholar |
Lamont BB, Miller BP, Enright NJ, Yan ZG (2022) Isolation and self-regulation processes in simulated postfire microsites promote plant species diversity. Acta Oecologica 114, 103795
| Isolation and self-regulation processes in simulated postfire microsites promote plant species diversity.Crossref | GoogleScholarGoogle Scholar |
McKemey M, The Banbai Rangers Patterson ML, Hunter J, Ridges M, Ens E, Miller C, Castello O, Reid N (2021) Indigenous cultural burning had less impact than wildfire on the threatened Backwater grevillea (Grevillea scortechinii subsp. sarmentosa) while effectively decreasing fuel loads. International Journal of Wildland Fire 30, 745–756.
| Indigenous cultural burning had less impact than wildfire on the threatened Backwater grevillea (Grevillea scortechinii subsp. sarmentosa) while effectively decreasing fuel loads.Crossref | GoogleScholarGoogle Scholar |
Russell-Smith J, Ryan PG, Klessa D, et al. (1998) Fire regimes, fire-sensitive vegetation, and fire management of the sandstone Arnhem Plateau, monsoonal northern Australia. Journal of Applied Ecology 35, 829–846.
| Fire regimes, fire-sensitive vegetation, and fire management of the sandstone Arnhem Plateau, monsoonal northern Australia.Crossref | GoogleScholarGoogle Scholar |
Zylstra PJ (2018) Flammability dynamics in the Australian Alps. Austral Ecology 43, 578–591.
| Flammability dynamics in the Australian Alps.Crossref | GoogleScholarGoogle Scholar |