

Establishment of native and exotic grasses on mine overburden and topsoil in the Hunter Valley, New South Wales
C. H. A. Huxtable A D , T. B. Koen B and D. Waterhouse A CA Department of Natural Resources (DNR), PO Box 2185 Dangar, NSW 2309, Australia.
B DNR, PO Box 445, Cowra, NSW 2794, Australia.
C Current address: Plant Breeder’s Rights, Department of Agriculture, Fisheries and Forestry Australia, PO Box 858 Canberra, ACT 2601, Australia.
D Corresponding author. Email: Charles.Huxtable@dipnr.nsw.gov.au
The Rangeland Journal 27(2) 73-88 https://doi.org/10.1071/RJ05006
Submitted: 30 July 2004 Accepted: 15 October 2004 Published: 21 November 2005
Abstract
Native grasses have an important role to play in mine rehabilitation throughout Australia, but there have been few scientifically designed studies of field establishment of native grasses from sown seed in this country. Current recommendations for rehabilitation of open-cut coal mines in the Hunter Valley involve the sowing of exotic pasture species to reinstate mined land to Class IV and V under the Rural Land Capability System. Despite the importance of native grasses in the pre-mined landscape, they are currently not widely included in mine rehabilitation. To address this issue a project was conducted between 1994 and 2000 to research the use of native grasses for rehabilitation of open-cut coal mines in the Hunter Valley. This paper reports on 2 mine site experiments that aimed to assess establishment and persistence of a broad range of native and exotic grass species from an autumn sowing in both topsoil and raw spoil over a period of 61 months. The most promising natives in terms of early establishment, persistence and spread over time, included six C3 accessions (five Austrodanthonia spp. and Austrostipa bigeniculata) and one C4 accession (Cynodon dactylon). Persistence of these accessions was better in raw spoil than topsoil, despite initial low numbers, due to a lack of weed competition and their ability to spread by self-seeding. In topsoil, and in the absence of any biomass reduction, native species were mostly out-competed by vigorous exotic perennial grasses which were sown in these experiments and from seed influx from adjacent rehabilitation areas or from the soil seed bank. The effects of climatic conditions and differences in soil physical, chemical and seed bank characteristics at the 2 mine sites are also discussed.
Additional keywords: germination, rehabilitation, weeds.
Introduction
Native grasses have an important role to play in mine rehabilitation throughout Australia and their past use has been reviewed by Huxtable (2000) and Langkamp and Plaisted (1987). The main ways that native grasses can be established on minesites is from sowing seed or from seed present in the topsoil, which is spread on a rehabilitation area. This topsoil seed bank can be the most important source of native seed, especially where a high species diversity is required as part of the rehabilitation program (Bellairs 1996). At Tarong minesite in south-east Queensland, 18–68% of the total seed bank was found to consist of native grass seed, including that of Oplismenus aemulus1 and Themeda australis (Bellairs 1996). At Mt Isa in Queensland, seed of four species of native grasses were found in the stockpiled topsoil at a minesite (Barrs et al. 1996). Conversely, rehabilitation with native species can be made extremely difficult if the seed bank contains many weed seeds. In comparative experiments in the Hunter Valley, a good cover of a range of native grasses and forbs was established at one minesite using topsoil freshly stripped from an area of native pasture, while at another minesite where stockpiled topsoil from a fertilised improved pasture was used, there was extensive weed establishment but very few native species established (Huxtable 1999).
There are relatively few examples of sowing seed or seed bearing material. Exceptions to this include the use of Themeda australis in gold mine rehabilitation (Windsor et al. 2000) and the use of Spinifex sericeus for dune rehabilitation following sand mining (Barr 1965). To the author’s knowledge, there have been no scientifically designed studies in Australia which test field establishment from sown seed of a broad range of native grasses for mine rehabilitation purposes under natural climatic conditions.
Recent figures show that there has been about 10 000 ha of land disturbed by open-cut coal mining in the Hunter Valley of New South Wales, of which 3000 ha had been rehabilitated (NSW Department of Mineral Resources 1999). This mined area represents only 3.8% of land in the Hunter coalfields region (covered by eight 1 : 25 000 map sheets) and is small relative to the area used for agriculture. Despite this, mining activities have had a major environmental impact on surrounding areas for which the community has held the mining companies accountable.
Rehabilitation goals in the Hunter Valley have traditionally been based on the Rural Land Capability System (Emery 1985). Using this system, most pre-mined land in the Hunter is classed as Class IV and V, i.e. suitable for grazing but not for cultivation. There is a government requirement that, following mining, land is rehabilitated with pasture to these standards or better (Hannan 1995). Current recommendations for Hunter Valley coal mines are based on the use of a mix of exotic species including Chloris gayana, Panicum maximum, Medicago sativa and Medicago truncatula (Hannan 1995). While these species have been highly successful in providing rapid, effective surface stabilisation, they tend towards a dominance of C. gayana (Rhodes grass) when not grazed, often require high inputs of maintenance fertiliser and have low biodiversity value (Marschke et al. 1994). In addition to exotic pastures, smaller areas have been hand planted or direct seeded with native trees and this area is expected to increase (NSW Department of Mineral Resources 1999). In addition, the use of native trees in a more landscape-oriented approach to rehabilitation is being encouraged by the production of a synoptic plan that aims for mining companies to coordinate their rehabilitation to enhance landscape connectivity (NSW Department of Mineral Resources 1999).
Despite the importance of native grasses in the pre-mined landscape, they have not been widely utilised in mine rehabilitation in the Hunter Valley. Hannan (1995) considered slow establishment, poor erosion protection during establishment and low biomass and seed production compared with exotic species as reasons for their non-use. However, early comparisons of the nutritive value of exotic and native grasses for potential use in mine rehabilitation (Dyson et al. 1987) were based on newly sown exotic pasture species compared with old native pasture of unspecified species composition. The validity of such comparisons is questionable as plants of similar ages and stages of growth should be used to give a true comparison, as demonstrated by Jones (1996). A detailed scientific research program had not been undertaken to fully investigate the potential of native grasses in mine rehabilitation. Priorities for rehabilitation have changed since the 1980s, when the original guidelines were developed and increasing importance is now being placed on native species and ecosystems and the benefits of using them (NSW Department of Mineral Resources 1999). Perceived benefits of using native grasses in mine rehabilitation include tolerance of drought, adaptation to low soil fertility and the ability of some species to establish in disturbed environments. The latter could represent substantial savings to the mining company in terms of reduced costs of fertiliser and spreading of topsoil if native grasses could establish in raw spoil. In addition, there is an increasing move towards establishing diverse and self-sustaining native ecosystems on mine sites, which will have a range of aesthetic and biodiversity values in addition to providing surface stabilisation (NSW Department of Mineral Resources 1999).
To address these issues, a 5-year project funded by the Australian Coal Association Research Program (ACARP) commenced in 1994, to research the use of native grasses in open-cut coal mine rehabilitation in the Hunter Valley of New South Wales (Huxtable et al. 1997; Huxtable 1999).
This paper reports on the results of 2 experiments in which the establishment, persistence and dynamics of 24 native and exotic grasses were monitored over a period of 61 months between 1995 and 2000, at 2 Hunter Valley mine sites. The aim of these experiments was to screen a broad range of species for their suitability for rehabilitating spoil dumps produced during open-cut coal mine rehabilitation in the Hunter Valley, with and without the application of topsoil.
Materials and methods
Study sites
The two experimental sites, at Ravensworth South and Howick mines, are situated about 10 km apart and half way between Singleton and Muswellbrook in the upper Hunter Valley. The sites were selected because they were considered typical of rehabilitation sites in the Hunter Valley. Both sites were located on artificial landscapes, formed when spoil (weathered or unweathered rock) from the open-cut mine is relocated and shaped in a designated rehabilitation area. Topsoil, which had been stripped from the area to be mined and stockpiled for up to 10 years, is generally then spread over the site to a depth of between 10 and 20 cm. The site is then generally ripped to a depth of 60 cm.
Average rainfall is summer dominant with an annual total of about 630 mm (Commonwealth Bureau of Meteorology 2003). The region experiences hot summers with an average range of between 17 and 30°C and moderate winters ranging between 5 and 18°C. Geology of the area is part of the Wittingham coal measures, consisting primarily of permian sediments that are predominantly alkaline laid down under fresh water conditions (Marschke et al. 1994). Vegetation in the pre-mined landscape consists of predominantly native pastures with scattered trees, apart from alluvial lands along creeks that often comprise exotic pastures or lucerne. The native grass component is dominated by species in the genera Bothriochloa, Chloris, Aristida and Cymbopogon on the lower fertility soils and Austrodanthonia, Austrostipa, Panicum and Dichanthium on the higher fertility soils.
Measurement of climatic data
Seasonal rainfall and temperature data for the period from January 1995 to December 2000 and 12-year averages were obtained from the meteorological station at Ravensworth South mine.
Analyses of raw spoil and topsoil material at the sites
Before sowing, a composite sample of each raw spoil and each topsoil was collected by bulking material collected from several spots within each of the 3 replicates of topsoil and raw spoil. These samples were tested for pH (in water), electrical conductivity (EC) and extractable phosphorus (P available to plants). Soil samples were also collected in the same manner in March 1999 (48 months after sowing) and analysed for pH, EC and P as well as equivalent cation exchange capacity (eCEC), percentage cations (sodium, potassium, calcium and magnesium), calcium/magnesium ratio and texture. All soil tests were conducted by the Department of Land and Water Conservation (DLWC), a NSW government department, at their soils laboratory at Scone. All soil testing procedures followed those described by Hazelton and Murphy (1992).
Experimental design
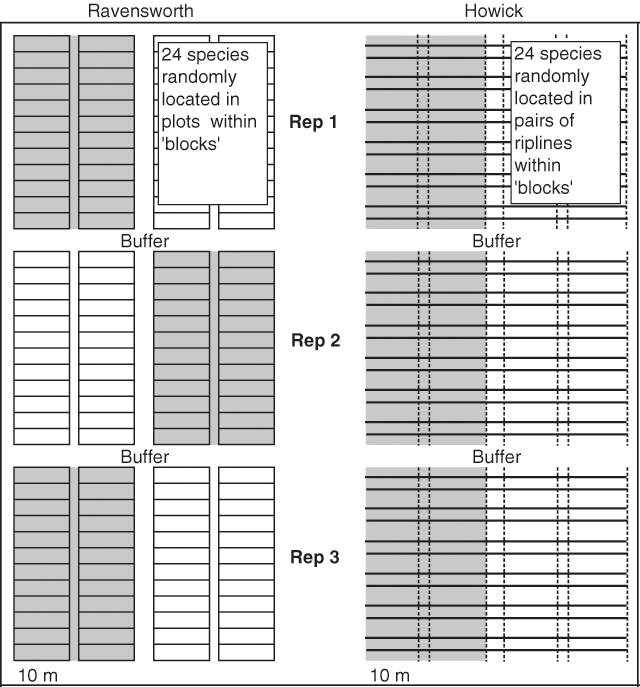
The two experiments had different soil types, plot sizes and dimensions. Because of these differences, they will be treated in this paper as two separate experiments and hereafter referred to as ‘Ravensworth’ and ‘Howick’. Figure 1 shows the experimental layout of the Ravensworth and Howick sites and explains how they differ.
|
Both experiments followed a factorial design of 24 species by 2 soil treatments, replicated 3 times as blocks, to give a total of 144 ‘experimental units’. Both were of a split plot design, the whole plots (blocks) being the soil treatments, ‘raw spoil’ and ‘topsoil’. ‘Raw spoil’ refers to subsoil overburden, consisting of weathered and unweathered rock of various aggregate, ranging from extremely fine clay to small rocks, with no topsoil added. ‘Topsoil’ refers to where topsoil was spread over the raw spoil to a depth of 20 cm. Species were randomly allocated to experimental units within the whole plots. Each whole plot was separated by a 5 m buffer, and experimental units were arranged in two tiers of 12 within each whole plot. Experimental units at Ravensworth consisted of 2 × 10 m plots and were separated by 1 m walkways (Fig. 1). At Howick, experimental units consisted of 10 m sections of deep rip lines arranged in pairs 1 m apart with each pair of rip lines separated by a 2 m walkway.
Species selection, sowing rate and seed preparation
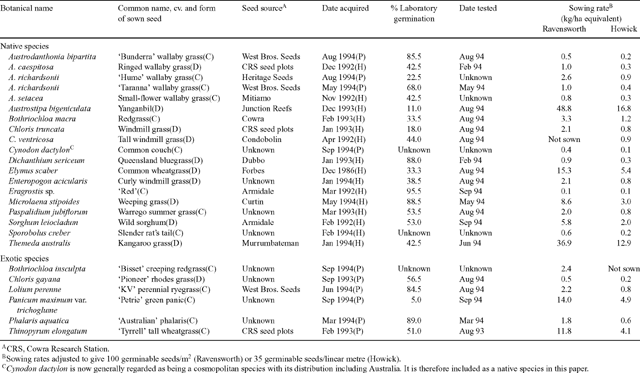
Species were selected on the basis of their potential for mine rehabilitation and seed availability. Seed was collected by hand, brush harvester or obtained from commercial suppliers. Table 1 lists the species, source of seed, percent laboratory germination, date of germination tests, sowing rate and form in which the seed was sown. Seed was sown on the basis of weight of seed required to give 100 germinable seeds per square metre (Ravensworth) or 35 germinable seeds per linear metre (Howick). This was calculated for each species by germination percentage in an incubator (light 30°C 12 h/dark 20°C 12 h) and average seed weight. The reason for the different sowing rates was that experimental units at Ravensworth were much larger (20 m2) than at Howick (10 m linear section of a ripline). It was reasoned that these different sowing rates would closely approximate a similar plant density if seed germinated in the field at rates equivalent to those in the laboratory. To assist ease of sowing, small-seeded species were bulked with sterile phalaris seed. All seed was treated with Ficam® (as Bendiocarb, East Hawthorn, Vic., Australia) at a rate equivalent to 200 g per 100 kg seed, to prevent ant predation.
|
Site preparation and sowing
At Ravensworth, topsoil was spread on designated main plots and the whole site deep ripped in October 1994, with the intention of sowing shortly afterwards. Because of drought conditions however, sowing was postponed until the following autumn. The site was sown during 3–5 April 1995. At Howick, topsoil was spread on designated main plots and the whole site deep ripped to produce furrows on 24 March 1995, seed was sown on 28 March. Seed was mixed with coarse sawdust as a carrier and Starter 15® fertiliser (at 100 kg/ha) and hand broadcast into the experimental units. Plots were then hand raked to cover the seed to a shallow depth (Ravensworth), or covered by tamping down gently onto the loose soil in the furrows (Howick).
Weed control
Because the Ravensworth site had been prepared in October 1994, weed control was necessary before sowing in early April 1995. The site was therefore sprayed in March 1995 with picloram and triclopyr (as Grazon®, Sydney, NSW, Australia) and glyphosate (as Roundup®, Laverton North, Vic., Australia). In addition, both Ravensworth and Howick were sprayed with picloram and triclopyr in October 1995 to control severe infestations of Galenia pubescens at Ravensworth and Hirshfeldia incana at Howick. All sprays were applied at the rate of 2.0 L/ha.
Plant sampling
In both experiments, establishment, survival and recruitment of sown species was measured in terms of number of plants or ground cover in June 1995, July 1996, April 1997 and April 2000 (corresponding to 2, 15, 24 and 61 months, respectively, after sowing).
At Ravensworth, plant counts were made on two 50 × 50 cm quadrats randomly located within each experimental unit. These quadrat count data were averaged and re-expressed as plants/m2. In some situations where rhizomatous and stoloniferous species, or in some cases tussocky species, had formed dense groundcover (which was nearly always 100%), it was impossible to accurately record plant numbers in one or both quadrats. In cases where dense cover existed in only one of the two quadrats, count data from the single quadrat was favoured, and a conservative estimate of plants/m2 formed. In cases where both quadrats had high cover levels, the species concerned was omitted from statistical analysis for that time of measurement.
At Howick in June 1995, July 1996 and April 1997, average plant numbers per metre of row were estimated from plant counts in 2 randomly located 1-m sections within each experimental unit. In April 2000, percentage cover for the entire 10-m experimental unit was estimated.
Presence of sown species outside their designated experimental units at both Ravensworth and Howick was recorded in April 2000. From this was calculated percentage occurrence of each species, which was used as a measure of spread of each species across experimental units at each mine site.
At each sampling, presence of unsown native and exotic species was also recorded for all experimental units at both sites. These data were expressed both as a measure of species richness and as percentage frequency of occurrence of individual species within each site.
Statistical analysis
Establishment and survival data expressed as plants/m2, plants/linear metre or, in the case of Howick in April 2000, % cover/10-m row, were subjected to an analysis of variance (ANOVA) using a split plot model appropriate for the experimental layout. Data were analysed separately for the four observation times of each site since the combination of species differed among times and sites. This was because some species had zero mean plant counts throughout the full experimantal period, others varied in initial establishment and persistence over time and others, on some occasions, were recorded using groundcover rather than plant numbers. Data displayed variances proportional to the mean, and were stabilised by transforming to the scale of natural logarithms. Where soil type, species or their interaction was statistically significant (P < 0.05), means were compared using the least significant difference (l.s.d.) multiple comparison procedure.
Results
Climatic data
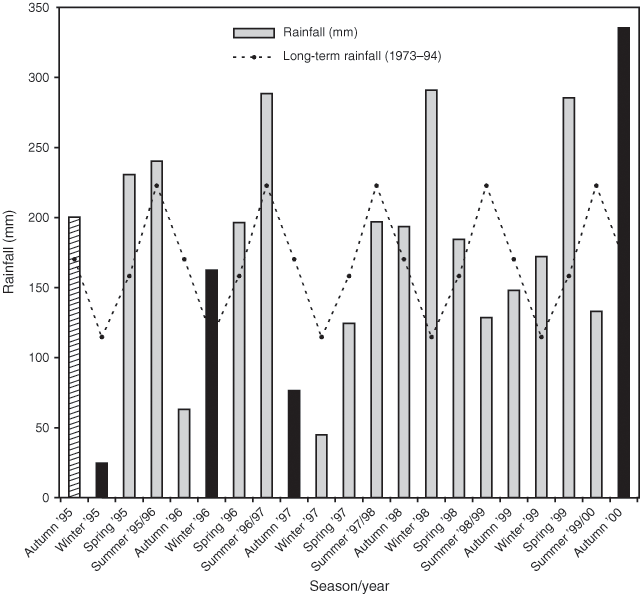
Seasonal rainfall varied considerably from long-term mean figures during the experimental period (Fig. 2). Spring and summer rainfall during the first 24 months of the experiment was higher than the 21-year average. Autumn and winter rainfall during the first 36 months was much lower than average, except for autumn 1995 (when seed was sown) which had 200 mm and winter 1996 which had 163 mm. Winter 1998, spring 1999 and autumn 2000 each had much higher than average rainfall of 280 mm rain and the final three summers of the experimental period each had lower than average falls (Fig. 2).
|
During the experimental period, mean temperatures ranged from 4.8 to 20.5°C in winter and 15 to 34.4°C in summer.
Analyses of raw spoil and topsoil material
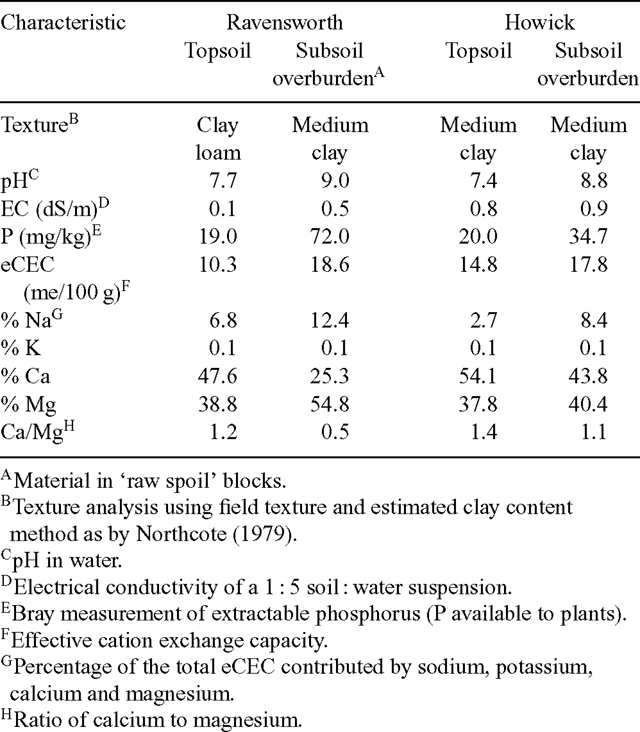
Mean pH and EC levels were similar at the two times of sampling, and P levels only varied minimally (± 0.3 mg/kg). Results from the March 1999 sample are presented (Table 2). All material was classified as medium clay, except the topsoil at Ravensworth, which was clay loam. Both sites had mildly alkaline topsoil and strongly alkaline raw spoil. Topsoil and raw spoil at both sites can be classified as non-saline, although the electrical conductivities (ECs) of raw spoil materials were higher than either topsoil. In addition, topsoil from Howick had higher EC than topsoil from Ravensworth (Table 2). Effective cation exchange capacity (eCEC) was low at Ravensworth topsoil and moderate in all other materials. Calcium/magnesium ratios indicate that all topsoil and raw spoil materials had low calcium levels, with raw spoil at Ravensworth being classified as calcium deficient. Raw spoil material was highly sodic at Ravensworth and marginally sodic at Howick, while topsoil at Ravensworth was marginally sodic and at Howick it was non-sodic. Extractable phosphorus levels were higher in raw spoil than in topsoil, and markedly so at Ravensworth (72 mg/kg).
|
Interpretation of plant data
Measurements of plant numbers or groundcover of sown species had two main components: first, plants that were derived from the seed initially sown and second, plants from subsequent seedling recruitment.
Assessment of sown species in their designated plots
Ravensworth
Of the 24 species sown, nine had nil or extremely low plant numbers or cover at all four times of data collection (Table 3) and are not further included in this section of the results. Two exotic species, Chloris gayana and Bothriochloa insculpta, while not recorded in June 1995, had formed dense groundcover in topsoil units from July 1996 onwards, and C. gayana had formed dense groundcover in raw spoil plots by April 2000 (Table 3). In addition, the native species Cynodon dactylon had high cover levels from April 1997 onwards and Austrostipa bigeniculata had high levels in April 1997. No statistical analyses were undertaken on these species at those times of measurement.
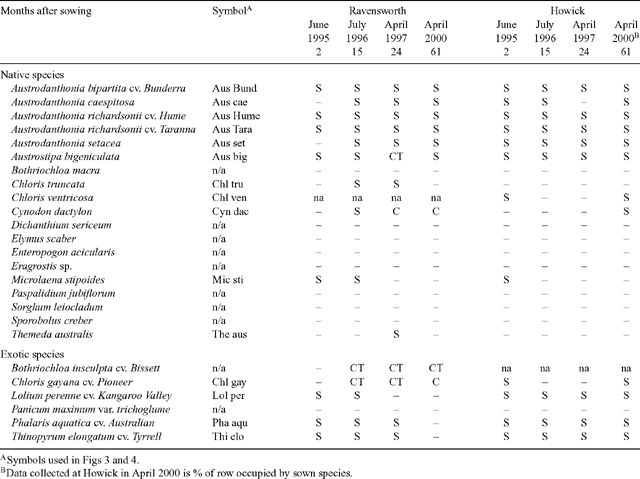
|
Thirteen species had plants recorded in sufficient numbers on at least one occasion to enable statistical analysis and the results are presented in Fig. 3. For ease of comparison, all 13 species are shown for each time of measurement, even when fewer species than this were included in the analysis for any one time. Consequently, species showing zero plant numbers indicate that that species was not included in the analysis at that time of measurement.
|
In general, the most successful native species in terms of initial establishment and persistence, included the 5 accessions of Austrodanthonia, A. bigeniculata and C. dactylon, with mean plant numbers generally fluctuating variously between 5 and 150 plants/m2 during the experimental period (Fig. 3). Microlaena stipoides, C. truncata and Themeda australis had between about 3 and 25 plants/m2, but did not persist in either topsoil or raw spoil through to April 2000. The exotic species Lolium perenne, Phalaris aquatica and Thinopyrum elongatum all had relatively high plant numbers in June 1995 (50–100 plants/m2), but none persisted through to April 2000.
In June 1995, with the exception of ‘Bunderra’ wallaby grass, there were no significant differences (P < 0.05) in plant numbers between topsoil and raw spoil for any one species, whereas in July 1996, all native species and L. perenne had significantly more plants in topsoil than raw spoil (Fig. 3). In April 1997 there were no significant differences (P < 0.05) between soils for any one species, whereas among native species in April 2000 there were significantly greater numbers of plants on raw spoil than in topsoil, while L. perenne, P. aquatica and T. elongatum were not recorded.
Comparisons among species showed that in June 1995, A. bigeniculata had significantly more plants (P < 0.05) than all other native species and did not significantly differ from the 3 exotic species (Fig. 3). Microlaena stipoides had significantly fewer plants than ‘Taranna’ wallaby grass, with ‘Bunderra’ and ‘Hume’ wallaby grasses being intermediate. In July 1996, plant numbers among native species in topsoil did not differ significantly (P < 0.05), with the exception of M. stipoides, which was significantly lower than most of the others. In raw spoil plots at this time, A. bigeniculata had significantly more plants than the other native species, with the exception of ‘Taranna’ wallaby grass, which in turn was significantly higher than ‘Bunderra’ wallaby grass, Austrodanthonia caespitosa, A. setacea and C. truncata (Fig. 3). In July 1996, there were no significant differences (P < 0.05) among the 3 exotic species in either topsoil or raw spoil, except between Lolium perenne in topsoil and both L. perenne and P. aquatica in raw spoil. In April 1997, A. bigeniculata had full cover in some replicates and could not be included in the analysis. At this time, most of the native species did not differ significantly among one another or between soil types, with the exceptions of C. truncata, which was significantly lower than ‘Taranna’ wallaby grass and M. stipoides, which had was not present (Fig. 3). In April 1997, P. aquatica and T. elongatum did not vary significantly from one another in either soil type, or from most native species in raw spoil. In April 2000, there were virtually no significant differences among the 5 accessions of Austrodanthonia and A. bigeniculata, with generally less than 20 plants/m2 persisting in raw spoil units, with fewer or none in topsoil units.
Howick
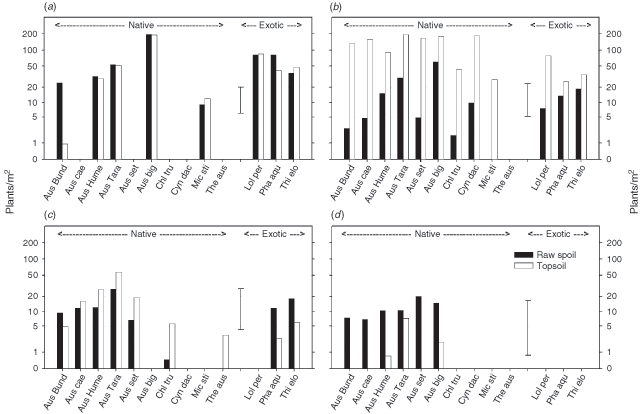
Of the 24 species sown, 11 had nil or extremely low plant numbers or cover at all four times of data collection (Table 3) and are not further included in this section of the results. The remaining 13 species were recorded in sufficient numbers or cover levels on at least one occasion to enable statistical analysis, and the results are presented in Fig. 4. In Fig. 3, all 13 species are included in each histogram, even when fewer species than this were included in the analysis for any one time of measurement.
|
In general, the most successful native species in terms of initial establishment and persistence, included the five accessions of Austrodanthonia and A. bigeniculata, with mean plant numbers generally fluctuating variously between one and 75 plants/linear metre, or cover of between 3 and 83% of the experimental unit (April 2000 only, Fig. 4). Chloris ventricosa and M. stipoides had relatively small mean numbers of plants (<7) in June 1995 and C. ventricosa and C. dactylon had relatively high cover levels in April 2000. All four exotic species persisted until April 2000, and generally ranged between five and 60 plants/linear metre, and between 5 and 60% cover.
In June 1995, there were significant differences (P < 0.05) in plant numbers between topsoil and raw spoil for the native species ‘Bunderra’ wallaby grass, ‘Hume’ wallaby grass, A. bigeniculata and C. ventricosa, and the exotic species C. gayana and P. aquatica (Fig. 4). In July 1996, this trend continued only for A. bigeniculata and P. aquatica, while for all other species there were no significant differences between topsoil and raw spoil. In April 1997, there were no significant differences (P < 0.05) for any of the eight species included in the analysis. In April 2000, all five accessions of Austrodanthonia had higher cover levels in raw spoil than in topsoil, although this effect was only significant (P < 0.05) with ‘Bunderra’, A. caespitosa and ‘Taranna’ accessions. Chloris ventricosa had significantly higher cover on topsoil than in raw spoil, which had zero cover (Fig. 4).
Austrostipa bigeniculata had more plants than all other species in both June 1995 and July 1996, with more than 50 plants/linear metre in topsoil and more than five plants/linear metre in raw spoil (Fig. 4). In June 1995, this effect was significant for all the native species in both topsoil and raw spoil but not significant when compared to the four exotic species. In July 1996, the effect was significant for all native and exotic species in topsoil, with the exception of L. perenne, which had over 50 plants/linear metre in topsoil. In July 1996, L. perenne had significantly more (P < 0.05) plants/linear metre in raw spoil than all other species, with the next highest being T. elongatum and A. bigeniculata, which were not significantly different from one another (Fig. 4). In June 1995, ‘Bunderra’ and ‘Hume wallaby grasses had significantly higher (P < 0.05) numbers of plants/linear metre in topsoil than A. caespitosa and A. setacea with ‘Taranna’ intermediate, while plant numbers did not differ significantly among these accessions in raw spoil. In July 1996, all five accessions of Austrodanthonia had significantly lower plant numbers in raw spoil than T. elongatum but not significantly different to A. bigeniculata, except for A. caespitosa, which was significantly lower. In April 1997, there were no significant differences in plant numbers/linear metre for any species on raw spoil, while ‘Bunderra; and A setacea had significantly lower numbers on topsoil than all other species (Fig. 4). In April 2000, C. dactylon had the highest percentage groundcover of any species on raw spoil, however this effect was not significant, except in comparison to A. setacea, which was significantly lower. On topsoil, C. dactylon had significantly more (P < 0.05) groundcover than the five Austrodanthonia accessions however did not differ significantly from A. bigeniculata and C. ventricosa, which both had over 20% mean cover (Fig. 4). Percentage cover in April 2000 did not differ significantly among the exotic species in either soil type, with the exception of C. gayana, which had significantly lower cover in topsoil than P. aquatica in topsoil. With the exception of A. setacea, percentage groundcover on raw spoil of each native species exceeded that of each exotic species (Fig. 4).
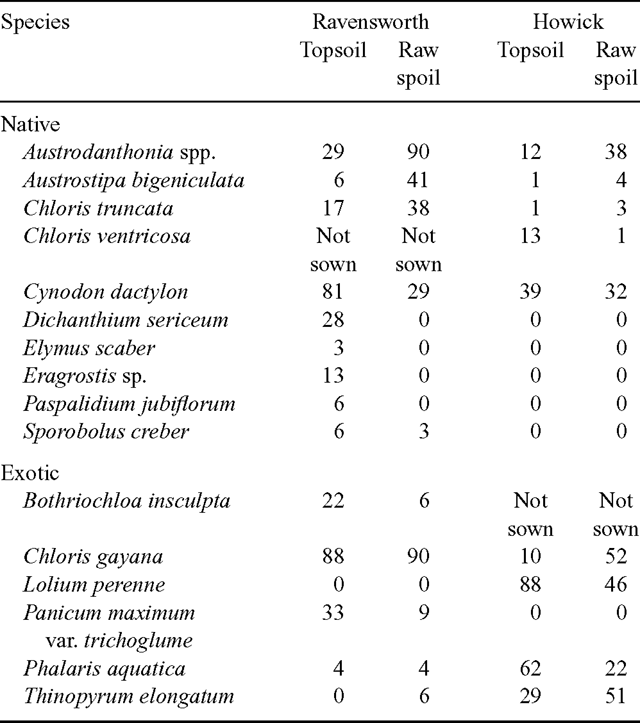
Sown grass species outside experimental units
Occurrence of sown species outside their designated experimental units varied considerably between the two sites and among species and soil treatments (Table 4). Among native species, Austrodanthonia spp.2, A. bigeniculata, C. truncata, C. ventricosa and C. dactylon had the greatest spread at one or both sites. At both sites, the former three spread more in raw spoil than in topsoil, while with C. dactylon at both sites and C. ventricosa at Howick, the opposite occurred.
|
Dichanthium sericeum, Elymus scaber, Eragrostis sp. ‘A’, Paspalidium jubiflorum and Sporobolus creber, which were absent or had extremely low numbers in the units in which they were sown, occurred mainly in other topsoil units at Ravensworth but not at Howick (Table 4).
Among exotic species, C. gayana had the greatest spread at Ravensworth and to a lesser extent at Howick, where L. perenne and P. aquatica were the most ubiquitous exotic species (Table 4). While C. gayana had relatively greater spread at both sites in raw spoil, for L. perenne and P. aquatica it was greater in topsoil. At Ravensworth, spread of Bothriochloa insculpta and Panicum maximum were much higher in topsoil than it was in raw spoil.
Unsown species
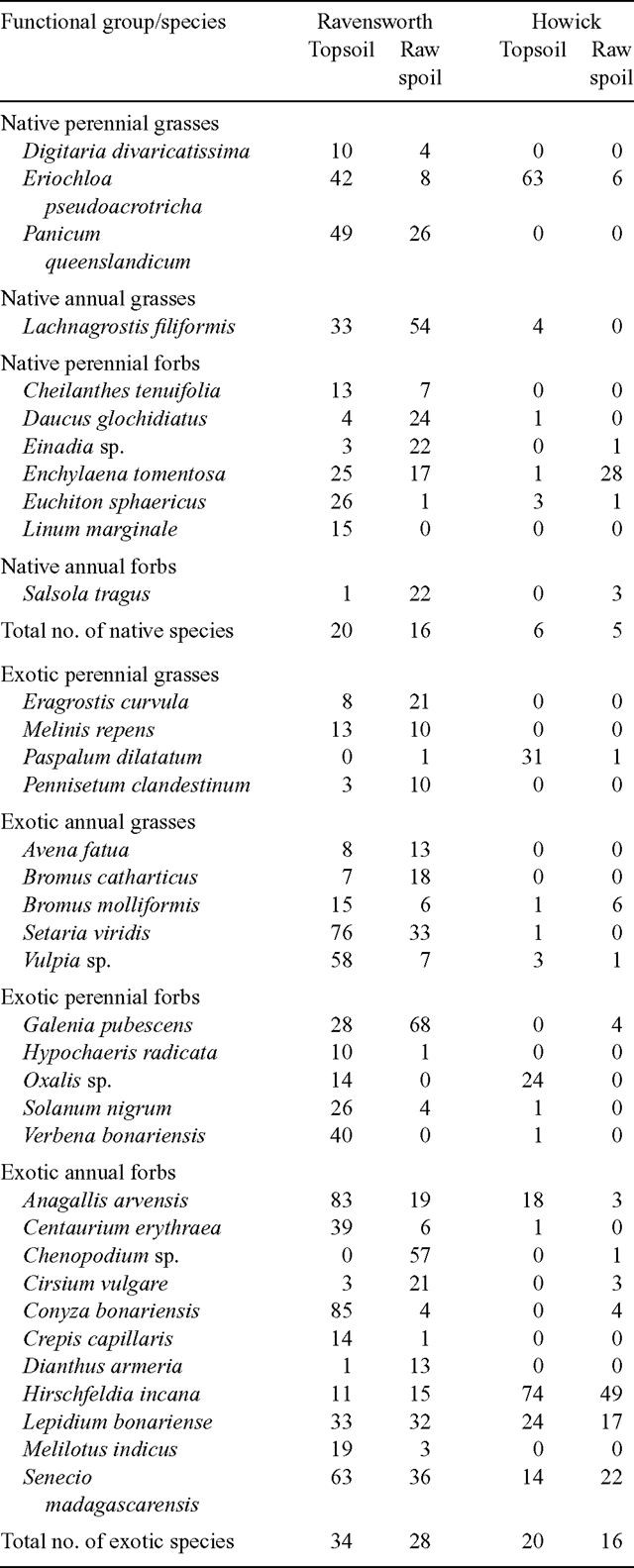
The two sites varied considerably in the numbers and abundances of unsown species that were recorded during the experiments (Table 5). In general, however, there were more unsown native and exotic species recorded at Ravensworth than at Howick and more species recorded in topsoil than in raw spoil at both sites.
|
Eriochloa pseudoacrotricha was the most commonly occurring unsown native perennial grass in topsoil at both sites while Panicum queenslandicum and Digitaria divaricatissima occurred in both topsoil and raw spoil at Ravensworth only, the former species being common. The native annual grass Lachnagrostis filiformis was common at Ravensworth while almost absent at Howick. Enchylaena tomentosa, a native perennial forb, was common at Ravensworth and had a preference for raw spoil at Howick (Table 5). Daucus glochidiatus, Einadia sp. and Salsola tragus were found mostly on raw spoil at Ravensworth while Cheilanthes tenuifolia, Euchiton sphaericus and Linum marginale were mostly on topsoil at Ravensworth.
The most common unsown exotic perennial grasses were Paspalum dilatatum (Howick) and Eragrostis curvula, Melinis repens and Pennisetum clandestinum (Ravensworth) (Table 5). There were more species and a greater abundance of exotic annual grasses at Ravensworth than at Howick. At Ravensworth Setaria viridis and Vulpia spp. were very common on topsoil but virtually absent from Howick. The most ubiquitous exotic perennial forb at Ravensworth was Galenia pubescens, where it was most common on raw spoil, while at Howick it was Oxalis sp. in topsoil (Table 5). Other exotic perennial forbs including Hypochaeris radicata, Solanum nigrum and Verbena bonariensis were mostly restricted to topsoil at Ravensworth. The most common exotic annual forbs at both sites and on both soil types included Anagallis arvensis, Hirschfeldia incana, Lepidium bonariense and Senecio madagascarensis. Other exotic annual forbs occurred at Ravensworth, including Centaurium erythraea, Conyza bonariensis, Crepis capillaris and Melilotus indicus which occurred most commonly on topsoil and Chenopodium sp., Cirsium vulgare and Dianthus armeria most commonly on raw spoil (Table 5).
Discussion
Establishment of sown species in designated experimental units
Of the 24 species sown in each experiment, only half had levels of establishment that could be considered acceptable. With the exception of C. dactylon (both sites) and C. ventricosa (Howick only), all of the most successful natives were cool season perennials and these species had established by June 1995, 2 months after sowing. Many of the sown native warm season species had failed to establish by this time and many of these were not recorded on either site for the duration of the experiment. This can be attributed, at least partly, to the conditions around the time of sowing in autumn 1995, which included good rainfall and lower temperatures, which would have favoured cool season species rather than warm season species. Another attributing factor was the ability of seed of the sown warm season species to remain viable in the soil until conditions favoured germination. Seed of C. dactylon, C. gayana and B. insculpta, 3 species that mostly had not emerged by June 1995 but were highly successful overall, had presumably remained viable in the soil for some time. Seed of many sown native warm season species apparently did not survive in the soil or did not germinate for some other reason. Survival of seed of warm season native species in the soil for prolonged periods however, has been recorded for Dichanthium sericeum (Watt and Whalley 1982). In addition, Themeda australis has been successfully established on acidic, oxidised raw spoil on a mine rehabilitation site in central western NSW (Windsor et al. 2000).
While percentage emergence was not measured, the numbers of plants established by June 1995, 2 months after sowing, did not appear to closely match germination percentages achieved under controlled conditions in the laboratory, despite adjusting sowing rates to allow for dormant or non-viable seed. This is consistent with previous observations where percentage seedling establishment in the field is often much lower than germination percentages attained under controlled laboratory conditions (e.g. Jefferson et al. 1991; Huxtable and Whalley 1999) and is indicative of the many factors in the field that can affect seed germination and plant survival. Recommended sowing rates of pasture species for mine rehabilitation in the Hunter Valley are 20–60 kg/ha which equates to 2000–8500 seeds/m2 (Huxtable et al. 1997), figures that reflect a ready seed supply and ensure rapid ground cover formation. Sowing rates in these experiments were much lower than this (100 seeds/m2 and 35 seeds/linear metre at Ravensworth and Howick, respectively).
Among sown native grasses, the five accessions of Austrodanthonia and A. bigeniculata had the most plants established by June 1995 (2 months after sowing). Both Austrodanthonia bipartita and A. richardsonii are recognised as being good colonising species, and are often associated with soil disturbance and can respond positively to soil phosphorus (Scott and Whalley 1982). Both cultivars of A. richardsonii ‘Hume’ (Jefferson et al. 1991) and ‘Taranna’ (Huxtable and Whalley 1999), have been successfully established on roadsides, so their success on disturbed mine sites concurs with these findings. In addition, A. richardsonii is reported as tolerating saline soils and neutral to alkaline pH (Scott and Whalley 1982). These factors concur with its success on the slightly alkaline soils in these experiments, and with the relatively high P levels in the raw spoil [72 and 34.7 mg/kg (Bray) at Ravensworth and Howick, respectively]. Both Austrodanthonia caespitosa and A. setacea are widespread species in NSW and often co-occur in pastures. Studies in the Riverina region of NSW have shown that extensive grasslands dominated by A. caespitosa have been derived from an earlier vegetation type, partly as a result of heavy grazing (Moore 1953; Williams 1961). Its colonising ability, drought tolerance and prolific seeding ability (Williams 1961), are features that make this species likely to do well in mine rehabilitation in the Hunter Valley. In these experiments, A. bigeniculata also established in high numbers in both topsoil and raw spoil and apparently has a tolerance to alkaline soils, as has a similar species Austrostipa aristiglumis (R. D. B. Whalley, pers. comm.). This species has hygroscopically active awns that aid seed movement to favourable microsites (Peart 1979) and, combined with its comparatively large seed size, may have contributed to its successful initial establishment in these experiments.
Exotic species that had the best initial establishment, P. aquatica, T. elongatum and L. perenne, were all C3 cool season species. As was the case for the native species, this reflects the seasonal conditions at the time of sowing and for the subsequent 2 months. The cultivar of P. aquatica sown in these experiments, ‘Australian’, is regarded as being tolerant of harsh conditions. Thinopyrum elongatum is well recognised as being tolerant of saline and poorly drained soils, is slow to establish and forms tall unpalatable growth if ungrazed (NSW Agriculture 2004). Lolium perenne is regarded as having low drought tolerance, prefers fertile soils and regenerates readily from seed following dry periods (NSW Agriculture 2004). At Ravensworth, C. gayana, an exotic C4 warm season species, was not recorded until July 1996, 15 months after sowing while at Howick, C. gayana had established by June 1995. It is difficult to explain this result, other than that some difference in soils at the 2 sites may have caused a difference in germination rates. Chloris gayana is recorded as being tolerant of a wide range of soils including saline soils, capable of rapidly establishing groundcover and is not tolerant of drought (Queensland Department of Primary Industries and Fisheries 2004). Bothriochloa insculpta cv. ‘Bisset’, another exotic warm season species is adapted to low fertility soils, forms rapid groundcover and should be sown in January or February (Queensland Department of Primary Industries and Fisheries 2004). Its success in the experiment at Ravensworth is consistent with this suggestion.
A few native C4 warm season species established in very low numbers, generally months, or even years after being sown. This may reflect more favourable soil temperatures and moisture, an improvement in soil characteristics over time and/or loss of seed dormancy.
Survival and spread of sown species over time
At Ravensworth in topsoil, there were a large number of seedlings of the five Austrodanthonia accessions, A. bigeniculata and C. dactylon recorded in July 1996. With the exception of Cynodon, which had established later than the other native species, these seedlings represented a substantial recruitment from seed produced by the first cohort of plants established at the site. High rainfall during autumn 1996 and in the preceeding spring and summer (over 220 mm each), may have contributed to this result. Given favourable seasonal conditions, young grass plants are capable of setting many more times the amount of seed than was used to establish them (Cole and Johnston 2005). Similar substantial seedling recruitment was not apparent at Howick, or in raw spoil at Ravensworth and may be due to poorer moisture retention in these other substrates, although this was not measured.
Initial establishment levels of A. bigeniculata and the five accessions of Austrodanthonia in raw spoil generally never exceeded those in topsoil and were generally poorer. As time passed however, many topsoil units were invaded by C. gayana, C. dactylon and to a lesser extent B. insculpta (Ravensworth), or L. perenne, P. aquatica and C. dactylon (Howick). The vigorous rhizomatous and/or stoloniferous species particularly, were able to form dense groundcover in many experimental units and native species disappeared or were greatly reduced both in numbers and contribution to cover. This can be attributed mainly to competition effects and the absence of grazing or any other form of biomass reduction. In raw spoil units however, competition from these species was much reduced or absent and A. bigeniculata and Austrodanthonia spp. spread extensively out of their designated areas and generally increased in number and cover. An exception to this however, was C. gayana at Ravensworth, that spread extensively to raw spoil areas and, while initially sparse in cover, by April 2000 it occurred in about 90% of experimental units and had dense groundcover in many of these. At Howick, however, C. gayana occurred in fewer topsoil and raw spoil units and contributed far less to groundcover. A second exception was C. ventricosa, which maintained cover levels of 21–36% in designated topsoil units and spread to some adjacent units, but did not establish or persist in raw spoil. Lolium perenne spread to between 80 and 100% of topsoil experimental units and competed through having large numbers of small plants. Phalaris aquatica occurred in between 51 and 62% of units, having had fewer, but larger, more robust plants. In comparison to A. bigeniculata and the five species and cultivars of Austrodanthonia, C. ventricosa had very few plants establish or persist in raw spoil at Howick.
Several sown species that were not recorded, or had extremely low numbers initially, appeared later either inside or outside their designated experimental units at Ravensworth. These included the native species C. truncata, Dichanthium sericeum, Eragrostis sp. ‘A’, Paspalidium jubiflorum, Sporobolus creber and Elymus scaber and the exotic species Panicum maximum var. trichoglume. This could be due to a loss of dormancy and/or the seed bed environment becoming more amenable to germination and establishment over time. It is also possible that seed of these same species was already present in the soil seed bank before sowing. With the exceptions of C. truncata and P. maximum var. trichoglume, none of these species were recorded until a degree of vegetative groundcover had been established. The ability of native grass seed to remain viable in the soil for prolonged periods until conditions are suitable for germination has been documented for a range of species (e.g. Watt and Whalley 1982; Silcock et al. 1990; Huxtable and Whalley 1999) and such a process may have occurred at Ravensworth. Recruitment of these species was absent at Howick, highlighting differences in soil chemistry and soil seed bank characteristics.
Differences in soil characteristics
The differences in results between Ravensworth and Howick can at least partly be attributed to a combination of soil physical and chemical characteristics. Both sites had topsoil that was classed as mildly alkaline, but Ravensworth topsoil was slightly more alkaline (pH 7.7), was a clay loam and was less saline (eCEC 10.3) than Howick topsoil (pH 7.4, medium clay, eCEC 14.8). In addition, while soil samples from different replicates were bulked for simplification before analysis, results in the field indicated that variation must have existed. In the process of topsoil and raw spoil placement, material from very different sources can be placed adjacently on an area designated for rehabilitation. In this case, one truck load of topsoil was enough for a topsoil block in one replicate, so that the next replicate topsoil block would come from a different truckload and probably from a different source.
Seed bank characteristics
While secondary to the main aim of these experiments, which was to assess establishment from sown seed, soil seed banks were an important factor influencing results. Both desirable and undesirable species influenced competition for light, nutrients and water of the sown species, as well as contributing to the composition of vegetation at the sites.
Ravensworth had a greater number and abundance of unsown native and exotic species than Howick. It is reasonable to assume that plants of these species were derived primarily from the seed bank, rather than influx from adjacent areas, which were vegetated with a dense sward of C. gayana and P. maximum var. trichoglume. Stockpiling of topsoil is a common practice in coal mine rehabilitation and the topsoil at Howick had been stockpiled for several years, before spreading on site (B. Foster, pers. comm.). In comparison, topsoil at Ravensworth had been freshly stripped and placed on the rehabilitation site. Native grasses such as the perennials Eriochloa pseudoacrotricha and P. queenslandicum and the annual species Lachnagrostis filiformis which became established in abundance during the experiments, provided ground cover and litter. Such an environment may also have enhanced subsequent seedling recruitment of sown species. In addition, native forbs became established and added to the diversity of the vegetation sward at Ravensworth. The benefits of selecting topsoil with a viable native seed bank are obvious.
The soil seed banks in these experiments also contained aggressive weed species, notably Galenia pubescens and Hirshfeldia incana. Spraying for broad-leaved weeds at Ravensworth was undertaken in March 1995 (Grazon® and Roundup®, both at 2.0 L/ha), before sowing in early April 1995. In addition, both Ravensworth and Howick were sprayed with Grazon® in October 1995 to control severe infestations of Galenia pubescens at Ravensworth and Hirshfeldia incana at Howick. Grazon® is a residual herbicide and these treatments no doubt greatly influenced the abundance and types of weeds occurring after this time. The effects of weeds can be minimised by selection of topsoil containing less weed seed, increasing the sowing rate of native seed and by strategic spraying during at least the first year.
It is likely that unsown species that occurred in raw spoil units had arisen from seed blown in from adjacent areas, or was brought in on the wheels of machinery used in site preparation. While often being considered inferior to topsoil as a growth medium for plants, the lack of weed competition confers an advantage to native grasses, which are often slow to establish. This has been similarly reported by Windsor et al. (2000) with successful establishment of T. australis in the absence of weeds, on oxidised raw spoil on a minesite in the central west of NSW. In addition, raw spoil in Hunter Valley coal mines can also be highly variable in its physical and chemical characteristics and in fact, sometimes provides a better growth medium that topsoil (Elliott and Veness 1981). By increasing the sowing rate of native grasses in raw spoil, the promising results obtained during these experiments could be further enhanced.
Management implications
These experiments have indicated that several native species have potential for use in rehabilitation of coal mine spoil in the Hunter Valley. The study has also indicated the relative advantages and disadvantages of using topsoil compared to sowing into raw spoil material. Some native species used in these experiments successfully survived and increased after initially poor establishment in raw spoil, while competition from exotic pasture grasses and weeds proved to be a problem as the experiment progressed, despite initially good establishment. Costs of transport and spreading topsoil to the required depth of 0.2 m on a rehabilitation site vary depending on how far the material is transported, but can be in the order of up to $6000/ha (D. Stewart, pers. comm.). Not using topsoil can therefore represent considerable savings to the mining company. Additional savings can be made in reduced fertiliser and weed control costs which are estimated at $500/ha and over $500/ha for weed control respectively (NSW Department of Mineral Resources 1999). In these experiments we applied fertiliser at the rate of 100kg/ha, which is approximately half the recommended rate and spraying for weeds was only necessary on the topsoil areas.
The costs of seeding rehabilitation sites using the current recommended species is estimated at $1800/ha, but this would also include the cost of the machinery used to do it (NSW Department of Mineral Resources 1999). At present native grass seed is considered to be expensive, largely as a result of its scarcity. Native grass seed can cost anywhere between $20/kg and $800/kg depending on the level to which it has been cleaned, purity, viability and germination percentage (I. Chivers, S. Rahilly, pers. comm.). It is likely that the higher cost of native grass seed in comparison to exotic species can be offset by reduced costs of not using topsoil, applying less fertiliser and reduced weed control.
Conclusions
These experiments tested the establishment and persistence of a broad range of species from an autumn sowing, over a long period in two typical mine rehabilitation environments. From this study several conclusions can be drawn.
The most successful sown native species at the two sites included the C3 species A. bigeniculata, A. bipartita cv. ‘Bunderra’ A. caespitosa, A. richardsonii cvv. ‘Taranna’ and ‘Hume’ and A. setacea; and the C4 species C. truncata and C. ventricosa. Based on recruitment from the seed bank observed in these experiments, the C4 species E. pseudoacrotricha and P. queenslandicum and the C3 species L. filiformis may also have potential as sown species.
Given the desirability of establishing native warm season species on mine sites in the Hunter Valley, further attempts to improve on the results obtained in these experiments are warranted. These include sowing in late winter or early spring to enable seedlings to establish adequately before the hot dry summer and trialling with other seed sources.
In general, sowing rates need to be much higher to attempt to out-compete weeds and undesirable exotic pasture species. Given the success of the best native species and the extremely low sowing rates used in these experiments, even better results could be expected if sowing rates were increased.
While establishment from topsoil sowings of the above-mentioned species is better initially, the lack of weed competition in raw spoil can provide better prospects for establishment in the long term. By sowing at higher rates, this advantage may be further enhanced.
Careful selection of topsoil can greatly enhance successful establishment of native grasses, both from sown seed and from the soil seed bank. Where possible, topsoil should be stripped from areas of predominantly native grassland and spread immediately on site, rather than being stockpiled.
Acknowledgments
The authors thank the Australian Coal Association Research Program (ACARP), who funded the 5-year project of which this paper is part. The Hunter Coal Environment Group initiated the project and members of the steering committee (Peter Brennan, Tony Voller, Geoff Marschke, Greg Summerhayes, Doug Stewart, Brian Medhurst, James Bailey and Bruce Foster) provided direction for the project. Thanks go to Environmental Officers at Howick and Ravensworth South mines for providing and preparing sites. Valuable technical assistance in the field was provided by Marianne Close and Andrew Sawicki and was much appreciated. Thanks also go to Ken Reynolds, Steven Young, Glenda Holman and Kerry Hindle and others at the Scone Soils Laboratory for conducting soil analyses. Thanks to Bill Semple and Ian Cole for critical review and helpful suggestions to earlier drafts of this paper.
Barr D. A.
(1965) Restoration of coastal dunes after beach mining. Journal of the Soil Conservation Service of NSW 21, 199–209.
(accessed 24/02/2004).
Dyson, J. R. ,
Elliott, G. L. ,
Reynolds, K. C. ,
Dight, D. C. G. ,
and
Hill, M. J. (1987).
Elliott G. L., Veness R. A.
(1981) Selection of topdressing material for rehabilitation of disturbed areas in the Hunter Valley. Journal of Soil Conservation Service NSW 37, 37–41.
(accessed 31/03/2004).
NSW Department of Mineral Resources (1999).
Northcote, K. H. (1979).
Peart M. H.
(1979) Experiments on the biological significance of the morphology of seed-dispersal units in grasses. Journal of Ecology 67, 843–863.
(accessed 31/03/2004).
Scott A. W., Whalley R. D. B.
(1982) The distribution and abundance of species of Danthonia DC on the New England Tablelands (Australia). Australian Journal of Ecology 7, 239–248.
and 2000) or Royal Botanic Gardens Sydney Plantnet (2004).
2Because of the difficulty of identification in the field, Austrodanthonia accessions are grouped and will be referred to collectively as ‘Austrodanthonia spp.’ in this section of the results.