

Evidence for palatability variation between the sex phenotypes of bladder saltbush from small-plot grazing trials
D. Walsh A D , M. H. Andrew B , R. Sinclair C and D. Coleman CA Centralian Land Management Association, PO Box 2534, Alice Springs, NT 0871, Australia.
B URS Australia Pty Ltd, 25 North Tce, Hackney, SA 5069, Australia.
C University of Adelaide, Adelaide, SA 5005, Australia.
D Corresponding author. Email: ems@clma.com.au
The Rangeland Journal 27(2) 89-96 https://doi.org/10.1071/RJ05007
Submitted: 23 February 2005 Accepted: 24 August 2005 Published: 21 November 2005
Abstract
This paper reports the results of the first intensive grazing trials to study palatability variation between the sex phenotypes of bladder saltbush (Atriplex vesicaria Hew. ex Benth.). The results show that Merino sheep preferentially graze female shrubs over male and bisexual shrubs in South Australia, which supports earlier paddock-scale observations made in New South Wales. Male shrubs with higher densities of flower spikes were consumed less than those with lower densities. However, when male flowers were small and/or sparse, the removal of male flowers by clipping did not significantly increase the consumption of male shrubs. Female shrubs with very low or very high densities of fruit tended to be consumed significantly more than those with moderate fruit densities. Other physical aspects such as plant growth habit and foliage density did not have a consistent influence on grazing behaviour. Chemical differences between the sex phenotypes are implicated.
The presence of palatability variation between the sex phenotypes suggests that there is potential to refine the existing use of bladder saltbush as an indicator species in southern Australia.
Additional keywords: Atriplex vesicaria, rangelands, selective grazing.
Introduction
Atriplex vesicaria (bladder saltbush) is a low-growing, subdioecious shrub with a lifespan of about 25 to 30 years (Crisp 1978). It has long been recognised as an important species for ecosystem stability and pastoral production in the rangelands of southern Australia (Wilson and Graetz 1979). Due to its susceptibility to grazing and trampling, bladder saltbush has also assumed importance as an ‘early-warning’ indicator of change in range condition in chenopod shrublands (Lange et al. 1994).
Before this study, a growing body of evidence suggested that palatability variation occurs within the species. Palatability variation has been noted between ecotypes (Knowles and Condon 1951; Graetz and Wilson 1984), age classes (Knowles and Condon 1951; Lange et al. 1992; Maywald 1993) and between plants of different geographic origin (Pearson et al. 1990). Most of the research, however, has focused on the differential acceptability of the sex phenotypes to Merino sheep. In separate studies, Williams et al. (1978) and Graetz (1978) found that the sex ratio of grazed saltbush populations was strongly biased in favour of the male phenotype. Graetz (1978) found up to a 5-fold increase in the female to male sex ratio with increasing distance from water. Maywald et al. (1999) reported that the ratio of male to female plants on grazed sites was up to 12 times higher than on adjacent ungrazed sites. Furthermore, it has been found that female shrubs are more likely to be grazed than male shrubs at all distances from water (Graetz and Wilson 1979). Williams (1972) and Williams et al. (1978) found that male plants in grazed populations were vegetatively and reproductively more vigorous than females of the same lifestage, and noted that this could be explained by selective grazing. Graetz (1978) noted that preferential grazing of females reduced their size in comparison to males. Similar results have been reported by Hunt (1995) and Maywald (1998). If the sex phenotypes do indeed differ in their susceptibility to grazing, the use of bladder saltbush as an indicator of rangeland condition and trend could potentially be refined.
Possible reasons for the selectivity have been subject to debate. Graetz (1978) suggested that sheep avoid the male shrub because the terminally placed flowering spike acts as a mechanical obstruction to the preferred foliage below. However, Williams (1972) believed that sheep seek out and preferentially graze the succulent unripe fruits on female shrubs.
This paper reports the findings of three small-plot grazing trials undertaken to:
-
(a) verify that the sexes vary in their palatability in the rangelands of South Australia, and
-
(b) to test the influence of some physical plant attributes thought to influence grazing behaviour.
Materials and methods
Study site
Three grazing trials were conducted in Little One Mile Paddock, Middleback Station (33°10′S, 137°20′E), South Australia (Fig. 1). The site, described by Lange et al. (1992), occupied a broad, flat swale dominated by an Atriplex vesicaria–Maireana pyramidata (Benth.) Paul G. Wilson shrubland, with shrub densities of about one shrub per square metre. Little One Mile is a small paddock which is rarely stocked and was thus considered to be unmodified by grazing (A. W. Nicolson, pers. comm.). As the site had not been modified by continuous grazing by sheep, the saltbush population was expected to encompass a large range of palatability variation.
|
Trials were conducted in September 1994, February 1995 and January 1996. This ensured that hypotheses were tested in a range of seasonal conditions and allowed the testing of new hypotheses as they arose. The first two trials were conducted in the same sheep-proof enclosure, which was 90 m long, 10 m wide and oriented north–south. The enclosure was treated as nine contiguous 10 × 10 m ‘cells’ and a holding yard was situated at the southern end of the enclosure. A new enclosure was built for the January 1996 trial to provide a dataset independent of the previous trials. The enclosure used in January 1996 was 70 m long, 10 m wide and was situated directly adjacent to the other plot. Small-plot grazing trials using similar methods have been used for studying the grazing behaviour of Merino sheep in the rangelands of semi-arid South Australia for many years (Noble 1975; Pearson et al. 1990; Lange et al. 1992; Maywald 1993).
Experimental shrubs
In September 1994, small aluminium tags were attached to the bases of about 24 sexually mature shrubs in each cell. The shrubs were chosen to ensure a relatively even distribution throughout the cell. Where possible, equal numbers of shrubs of both sexes were chosen. Although bladder saltbush populations are usually dominated by the male and female phenotypes (Williams 1979), the experimental population was dominated by female and bisexual plants in September 1994. As a result, the foliage consumption from 106 females and 108 bisexuals was compared. However, by February 1995, most of the bisexual shrubs had changed sexual phenotype, so the consumption of 93 females and 84 males was compared. The phenomenon of sexual lability has been found to occur quite naturally in both grazed and ungrazed populations of bladder saltbush (Maywald 1996; Walsh 2005). A small number of shrubs died after the September trial and some had remained bisexual. For these reasons, less shrubs were monitored in the February trial. In January 1996, ten sexually mature shrubs of each sex were tagged in each cell of the new plot, resulting in 70 female and 70 male shrubs.
Shrub size and habit
The maximum width and height of every experimental shrub was measured before grazing commenced. The measurements were made using a metre ruler and were recorded to the nearest centimetre. The growth habit of each shrub was also classified as either erect or spreading. Erect shrubs were those with no branches touching the ground and spreading shrubs had at least one branch touching the ground. Height, width and habit were used to compare the architecture of the sexes before grazing. Shrub habit was also included as an experimental factor in the analyses to determine whether there was any influence of habit on foliage consumption.
Additional treatments in the September 1994 and February 1995 trials
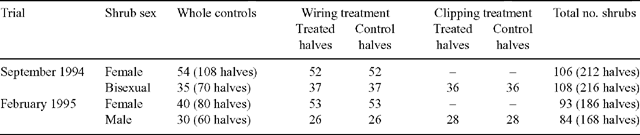
In addition to monitoring the consumption of shrubs on the basis of sex and habit, two extra treatments were included to determine the influence of foliage density and male flower spikes (Table 1). Observations in paddocks at Middleback suggested that these factors may influence the grazing behaviour of Merino sheep. To investigate these aspects, the following treatments were applied to the experimental shrubs:
|
Wired shrub halves with control halves
This treatment was included to test the effect of foliage density on selectivity. The underlying assumption was that grazing from densely packed foliage should maximise the intake per bite and, thus, may encourage preferential grazing. Randomly selected shrubs of both sexes were manipulated to create two choices in foliage density. For each of these shrubs, one half of the plant was left untreated and the other half was bunched together and tied with fine wire to produce a dense array of foliage. This allowed foliage density to be examined independently of underlying palatability. The untreated shrub halves acted as controls for the increase in foliage density, while the non-manipulated whole shrubs (described previously) acted as controls for the altered appearance of wired shrubs. Where possible, equal numbers of shrubs were chosen in each cell. The shrubs in even-numbered cells had the eastern halves wired, while shrubs in the odd-numbered cells had the western halves wired. This restricted randomisation was used to minimise the time required to relocate treatments, while acting as a control for aspect.
Clipped shrub halves with control halves
This treatment was included to test whether male flower spikes influence consumption and was thus applied to bisexual and male shrubs only. For each of the chosen shrubs, the spikes were clipped from one half while the other half was left as an untreated control. As with the wiring treatment, the untreated halves acted as a control for the removal of male spikes and whole shrubs (described previously) acted as a control for the altered appearance of the treated shrubs. The same restricted-randomisation pattern used to assign treatments to wired shrubs was also used for the clipped shrubs. The wiring and clipping treatments were assigned to different shrubs so that no shrubs received both treatments.
Additional treatments in the January 1996 trials
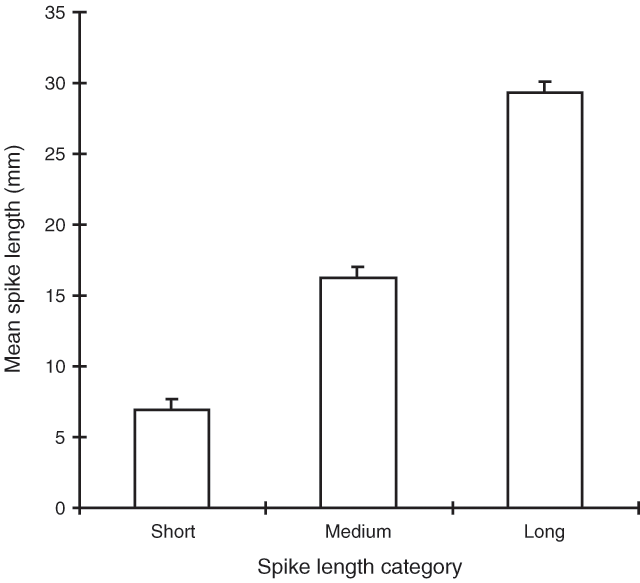
The results of the first two trials indicated that increased foliage density and the removal of male spikes did not significantly influence selectivity. For this reason, the wiring and clipping treatments were not included in the January 1996 experiment. Observations from the first two trials indicated, however, that sheep did tend to reject male shrubs with the longest and/or densest flower spikes. To quantify whether spike length was influential, a spike length category was assigned to every male shrub after visual inspection. The three categories were ‘short’, ‘medium’ and ‘long’. For those shrubs with a mixture of spike lengths, the numerically dominant category was recorded. A calibration of spike lengths was undertaken to quantify the means of the three size classes. The calibration shrubs were chosen randomly from the population directly adjacent to the experimental enclosures. One flowering branch was cut from each of ten shrubs of each spike size class to provide a total sample of 30 branches. These branches were bagged immediately and returned to the laboratory. The ten branches of a single size class were then spread evenly over a clean piece of white paper and 25 spikes were chosen from the material as follows. With eyes closed, a drawing compass was used to select a point on the paper and the spike closest to the compass point was cut from its branch using dissection scissors. This was repeated 25 times and the lengths of the selected spikes were then measured to the nearest millimetre. All material was then discarded and the area thoroughly cleaned before the next size class was prepared. The relationship between category and spike length is shown in Fig. 2. The mean spike lengths for all three categories were significantly different (P < 0.001).
|
In addition to male spike length, the percentage of branches with spikes (males) or fruits (females) was estimated for each experimental shrub. Five classes were used: 1–20, 21–40, 41–60, 61–80, and 81–100%. These classes were used to determine if spike and fruit density influenced consumption in the January 1996 trial.
Biomass estimation
Immediately before each trial, the twig and leaf biomass of each experimental shrub was estimated using the Adelaide technique (Andrew et al. 1979). This non-destructive method relies on comparisons between the experimental shrubs and a visual standard. In the first two trials, estimates were made separately for the east and west halves of each experimental shrub because the treatments had been applied to different halves.
Biomass consumption
Consumption was estimated using the microdot technique of Lange (1984). Five small paint dots were applied to the accessible foliage of each shrub half to provide a total of ten dots per shrub. The loss of microdots was monitored daily throughout each trial. The percentage of microdots lost was assumed to be directly proportional to the percentage of biomass lost (as demonstrated by Lange 1984). Microdots damaged by the sheep throughout the course of a trial were recorded and then discarded away from the site. This damage was subtracted from total biomass loss to provide an estimate of loss due to consumption. The new starting biomass was calculated for each day by subtracting biomass consumed and biomass damaged from the starting biomass of the previous day.
Experimental sheep
Merino wethers (12–18 months old) were used in all trials because they were always available, where as ewes were sometimes unavailable due to lambing. Eight sheep, selected from the larger station flock and kept overnight in the holding yard at the trial site, were used in the first two trials. Seven sheep were used in the third trial to account for the smaller enclosure size. The sheep were let into the enclosure the following morning and left to graze for 24 h. The sheep had unlimited access to water from a trough during the trials. At the end of each 24 h period the sheep were quietly mustered into the holding yard and kept there while the microdots were recounted. Counting usually took about 2 h and the sheep were let back into the enclosure after counting was completed. These procedures were repeated every day. All trials were terminated when the percentage of microdots fell below 20% of the initial number available. Typically, this took 4 days.
The high stocking rates used in these small-plot grazing trials did not attempt to reflect the average stocking rate of the pastoral district. In fact, the method relies on the very high stocking pressure to investigate patterns and processes that may take months or years to develop in real station paddocks. The validity of using a high stocking pressure has not been rigorously tested, however, the preferences measured in small-plot trials are highly consistent with those measured in other contexts (Jessup 1951; Leigh and Mulham 1966; Robards et al. 1967; Andrew and Lange 1986).
Analyses
Log10 transformations of height, width and initial biomass were performed after Shapiro–Wilk W tests showed that the raw data were not distributed normally. In all cases, transformation resulted in normally distributed datasets. After transformation, the heights, widths and initial biomass of the sexes were compared with Student’s t-test.
Relative biomass consumption was calculated for each shrub by adding the consumption for the first 2 days and dividing it by the initial biomass. The data for the first 2 days were used because herbivores are most selective when the choice of forage is highest (Vallentine 1990). For the first two trials, relative biomass consumption was regressed with each experimental factor (sex, habit and treatment) in a generalised linear model. The number of interaction terms was restricted because the clipping treatment was applied to bisexuals and males only. The interaction terms were, thus, sex × habit and sex × habit × wiring for both sexes and sex × habit × clipping for males and bisexuals only. For the third trial, analysis of variance (ANOVA) was used to examine the influence of spike length, spike density and fruit density on relative biomass consumption. After ANOVA, the Tukey–Kramer Honestly Significant Difference (HSD) test was used to identify the means that were significantly different. All analyses were performed using the JMP program (SAS Institute, Cary, NC).
Results
September 1994
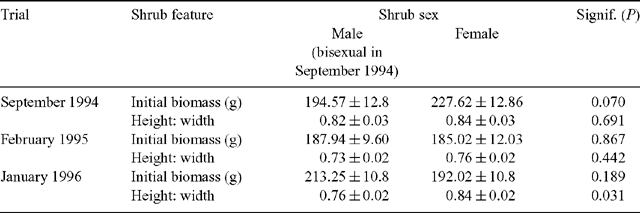
The initial biomass of the separate halves of the tagged shrubs were not significantly different (P > 0.1), therefore, the total biomass of each shrub was used to compare the sexes. The results indicated that the bisexual shrubs had slightly lower biomass compared with females, but this was not statistically significant (P > 0.05; Table 2). The height to width ratio of the sexes was also not significantly different (P > 0.1; Table 2).
|
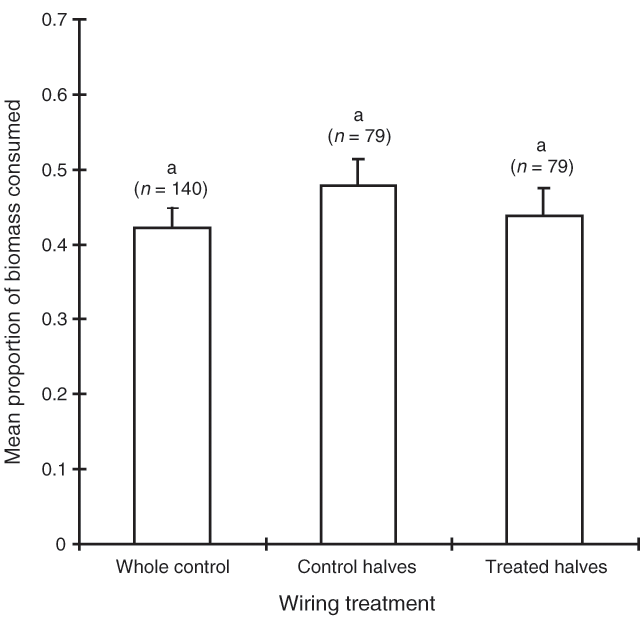
Sex was the most influential factor determining consumption of bladder saltbush in the September 1994 trial (Table 3). Sheep consumed 36% of the female biomass available and only 21% of the bisexual biomass. Shrub habit was of borderline significance with spreading shrubs being consumed more than erect shrubs (Table 3). However, there was no significant interaction effect between sex and habit (Table 3). Neither the clipping nor wiring treatments had a significant effect on consumption (Figs 3 and 4) and these treatments were, thus, pooled in the model (Table 3).
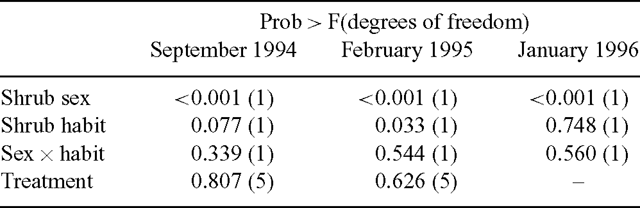
|
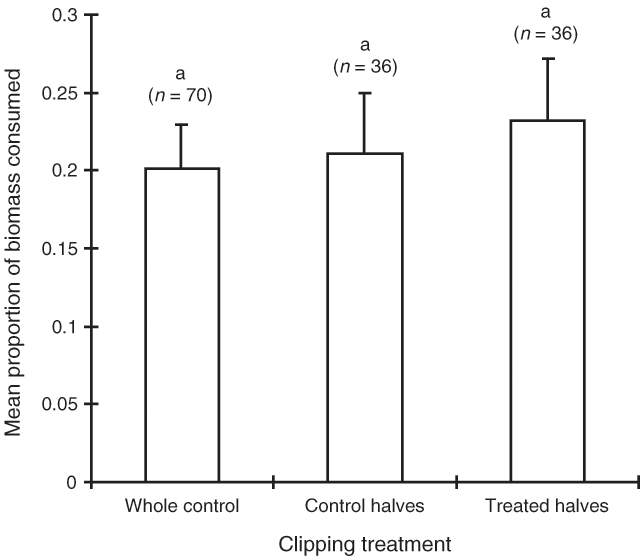
|
|
February 1995
Shrub biomass before the February 1995 trial was not significantly different from that at the start of the September 1994 trial (P > 0.1). This indicates that the biomass grazed from the shrubs in September was replaced by new growth during the rest period. Inspection of the data confirmed that 96% of the shrubs increased in biomass between the end of the September trial and the beginning of the February one. As in the previous trial, the biomass of the east and west halves was not significantly different (P > 0.1) and the biomass of whole shrubs was thus used to compare the sexes. Male and female shrubs did not differ significantly in initial biomass (P > 0.1; Table 2) or height to width ratio (P > 0.1; Table 2).
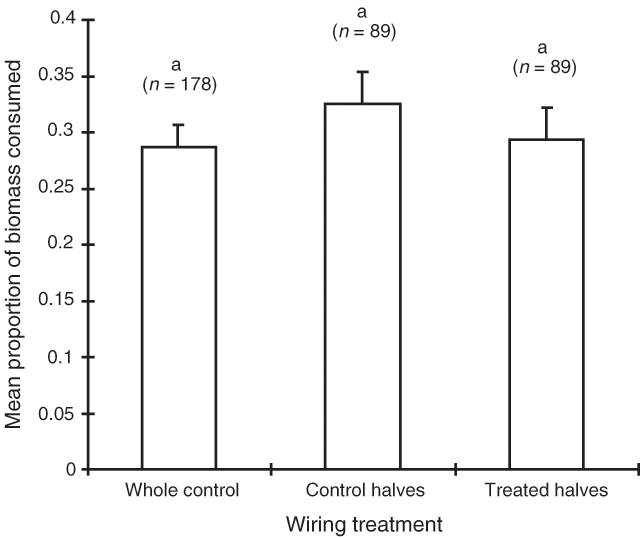
The consumption results of the February 1995 trial mirror those of September 1994 (Table 3). Once again, shrub sex was the most influential factor determining consumption of bladder saltbush, with 54% of the female biomass being consumed compared with 30% of the male biomass. Spreading shrubs were consumed more than erect shrubs, however, there was no interaction between sex and habit (Table 3). Once again, the clipping and wiring treatments had no significant effect on consumption (Figs 5 and 6).
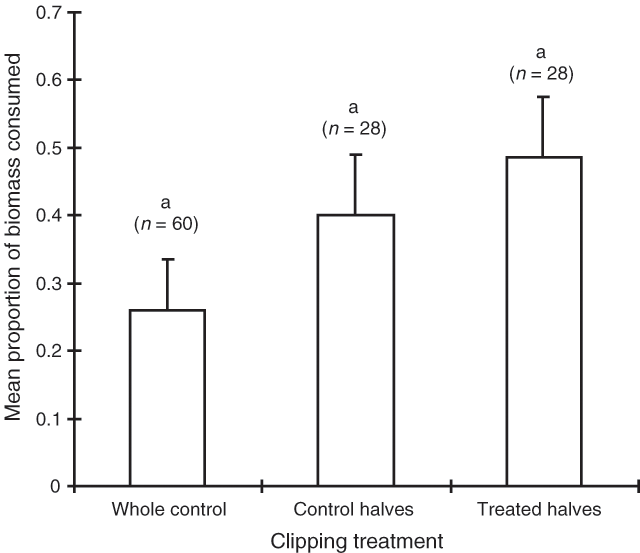
|
|
January 1996
The initial biomass of female and male plants was not significantly different (P > 0.1; Table 2). Unlike the previous 2 trials, however, the height to width ratios of the sexes differed significantly (P < 0.05; Table 2). On average, females had greater height to width ratios than males.
The January 1996 trial was the third demonstration that sex was the most important determinant of consumption (Table 3). The relative consumption of the sexes was similar to February 1995, with 57% of the initial female biomass consumed compared with 30% of the initial male biomass. Shrub habit was not a significant determinant of consumption in this trial.
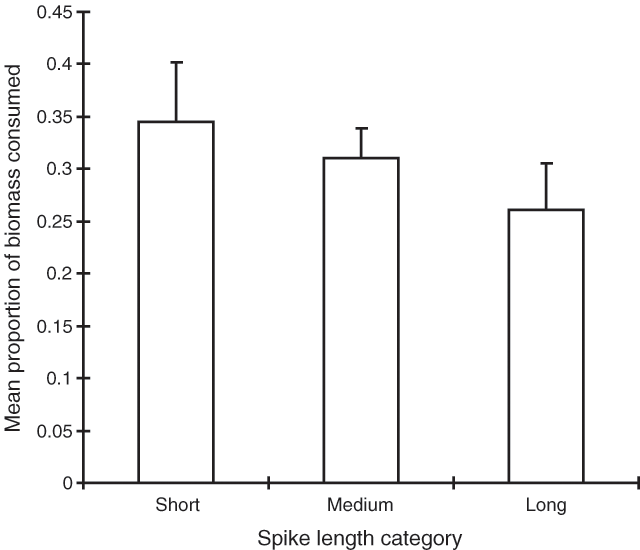
More male shrubs had medium length (n = 82) spikes than long (n = 36) or short (n = 22) ones. The mean relative consumption of the classes is shown in Fig. 7. Although the data suggest a trend towards decreasing consumption with increasing spike length, this was not significant (P > 0.1).
|
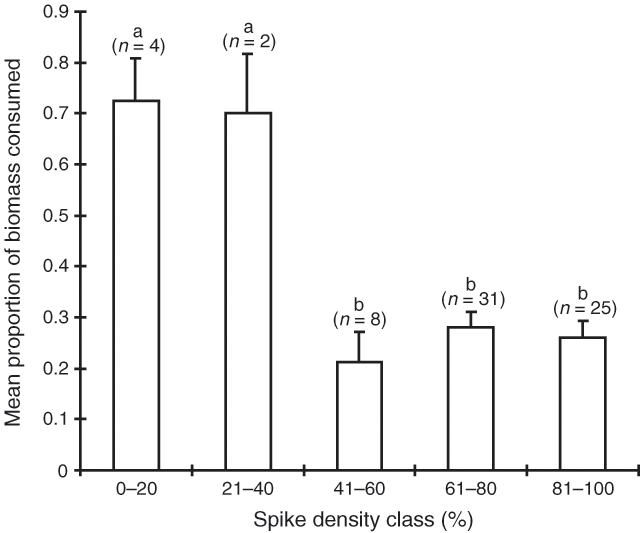
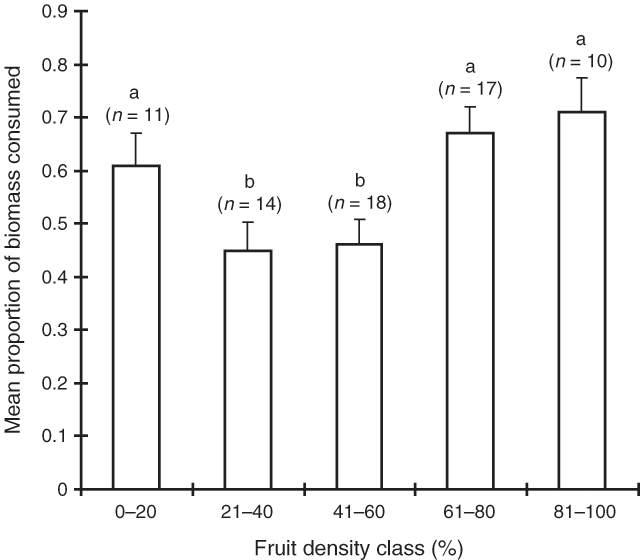
Consumption on the basis of spike and fruit density is shown in Figs 8 and 9. Male shrubs with lower densities of spikes were consumed more than those with higher densities (P < 0.001), however, the sample sizes in the lower density categories were low and this result should be treated with some caution. Female shrubs with moderate densities of fruit were eaten less than those with lower or higher densities (P < 0.01).
|
|
Discussion
The first aim in these trials was to quantify whether the sexes vary in their palatability in the semi-arid rangelands of South Australia. The results show that sex was indeed the most important of the factors measured. Females were consumed about 70% more than bisexuals in the first trial, 80% more than males in the second and 90% more than males in the third. This is consistent with observations made elsewhere and suggests that a preference for female shrubs is widespread throughout southern Australia.
Foliage density was not a significant determinant of consumption in these trials, even though it is known to influence herbivores in other contexts (Heady 1964; Vallentine 1990). The growth habit of shrubs appeared to be of some influence in the first two trials, but not in the third. The first two trials showed that spreading shrubs tended to be grazed more than erect shrubs. This may be because spreading shrubs allow sheep to access foliage more readily.
The third trial showed that male flower spikes may influence grazing, even though the clipping treatment in the first two trials did not support this. We suspect that the clipping treatment may not have been a sound way to test the influence of spikes. If spikes deter sheep (as proposed by Graetz 1978), then the removal of them from one half of a shrub may not result in an increase in consumption of that half. This is similar to the situation where a plant which is considered to be relatively palatable is left ungrazed due to its proximity to a less palatable one (Heady 1964; Arnold 1987). Thus, the clipped half of a saltbush may not be grazed because of the close proximity of spikes on the non-clipped half. Furthermore, because of a large rainfall event before the third trial, the saltbushes at the site flowered and fruited more heavily than in either of the previous trials. Given that the spikes were relatively smaller and sparser in the first two trials, the clipping treatment may not have greatly altered the clipped halves compared with the unclipped halves. This is supported by the fact that it was the males with the most densely packed spikes that were consumed the least in the third trial. Inclusion of the clipping treatment in the third trial may have helped to clarify this issue. Female shrubs with very low or very high densities of fruit tended to be consumed significantly more than those with moderate fruit densities. If sheep preferentially graze female shrubs because they are attracted to the fruits (as suggested by Williams 1972), we would have expected a pattern of increasing consumption with increasing fruit density. Instead, these trials lend support to the alternative hypothesis (Graetz 1978) that sheep avoid the male flower spikes. Later research using cafeteria trials confirms indeed that sheep use the male flower spike as a visual cue to an underlying chemical deterrent in male shrubs (Maywald 1998; Walsh et al. 2005).
Although the small-plot trials reported here were designed to investigate physical influences on grazing behaviour, it is worth foreshadowing the issue of an underlying chemical basis for the selectivity. On a world scale, the genus Atriplex is considered to be relatively benign in terms of chemical defences (Sanderson and Stutz 1994). This suggests that research into chemical differences between the sex phenotypes of A. vesicaria should focus initially on the simpler chemical groups or on chemicals already known to occur in Atriplex. For example, some Australian Atriplex are known to contain unpalatable concentrations of salts, saponins, triterpenes, oxalates and steroids (Simes et al. 1959; Osmond 1963; Sanderson and Stutz 1994). However, palatability differences between plants are not always due to toxic or secondary compounds. Nutritional factors such as protein, fibre and fats are also known to influence the selective grazing behaviour of animals (Malecheck and Provenza 1983). Although the relationship between palatability and chemical composition is highly complex and proof of causation is difficult to verify (Molyneux and Ralphs 1992), the chemical factors outlined above may provide a starting point for investigating differences between the sex phenotypes.
These three trials demonstrated that the relative palatability of the sexes of bladder saltbush is quite consistent through time. The three trials were conducted in three different years, in contrasting seasonal conditions, in two different plots, using three different flocks of sheep. A single experiment with three replicates would not have revealed the consistency of the behaviour over time. The results support the findings from piosphere studies in western New South Wales and the anecdotal observations made throughout southern Australia. This suggests that the cues influencing the sheep are quite strong and appear to elicit the same grazing behaviour from sheep separated through space, time and edaphic context. Given the accumulating evidence for palatability variation between the sexes across the species’ range, we hypothesise that the use of bladder saltbush as an indicator could potentially be refined. Measures that focus on the status of female shrubs (such as size and vegetative and reproductive vigour) may allow adverse changes to be detected even sooner than at present. Furthermore, the measurement of sex ratios may offer a rapid way of assessing rangeland condition in areas containing Atriplex vesicaria. We have since investigated these concepts as part of a larger study and will present our findings in future papers.
Acknowledgments
We are indebted to the Nicolson families of Middleback and Roopena stations, the many people who assisted with the field work and the undergraduate students from the 1994 and 1996 ‘Ecology and Management of Rangelands’ courses (The University of Adelaide). We thank Ian Watson and Alan Payne for reviewing an earlier draft of the manuscript. This research was funded by an International Wool Secretariat Postgraduate Scholarship and The University of Adelaide, and was conducted as part of the Middleback Field Centre research program. Animal ethics committee approval S/14/94.
Andrew M. H., Lange R. T.
(1986) Development of a new piosphere in arid chenopod shrubland grazed by sheep. 2: Changes to the vegetation. Australian Journal of Ecology 11, 411–424.

Andrew M. H.,
Noble I. R., Lange R. T.
(1979) A non-destructive method for estimating the weight of forage on shrubs. Australian Rangeland Journal 1, 225–231.
Arnold G. W.
(1987) Influence of the biomass, botanical composition and sward height of annual pastures on foraging behaviour by sheep. Journal of Applied Ecology 24, 759–772.
Crisp M. D.
(1978) Demography and survival under grazing of three Australian semi-desert shrubs. Oikos 30, 520–528.
Graetz R. D.
(1978) The influence of grazing by sheep on the structure of a saltbush (Atriplex vesicaria Hew. ex Benth.) population. Australian Rangeland Journal 1, 117–125.
Graetz R. D., Wilson A. D.
(1979) An assessment of herbivore diets in the chenopod shrublands. : ‘Studies of the Australian arid zone. IV: Chenopod shrublands’. (Eds R. D. Graetz, K. M. N. Howes)
pp. 144–159. (CSIRO Publishing: Melbourne: Vic.)
Graetz R. D., Wilson A. D.
(1984) Saltbush and bluebush. : ‘Management of Australia’s rangelands’. (Eds G. N. Harrington, A. D. Wilson, M. D. Young)
pp. 209–222. (CSIRO Publishing: Melbourne, Vic.)
Heady H. F.
(1964) Palatability of herbage and animal preference. Journal of Range Management 17, 76–82.
Hunt L. P.
(1995) Spatial variation in the population dynamics of Atriplex vesicaria under sheep grazing. PhD thesis, The University of Adelaide, Australia.
Jessup R. W.
(1951) The soils, geology and vegetation of north-western South Australia. Transactions of the Royal Society of South Australia 74, 189–273.
Knowles G. H., Condon R. W.
(1951) The perennial saltbush (Atriplex vesicarium Heward). New South Wales Soil Conservation Service Journal 7, 123–130.
Lange R. T.
(1984) Leaf marking in rangeland grazing studies. Transactions of the Royal Society of South Australia 108, 213–214.
Lange R. T.,
Coleman D., Cowley K. R.
(1992) Survival of saltbush (Atriplex vesicaria) seedlings in heavily grazed experimental plots. Australian Journal of Ecology 17, 99–101.
Lange R. T.,
Lay B. G., Tynan R. W.
(1994) Evaluation of extensive arid rangelands: the Land Condition Index (LCI). Transactions of the Royal Society of South Australia 118, 125–131.
Leigh J. H., Mulham W. E.
(1966) Selection of diet by sheep grazing semi-arid pastures on the Riverine Plain. 1: A bladder saltbush (Atriplex vesicaria)–cotton bush (Kochia aphylla) community. Australian Journal of Experimental Agriculture and Animal Husbandry 6, 460–467.
| Crossref | GoogleScholarGoogle Scholar |
Malecheck J. C., Provenza F. D.
(1983) Feeding behaviour and nutrition of goats on rangelands. World Animal Review. 47, 38–48.
Maywald D. L.
(1993) The importance of context in understanding the plant-herbivore interaction in the South Australian arid zone. Honours thesis, The University of Adelaide, Australia.
Maywald D. L.
(1996) Sexual lability in bladder saltbush (Atriplex vesicaria).
: ‘Conference papers ninth Biennial Australian Rangeland Conference. Focus on the future – the heat is on!’. (Australian Rangeland Society: Perth, W. Aust.)
Maywald D. L.
(1998) Palatability variation between the sex phenotypes of bladder saltbush (Atriplex vesicaria). PhD thesis, The University of Adelaide, Australia.
Maywald D. L., Andrew M. H., Sinclair R., Coleman D.
(1999) Palatability differences between the sexes of Atriplex vesicaria in southern Australia.
: ‘People and rangelands: building the future. Proceedings VI International Rangeland Congress’. (Eds D. Eldridge, D. Freudenberger)
pp. 455–456. (VI International Rangeland Congress Inc: Townsville)
Molyneux R. J., Ralphs M. H.
(1992) Plant toxins and palatability to herbivores. Journal of Range Management 45, 13–18.
Noble I. R.
(1975) Computer simulations of sheep grazing in the arid zone. PhD thesis, The University of Adelaide, Australia.
Osmond B.
(1963) Oxalates and ionic equilibria in Australian saltbushes (Atriplex). Nature 198, 503–504.
Pearson J. T.,
Sparrow A. D., Lange R. T.
(1990) Prolonged exposure to sheep grazing reduces the palatability of Australian saltbush populations. Australian Journal of Ecology 15, 337–344.
Robards G. E.,
Leigh J. H., Mulham W. E.
(1967) Selection of diet by sheep grazing semi-arid pastures on the Riverine Plain IV: A grassland (Danthonia caespitosa) community. Australian Journal of Experimental Agriculture and Animal Husbandry 7, 426–433.
| Crossref | GoogleScholarGoogle Scholar |
Sanderson S. C., Stutz H. C.
(1994) Woody chenopods useful for rangeland reclamation in western North America. : ‘Proceedings: Ecology and Management of Annual Rangelands’. (Eds B. Monsen, S. G. Kitchen)
pp. 374–378. (USDA Forest Service General Technical Report INT-GTR-313: Utah.)
Simes J. J. H.,
Tracey J. G.,
Webb L. J., Dunstan W. J.
(1959) An Australian phytochemical survey. 3: Saponins in eastern Australian flowering plants. CSIRO Bulletin 281, 5–31.
Vallentine, J. F. (1990).
Walsh D.
(2005) Sex lability discovered in Atriplex vesicaria Heward ex Benth. (Chenopodiaceae). Journal of Arid Environments 60, 201–210.
| Crossref | GoogleScholarGoogle Scholar |
Walsh D.,
Sinclair R.,
Andrew M. H., Coleman D.
(2005) The role of the male flower spike as a cue for selective grazing in bladder saltbush. The Rangeland Journal 27, 97–103.
Williams D. G.
(1972) Ecological studies on shrub-steppe of the western Riverina, New South Wales. PhD thesis, The Australian National University.
Williams D. G.
(1979) The comparative ecology of two perennial chenopods.
: ‘Studies of the Australian arid zone. IV: Chenopod shrublands’. (Eds R. D. Graetz, K. M. N. Howes)
pp. 29–40. (CSIRO Publishing: Melbourne, Vic.)
Williams D. G.,
Anderson D. J., Slater K. R.
(1978) The influence of sheep on pattern and process in Atriplex vesicaria populations from the Riverine Plain of New South Wales. Australian Journal of Botany 26, 381–392.
Wilson A. D., Graetz R. D.
(1979) Management of the semi-arid rangelands of Australia.
: ‘Management of semi-arid ecosystems’. (Ed. B. H. Walker)
pp. 83–111. (Elsevier: Amsterdam.)