

Age estimation of striped marlin (Kajikia audax) in the eastern North Pacific using otolith microincrements and fin spine sections
Tamaki Shimose A C and Kotaro Yokawa BA Research Center for Subtropical Fisheries, Seikai National Fisheries Research Institute, Japan Fisheries Research and Education Agency, 148 Fukai-Ohta, Ishigaki, Okinawa 907-0451 Japan.
B National Research Institute of Far Seas Fisheries, Japan Fisheries Research and Education Agency, 5-7-1 Orido, Shimizu, Shizuoka 424-8633, Japan.
C Corresponding author. Email: shimose@affrc.go.jp
Marine and Freshwater Research 70(12) 1789-1793 https://doi.org/10.1071/MF18322
Submitted: 31 August 2018 Accepted: 20 December 2018 Published: 27 February 2019
Journal Compilation © CSIRO 2019 Open Access CC BY-NC-ND
Abstract
The age of striped marlin (Kajikia audax) in days and years (daily and yearly ages) were estimated by counts of otolith microincrements (n = 25) and dorsal fin spine annuli (n = 175) using specimens caught in the tropical eastern North Pacific between September and November 2004. Daily ages of small striped marlin (87.0–145.5 cm lower jaw–fork length) were estimated to range from 81 to 239 days, which indicates that the species in the area grow, on average, to over 100 cm within 4 months. Back-calculated hatch dates were estimated to be from March to July; this period is earlier than the known spawning season in a slightly more northern area. Approximately 20% of these year-0 striped marlin had one or more growth bands in the sectioned spine. They were thought to be false annual growth bands and excluded from yearly age estimation. The estimated yearly age of 175 individuals (87.0–228.4 cm) ranged from 0 to 5 and was dominated by ages-0 to 3 (>90%). Age composition was different among three subregions in the eastern North Pacific.
Additional keywords: billfish, Istiophoridae.
Introduction
The age of billfishes (families Istiophoridae and Xiphiidae) has been estimated using fin spine (or ray) sections for decades (e.g. Hill et al. 1989; Ehrhardt 1992; Speare 2003; Chiang et al. 2004; Kopf et al. 2005). However, early growth rates (up to age-1) of billfishes estimated by otolith microincrements are known to differ to those estimated by fin spines (Prince et al. 1991; DeMartini et al. 2007; Kopf et al. 2011). Recent studies have used otolith microincrements to estimate the early growth of billfishes, and these results have also been used in the estimation of the position of the first-year annual growth band in sectioned fin spines (DeMartini et al. 2007; Kopf et al. 2011; Shimose et al. 2015). This method successfully eliminates counting false annual growth bands formed in younger ages (e.g. Kopf et al. 2011; Shimose et al. 2015). However, otolith microincrements can only be counted up to age-1 (or age-2) in most billfishes (Prince et al. 1991; DeMartini et al. 2007). Furthermore, fin spine sections are still used in the ageing of individuals older than 1 or 2 years (e.g. DeMartini et al. 2007; Kopf et al. 2011; Shimose et al. 2015).
The striped marlin (Kajikia audax) is a large istiophorid billfish species targeted by commercial and recreational fisheries in the Pacific Ocean (Nakamura 1985; Whitelaw 2003). The age and growth of striped marlin, which provide important information for stock assessment, have been estimated in several previous studies (e.g. Skillman and Yong 1976; Melo-Barrera et al. 2003; Kopf et al. 2005), and a recent study provided reliable growth parameters for the western South Pacific population (Kopf et al. 2011). There are several genetically discrete stocks of striped marlin in the Pacific, and the western South Pacific stock is isolated from stocks in the North Pacific (McDowell and Graves 2008). Average body size of striped marlin also differs, and is smaller in the eastern North Pacific (González-Armas et al. 2006) than in the western South Pacific (Kopf et al. 2005). Therefore, the age composition and growth patterns may be different among regions and should be estimated independently. The aim of this study was to estimate the age of striped marlin in the eastern North Pacific simultaneously in days and years (daily and yearly ages). The daily age of young striped marlin was estimated by counting otolith microincrements, and the growth rate was compared with that in the western South Pacific (Kopf et al. 2011). Yearly age of young to adult striped marlin was estimated by counting annual growth bands in sectioned fin spines, and age composition was compared among subregions in the tropical eastern North Pacific.
Materials and methods
Sample collection
In all, 26 otolith and 175 fin spine samples of striped marlin were collected during the longline research cruise by RV Shoyo-Maru in the tropical areas of the eastern North Pacific Ocean (5–18°N, 103–134°W) between September and November 2004. All 26 otolith samples were from same fish used for fin spine samples. The sampling area was divided into three subregions based on previous studies (Shimose et al. 2010, 2013), namely an open ocean area (16–18°N, 118–134°W), a near-continental area (13–16°N, 103–107°W) and a near-equatorial area (5°N, 104–120°W). Bodyweight (BW) was recorded to the nearest 0.05 kg, and the lower jaw–fork length (LJFL) was measured to the nearest 0.1 cm for each specimen. Sex was also determined when possible through visual inspection of gonad morphology for potential use in a future study. However, sexual differences were not examined in the present study because of the insufficient sample size. The LJFL and BW of the specimens used for the otolith microincrement analysis were in the range 87.0–145.5 cm and 2.00–17.40 kg. The LJFL and BW of the specimens used for yearly age estimation based on fin spines were 87.0–228.4 cm and 2.00–78.40 kg. Much larger blue marlin (265 cm LJFL, >160 kg BW) were caught in this research cruise (Shimose et al. 2010, 2013), and the currently used longline gear can catch juvenile (~2.0 kg) to adult (~ > 100 kg) striped marlin.
Observation of otolith microincrements
Right or left sagittal otoliths were used in the otolith microincrement analysis. In this study, transverse sections were observed under light microscopy. The otoliths were embedded in resin and transversely sectioned to ~0.5 mm before being attached to glass slides. These sections were roughly polished until the core appeared on the surface, and polished again with 1-μm abrasive powder. Polished sections were observed under an optical microscope with transmitted light, and microincrements were counted three times (in December 2017 and in June and August 2018) by one reader (T. Shimose) without previous information on the individual specimen (Fig. 1). Otolith microincrements were identified referring to Prince et al. (1991) and Shimose et al. (2015). Otolith microincrements were assumed to be formed daily as in blue marlin (Makaira nigricans; Prince et al. 1991) and striped marlin (Kopf et al. 2011). The precision of the three counts was quantified by the average percentage error (APE; Beamish and Fournier 1981) and the CV (Campana 2001). Many ageing studies have been conducted with CVs of <7.6, which corresponds to an APE of 5.5 (Campana 2001). The median value of the three counts was used as the daily age. Back-calculated hatch dates were compared with the known spawning season off Mexico (24°N, 110°W; González-Armas et al. 2006).
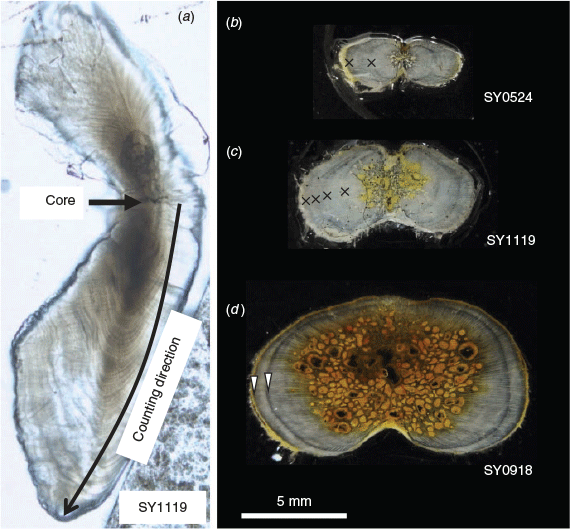
|
Observation of fin spines
Sample processing and terminology of growth bands followed Shimose et al. (2015). The longest spine of the first dorsal fin was boiled to remove tissues and was preserved in dry storage conditions for later analysis. The fin spines were sectioned to ~0.5 mm. The position of sectioning was at a distance of half a condyle base width up from the condyle base (Sun et al. 2002; Chiang et al. 2004). Spine sections were attached to glass slides and photographed under a microscope. The translucent zone was treated as a growth band, and the annual growth band was identified according to Speare (2003) and Kopf et al. (2010, 2011) to avoid the counting of false annual growth bands (Fig. 1). Assumed annual growth bands (distinct translucent growth bands) were counted once by one reader (T. Shimose). Prior to the count, all sections were carefully checked for their growth bands and compared with previous studies (Kopf et al. 2010, 2011). Some specimens aged <1 year by the otolith microincrement count had some translucent growth bands, and they were assumed to have been formed before 1 year of age. The diameter of these growth bands (up to ~8 mm) was considered as a criterion of non-annual growth bands. Periodicity in growth band formation was not assessed in this study because of the limited sampling season.
Results and discussion
Daily age
The sectioned otoliths of striped marlin exhibited microincrements from the core to the edge except for one individual male (141.9 cm). The daily age of the remaining 25 individual striped marlin was successfully estimated (Table 1). The mean APE and CV of three counts in 25 individuals were 4.31 and 5.89 respectively; these values were less than the average values (APE = 5.5, CV = 7.6) reported in the literature (Campana 2001), and imply that the current counts are precise. The number of otolith microincrements of the 25 individuals (87.0–145.5 cm, mean 104.1 cm) ranged from 81 to 239 (mean 124), which indicates that, on average, striped marlin in the area grow over 100 cm in 4 months (Fig. 2). This result of the age–length relationship overlapped with the formerly estimated relationship in the western South Pacific (Kopf et al. 2011), and the growth rate is thought to be similar between the two areas at least up to 6 months. For the one individual male for which otolith microincrements could not be counted, this was due to the low legibility after inflection of the dorsal and ventral arms of the sectioned otolith.

|
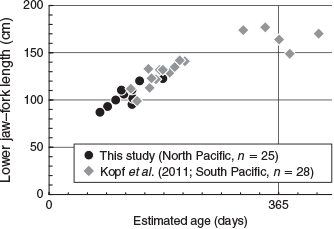
|
Otolith length increases as fish body length increase, and only otolith weight continues to increase with age after growth in body length has almost stopped (Newman et al. 1996). Otolith weight is linked to otolith thickness, which continues to increase after inflection of the dorsal and ventral arms of the otolith in billfishes (Wilson et al. 1991). This means that the ageing of billfish for the same body size but older ages (i.e. slower growing) tends to fail more through counting their daily age from the core to the edge. This can lead to an overestimation of the growth rate because slower-growing individuals (older individuals within the same body size group) are excluded from the estimation.
The back-calculated hatch dates were between 3 March and 17 July and concentrated in May and June (Fig. 3). Maturing and mature females are known to be abundant in July and August in an area at a higher latitude off Mexico (24°N, 110°W; González-Armas et al. 2006) than the present study area (5–18°N, 103–134°W). Back-calculated hatch dates of the present study (May and June) are also earlier than the spawning season off Mexico. Spawning season is possibly early in the lower-latitude area, where temperature starts to increase earlier during the spring to summer transition.
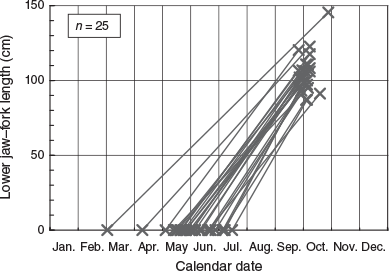
|
Yearly age
Five of 25 observed small individuals (20%) for which daily age was estimated to be less than 365 (<1 year) had some growth bands in their sectioned fin spine (Fig. 1). Therefore, these growth bands (up to ~8 mm in diameter) were not annuli and should not be included in the count as annuli. Similar growth bands that formed before 1 year of age have also been reported in blue marlin (Shimose et al. 2015) and they are thought to be a common phenomenon in billfishes.
Estimated ages of the 175 striped marlin ranged from 0+ to 5+ years, with >90% of samples dominated by 0+ to 3+ years. Older striped marlin (up to 8 years of age) have been reported in the western South Pacific (Kopf et al. 2011), and the specimens used in the present study may not cover the full size range of striped marlin in the eastern North Pacific. However, different age compositions were observed among the three subregions (Fig. 4). In the near-continental area, a significant number of year-0 striped marlin occurred, but there were few year-0 individuals in the open ocean area. Striped marlin aged from 1+ to 3+ years were observed with high frequency in all three areas. Spawning grounds occur near the continent off Mexico (González-Armas et al. 2006) and in the near-continental area (Shimose et al. 2013). The age composition in the near-continental area is thought to reflect the occurrence of both juveniles and adults. Prey items are more abundant in the open ocean area than in the near-continental area (Shimose et al. 2010), and striped marlin start migrating to the open ocean area for feeding after reaching 1 year of age.
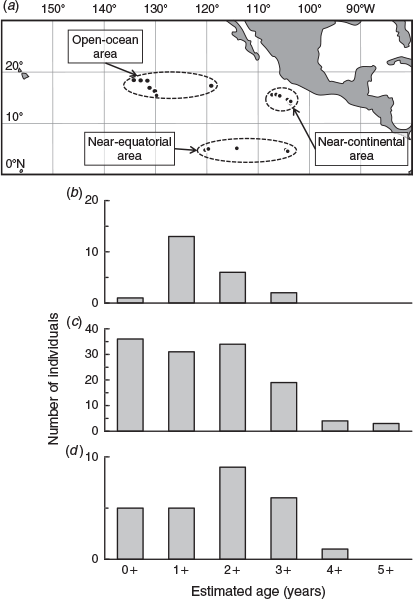
|
The results of this study will contribute to further our understanding of the life history of striped marlin relating to earlier growth and ontogenetic migration in the eastern North Pacific. Growth parameters of striped marlin were not estimated in this study because the sampling coverage was at specific points, both temporally and spatially, and the range of sizes sampled was limited due the sampling method. Further studies on age composition and growth curve estimation of striped marlin are needed based on representative samples in the eastern North Pacific.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This study was funded by a grant from the Japan Fisheries Agency.
Acknowledgements
The authors thank the captain and crew of RV Shoyo-Maru and H. Saito for their cooperation with sampling specimens. The authors also thank Y. Semba (National Research Institute of Far Seas Fisheries) and the anonymous reviewers for their comments on an earlier version of this manuscript.
References
Beamish, R. J., and Fournier, D. A. (1981). A method for comparing the precision of a set of age determinations. Canadian Journal of Fisheries and Aquatic Sciences 38, 982–983.| A method for comparing the precision of a set of age determinations.Crossref | GoogleScholarGoogle Scholar |
Campana, S. E. (2001). Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. Journal of Fish Biology 59, 197–242.
| Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods.Crossref | GoogleScholarGoogle Scholar |
Chiang, W.-C., Sun, C.-L., Yeh, S.-Z., and Su, W.-C. (2004). Age and growth of sailfish (Istiophorus platypterus) in waters off eastern Taiwan. Fishery Bulletin 102, 251–263.
DeMartini, E. E., Uchiyama, J. H., Humphreys, R. L., Sampaga, J. D., and Williams, H. A. (2007). Age and growth of swordfish (Xiphias gladius) caught by the Hawaii-based pelagic longline fishery. Fishery Bulletin 105, 356–367.
Ehrhardt, N. M. (1992). Age and growth of swordfish, Xiphias gladius, in the northwestern Atlantic. Bulletin of Marine Science 50, 292–301.
González-Armas, R., Klett-Traulsen, A., and Hernández-Herrera, A. (2006). Evidence of billfish reproduction in the southern Gulf of California, Mexico. Bulletin of Marine Science 79, 705–717.
Hill, K. T., Cailliet, G. M., and Radtke, R. L. (1989). A comparative analysis of growth zones in four calcified structures of Pacific blue marlin, Makaira nigricans. Fishery Bulletin 87, 829–843.
Kopf, R. K., Davie, P. S., and Holdsworth, J. C. (2005). Size trends and population characteristics of striped marlin, Tetrapturus audax caught in the New Zealand recreational fishery. New Zealand Journal of Marine and Freshwater Research 39, 1145–1156.
| Size trends and population characteristics of striped marlin, Tetrapturus audax caught in the New Zealand recreational fishery.Crossref | GoogleScholarGoogle Scholar |
Kopf, R. K., Drew, K., and Humphreys, R. L. (2010). Age estimation of billfishes (Kajikia spp.) using fin spine cross-sections: the need for an international code of practice. Aquatic Living Resources 23, 13–23.
| Age estimation of billfishes (Kajikia spp.) using fin spine cross-sections: the need for an international code of practice.Crossref | GoogleScholarGoogle Scholar |
Kopf, R. K., Davie, P. S., Bromhead, D., and Pepperell, J. G. (2011). Age and growth of striped marlin (Kajikia audax) in the Southwest Pacific Ocean. ICES Journal of Marine Science 68, 1884–1895.
| Age and growth of striped marlin (Kajikia audax) in the Southwest Pacific Ocean.Crossref | GoogleScholarGoogle Scholar |
McDowell, J. R., and Graves, J. E. (2008). Population structure of striped marlin (Kajikia audax) in the Pacific Ocean based on analysis of microsatellite and mitochondrial DNA. Canadian Journal of Fisheries and Aquatic Sciences 65, 1307–1320.
| Population structure of striped marlin (Kajikia audax) in the Pacific Ocean based on analysis of microsatellite and mitochondrial DNA.Crossref | GoogleScholarGoogle Scholar |
Melo-Barrera, F. N., Félix-Uraga, R., and Quiñónez-Velázquez, C. (2003). Growth and length–weight relationship of the striped marlin, Tetrapturus audax (Pisces: Istiophoridae), in Cabo San Lucas, Baja California Sur, Mexico. Ciencias Marinas 29, 305–313.
| Growth and length–weight relationship of the striped marlin, Tetrapturus audax (Pisces: Istiophoridae), in Cabo San Lucas, Baja California Sur, Mexico.Crossref | GoogleScholarGoogle Scholar |
Nakamura, I. (1985). FAO species catalogue. Vol. 5. Billfishes of the world. An annotated and illustrated catalogue of marlins, sailfishes, spearfishes and swordfishes known to date. FAO Fisheries Synopsis number 125, FAO, Rome, Italy.
Newman, S. J., Williams, D. M., and Russ, G. R. (1996). Age validation, growth and mortality rates of the tropical snappers (Pisces: Lutjanidae) Lutjanus adetii (Castelnau, 1873) and L. quinquelineatus (Bloch, 1790) from the central Great Barrier Reef, Australia. Marine and Freshwater Research 47, 575–584.
| Age validation, growth and mortality rates of the tropical snappers (Pisces: Lutjanidae) Lutjanus adetii (Castelnau, 1873) and L. quinquelineatus (Bloch, 1790) from the central Great Barrier Reef, Australia.Crossref | GoogleScholarGoogle Scholar |
Prince, E. D., Lee, D. W., Zweifel, J. R., and Brothers, E. B. (1991). Estimating age and growth of young Atlantic blue marlin Makaira nigricans from otolith microstructure. Fishery Bulletin 89, 441–459.
Shimose, T., Yokawa, K., and Saito, H. (2010). Habitat and food partitioning of billfishes (Xiphioidei). Journal of Fish Biology 76, 2418–2433.
| Habitat and food partitioning of billfishes (Xiphioidei).Crossref | GoogleScholarGoogle Scholar | 20557600PubMed |
Shimose, T., Ashida, H., and Yokawa, K. (2013). Sex ratio and reproductive condition of four istiophorid billfishes in tropical regions of the eastern North Pacific Ocean: with special reference to striped marlin Kajikia audax (Philippi, 1887). Journal of Applied Ichthyology 29, 1247–1251.
| Sex ratio and reproductive condition of four istiophorid billfishes in tropical regions of the eastern North Pacific Ocean: with special reference to striped marlin Kajikia audax (Philippi, 1887).Crossref | GoogleScholarGoogle Scholar |
Shimose, T., Yokawa, K., and Tachihara, K. (2015). Age determination and growth estimation from otolith micro-increments and fin spine sections of blue marlin (Makaira nigricans) in the western North Pacific. Marine and Freshwater Research 66, 1116–1127.
| Age determination and growth estimation from otolith micro-increments and fin spine sections of blue marlin (Makaira nigricans) in the western North Pacific.Crossref | GoogleScholarGoogle Scholar |
Skillman, R. A., and Yong, M. Y. Y. (1976). Von Bertalanffy growth curves for striped marlin, Tetrapturus audax, and blue marlin, Makaira nigricans, in the central North Pacific Ocean. Fishery Bulletin 74, 553–566.
Speare, P. (2003). Age and growth of black marlin, Makaira indica, in east coast Australian waters. Marine and Freshwater Research 54, 307–314.
| Age and growth of black marlin, Makaira indica, in east coast Australian waters.Crossref | GoogleScholarGoogle Scholar |
Sun, C.-L., Wang, S.-P., and Yeh, S.-Z. (2002). Age and growth of swordfish (Xiphias gladius L.) in the waters around Taiwan determined from anal-fin rays. Fishery Bulletin 100, 822–835.
Whitelaw, W. (2003). Recreational billfish catches and gamefishing facilities of Pacific Island nations in the Western and Central Pacific Ocean. Marine and Freshwater Research 54, 463–471.
| Recreational billfish catches and gamefishing facilities of Pacific Island nations in the Western and Central Pacific Ocean.Crossref | GoogleScholarGoogle Scholar |
Wilson, C. A., Dean, J. M., Prince, E. D., and Lee, D. W. (1991). An examination of sexual dimorphism in Atlantic and Pacific blue marlin using body weight, sagittae weight, and age estimates. Journal of Experimental Marine Biology and Ecology 151, 209–225.
| An examination of sexual dimorphism in Atlantic and Pacific blue marlin using body weight, sagittae weight, and age estimates.Crossref | GoogleScholarGoogle Scholar |