

Assessment of humpback whale swimming speeds in two eastern Australian bays
H. Kela A B C , J. de Bie A B , K. H. W. Paas D , S. Stack E F , W. Franklin G H I , T. Franklin G H I and J.-O. Meynecke A B C *
A B C , J. de Bie A B , K. H. W. Paas D , S. Stack E F , W. Franklin G H I , T. Franklin G H I and J.-O. Meynecke A B C *
A
B
C
D
E
F
G
H
I
Abstract
Humpback whales (Megaptera novaeangliae) are capital breeders, travelling several thousand kilometres between their breeding and feeding sites annually. Along the migration route, humpback whales utilise locations such as Hervey Bay and the Gold Coast bay for resting.
Here we used sightings data from 2066 boat-based surveys spanning 11 years.
To compare and analyse the sighting data and evaluate the swimming speed in Hervey Bay and the Gold Coast bay.
During southward migration, swimming speeds observed in Hervey Bay and the Gold Coast bay for both pods with and without calves were slower than are migration speeds reported in literature for each pod type. Whales swam faster in the Gold Coast bay (mean = 4.14 km h−1) than Hervey Bay (mean = 3.32 km h−1); however, the effect of location on swimming speed was small. In Hervey Bay, swimming speeds of pods without calves were faster (mean = 3.48 km h−1) than swimming speeds of pods with calves (mean = 3.15 km h−1).
Our findings contribute to the understanding of swimming speeds in different humpback whale cohorts and their movements in nearshore environments.
Understanding the importance of the role of humpback whale swimming speed in nearshore environments for assessment of habitat use can help management of such areas for effective protection of humpback whales.
Keywords: Australia, cetaceans, citizen science, conservation, humpback whale, migration, resting area, swimming speed.
Introduction
Humpback whale (Megaptera novaeangliae) is a cosmopolitan species, migrating seasonally between its summer feeding grounds in high latitudes and its winter breeding grounds in the tropics and subtropics (Dawbin 1966; Clapham and Mead 1999). The breeding stock E1 southward migration begins in late July to early August (Paterson 1991), largely following the eastern Australian coastline from the breeding sites in the Great Barrier Reef to Eden, before swimming to their feeding sites in Antarctica (Andrews-Goff et al. 2018). During migration, mothers and calves seek shallow and warm waters to rest, and minimise disturbance from male humpback whales and predators (Ersts and Rosenbaum 2003; Rasmussen et al. 2007; Bruce et al. 2014; Franklin et al. 2018, 2021; Stack et al. 2020). Model estimates have suggested mothers and calves rest, on average, every second day of their migration (Braithwaite et al. 2015).
Along the eastern coast of Australia, previously described resting areas, such as Hervey Bay and Jervis Bay, carry particular importance to mothers and calves of the E1 population (Bruce et al. 2014; Franklin et al. 2021). Between 23 and 46% (mean = 34%, s.d. = 5.3%) of the estimated eastern Australian migration enters Hervey Bay from early August to mid-October each year (Franklin 2014). The average residency time in the bay is 1–3 days (Corkeron et al. 1994; Franklin 2012; Stack et al. 2020); however, extended stays of up to 40 days (Stack et al. 2020) and 51 days (Franklin 2012) have been reported.
More recently, the importance of the Gold Coast bay for resting mothers and calves has been recognised through the analysis of surface behaviours (McCulloch et al. 2021). It has been suggested that mother and calf pairs account for 11.5–30% of all sightings annually in the Gold Coast bay, peaking at 40% in October (Valani et al. 2020).
Evaluation of swimming speed of humpback whales can provide valuable insights into aspects of their behaviour and ecology. For instance, it has been shown that in calm conditions, resting whales drift or move slowly on the surface (Bejder et al. 2019; Sprogis et al. 2020a); several swimming-related behavioural states, such as ‘directed travel’ and ‘mill’, have been identified on Hawaiian breeding grounds (Henderson et al. 2018); and behavioural responses to disturbances as a result of swim-with-whale tourism were quantified (Sprogis et al. 2020b). Swimming speeds have also been shown to depend on age, with calves unable to move fast, resulting in mother–calf pairs being slower than whales without a calf during migration (Chittleborough 1953; Noad and Cato 2007), and at breeding areas (Currie et al. 2021; Villagra et al. 2021).
Finally, swimming speed varies on the basis of the stage of migration, with those migrating northward travelling faster than southward-migrating individuals (Salgado Kent et al. 2012). Swimming speeds in feeding and breeding areas have been shown to be slower than those estimated during migration. Whales spend considerable amount of time resting on the surface (Bejder et al. 2019; Ejrnæs and Sprogis 2021), drifting with the current at a speed of 0.3 m s−1 (<1.1 km h−1) (Sprogis et al. 2020a). A multi-day satellite tagging study found that migrating whales from various cohorts slowed down to an average speed of 2.19 km h−1 after reaching the feeding grounds in Antarctica (Andrews-Goff et al. 2018). Similar swimming speeds were observed from tagged humpback whales in New Caledonian breeding grounds averaging 2.01 km h−1 (Garrigue et al. 2015).
Slow movement patterns at the surface pose several risks to humpback whales. In addition to the increasing popularity of swimming with whales (Sprogis et al. 2020b), these can lead to vessel collisions and subsequent injuries or mortality (Schoeman et al. 2020; Mayaud et al. 2022), or prolonged presence in areas where fishing gear has been deployed (Johnson et al. 2005; Hamilton and Baker 2019). An improved understanding of humpback whale swimming speeds and the nearshore habitat utilisation is thus important for recognising potential hazards and ensuring the protection of vulnerable mother–calf pairs during their migration pathways.
Long-term monitoring of migratory animals can be difficult, and systematic research is not always feasible. Citizen science allows large-scale data collection over multiple years, and the use of citizen science data has become more prevalent in recent years (Conrad and Hilchey 2011; Franklin et al. 2020). It has many applications, including research into humpback whale behaviours (McCulloch et al. 2021), abundance (Tonachella et al. 2012; Pirotta et al. 2020), or distribution patterns (Bruce et al. 2014). Citizen science projects in Hervey Bay and the Gold Coast bay have been collecting humpback whale sighting data since 1992 and 2010 respectively. These long-term datasets were used to analyse and compare the temporal trends in sighting data as well as swimming speed at Hervey Bay and the Gold Coast bay. Specifically, we aimed to (1) analyse overall trends in sighting data by comparing trends within season, and between seasons in Hervey Bay and the Gold Coast bay, (2) investigate the effect of calf presence on swimming speed of a humpback whale pod within and between locations, and (3) compare the swimming speeds with known travel times based on sighting data.
Methods
Survey sites
The survey sites, Hervey Bay and the Gold Coast bay, are located in south-eastern Queensland, Australia, between 24.5 and 28.5°S (Fig. 1). Hervey Bay is a bowl-shaped bay separated from the Coral Sea on the east by K’gari. The bay covers an area of ~4000 km2, with the opening facing north, with a width of ~80 km. The bay has a mean depth of 20 m (Ribbe 2014), and calm waters being protected by K’gari (Fig. 1).
Map showing study sites in south-eastern Queensland, Australia. Surveys were conducted on the eastern side of Hervey Bay and in the vicinity of the Gold Coast seaway.
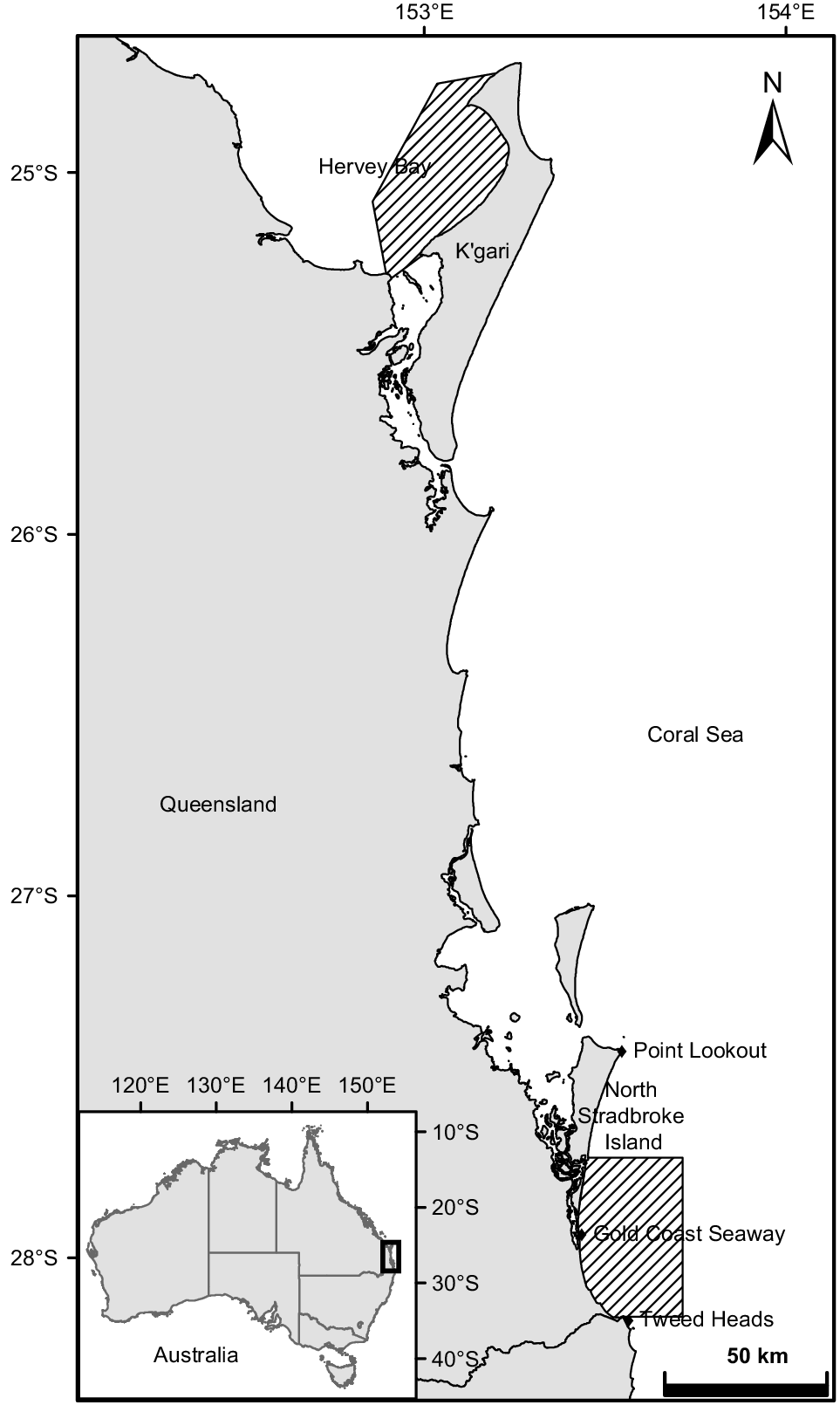
The study site is located 300 km south of Hervey Bay, off the Gold Coast shore. The study site spans roughly 48 km from Jumpinpin to Tweed Heads at the border of Queensland and New South Wales and extends 26 km to the sea from the Gold Coast seaway. It has an area of ~1000 km2 and a mean depth of 40 m (Fig. 1). The waters form an open embayment (Bird 2008) that provides shelter from the dominant south-easterly trade winds in the southern part of the region.
Definitions
For the analyses, pod is defined as any group of whales sighted within 100 m of each other and moving in the same general direction. Individual is defined as any individual whale sighted, including all whales from calves to mature adults, whereas calf refers to any small whale (including yearling) (<10 m) with a constant close association with a whale approximately twice their size, assumed to be the mother. ‘Calf pod’ is used when referring to a pod that includes at least one calf in the composition, whereas ‘non-calf pod’ refers to any pod that does not contain a calf. Follow refers to observers encountering and focusing on one pod or individual, recording their behaviour and movements for a period of time, usually less than 30 min. Survey consists of one or more follows within a specific observational period that varied between organisations (2.5 to 9 h). ‘Humpback whale - season’ is considered the period of time between early June and late October when humpback whales are migrating through the waters around the study locations to the breeding grounds and back to Antarctica. Swimming speed is defined as the calculated rate of movement between two sighting points.
Data collection
In Hervey Bay, humpback whale sighting data were collected by The Oceania Project on motorised dedicated research vessels up to 6 days per week from 2010 to 2017 (excluding 2014), and by Pacific Whale Foundation (PWF) on a motorised dedicated research vessel from 2010 to 2013, and aboard whale watching vessels from 2014 to 2020. The Oceania Project conducted surveys in August, September, and October, utilising three different catamarans ranging in length from 11.92 to 13.23 m and powered by two 27–503-horsepower (HP) (~20–375-kW) engines. Average time spent at sea for each survey was 9 h. These surveys were performed on a ‘first pod available’ basis, where the boat would travel in a random direction in the morning, with a minimum of six surveyors scanning for any signs of whales. If location information for a humpback whale pod was available from a local whale-watching operator, the boat would head in the direction of the pod until approached, or a different pod was sighted en route. PWF surveys were performed during the southward migration every day during daylight hours, up to twice per day, weather permitting, in Beaufort sea states of ≤4. Sighting data were collected on rigid hull inflatable vessels of 5.3 or 5.8 m in length, equipped with a 70-HP (~52-kW) engine, 12.5-m vessel with four 300-HP (~223-kW) engines, and 20-m vessel with two 350-HP (~260-kW) engines. Surveys commenced once the boat left the Hervey Bay marina, averaging 3 h per survey. Humpback whales were searched opportunistically, with surveyors scanning around the vessel with a naked eye and binoculars for signs of humpback whales. Once a pod was located by surveyors, the vessel approached and followed it, while keeping a distance over 100 m. The vessel speed and direction were recorded to match the movement of the pod. Date, time, and global positioning system (GPS) coordinates of the vessel were recorded at the start and end of observations. Additionally, pod composition, whale size, and sex, when possible, were recorded.
The Gold Coast bay sighting data were collected by Humpbacks and High-rises (HHR), an non-governmental organisation based on the Gold Coast, from 2010 to 2020 throughout the whole migration. Two or three trained observers collected data aboard whale-watching vessels during daylight hours every day, depending on weather conditions and observer availability. Vessel length varied from 15 to 24 m, and was powered by 510–650-HP (~380–484-kW) engines. Humpback whales were searched opportunistically with a naked eye and binoculars once the vessel had travelled through Southport seaway (Fig. 1). The average observation time at sea for each trip was 2.5 h. Once a pod was located, the vessel approached the pod, observations commencing once the pod was within 300 m of the vessel. The date, time and the GPS location of the pod was recorded at the start of observation, and on departure. Additionally, the pod composition, whale size and sex were recorded, when possible.
Research activities were authorised under permit and animal ethics authority. The Hervey Bay data are covered by permits associated with previously published data (e.g. Stack et al. 2020; Franklin et al. 2021).
Migration directionality in the Gold Coast bay
Because humpback whales generally do not enter Hervey Bay during the northward migration (Paterson 1991), the Gold Coast bay sighting data were divided into northward and southward migration to allow better comparisons between the sites. Bearing of humpback whale pods sighted in the Gold Coast bay were calculated on the basis of the GPS location of the vessel at the start and end of each follow to distinguish between southward- and northward-migrating pods. A pod was considered to be travelling south if the bearing was between 90 and 270°, and north for other values (Noad and Cato 2007). All bearings over the 11-year period were then used to estimate the day when the migration direction begins to switch from north to south for the majority of pods. This information was used to divide the Gold Coast bay humpback whale sightings into northward- and southward-migration data.
Sightings per unit effort
Data from The Oceania Project and PWF were combined into one Hervey Bay dataset. Because survey effort varied slightly across seasons and daily survey hours between the organisations, sightings per unit effort (SPUE) was calculated to minimise the effect of variable effort across sites, and between years. SPUE was calculated separately for pods, individuals, and calves at each location for each day of the migration, by dividing the number of sightings in each of these three categories by the effort hours per day. Surveys were undertaken in Hervey Bay and the Gold Coast bay at least every 3 days; thus, the survey days were grouped into 3-day intervals. For those intervals, average SPUE was calculated for seasonal analysis. The 3-day averages are hereafter referred to as ‘blocks’, with block numbers referring to the first day of the year included in the block; e.g. 3–5 January would be block Number 3. Average SPUE per each survey year was calculated for pods, individuals, and calves for both sites, allowing comparisons between seasons.
Swimming speed
Swimming speeds for all pods were estimated using the track duration, and vessels start and end location for each observation (e.g. see Noad and Cato 2007; Craig et al. 2014). The calculated swimming speeds here represent the minimum constant swimming speed.
Data were grouped by calf presence in each location, and whale-sighting records without valid beginning and end times or GPS locations for the follow were excluded from swimming-speed analyses. All resting whales that drifted were not included. Some uncertainty was introduced by using the vessel GPS location indicative of the whale location, but no more than ±300 m. The effect of uncertain GPS locations was minimised by including only observations longer than 10 min in the swimming-speed calculation. Resulting speeds over 15 km h−1 (n = 38) were considered unreliable because they were likely to be a result of inaccuracies in recording GPS location or time, and were excluded from the analyses on the basis of previously published maximum swimming speeds (e.g. Tyack 1983; Frankel et al. 1995). Humpback whales have been documented to change swim directions underwater while feeding (Friedlaender et al. 2009; Burrows et al. 2016). Calculating the swimming speeds using only track start and end method is likely to underestimate the swimming speed because it assumes that whales swim in a straight trajectory.
A pod was assumed to be migrating south if it had a southward bearing and it was sighted after the day most pods were observed to be migrating south. Similarly, pods with a bearing north when most pods were migrating north were assumed to be northward-migrating pods. These definitions were used for comparisons of swimming speed between north- and south-migrating calf pods and non-calf pods sighted in the Gold Coast bay. Because humpback whales rarely enter Hervey Bay during northward migration (Paterson 1991), only southward-migrating calf pods and non-calf pods were considered in the analyses between Hervey Bay and the Gold Coast bay.
Kruskal–Wallis H-tests were used for all statistical analyses of swimming speed between and within the study locations, owing to violations of normality and homogeneity. The tests were followed with a Dunn’s multiple comparison post hoc test with a Bonferroni-adjustment to compare all pairs of groups. The effect size was calculated for all Kruskal–Wallis tests. All statistical testing was performed in R (ver. 2.11, R Foundation for Statistical Computing, Vienna, Austria, see https://www.r-project.org/), using package stats (ver. 4.3.2, see https://stat.ethz.ch/R-manual/R-devel/library/stats/html/00Index.html), for the Kruskal–Wallis tests and the package rstatix (ver. 0.7.2, see https://cran.r-project.org/package=rstatix) for Dunn’s tests and effect-size calculations.
Results
Migration directionality in the Gold Coast bay
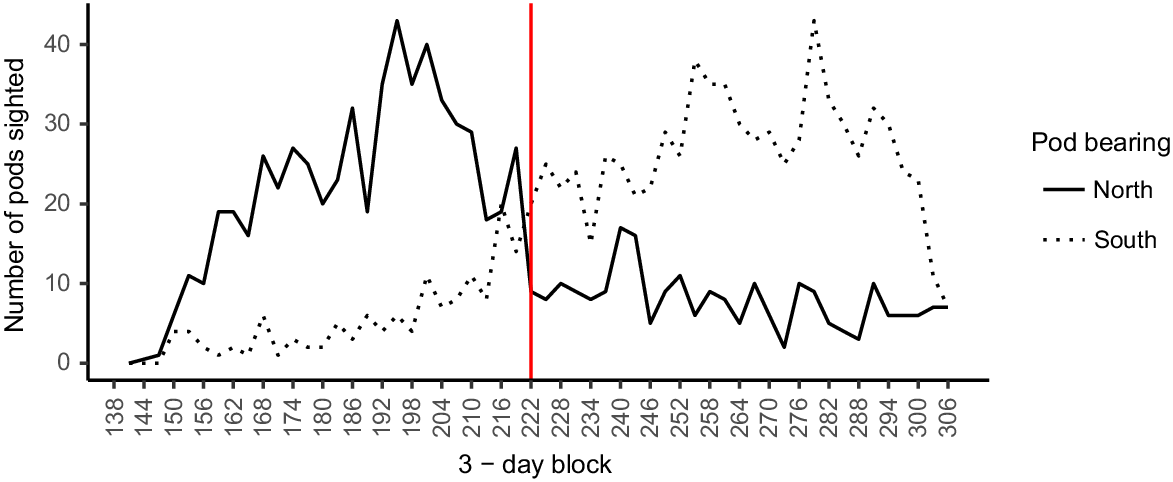
The average northward migration between 2010 and 2020 peaked in mid-July (Block 195), switching to predominantly southward-migrating humpback whales in early- to mid-August (Block 222) (Fig. 2). The peak of southward migration occurred in early October (Block 279). On the basis of these estimates, all pods were considered travelling south from 10 August (start of Block 222) until the end of the season, and north at other times, for analysis of trends in the Gold Coast bay survey data. Only records of pods with bearing north before 10 August and records with bearing south from 10 August were considered in the swimming-speed analyses.
Survey effort
During the 11-year period of data collection, 961 and 1105 surveys were undertaken in Hervey Bay and the Gold Coast bay respectively (Supplementary Table S1). The Hervey Bay survey effort was greater, totalling 3729 h of surveys during the southward migration, compared with a total of 2762 h of surveys in the Gold Coast bay, 1225 h over northward migration, and 1537 h over the southward migration. The effort varied each year between 123 and 525 h in Hervey Bay, and between 50 and 200 h during northward migration and between 29 and 207.5 h during southward migration in the Gold Coast bay. First day of surveys was between 21 May and 18 June each year in the Gold Coast bay, and, in Hervey Bay, between 13 July and 26 August, depending on the year.
Over four times as many humpback whales were sighted in Hervey Bay as in the Gold Coast bay during southward migration during the 11-year period (Table S1). The whale sightings in Hervey Bay were concentrated within 20 km of the coast of K’gari. In the Gold Coast bay, most of the whale sightings were within 15 km of the Gold Coast seaway. On average, the distance covered during a follow was shorter in Hervey Bay (1.2 km) than Gold Coast bay (north 3.4 km, south 2.7 km). Similarly, the follow duration, on average, was shorter in Hervey Bay (20 min) than in the Gold Coast bay (north 41 min, south 43 min).
Sighting per unit effort
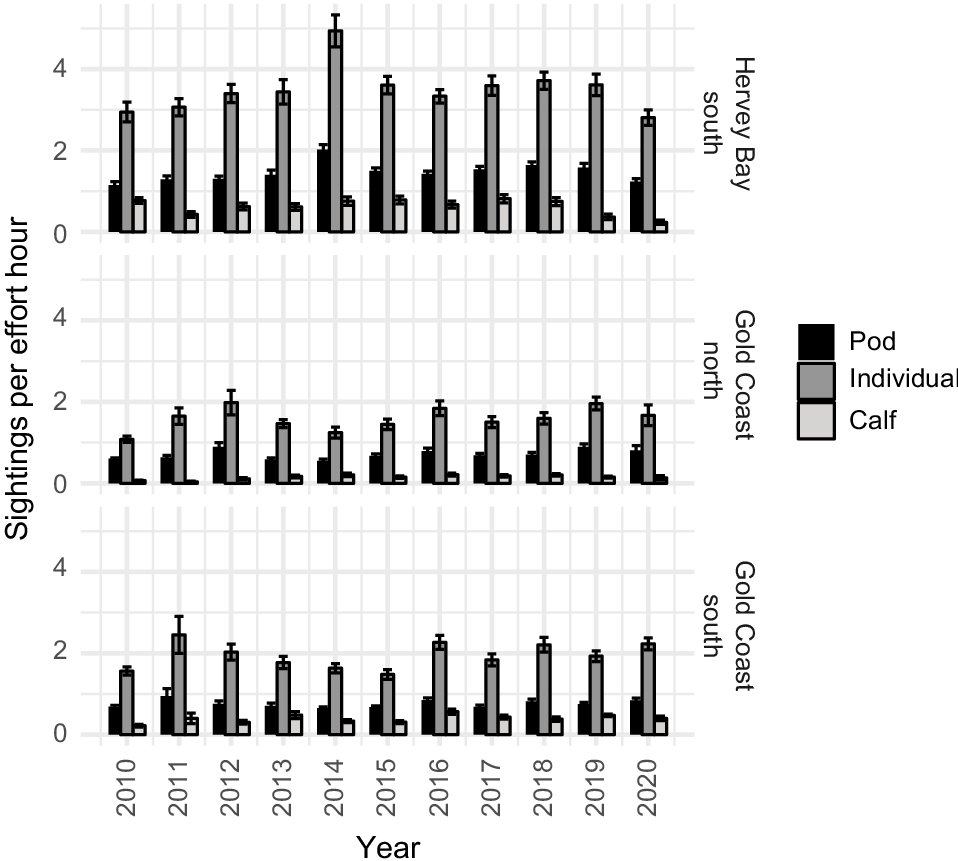
SPUE was higher for individuals, pods, and calves in Hervey Bay than in the Gold Coast bay each year during the study period (Fig. 3). In Hervey Bay, the average SPUE between 2010 and 2020 was 1.49 for pods, 3.55 for individuals, and 0.64 for calves. In the Gold Coast bay, the average SPUE across the years during northward migration was 0.71 for pods, 1.58 for individuals, and 0.15 for calves, and during southward migration 0.75 for pods, 1.92 for all individuals, and 0.40 for calves. Highest average SPUE for pods and individuals was observed in 2014 in Hervey Bay, and in 2011 in the Gold Coast bay during the southward migration. Northward migration in the Gold Coast bay had the highest SPUE for pods in 2019, and in 2012 for individuals. Interestingly, highest SPUE for calves was in 2016 for both northward and southward migration in the Gold Coast bay and 2017 in Hervey Bay.
Average SPUE per year for pods, individuals and calves surveyed in Hervey Bay and the Gold Coast bay. The error bars indicate one standard error around the mean.
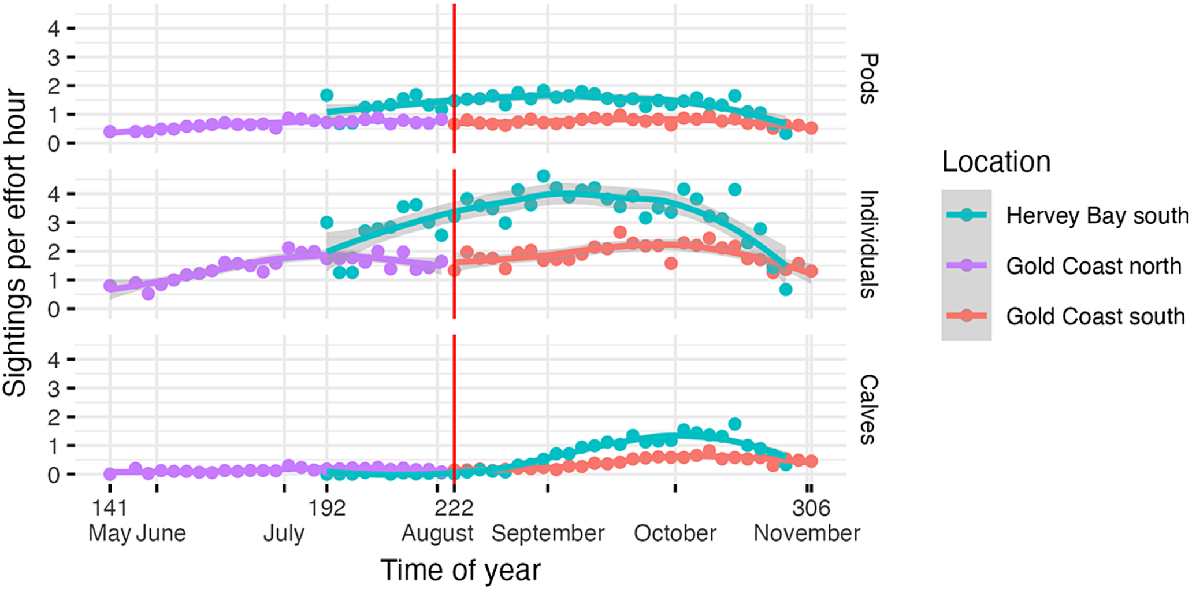
In both locations, the average SPUE varied similarly within season, although the SPUE was higher in Hervey Bay than in Gold Coast bay throughout the southward migration (Fig. 4). Pod SPUE showed some variability during the season in Hervey Bay, stayed almost constant in the Gold Coast bay throughout the migration. The average SPUE for individuals showed more fluctuations within the season. In the Gold Coast bay, the northward migration peaked at the start of July, whereas the southward migration reached its peak in early September in Hervey Bay, and by mid-September in the Gold Coast bay. SPUE for calves was close to zero prior to late August in Hervey Bay, increasing until beginning of October, after which the average calf SPUE slowly tapered. A similar, albeit delayed and less prominent, trend could also be observed in the Gold Coast bay. SPUE for calves slowly increased from the start of August until early October, after which it slowly started to decrease towards the end of the whale season.
Line graphs with LOESS smoothing and 95% confidence intervals (grey shading) illustrate the seasonal variation of average SPUE of pods, individuals, and calves per 3-day period, surveyed during southern migration in Hervey Bay, and both southward and northward migration in the Gold Coast bay between 2010 and 2020. Note that Hervey Bay 192 is a 1-day period. The red line indicates the transition from northern to southern migration.
Swimming speed
Between 2010 and 2020, the mean annual swimming speed was generally higher in the Gold Coast bay (4.14 km h−1) than in Hervey Bay (3.32 km h−1), Kruskal–Wallis H(1) = 103.86, N = 3504, P < 0.001, η2 = 2.94 × 10−2 (Fig. 5). Average swimming speeds were usually lower for calf pods than non-calf pods within both study sites. Furthermore, there was more variability in means among years before 2015 than after, especially in non-calf pods in the Gold Coast bay.
Mean yearly swimming speed of calf pods and non-calf pods in Hervey Bay and Gold Coast bay between 2010 and 2020. Error bars represent one standard error of the mean.
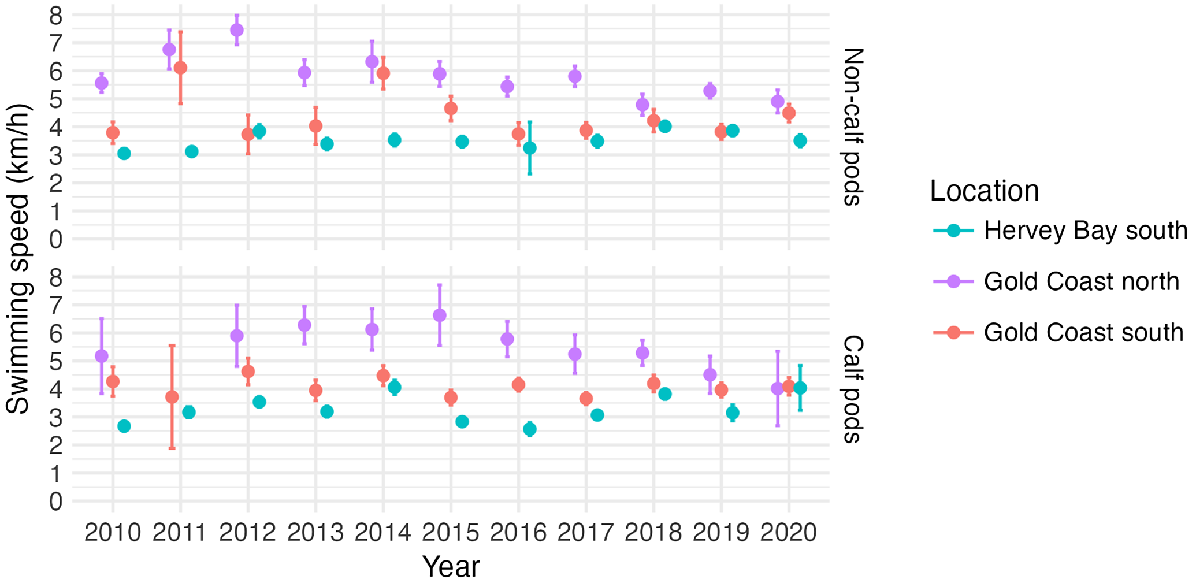
There was a difference in swimming speeds between calf pods and non-calf pods in the Gold Coast bay and Hervey Bay (Kruskal–Wallis H(3) = 120.27, N = 3504, P < 0.001, η2 = 3.35 × 10−2). Dunn post hoc test showed that there was difference in swimming speed between calf pods in the Gold Coast bay and Hervey Bay, and between non-calf pods in the Gold Coast bay and Hervey Bay (Table 1). Additionally, calf pod and non-calf pod swimming speeds in Hervey Bay were different. However, no difference was detected between the Gold Coast bay calf pod and non-calf pod swimming speeds. The effect of location and calf presence was small (η2 < 0.06) on all comparisons between the groups.
| n | Mean | Median | Dunn’s post hoc | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Z | P | η2 | |||||||
| Between sites | Non-calf pods | HB | 1424 | 3.48 | 3.15 | 5.73 | <0.001 | 0.018 | |
| GC south | 326 | 4.26 | 3.97 | ||||||
| Calf pods | HB | 1385 | 3.15 | 2.84 | 8.76 | <0.001 | 0.044 | ||
| GC south | 369 | 4.04 | 4.04 | ||||||
| Within sites | Non-calf pod | HB | 1424 | 3.48 | 3.15 | −4.05 | <0.001 | 0.005 | |
| Calf pod | 1385 | 3.15 | 2.84 | ||||||
| Non-calf pod | GC north | 415 | 5.63 | 5.77 | −0.45 | ns | −0.001 | ||
| Calf pod | 113 | 5.52 | 5.49 | ||||||
| Non-calf pod | GC south | 326 | 4.26 | 3.97 | 0.11 | ns | −0.001 | ||
| Calf pod | 369 | 4.04 | 4.04 | ||||||
| Between northward and southward migration at the GC | Non-calf pod | Northward migration | 415 | 5.63 | 5.77 | −8.22 | <0.001 | 0.082 | |
| Southward migration | 326 | 4.26 | 3.97 | ||||||
| Calf pod | Northward migration | 113 | 5.52 | 5.49 | −5.64 | <0.001 | 0.071 | ||
| Southward migration | 369 | 4.04 | 4.04 | ||||||
GC, Gold Coast bay; HB, Hervey Bay.
The swimming speeds of north- and south-migrating humpback whales in the Gold Coast bay were different between calf pods and non-calf pods (Kruskal–Wallis H(3) = 116.6, n = 1223, P < 0.001, η2 = 9.32 × 10−2). Calf pods were faster swimming north than south, and similarly non-calf pods swam faster north than south (Table 1). Swimming direction had a moderate effect on swimming speed. Speeds of calf pods and non-calf pods were statistically similar when migrating in the same direction.
Discussion
This study used over a decade of humpback whale observation data to analyse temporal trends in sightings (SPUE) and swimming speeds in Hervey Bay, and the Gold Coast bay. We found that SPUE for individuals, pods, and calves was higher in Hervey Bay, but variation of this variable was similar within seasons at both locations. Swimming speeds in Hervey Bay were slower than those observed in the Gold Coast bay, and at both sites, these were slower than are the published swimming speeds during migration.
We found that mean swimming speed for south-migrating non-calf pods was 4.27 km h−1 in the Gold Coast bay and 3.48 km h−1 in Hervey Bay, whereas calf pod swimming speeds were 4.04 km h−1 in the Gold Coast bay and 3.15 km h−1 in Hervey Bay. These mean swimming speeds are in line with previous observations, albeit slower (Table 2). Comparable mean swimming speeds for southward-migrating humpback whales in Western Australia and a different part of eastern Australia were found to be between 3.58 and 6.48 km h−1, with those for calf pods averaging slower at 4.67 km h−1 than did non-calf pod speeds at 5.32 km h−1 (Noad and Cato 2007; Salgado Kent et al. 2012; Sprogis et al. 2020b). Hervey Bay and Gold Coast bay are both used by resting whales (McCulloch et al. 2021), and the associated surface movements can explain slower swimming speeds within the study areas than elsewhere on the migration route.
| Location | Reference | Speed (km h−1) | Cohort | N | Habitat | Migration direction | Method | Distance to vessel | |
|---|---|---|---|---|---|---|---|---|---|
| Eastern coast | Noad and Cato (2007) | 4.17 | Pods without calves | 87 | Migration | S | Land-based survey and acoustic tracking | – | |
| Noad and Cato (2007) | 3.58 | Pods with calves | 28 | Migration | S | Land-based survey and acoustic tracking | – | ||
| Pirotta et al. (2016) | 7.2 | 57 | Migration | N | Land-based survey | – | |||
| Western coast | Salgado Kent et al. (2012) | 4.07 | 44 | Migration | S | Boat-based survey | NA | ||
| Sprogis et al. (2020b) | 5.76 | 86 | Migration | S | Boat-based survey | ~200 m | |||
| Chittleborough (1953) | 9.19 | Pods without calves | 6 | Migration | N | Aerial | – | ||
| Salgado Kent et al. (2012) | 5.65 | Migration | N | Boat-based survey | NA | ||||
| Chittleborough (1953) | 7.56 | Pods with calves | 16 | Migration | N | Aerial | – | ||
| Sprogis et al. (2020a) | 1.08 | Pods with calves | 13 | Nursing ground | – | UAV | – |
Another possible factor influencing swimming speed is the level of human interactions and associated disturbances. For instance, the use of motorised vessels during data collection may have influenced the swimming speeds, because humpback whales have shown to speed up and change course because boating activities (Senigaglia et al. 2016; Sprogis et al. 2020a). In Western Australia, an increase in swimming speed was linked with noise levels of vessels, with high (172 dB re 1 μPa RMS) levels of vessel noise provoking a change in resting duration and swimming speed. Conversely, boat disturbances and vessel noise can induce changes in dive patterns, with whales seeking shelter elsewhere, leading to slower overall swimming speeds (Sprogis et al. 2020a). In the two study sites, operators adhering to vessel speed and approach guidelines (Commonwealth Regulations 2019) are unlikely to have influenced swimming speeds of the whales they follow. The use of boat-based data may further have resulted in a slight overestimation of whale swimming speed owing to swimming and diving under water taking place outside the observer’s field of vision; however, the similar methods used at both locations and the large amount of data available ensured that the swimming speeds between the study sites are comparable, as well as the interpretation of the statistical tests employed. Finally, humpback whales are known to avoid currents flowing in the opposite direction (Findlay et al. 2011), and there is evidence of humpback whales swimming slower in high wind speeds (Kavanagh et al. 2017). The effects of surface currents on humpback whale movements were out of scope for the analyses here; however, they may be an interesting area for future research.
There was a difference between swimming speeds in both sites, with whales swimming faster in the Gold Coast bay. This may be related to the local geography, because Hervey Bay is an enclosed bay that requires deviating from the main migratory pathway to enter (Paterson 1991), and is known to be utilised by mother–calf pairs (Franklin et al. 2021), whereas the Gold Coast bay as an open embayment is less sheltered, allowing migration through the study site. Whales have been estimated to reside in Hervey Bay for up to several weeks (Franklin 2014; Stack et al. 2020), whereas anecdotal reports and sighting data suggest residency periods of 1–2 days in the Gold Coast bay. Faster swimming speeds may reflect whales sighted off the Gold Coast bay migrating through the area without stopping, or only staying for a short period of time. Additionally, activities differ between the locations, with higher proportion of agonistic behaviour, breaching and social communication being observed in the Gold Coast bay, whereas non-aggressive social behaviour was more common in Hervey Bay (McCulloch et al. 2021). Non-agonistic pods, present in Hervey Bay, have been associated with a slower swimming speed (Franklin et al. 2021). Pod size may also influence swimming speed, and this variable could be incorporated in future studies.
Significant difference in swimming speed between cohorts was found in Hervey Bay, with non-calf pods swimming faster, which aligns with previous research (Chittleborough 1953; Noad and Cato 2007; Currie et al. 2021). However, no difference was found between calf pod and non-calf pod speeds in the Gold Coast bay when the pods were travelling in the same direction. Additionally, we found that whales swimming north in both cohorts were faster in the Gold Coast bay than were whales swimming south. Whereas previous literature on humpback whale speeds in Australia has shown northbound speeds to be generally faster (5.65– 9.2 km h−1) (Chittleborough 1953; Salgado Kent et al. 2012) than southbound speeds (4.07–6.48 km h−1) (Salgado Kent et al. 2012; Sprogis et al. 2020a), a study on singing humpback whales reported slower northbound speeds (Noad and Cato 2007). For singers, such a strategy would allow more access to females on the way to the breeding grounds (Noad and Cato 2007). Incorporation of acoustics would be an interesting avenue of future research on humpback whales in Hervey Bay and the Gold Coast bay.
SPUE was higher each year and throughout the season for all categories in Hervey Bay and a higher proportion of south-migrating whales were observed from 10 August onward. Similarly, Paterson (1991) observed more whales travelling south than north from mid-August. The lack of a clear peak in the southbound SPUE at both locations for individuals and pods is likely to be a result of calf pods being the last cohort to leave the breeding grounds and start their migration. Sightings data from The Oceania Project generally concentrated on the peak abundance times and had longer effort per day than did PWF and HHR, resulting in sampling bias. This is likely reflected in higher SPUE for Hervey Bay and also explains the yearly variation of SPUE here, especially before 2015. For the Gold Coast bay, survey effort was low here in some years (2011 and 2012), and the timing of these surveys close to peak abundance explains higher values for SPUE in 2011 and 2012 than in the adjacent years.
It has been suggested that as the humpback whale population increases, so does the use of habitats previously deemed less important, especially by mothers and calves (Bruce et al. 2014). Presently, it is unclear what proportion of mother and calf pairs visiting Hervey Bay also spend time in the Gold Coast bay during their southward migration. Most whales are expected to continue their southward migration from Hervey Bay to the Gold Coast bay following the Australian coastline. There is some evidence of whales departing the coastal areas to continue their swim to New Zealand (Garrigue et al. 2020; Andrews-Goff et al. 2023) or New Caledonia; however, the rate of exchange among the eastern Australian (E1), New Caledonian (E2) and Tongan (E3) breeding grounds is small (Garrigue et al. 2020). Some confirmed unpublished sightings exist between the study sites that provide insight in transition times. For example, a humpback whale calf with a lower jaw deformity was sighted in Hervey Bay and again in the Gold Coast bay 9 days later in 2021 (Table S2, Supplementary Fig. S1). The shortest distance between the two bays is ~370 km, equating to a mean minimum constant speed of 1.7 km h−1 between bays for this calf. Similarly, mean minimum migration speeds obtained from fluke photos between Hervey Bay and Ballina, a town ~400 km south of Hervey Bay, suggest minimum constant swimming speeds for calf pods to be from 1.3 to 1.9 km h−1 (Burns et al. 2014). Because Ballina is further south than the Gold Coast bay, any resting in the Gold Coast bay would increase time between sightings and result in a lower estimated migration speed. To investigate the connectivity between Hervey Bay and the Gold Coast bay further, residency times in the Gold Coast bay should be assessed, as well as a comparison of individuals sighted in both Hervey Bay and in the Gold Coast bay.
Conclusions
By analysing over 10 years of sighting data, this study found that whales in Hervey Bay travelled at slower speeds than did whales in the Gold Coast bay and that pods containing a calf consistently travel at slower speeds than do pods without a calf. These results have important implications for techniques aimed at modelling migration or movements of humpback whales between core habitats. Swim speeds of different cohorts of humpback whales may be used in future studies to assess the role of vessel speeds, especially in areas where mothers and calves rest. Furthermore, knowledge of swimming speeds and related transition times between different locations may inform tourism operators about the possible start and length of the whale-watching season and aid the conservation and management of this species in a sustainable way.
Declaration of funding
This work was supported by a grant to Griffith University from a private charitable trust as a part of the Whales and Climate Research Program. Pacific Whale Foundation’s research in Hervey Bay was funded by PacWhale Eco-Adventures Hawaii, members of Pacific Whale Foundation, and a number of private donors.
Acknowledgements
We thank all volunteers who participated in The Oceania Project, and Humpbacks and High-Rises for dedicating their time collecting humpback whale sighting data, and the many team members, both staff and volunteers, that contributed to Pacific Whale Foundation’s long-term humpback whale research. Humpbacks and High-Rises data collection between 2011 and 2020 was supported by Whales in Paradise, Sea World Cruises, Spirit of Gold Coast, Boat Time Yacht Charter and Tallships.
References
Andrews-Goff V, Bestley S, Gales NJ, Laverick SM, Paton D, Polanowski AM, Schmitt NT, Double MC (2018) Humpback whale migrations to Antarctic summer foraging grounds through the southwest Pacific Ocean. Scientific Reports 8(1), 12333.
| Crossref | Google Scholar | PubMed |
Andrews-Goff V, Gales N, Childerhouse SJ, Laverick SM, Polanowski AM, Double MC (2023) Australia’s east coast humpback whales: Satellite tag-derived movements on breeding grounds, feeding grounds and along the northern and southern migration. Biodiversity Data Journal 11, e114729.
| Crossref | Google Scholar |
Bejder L, Videsen S, Hermannsen L, Simon M, Hanf D, Madsen PT (2019) Low energy expenditure and resting behaviour of humpback whale mother–calf pairs highlights conservation importance of sheltered breeding areas. Scientific Reports 9(1), 771.
| Crossref | Google Scholar | PubMed |
Braithwaite JE, Meeuwig JJ, Hipsey MR (2015) Optimal migration energetics of humpback whales and the implications of disturbance. Conservation Physiology 3(1), cov001.
| Crossref | Google Scholar |
Bruce E, Albright L, Sheehan S, Blewitt M (2014) Distribution patterns of migrating humpback whales (Megaptera novaeangliae) in Jervis Bay, Australia: a spatial analysis using geographical citizen science data. Applied Geography 54, 83-95.
| Crossref | Google Scholar |
Burns D, Brooks L, Harrison P, Franklin T, Franklin W, Paton D, Clapham P (2014) Migratory movements of individual humpback whales photographed off the eastern coast of Australia. Marine Mammal Science 30(2), 562-578.
| Crossref | Google Scholar |
Burrows JA, Johnston DW, Straley JM, Chenoweth EM, Ware C, Curtice C, DeRuiter SL, Friedlaender AS (2016) Prey density and depth affect the fine-scale foraging behavior of humpback whales Megaptera novaeangliae in Sitka Sound, Alaska, USA. Marine Ecology Progress Series 561, 245-260.
| Crossref | Google Scholar |
Chittleborough RG (1953) Aerial observations on the humpback whale, Megaptera nodosa (Bonnaterre), with notes on other species. Marine Freshwater Research 4, 219-226.
| Crossref | Google Scholar |
Clapham PJ, Mead JG (1999) Megaptera novaeangliae. Mammalian Species 604, 1-9.
| Google Scholar |
Conrad CC, Hilchey KG (2011) A review of citizen science and community-based environmental monitoring: issues and opportunities. Environmental Monitoring and Assessment 176, 273-291.
| Crossref | Google Scholar | PubMed |
Corkeron PJ, Brown M, Slade RW, Bryden MM (1994) Humpback whales, Megaptera novaeangliae (Cetaceaa : Balaenopteridae), in Hervey Bay, Queensland. Wildlife Research 21(3), 293.
| Crossref | Google Scholar |
Craig AS, Herman LM, Pack AA, Waterman James O (2014) Habitat segregation by female humpback whales in Hawaiian waters: avoidance of males? Behaviour 151(5), 613-631.
| Crossref | Google Scholar |
Currie JJ, McCordic JA, Olson GL, Machernis AF, Stack SH (2021) The impact of vessels on humpback whale behavior: the benefit of added whale watching guidelines. Frontiers in Marine Science 8, 601433.
| Crossref | Google Scholar |
Ejrnæs DD, Sprogis KR (2021) Ontogenetic changes in energy expenditure and resting behaviour of humpback whale mother–calf pairs examined using unmanned aerial vehicles. Wildlife Research 49(1), 34-45.
| Crossref | Google Scholar |
Ersts PJ, Rosenbaum HC (2003) Habitat preference reflects social organization of humpback whales (Megaptera novaeangliae) on a wintering ground. Journal of Zoology 260(4), 337-345.
| Crossref | Google Scholar |
Findlay KP, Best PB, Meÿer MA (2011) Migrations of humpback whales past Cape Vidal, South Africa, and an estimate of the population increase rate (1988–2002). African Journal of Marine Science 33(3), 375-392.
| Crossref | Google Scholar |
Frankel AS, Clark CW, Herman LM, Gabriele CM (1995) Spatial distribution, habitat utilization, and social interactions of humpback whales, Megaptera novaeangliae, off Hawai’i, determined using acoustic and visual techniques. Canadian Journal of Zoology 73(6), 1134-1146.
| Crossref | Google Scholar |
Franklin T, Franklin W, Brooks L, Harrison P (2018) Site-specific female-biased sex ratio of humpback whales (Megaptera novaeangliae) during a stopover early in the southern migration. Canadian Journal of Zoology 96(6), 533-544.
| Crossref | Google Scholar |
Franklin T, Franklin W, Brooks L, Harrison P, Burns D, Holmberg J, Calambokidis J (2020) Photo-identification of individual Southern Hemisphere humpback whales (Megaptera novaeangliae) using all available natural marks: managing the potential for misidentification. Journal Cetacean Research and Management 21(1), 71-83.
| Crossref | Google Scholar |
Franklin T, Franklin W, Brooks L, Harrison P, Pack AA, Clapham PJ (2021) Social behaviour of humpback whales (Megaptera novaeangliae) in Hervey Bay, Eastern Australia, a preferential female stopover during the Southern Migration. Frontiers in Marine Science 8, 652147.
| Crossref | Google Scholar |
Friedlaender AS, Hazen EL, Nowacek DP, Halpin PN, Ware C, Weinrich MT, Hurst T, Wiley D (2009) Diel changes in humpback whale Megaptera novaeangliae feeding behavior in response to sand lance Ammodytes spp. behavior and distribution. Marine Ecology Progress Series 395, 91-100.
| Crossref | Google Scholar |
Garrigue C, Clapham PJ, Geyer Y, Kennedy AS, Zerbini AN (2015) Satellite tracking reveals novel migratory patterns and the importance of seamounts for endangered South Pacific humpback whales. Royal Society Open Science 2(11), 150489.
| Crossref | Google Scholar | PubMed |
Garrigue C, Franklin T, Constantine R, Russell K, Burns D, Poole M, Paton D, Hauser N, Oremus M, Childerhouse S, Mattila D, Gibbs N, Franklin W, Robbins J, Clapham P, Baker CS (2020) First assessment of interchange of humpback whales between Oceania and the east coast of Australia. Journal of Cetacean Research and Management 3, 269-274.
| Crossref | Google Scholar |
Hamilton S, Baker GB (2019) Technical mitigation to reduce marine mammal bycatch and entanglement in commercial fishing gear: lessons learnt and future directions. Reviews in Fish Biology and Fisheries 29, 223-247.
| Crossref | Google Scholar |
Henderson E, Helble TA, Ierley G, Martin S (2018) Identifying behavioral states and habitat use of acoustically tracked humpback whales in Hawaii. Marine Mammal Science 34(3), 701-717.
| Crossref | Google Scholar |
Johnson A, Salvador G, Kenney J, Robbins J, Kraus S, Landry S, Clapham P (2005) Fishing gear involved in entanglements of right and humpback whales. Marine Mammal Science 21(4), 635-645.
| Crossref | Google Scholar |
Kavanagh AS, Noad MJ, Blomberg SP, Goldizen AW, Kniest E, Cato DH, Dunlop RA (2017) Factors driving the variability in diving and movement behavior of migrating humpback whales (Megaptera novaeangliae): implications for anthropogenic disturbance studies. Marine Mammal Science 33(2), 413-439.
| Crossref | Google Scholar |
McCulloch S, Meynecke J-O, Franklin T, Franklin W, Chauvenet ALM (2021) Humpback whale (Megaptera novaeangliae) behaviour determines habitat use in two Australian bays. Marine and Freshwater Research 72(9), 1251-1267.
| Crossref | Google Scholar |
Mayaud R, Castrillon J, Wilson C, Peel D, Smith JN, Luche GD, Allen J, Nash SB (2022) Traffic in a nursery: ship strike risk from commercial vessels to migrating humpback whales (Megaptera novaeangliae) in a rapidly developing Australian urban embayment. Marine Policy 146, 105332.
| Crossref | Google Scholar |
Noad MJ, Cato DH (2007) Swimming speeds of singing and non-singing humpback whales during migration. Marine Mammal Science 23(3), 481-495.
| Crossref | Google Scholar |
Paterson RA (1991) The migration of humpback whales Megaptera novaeangliae in east Australian waters. Memoirs of the Queensland Museum 30, 333-341.
| Google Scholar |
Pirotta V, Slip D, Jonsen ID, Peddemors VM, Cato DH, Ross G, Harcourt R (2016) Migrating humpback whales show no detectable response to whale alarms off Sydney, Australia. Endangered Species Research 29(3), 201-209.
| Crossref | Google Scholar |
Pirotta V, Reynolds W, Ross G, Jonsen I, Grech A, Slip D, Harcourt R (2020) A citizen science approach to long-term monitoring of humpback whales (Megaptera novaeangliae) off Sydney, Australia. Marine Mammal Science 36(2), 472-485.
| Crossref | Google Scholar |
Rasmussen K, Palacios DM, Calambokidis J, Saborio MT, Dalla Rosa L, Secchi ER, Steiger GH, Allen JM, Stone GS (2007) Southern Hemisphere humpback whales wintering off Central America: insights from water temperature into the longest mammalian migration. Biology Letters 3(3), 302-305.
| Crossref | Google Scholar | PubMed |
Salgado Kent C, Jenner K, Jenner M, Bouchet P, Rexstad E (2012) Southern hemisphere breeding stock D humpback whale population estimates from North West Cape, Western Australia. Journal of Cetacean Research and Management 12, 29-38.
| Crossref | Google Scholar |
Schoeman RP, Patterson-Abrolat C, Plön S (2020) A global review of vessel collisions with marine animals. Frontiers in Marine Science 7, 292.
| Crossref | Google Scholar |
Senigaglia V, Christiansen F, Bejder L, Gendron D, Lundquist D, Noren DP, Schaffar A, Smith JC, Williams R, Martinez E, Stockin K, Lusseau D (2016) Meta-analyses of whale-watching impact studies: comparisons of cetacean responses to disturbance. Marine Ecology Progress Series 542, 251-263.
| Crossref | Google Scholar |
Sprogis KR, Videsen S, Madsen PT (2020a) Vessel noise levels drive behavioural responses of humpback whales with implications for whale-watching. Elife 9, e56760.
| Crossref | Google Scholar |
Sprogis KR, Bejder L, Hanf D, Christiansen F (2020b) Behavioural responses of migrating humpback whales to swim-with-whale activities in the Ningaloo Marine Park, Western Australia. Journal of Experimental Marine Biology and Ecology 522, 151254.
| Crossref | Google Scholar |
Stack SH, Currie JJ, McCordic JA, Machernis AF, Olson GL (2020) Distribution patterns of east Australian humpback whales (Megaptera novaeangliae) in Hervey Bay, Queensland: a historical perspective. Australian Mammalogy 42(1), 16-27.
| Crossref | Google Scholar |
Tonachella N, Nastasi A, Kaufman G, Maldini D, Rankin RW (2012) Predicting trends in humpback whale (Megaptera novaeangliae) abundance using citizen science. Pacific Conservation Biology 18(4), 297-309.
| Crossref | Google Scholar |
Tyack P (1983) Differential response of humpback whales, Megaptera novaeangliae, to playback of song or social sounds. Behavioral Ecology and Sociobiology 13(1), 49-55.
| Crossref | Google Scholar |
Valani R, Meynecke J-O, Olsen MT (2020) Presence and movement of humpback whale (Megaptera novaeangliae) mother–calf pairs in the Gold Coast, Australia. Marine and Freshwater Behaviour and Physiology 53, 251-263.
| Crossref | Google Scholar |
Villagra D, García-Cegarra A, Gallardo DI, Pacheco AS (2021) Energetic effects of whale-watching boats on humpback whales on a breeding ground. Frontiers in Marine Science 7, 600508.
| Crossref | Google Scholar |