

Polymicrobial nature of chronic oral disease
Stuart Dashper A , Helen Mitchell A , Geoff Adams A and Eric Reynolds A BA Oral Health Cooperative Research Centre, Melbourne Dental School, The University of Melbourne, Parkville, Vic. 3052, Australia
B Corresponding author. Tel: +61 3 9341 1547, Fax: +61 3 9341 1596, Email: e.reynolds@unimelb.edu.au
Microbiology Australia 36(1) 22-24 https://doi.org/10.1071/MA15007
Published: 6 March 2015
Recent microbiome studies have shown that the human oral microbiome is composed of over 260 abundant bacterial species that predominantly live as polymicrobial biofilms accreted to the non-shedding hard surfaces of the teeth. In addition representatives of both Archaea and Fungi are found in the oral cavity and there is considerable colonisation of the soft tissues of the mouth. Most of these species are commensal and form complex biofilm communities that restrict the colonisation of the oral cavity by exogenous bacteria. Changes in the polymicrobial biofilm microenvironment such as those resulting from the effects of chronic inflammation for subgingival plaque, can lead to the emergence of opportunistic pathogens resulting in dysbiosis and the development of chronic diseases such as periodontitis in a susceptible host. The application of microbiomic studies to the analysis of these complex and dynamic communities in rigorously designed human clinical studies will provide valuable mechanistic insight into the bacterial succession and complex interactions involved in the development of dysbiosis and disease.
The human oral cavity is the entry point of the gastrointestinal tract and offers a number of microenvironments that enable the proliferation of a wide range of largely commensal bacteria, the vast majority of which are endemic to the human oral cavity. Considerable effort has been expended to identify the approximately 700 prokaryote species that compose the total human oral microbiome. Over one-third of these species remain uncultivated and less than half are officially named; however, draft genomes for approximately half of these taxa are now available from the Human Oral Microbial Database (www.homd.org/)1.
16S rRNA gene sequence surveys are providing a cost effective means of studying microbiomes, identifying and enumerating a relatively unbiased set of the prokaryotic species present, including uncultivable species. This technique has been adopted for studying the oral microbiota; however, the results produced have not yet been definitive, with some studies finding huge variation across individuals and limited or no differences between healthy and diseased states. Many factors of the design and analysis of these experiments can make it difficult to compare results between the different studies, including pooling of samples, DNA extraction method, marker gene or region used, primer sets, PCR conditions, sequencing platform, choice of taxonomic classifier and level of classification, clustering of read data into microbial groups, and the statistical methods used for diversity analysis2. The composition of the healthy oral microbiota can certainly vary considerably across sites within the mouth, at the same site over time, and from person to person3,4. Although the 16S rRNA survey techniques are available most clinical studies to date have been cross-sectional and have investigated a limited number of bacterial species using either real time PCR, checkerboard DNA-DNA hybridisation, or more recently, the Human Oral Microbe Identification Microarray (HOMIM) techniques. To compound these limitations samples taken from a limited number of sites within the mouth are often pooled which can obscure comparative results and is not recommended for diverse communities such as the oral microbiome. Generating less information from a larger number of samples is much more informative than generating more information from a small number of samples, particularly for the classification of diseased and healthy states5. Furthermore, clinical sampling techniques may result in only part of the polymicrobial biofilm being collected; with the surface of the biofilm most closely associated with the host and disease process poorly represented in the sample.
Chronic periodontitis is an inflammatory disease of the supporting tissues of the teeth with an endogenous polymicrobial aetiology. Over 47% of Americans over 30 (64.7 million adults) have chronic periodontitis, distributed as 8.7% mild, 30.0% moderate and 8.5% severe6. The prevalence and severity of periodontitis increases with age, with more than 64% of adults aged 65 and over likely to have moderate or severe periodontitis. Numerous cross-sectional and longitudinal epidemiological studies have shown associations between periodontal diseases and a greater risk of certain systemic diseases and disorders, such as cardiovascular diseases, diabetes, chronic kidney disease, metabolic syndrome, obesity, rheumatoid arthritis, Alzheimer’s disease, pre-term and underweight births, and some cancers, particularly pancreatic, head and neck and oesophageal cancers. These associations remain even after adjustment for medical and socio-economic confounding factors.
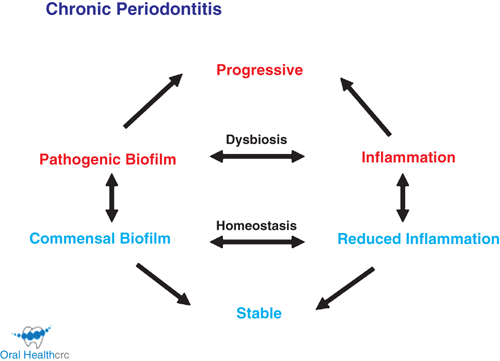
The concepts of the roles of particular oral bacterial species in chronic periodontitis have changed over the past two decades but there is wide consensus that anaerobic, proteolytic, amino acid fermenting species including Porphyromonas gingivalis, Treponema denticola and Tannerella forsythia play a crucial role in initiation and/or progression of disease. A process of bacterial succession in subgingival plaque has been described where mild inflammation of the gingival tissue (gingivitis) and the establishment of an appropriate microenvironment by early colonising bacteria allows late colonisers to emerge as opportunistic pathogens7,8. More recently P. gingivalis has been proposed to be a keystone pathogen that perturbs the ecological balance enabling the proliferation of other oral bacterial species resulting in the formation of a dysbiotic polymicrobial plaque, whilst remaining at very low levels itself9. The keystone pathogen concept was adapted from conservation biology’s keystone species defining a low abundance species which has a disproportionately large effect on its environment. While this theory does help explain the major role of a few species in a complex polymicrobial biofilm, it is not entirely consistent with prospective clinical trial data demonstrating that this bacterium, amongst others, proliferates during disease and can represent a significant proportion of subgingival plaque bacteria at diseased sites. Furthermore the imminent progression of chronic periodontitis at a site has been predicted by increases in the relative proportions of P. gingivalis and/or T. denticola in subgingival plaque at that site above threshold levels of 10–15%10. This and more recent research demonstrating mutualism and synergistic virulence in animal models of periodontitis of multiple late colonising species found closely associated with disease progression in humans has led to the more generally supported view of the emergence of an opportunistic pathogenic polymicrobial biofilm as the trigger for disease progression in susceptible individuals (Figure 1)11. A large number of disease-associated species have been identified, consistent with the dysbiosis-hypothesis12, although further studies are required to differentiate commensal species that benefit from the disease process as opposed to those that actually cause the disease.
|
In addition to the composition of the subgingival plaque polymicrobial biofilm, which will be revealed by microbiomic analyses, its architecture is important as it has been shown that the late colonising opportunistic periodontal pathogens are found as microcolonies in the outer layer of the biofilm adjacent to the epithelium of an inflamed periodontal pocket13. These findings indicate that the disproportionately large impact of the late colonising opportunistic pathogens in disease may be explained by their close proximity to the inflamed tissue and resorbing alveolar bone.
Chronic periodontitis is episodic in nature with acute exacerbations of destruction followed by periods of dormancy. Currently, diagnosis of periodontitis is achieved retrospectively by clinical assessment of attachment loss. This loss of attachment is a result of pathogenic events that have already occurred at the diseased site, and any sampling at the time of diagnosis, may fail to identify those species involved in active destruction. In addition different teeth within the same patient, as well as different sites around the same tooth can display varying degrees of disease severity, all undergoing periodontal disease progression at different rates. Therefore to conclusively determine the polymicrobial aetiology of chronic periodontitis and how opportunistic pathogens emerge and proliferate rigorously designed prospective human clinical trials coupled with microbiomic analyses are essential, followed by the testing of the bacterial species and communities identified in appropriate in vitro and animal models to determine their potential as polymicrobial biofilms to induce dysbiosis and disease.
References
[1] Dewhirst, F.E. et al. (2010) The human oral microbiome. J. Bacteriol. 192, 5002–5017.| The human oral microbiome.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsFCitbrP&md5=c33667927bc198989c9485de67a54bcfCAS | 20656903PubMed |
[2] Goodrich, J.K. et al. (2014) Conducting a microbiome study. Cell 158, 250–262.
| Conducting a microbiome study.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXhtFyjtLfO&md5=d886e997922f9d8200fae60a0a6df47cCAS | 25036628PubMed |
[3] Simón-Soro, Á. et al. (2013) Microbial geography of the oral cavity. J. Dent. Res. 92, 616–621.
| Microbial geography of the oral cavity.Crossref | GoogleScholarGoogle Scholar | 23674263PubMed |
[4] Ge, X. et al. (2013) Oral microbiome of deep and shallow dental pockets in chronic periodontitis. PLoS ONE 8, e65520.
| Oral microbiome of deep and shallow dental pockets in chronic periodontitis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXpvFKhuro%3D&md5=8f649acbe947fb35942fc1fc7326aa46CAS | 23762384PubMed |
[5] Hamady, M. and Knight, R. (2009) Microbial community profiling for human microbiome projects: tools, techniques, and challenges. Genome Res. 19, 1141–1152.
| Microbial community profiling for human microbiome projects: tools, techniques, and challenges.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXosVCktL8%3D&md5=528b70a8af05dde8c9685ef78c153c7eCAS | 19383763PubMed |
[6] Eke, P.I. et al. (2012) Prevalence of periodontitis in adults in the United States: 2009 and 2010. J. Dent. Res. 91, 914–920.
| Prevalence of periodontitis in adults in the United States: 2009 and 2010.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC38bivVCqug%3D%3D&md5=fff11861185f4ecba6758102fb70b77bCAS | 22935673PubMed |
[7] Socransky, S.S. et al. (1998) Microbial complexes in subgingival plaque. J. Clin. Periodontol. 25, 134–144.
| Microbial complexes in subgingival plaque.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK1c7lsFSjsg%3D%3D&md5=5573b48cbf40d873a3455401302a4c29CAS | 9495612PubMed |
[8] Kolenbrander, P.E. et al. (2010) Oral multispecies biofilm development and the key role of cell–cell distance. Nat. Rev. Microbiol. 8, 471–480.
| Oral multispecies biofilm development and the key role of cell–cell distance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmslWmurs%3D&md5=caebe80935d0d9ead616646ff6166c09CAS | 20514044PubMed |
[9] Hajishengallis, G. et al. (2011) Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement. Cell Host Microbe 10, 497–506.
| Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsV2msrzK&md5=c293d1172b0756078b711ad7b4b185d2CAS | 22036469PubMed |
[10] Byrne, S.J. et al. (2009) Progression of chronic periodontitis can be predicted by the levels of Porphyromonas gingivalis and Treponema denticola in subgingival plaque. Oral Microbiol. Immunol. 24, 469–477.
| Progression of chronic periodontitis can be predicted by the levels of Porphyromonas gingivalis and Treponema denticola in subgingival plaque.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1MnpsFShtA%3D%3D&md5=e0be1e620959eadacb395de23d735e89CAS | 19832799PubMed |
[11] Tan, K.H. et al. (2014) Porphyromonas gingivalis and Treponema denticola exhibit metabolic symbioses. PLoS Pathog. 10, e1003955.
| Porphyromonas gingivalis and Treponema denticola exhibit metabolic symbioses.Crossref | GoogleScholarGoogle Scholar | 24603978PubMed |
[12] Curtis, M.A. (2014) Periodontal microbiology—The lid’s off the box again. J. Dent. Res. 93, 840–842.
| Periodontal microbiology—The lid’s off the box again.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC2cbntV2ltA%3D%3D&md5=948211b2e94d25abca931751c9d6374fCAS | 25074493PubMed |
[13] Zijnge, V. et al. (2010) Oral biofilm architecture on natural teeth. PLoS ONE 5, e9321.
| Oral biofilm architecture on natural teeth.Crossref | GoogleScholarGoogle Scholar | 20195365PubMed |
Biographies
Stuart Dashper is a Professor in the Oral Health Cooperative Research Centre and The Melbourne Dental School, The University of Melbourne. Over the past 15 years he has developed a systems biology approach to the study of chronic oral diseases that incorporates the identification and characterisation of bacterial pathobionts, the composition and structures of the polymicrobial biofilm communities in which they dwell, the molecular characterisation of virulence-related traits and their interactions with other members of the bacterial community and the host.
Helen Mitchell is a researcher with the Oral Health CRC and Masters of Science student in Bioinformatics at The University of Melbourne. She is undertaking comparative genomics of the periodontal pathobiont Porphyromonas gingivalis to determine virulence characteristics and assist in vaccine development. She is using Next-Generation Sequencing techniques to determine bacterial biomarkers of early childhood caries in saliva.
Geoff Adams has over 30 years’ experience as a biostatistician and epidemiologist involved in consulting, teaching, and research. He has been employed by the Melbourne Dental School and the Oral Health CRC as a biostatistician and epidemiologist since 1999. Geoff manages the Oral and Systemic Disease program in the Oral Health CRC, which is investigating associations between periodontal disease and various systemic conditions.
Eric Reynolds AO PhD FICD FTSE FRACDS is a Melbourne Laureate Professor and CEO and Director of Research of the Oral Health CRC. He is also Head of the Oral Biology Section of the Melbourne Dental School. He has been researching and teaching for over 30 years on the aetiology and prevention of the two major oral diseases, dental caries and periodontal diseases, which are associated with polymicrobial biofilms.