

Simulating the population dynamics of barley grass (Hordeum spp.) and impacts of weed management strategies in a southern Australian lucerne (Medicago sativa) pasture
Jane E. Kelly A B * , Karl Behrendt B C and Jane C. Quinn A B
A B * , Karl Behrendt B C and Jane C. Quinn A B
A Gulbali Institute for Agriculture Water and the Environment, Charles Sturt University, Locked Bag 588, Wagga Wagga, NSW 2678, Australia.
B Charles Sturt University, Locked Bag 588, Wagga Wagga, NSW 2678, Australia.
C Harper Adams University, Newport, Shropshire TF10 8NB, UK.
Crop & Pasture Science 74(9) 888-897 https://doi.org/10.1071/CP22297
Submitted: 31 August 2022 Accepted: 25 January 2023 Published: 28 February 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Barley grass (Hordeum spp. L.) is an annual, invasive grass weed of southern Australian crops and pastures, frequently associated with weight loss and carcass damage in sheep due to its sharp seeds. Knowledge gaps exist regarding optimal density thresholds for effective control to reduce impacts on animal production. The value of integrated weed management (IWM) over individual control options for reducing barley grass populations in pasture is also unknown.
Aims: We aimed to develop a model for simulating the population dynamics of barley grass within lucerne (Medicago sativa L.) pastures of southern Australia and to test the hypothesis that combining herbicides with mowing will be more effective for removing barley grass seedbanks over time than individual control measures.
Methods: The model was developed within Microsoft Excel and adapted from other annual grass models. The model takes a Monte Carlo approach to simulate control impacts on weed seedbanks over 10 years using five weed-control density thresholds. It was parameterised using data from recent experiments and available literature.
Key results: The most effective long-term control strategy for barley grass occurred with a density threshold of 5 seedlings m−2 by combining early and late herbicide applications, and by combining early and late herbicides with mowing, reducing the seedbank by 86% and 89%, respectively.
Conclusions: Simulation results showed that IWM programs were more effective than individual control options in reducing the barley grass seedbanks over 10 years, particularly at low weed densities (≤50 seedlings m−2).
Implications: Incorporation of this model into a bioeconomic grazing systems model will be valuable for determining the economic impacts and optimal weed-control strategies for minimising the effects of barley grass seed contamination in lamb production systems.
Keywords: annual pasture, defoliation, herbicides, integrated weed mangement, lucerne, mowing, plant growth models, seed production.
Introduction
Barley grass (Hordeum spp. L.) is a southern Australian annual pasture weed and is currently listed as one of the top 20 residual weeds impacting Australian cereal crops (Llewellyn et al. 2016). Barley grass flourishes in nutrient-rich soils, rapidly becoming dominant under limited competition (Rossiter 1964; Groves et al. 2003). Consequently, barley grass typically invades degraded lucerne (Medicago sativa L.) pastures grazed by sheep, where the sharp seeds can penetrate body tissues, resulting in significant production loss and negative welfare implications for the flock (Kelly et al. 2018). Barley grass control in Australia requires significant intervention, typically repeated herbicide applications.
Grazing and mowing have been investigated for barley grassmanagement (Smith 1968a; Myers and Squires 1970; Campbell et al. 1972; Hartley et al. 1978; El-Shatnawi et al. 1999; Bowcher 2002), and differ with regard to their impacts on pasture botanical composition (Jantunen 2003). Effects of grazing are considered more complex than those of mowing (Dormaar et al. 1989), often being associated with grass dominance (Jantunen 2003). By contrast, mowing has been associated with legume dominance (Myers and Squires 1970), frequently reducing annual grass seed production when timed to coincide with reproductive development (El-Shatnawi et al. 1999; Bowcher 2002; Anderson and Frank 2003; Donald 2006; Brownsey et al. 2017). Additional beneficial effects of mowing have been documented considering that repeated mowing may further limit seed production (Donald 2006; Kelly et al. 2020), because barley grass experiences difficulty recovering from defoliation after the commencement of reproductive growth (El-Shatnawi et al. 1999). These effects are also typically exacerbated in dense populations (Kelly et al. 2020), likely resulting from pre-existing competitive stress exerted by neighbouring plants (Weiner 2004).
Despite much research on grass weeds in pasture, information is scarce regarding the long-term efficacy of integrated weed management (IWM) programs against the barley grass seedbank. Recent studies in Australia compared the impacts of single herbicide applications and of mowing with combinations of each on barley grass survival (Kelly et al. 2020), with treatment success shown to be dependent on the timing of applications to plant growth stage and seasonal weather patterns. Other studies investigating herbicide combinations with repeated mowing for control of other annual weeds have also shown some success (Donald et al. 2001). However, the demanding nature of field experiments to validate the effects of IWM strategies can make them costly and impractical. Mathematical modelling can overcome these limitations, facilitating an examination of weed population dynamics and novel weed management strategies over time without the costly consequences of management errors associated with equipment miscalibration or weather events (Pannell et al. 2004). When weed models are authenticated using field trial data, a better understanding of key parameters influencing stages of the weed life cycle is generated, highlighting areas where control strategies may be most effective for managing weed populations (Watkinson et al. 2000).
This paper describes the development of a barley grass population model, which simulates barley grass population dynamics within a typical degraded lucerne pasture in southern New South Wales (NSW), Australia. The paper also identifies the impacts of herbicide/mowing combinations on the weed seedbank. The model was implemented to address the following questions: (1) How does a barley grass seedbank behave in the absence of control over 10 years within an infested southern Australian lucerne pasture? (2) Is the combination of herbicide applications and mowing more effective in reducing the barley grass seedbank than either tactic applied individually over 10 years? (3) Does barley grass population density influence the degree of control exerted on the population by the implementation of weed control strategies over time?
Model description
Existing population models for various weed species have traditionally been based on species with similar life cycles (Gonzalez-Andujar and Fernandez-Quintanilla 2004). Consequently, the barley grass model equations were adapted from the existing models of two other annual grasses, Lolium rigidum Gaud. (Gonzalez-Andujar and Fernandez-Quintanilla 2004) and Avena sterilis L. (Gonzalez-Andujar and Fernandez-Quintanilla 1991). The conceptual model for barley grass growth is described as a simple annual life cycle model from seedling emergence through to seed entry into the seedbank (Fig. 1). Pasture and control efficacy parameters were obtained from experimental observations generated on an experimental site containing a degraded lucerne stand, located at Charles Sturt University, Wagga Wagga, NSW, as described by Kelly et al. (2020).

|
The barley grass model was constructed in Microsoft Excel 2013 (ver. 15.0.5153.1000; Microsoft, Redmond, WA, USA). The model has a 10-year simulation horizon, a time frame considered adequate for visualising annual weed seedbank trends (Gonzalez-Andujar and Fernandez-Quintanilla 1991, 1993, 2004). Additional pasture data were imported into the model from simulations of a Wagga Wagga lucerne–barley grass pasture using the widely accepted decision support tool GRASSGRO (ver. 3.3.9) (Moore et al. 1997).
The six key model equations comprising the barley grass population dynamics model (as adapted from Gonzalez-Andujar and Fernandez-Quintanilla (1991, 2004) are available within the Supplementary Materials (numbered Eqns 1–6). The adaptations that were applied to some of these equations are described as follows:
-
The addition of a climate scalar value. This value was added as a stochastic multiplier within Eqn 4 (Cacho et al. 1999) to adjust seed production based on the effects of seasonal conditions within a given year type. Eqn 4 can thus be re-written as follows:
 where CS is the climate scalar (encompassing values from 0 to 1), which adjusts seed production based on year type. Year type represents the percentage change in seed production occurring as a result of variable weather patterns in any given year. Year types from 1990 to 2017 were used in this model and are randomly selected for each year of the 10-year horizon covered during each simulation iteration.
where CS is the climate scalar (encompassing values from 0 to 1), which adjusts seed production based on year type. Year type represents the percentage change in seed production occurring as a result of variable weather patterns in any given year. Year types from 1990 to 2017 were used in this model and are randomly selected for each year of the 10-year horizon covered during each simulation iteration. -
The addition of control strategies and species competition. Many post-emergent herbicide labels for annual grass weeds recommend application at early tiller stage for optimal control. Certain herbicides, such as paraquat, can be applied somewhat later in the season to reduce seed set and thereby seed production (Powles 1986). Similarly, competition from other species also acts as a mechanism to reduce fecundity (Popay and Sanders 1982). If c1 represents the percentage reduction in seedling survival by early herbicide applications, and c2 and c3 represent the percentage reduction in fecundity as a result of a later herbicide application and competition from another species, respectively (Gonzalez-Andujar and Fernandez-Quintanilla 2004), Eqns 2 and 4 can be rewritten (as Eqns 8, 9, respectively) to include all effects as follows:

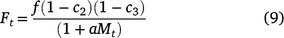 Defoliation by mowing is acknowledged as an effective method for reducing barley grass total fecundity when timed with the most susceptible stage of growth (Bowcher 2002). The proportional reduction in total fecundity due to mowing in the model is given by value c4, rewriting Eqn 5 as:
Defoliation by mowing is acknowledged as an effective method for reducing barley grass total fecundity when timed with the most susceptible stage of growth (Bowcher 2002). The proportional reduction in total fecundity due to mowing in the model is given by value c4, rewriting Eqn 5 as:
GRASSGRO simulations: competition and climate effects
To generate impacts on barley grass survival and seed production due to competition and climate in each simulated year, data for the lucerne–barley grass pasture were produced in GRASSGRO (Moore et al. 1997) using ‘lucerne’ and ‘annual grass – early’ species settings, historical climate data (1990–2017), and default soil settings for Wagga Wagga. This model has some limitations in simulating interspecies competition (Donnelly et al. 2002) and plant death due to stressors over time (Smith et al. 2017) – weaknesses common to all crop and pasture biophysical models (Smith et al. 2017). However, GRASSGRO was considered the most suitable method for modelling species competition within the barley grass model because GRASSGRO is a broadly accepted model, has the capability to simulate basic species competition, and also takes into account the significant and variable impacts of climate/soil moisture on plant growth over time (Clark et al. 2000). GRASSGRO simulations were conducted between 1990 and 2017 to obtain pasture data in response to the climate patterns for each of these years, generating a bank of data representing pasture production under each year ‘type’. Parameters used in the construction of the GRASSGRO pasture simulations are listed in Table 1.

|
For each year, daily total available biomass data (kg DM ha−1) was extracted from GRASSGRO for barley grass and lucerne species. The proportion of lucerne on a dry matter (DM) basis was then calculated per day and averaged across the year to provide the competition input value for c3 in Eqn 6 of the barley grass model, and which varied with year.
Because climate variability influences seed production in barley grass plants (Johnston et al. 2009), the climate scalar (i.e. CS) was introduced as a stochastic element into the barley grass model (Moore et al. 1997; Smith et al. 2017), enabling adjustment of seed production based on year type. Climate index values from 1990 to 2017 were derived by dividing the total barley grass seed production in each of these years by the long-term average seed production value obtained from yearly GRASSGRO simulations (pers. comm., A. Moore, 1 March 2019). Final plant fecundity in each year across the 10-year planning horizon was thus determined by multiplying plant fecundity by the CS index associated with the randomly drawn year (Eqn 5).
All other parameters used in the model are listed in Table 2 and were obtained from greenhouse experimental findings (Kelly et al. 2020) and the recently updated RIM model for barley grass (Monjardino and Llewellyn 2018).

|
Control strategy descriptions
Seven separate weed-control strategies were simulated in the barley grass model, plus a ‘No control’ option, all of which could be used in practice within a grazed lucerne paddock in Australia. Control strategies consisted of an annual early herbicide application, a late herbicide application, and a repeated mow. The remaining control programs consisted of combinations of these approaches. The various control strategies are shown in Table 3.

|
The ‘no control’ strategy provided a comparative baseline seedbank and barley grass population indicative of no suppression of plant growth or seedbank dynamics. ‘Early herbicide’ represents an annual systemic herbicide (e.g. ACCase inhibitor, propaquizafop) applied at 2–3-leaf stage during autumn at 99% efficacy, reflecting recent significant field results obtained during 2016 and 2017 (Kelly et al. 2020). The ‘late herbicide’ strategy represents an annual contact herbicide applied in winter (e.g. Photosystem I electron diverter, paraquat) at 58% efficacy, a value also signifying paraquat efficacy against barley grass obtained from recent field results during 2017 (Kelly et al. 2020). This treatment simulates an annual spraytop application and represents a reactive approach to barley grass control, that is, a herbicide applied later in the season to more mature plants (late vegetative to boot stage) to reduce seed set. The ‘mow’ strategy simulates the reduction in fecundity (removal of inflorescences) as a result of two mowings (i.e. a repeated mow), each of which is timed to coincide with all plants reaching post-inflorescence stage, for the initial defoliation and for the repeated defoliation on regrown plants.
Separate pasture data were generated in GRASSGRO for each strategy within each year to model the impact of control on botanical composition and competition between lucerne and barley grass. No grazing was simulated in the model, because each control strategy in this scenario would typically occur before grazing by spring-weaned lambs in this region. The impact of the herbicides was modelled in GRASSGRO by reducing the barley grass green DM percentage and seed DM parameters in GRASSGRO in each year by the percentage efficacy of each herbicide. This was implemented to simulate early systemic herbicide effects on seedling survival and later contact herbicide effects on seed set. The amount of green DM reduced by the early herbicide was added to the dead DM present at the time of application. Early herbicide parameters were initiated on 1 May in GRASSGRO to simulate control of seedlings. If <100 kg green barley grass DM was present at this time, the simulated application was delayed to the first of each month, thereafter until green DM reached ≥100 kg, or by 1 August, whichever came first. This ensured that the early herbicide application was ‘applied’ to young growing plants, considering that applications may also occur later in some years owing to the interaction of temperature and moisture.
The hay-cut function was utilised in GRASSGRO to simulate the effect of a repeat mowing on the proportion of lucerne in the simulated pasture. The hay-cut function in GRASSGRO was structured to include two defoliations, one on 1 August and the second on 1 September. These months were observed to coincide with inflorescence emergence at the simulated site in Wagga Wagga during 2016 and 2017 (Kelly et al. 2020).
Model parameterisation
Mature plant density and fecundity data obtained from recent greenhouse studies at Wagga Wagga (Kelly et al. 2020) were used to determine the parameters and relationships contained within the barley grass model. Additional data were also collected from greenhouse plants used in this study to determine particular parameters within the model equations. The development and description of these relationships (Supplementary Fig. S1a–d and associated Eqn 11) are available within Supplementary Materials. Parameters are listed in Table 2.
Model sensitivity
A sensitivity analysis incorporating methods by Gonzalez-Andujar and Fernandez-Quintanilla (1993, 2004) was used to assess model sensitivity (Pannell 1997). A sensitivity index was generated by the following relationship:
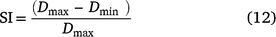
where Dmax and Dmin are the model seedbank output values at the end of the final year of the 10-year simulation when each parameter is set to 40% above (maximum) and below (minimum) the model parameter values (Table 4). The higher the sensitivity index, the more sensitive the model is to parameter variation. The starting seedbank was set at 100 seeds m−2 for all simulations. Variability occurs between model iterations because of climate variability between years; hence, 1000 iterations for each parameter variation were conducted and a mean final seedbank value at the end of the 10-year simulation period was determined. The results of this analysis showed that the barley grass model was most sensitive to changes in emergence, fecundity, seed loss, dormant seed mortality and propaquizafop (early herbicide) efficacy, and relatively insensitive to the value assigned to seedling survivorship, paraquat (late herbicide) and mowing efficacy (Table 4). Previous studies and observations of plants taken during the greenhouse study confirmed that the range of values selected for sensitivity analysis for each parameter was within a realistic set of values obtained across a wide range of conditions (Smith 1968b; Borchert and Jain 1978; Popay 1981; Perry and Ellerton 1983).

|
Model simulations
To examine the impacts of the control methods on the barley grass seedbank over time, a Monte Carlo simulation procedure was used. Different density threshold values (for seedling and mature plant density, per m2) were set to trigger the implementation of control measures based on typical timings of the control strategy in relation to plant growth stage. The selected density threshold value was assumed at seedling stage. The associated mature plant density threshold value was calculated by using Eqn 2 and was considered to be the proportion of seedlings surviving to maturity.
Early and late herbicide ‘applications’ were triggered when barley grass seedling and mature plant densities exceeded set density threshold values for seedlings and mature plants, respectively, during each simulation. The mowing treatment was also triggered in response to mature plant density exceeding a density threshold value, because each mature plant would be expected to produce at least one inflorescence during the reproductive stage, the time when mowing most effectively reduces seed production (Brownsey et al. 2017). In the simulations involving combined strategies, the model timed the implementation of control tactics sequentially as they would occur in practice, triggered whenever the density threshold values required for each strategy were reached. If density threshold values were not exceeded during a simulated year, the particular tactic was not implemented.
A factorial design was chosen, using five weed seedling density thresholds (WT = 5, 50, 250, 500, 1000 seedlings m−2) and eight control strategies, including an uncontrolled seedbank or ‘no control’ treatment (Table 3). The model was run for 1000 iterations per simulation and the starting value for the seedbank was set at 100 seeds m−2 for each iteration. Two-way analysis of variance was conducted using R software (R Core Team 2016) to determine the significance of individual factors (weed density threshold and strategy) in producing mean seedbank differences with significance threshold set at P = 0.05.
Model predictions
Significant differences in the mean seedbank overall occurred as a result of the interaction between strategy and weed density threshold (d.f. = 28, F = 156.9, P < 0.001). Combined strategies were generally found to be most effective in reducing the barley grass seedbank over 10 years (Fig. 2). The largest reductions in the weed seedbank compared with the control occurred when all control tactics were applied at the lowest density thresholds (≤50 seedlings m−2), whereas seedbank size converged towards that of the uncontrolled seedbank as the seedling density threshold increased (≥250 seedlings m−2; Fig. 2).

|
The uncontrolled seedbank reached a population equilibrium at 950 seeds m−2 (Fig. 2), representing the potential maximum density of barley grass invading a lucerne pasture under Wagga Wagga climatic conditions.
Compared with the uncontrolled population, the single early herbicide applications produced the smallest overall mean seedbank of all individual tactics when implemented at a density threshold of 5 seedlings m−2 (288 seeds m−2), reducing the barley grass seedbank by 70% (Fig. 2). By contrast, this tactic was the least effective individual tactic at all other density thresholds, reducing the mean seedbank by16–50% (between 435 and 746 seeds m−2; Fig. 2).
Together with the repeat mowing treatment, the late herbicide application was more effective than the early herbicide at densities >5 seedlings m−2 in reducing the mean seedbank by up to 56% of the uncontrolled population (Fig. 2).
At 5 seedlings m−2, repeat mowing resulted in the largest seedbank of all individual tactics (346 seeds m−2). When implemented at all other density thresholds, this tactic performed similarly to the late herbicide application, and was the most effective individual tactic at the highest density threshold, reducing the weed seedbank by 57% (Fig. 2).
The four combined programs generally resulted in smaller seedbanks than the individual tactics, particularly at the lower weed density thresholds of 5 and 50 seedlings m−2 (Fig. 2). The greatest impact overall was achieved at a density threshold of 5 seedlings m−2 by the early_late and early_late_mow programs, resulting in a 86% and 89% overall reduction in the uncontrolled seedbank, respectively (Fig. 2).
The early_late_mow program was more effective than all other strategies in reducing the seedbank across all density thresholds. Of all other combined strategies, the early_late strategy generally resulted in a lower overall seedbank at density thresholds of 5 and 50 seedlings m−2 only (Fig. 2), whereas the late_mow strategy performed most effectively at higher density thresholds. The early_mow strategy generally performed least effectively of all combinations over time at all density thresholds <1000 seedlings m−2, but surpassed performance of the early_late program within the largest population (Fig. 2).
Discussion
This study demonstrates the value of modelling for comparing the efficacy of integrated and individual control practices against barley grass populations in a lucerne pasture setting over time, and highlights the superiority of IWM for reducing barley grass seedbanks. The size of the barley grass seedbank obtained by model simulations in the absence of control (987 seeds m−2) was consistent with seedbanks measured in lucerne field studies conducted in southern Australia (Powles et al. 1992). Sustained seedbanks of this degree result in dense, mature barley grass stands each year (Powles et al. 1992), leading to prolific seed production, and pose significant contamination risks to grazing sheep (Atkinson and Hartley 1972; Hartley and Bimler 1975). Such risks demonstrate the importance of early control of barley grass populations in grazed pastures before high seedbank levels are attained.
The results of this study consist of model predictions and must therefore be considered as largely speculative; however, they can facilitate the identification of key principles that may inform the practical management of barley grass populations invading established lucerne pastures in southern Australia.
Although individual control strategies were predicted to reduce the seedbank compared with the uncontrolled population, a residual barley grass population remained, contributing seed to the seedbank in subsequent years. This is to be expected in practice, because the efficacy of singular control practices is rarely 100%. Further, when chemical control is suboptimal, such as encountered by the late herbicide treatment in this study (i.e. 58%), the practices typically lead to the formation of larger weed populations, greater investment in herbicides, and a higher risk of ensuing herbicide resistance (Diggle et al. 2003; Gonzalez-Andujar and Fernandez-Quintanilla 2004).
The repeat mowing strategy simulated in this study shows promise as an individual tool for reducing community-level seed production in barley grass populations, a predicted outcome supported by field studies investigating the impacts of defoliation on other annual grasses such as Avena spp. and Lolium perenne populations (Morris and Gardner 1958; Hebblethwaite and Clemence 1983). However, the timely application of mowing post inflorescence emergence will likely be crucial to success when implemented in practice, because defoliations of annual grasses prior to reproductive development under field study conditions have typically resulted in sustained or increased seed production (Hebblethwaite and Clemence 1983; El-Shatnawi et al. 1999; Brownsey et al. 2017; Kelly et al. 2020). Fewer viable seeds are typically available for dispersal and seedbank entry at this stage of growth, owing to incomplete seed development (Hempy-Mayer and Pyke 2008). Further, the repeated nature of the mowing strategy simulated in the model may also be important during field applications, considering that the greatest reductions in seed production were reported when repeated defoliations of barley grass plants were timed to occur at the same stage of growth (Kelly et al. 2020).
The superiority of the combined strategies in reducing the simulated barley grass seedbank was consistent with field results reported by Piltz et al. (2019), who reported superior control against barley grass in a clover pasture by combining grazing with herbicides. However, the model predictions were inconsistent with the results of defoliation and herbicide treatments applied under field conditions similar to the simulations (Kelly et al. 2020). This may be due to differences in the timing of treatments between the field study and simulations, where individual field mowing applications that were applied while some plants were still at the vegetative stage encouraged increased tillering and copious seed production.
Despite the predicted efficacy of the combined weed-control programs simulated in this model, some considerations and questions remain in relation to their use in practice. These include the potential for emerging herbicide resistance within weed populations as a result of repeated herbicide use and the economic advantages of such strategies for maximising production returns from livestock grazing barley grass-infested lucerne pastures. Hypothetically, herbicides applied prior to a repeated mowing post inflorescence emergence may in fact limit development of herbicide resistance because surviving plants cannot contribute seed to the seedbank owing to inflorescence removal by mowing. This may be likened to an effect similar to that of the ‘double knock’ system practiced in cropping systems. Further, considering the prevailing high lamb prices across southern Australia in recent years (Behrendt and Weeks 2019), it is possible that the benefits of weed seed control in pastures outweigh the input price of grass herbicide as well as the labour and machinery costs associated with mowing. If so, the early_late_mow program would show promise in yielding high economic benefits to a lamb enterprise grazing lucerne pastures with significant barley grass incursion, given that this strategy maintained the smallest weed population consistently over time at all weed density thresholds and would thus be expected to reduce the frequency of contact between seeds and grazing animals most significantly. However, in practice, consideration would need to be given to the phenological development of other species within the pasture, because applications timed to control barley grass may also inadvertently produce detrimental impacts on the co-existent lucerne population, resulting in a negative production outcome.
The outcomes from this study also highlight the important influence of weed seedling density on managed barley grass populations, owing to the direct relationship between seed density and the size of the residual population in the absence of any control intervention. Implementing effective population control at low density thresholds limited the size of the population and reduced the community-level seed production to the seedbank. This likely explains the greater efficacy of the early herbicide application than of the other individual tactics implemented at the lowest density threshold. Although control was not attained when implemented at high density thresholds owing to significant community-level seed production, individual late herbicide and mowing applications can effect greater control than an early herbicide because of their impact on seed production and the number of seeds entering the seedbank in the same year. Although more costly, better results may be achieved in practice via the optimal and timely application of herbicides and mowings, combined with re-establishment of the lucerne pasture in the following year. Model findings thus advocate the proactive uptake of cost-effective weed-control practices that preserve low barley grass seedbanks throughout the life of the pasture, facilitating greater control, less herbicide use over time, and the potential for competition by neighbouring species (Kemp and King 2001). Furthermore, such practices may limit the frequency of herbicide-resistance alleles in any emerging weed populations (Christoffers 1999).
The sensitivity analysis results reported in this study suggest the vulnerability of barley grass populations to changes in emergence, fecundity, seed loss and dormant seed mortality. Fecundity and seed loss results were consistent with the findings of Gonzalez-Andujar and Fernandez-Quintanilla (2004) for the similarly constructed annual ryegrass model, giving confidence to this model. Contrasting results concerning emergence and dormant seed mortality may be explained by the incorporation of climatic variability in the current model, which likely varied the response of these parameters from year to year. Unsurprisingly, seedling density (as influenced by emergence), and fecundity, were also two major population drivers that were influenced by each tactic within combined programs in this study, leading to greatest population control over time. Studying these areas in detail under field conditions at various locations will facilitate the validation of the model under different conditions. This will be particularly important in relation to the emergence parameter, which had been determined under controlled conditions. Such values could be expected to be lower under field conditions, where emergence can be regulated by the influence of early density dependence (Lortie and Turkington 2002) and the release of leachates by neighbouring seeds (Murray 1998). Lower emergence under field conditions may also result in greater plant size (and higher seed production) in later life stages, an effect observed in other annual species under field conditions (Lortie and Turkington 2002).
The predictions reported by the barley grass model suggest a variety of strategic approaches that could be used in practice for effective control of this weed in lucerne pastures. Control tactics that target emergence, fecundity, seed loss and seed dormancy within the barley grass life cycle in any given year (i.e. via species competition, controlling soil surface cover, prevention of seed production or inflorescence removal) were predicted to be most effective for seedbank control. Under field conditions, the correct timing of herbicide applications in low-density populations are likely to be particularly important for limiting barley grass seed production later in the season, and repeat mowing tactics applied during reproductive development may be useful when applied to high-density populations, reducing herbicide costs and constraining population size.
The model highlights the value of combined weed management strategies for reducing barley grass seedbanks in lucerne pastures over time. In dense stands, alternative measures in the first year may be required to reduce the residual population to more manageable levels. Assessment of control costs and the value of livestock products grazing such pastures under fluctuating market prices will also be necessary in determining optimal control measures in livestock grazing systems, models that may now be developed based on this founding model system.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study are available in the article and accompanying online supplementary material.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
The development of the barley grass model was supported by Meat & Livestock Australia (Grant number B.STU.1807) and a fellowship awarded to Jane Kelly by the AW Howard Memorial Trust.
Acknowledgements
The authors gratefully acknowledge the support of Prof. Leslie Weston, Dr John Broster and Dr Paul Weston for their oversight and support during model development, the assistance of Allison Chambers in the collection of data during greenhouse studies, and Simon McDonald for assistance with model parameterisation.
References
Anderson MT Anderson MT (2003) Defoliation effects on reproductive biomass: importance of scale and timing. Journal of Range Management 56, 501| Defoliation effects on reproductive biomass: importance of scale and timing.Crossref | GoogleScholarGoogle Scholar |
Atkinson G, Hartley M (1972) Damage caused to lambs by barley grass. In ‘Proceedings of the NZ weed and pest control conference’. 15–17 August, 1972. Hastings, New Zealand. Vol. 25. (Ed. M Hartley) pp. 29–33. (New Zealand Weed Plant Protection Society: Auckland, New Zealand)
Behrendt K, Weeks P (2017) How are global and Australian beef and sheepmeat producers performing? Global agri benchmark network results 2016. MLA market information report. Meat & Livestock Australia, North Sydney, NSW, Australia.
Borchert MI Borchert MI (1978) The effect of rodent seed predation on four species of California annual grasses. Oecologia 33, 101–113.
| The effect of rodent seed predation on four species of California annual grasses.Crossref | GoogleScholarGoogle Scholar |
Bowcher AJ (2002) Competition between temperate perennial pasture species and annual weeds: the effect of pasture management on population dynamics and resource use, PhD Thesis, Charles Sturt University, Wagga Wagga, NSW, Australia.
Brownsey P Brownsey P Brownsey P (2017) Using phenology to optimize timing of mowing and grazing treatments for medusahead (Taeniatherum caput-medusae). Rangeland Ecology & Management 70, 210–218.
| Using phenology to optimize timing of mowing and grazing treatments for medusahead (Taeniatherum caput-medusae).Crossref | GoogleScholarGoogle Scholar |
Cacho OJ Cacho OJ Cacho OJ (1999) Assessment of production risk in grazing models. Agricultural Systems 60, 87–98.
| Assessment of production risk in grazing models.Crossref | GoogleScholarGoogle Scholar |
Campbell R Campbell R Campbell R (1972) The effect of grass seed on sheep production. Proceedings of the Australian Society of Animal Production 9, 225–229.
Christoffers MJ (1999) Genetic aspects of herbicide-resistant weed management. Weed Technology 13, 647–652.
| Genetic aspects of herbicide-resistant weed management.Crossref | GoogleScholarGoogle Scholar |
Clark SG Clark SG Clark SG (2000) The GrassGro decision support tool: its effectiveness in simulating pasture and animal production and value in determining research priorities. Australian Journal of Experimental Agriculture 40, 247–256.
| The GrassGro decision support tool: its effectiveness in simulating pasture and animal production and value in determining research priorities.Crossref | GoogleScholarGoogle Scholar |
Crawford MC Crawford MC Crawford MC Crawford MC (1997) Root production of a barrel medic (Medicago truncatula) pasture, a barley grass (Hordeum leporinum) pasture, and a faba bean (Vicia faba) crop in southern Australia. Australian Journal of Agricultural Research 48, 1139–1150.
| Root production of a barrel medic (Medicago truncatula) pasture, a barley grass (Hordeum leporinum) pasture, and a faba bean (Vicia faba) crop in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Diggle AJ Diggle AJ Diggle AJ (2003) Herbicides used in combination can reduce the probability of herbicide resistance in finite weed populations. Weed Research 43, 371–382.
| Herbicides used in combination can reduce the probability of herbicide resistance in finite weed populations.Crossref | GoogleScholarGoogle Scholar |
Donald WW (2006) Mowing for weed management. In ‘Handbook of sustainable weed management’. (Eds HP Singh, DR Batish, RK Kohli) pp. 329–372. (Food Products Press: New York, NY, USA)
Donald WW Donald WW (2001) Between-row mowing + banded herbicide to control annual weeds and reduce herbicide use in no-till soybean (glycine max) and corn (Zea mays). Weed Technology 15, 576–584.
| Between-row mowing + banded herbicide to control annual weeds and reduce herbicide use in no-till soybean (glycine max) and corn (Zea mays).Crossref | GoogleScholarGoogle Scholar |
Donnelly JR, Freer M, Salmon L, Moore AD, Simpson RJ, Dove H, Bolger TP (2002) Evolution of the GRAZPLAN decision support tools and adoption by the grazing industry in temperate Australia. Agricultural Systems 74, 115–139.
| Evolution of the GRAZPLAN decision support tools and adoption by the grazing industry in temperate Australia.Crossref | GoogleScholarGoogle Scholar |
Dormaar JF Dormaar JF Dormaar JF (1989) Vegetation and soil responses to short-duration grazing on fescue grasslands. Journal of Range Management 42, 252–256.
| Vegetation and soil responses to short-duration grazing on fescue grasslands.Crossref | GoogleScholarGoogle Scholar |
El-Shatnawi MKJ El-Shatnawi MKJ El-Shatnawi MKJ El-Shatnawi MKJ (1999) Defoliation time and intensity of wall barley in the Mediterranean rangeland. Journal of Range Management 52, 258–262.
| Defoliation time and intensity of wall barley in the Mediterranean rangeland.Crossref | GoogleScholarGoogle Scholar |
Gonzalez-Andujar JL Gonzalez-Andujar JL (1991) Modelling the population dynamics of Avena sterilis under dry-land cereal cropping systems. Journal of Applied Ecology 28, 16–27.
| Modelling the population dynamics of Avena sterilis under dry-land cereal cropping systems.Crossref | GoogleScholarGoogle Scholar |
Gonzalez-Andujar JL Gonzalez-Andujar JL (1993) Strategies for the control of Avena sterilis in winter wheat production systems in central Spain. Crop Protection 12, 617–623.
| Strategies for the control of Avena sterilis in winter wheat production systems in central Spain.Crossref | GoogleScholarGoogle Scholar |
Gonzalez-Andujar JL Gonzalez-Andujar JL (2004) Modelling the population dynamics of annual ryegrass (Lolium rigidum) under various weed management systems. Crop Protection 23, 723–729.
| Modelling the population dynamics of annual ryegrass (Lolium rigidum) under various weed management systems.Crossref | GoogleScholarGoogle Scholar |
Groves R Groves R Groves R (2003) Competition between Australian native and introduced grasses along a nutrient gradient. Austral Ecology 28, 491–498.
| Competition between Australian native and introduced grasses along a nutrient gradient.Crossref | GoogleScholarGoogle Scholar |
Hartley M, Bimler K (1975) Barley grass damage to lambs. In ‘Proceedings of the 28th New Zealand weed and pest control conference, 5–7 August, Hastings, New Zealand’. (Ed. M Hartley) pp. 2–6. (The New Zealand Weed and Pest Control Conference Inc.: New Zealand)
Hartley M, Atkinson G, Bimler K, James T, Popay A (1978) Control of barley grass by grazing management. In ‘Proceedings of the NZ weed and pest control conference’. 8–10 August, 1978. New Plymouth, New Zealand. Vol. 31. (Ed. MJ Hartley) pp. 198–202. (New Zealand Plant Protection Society: Auckland, New Zealand)
Hebblethwaite P, Clemence T (1983) Effect of autumn and spring defoliation and defoliation method on seed yield of Lolium perenne. In ‘Proceedings of the 14th international grasslands congress’. 15–24 June 1983, Lexington, KY, USA. (Eds J Smith, V Hayes) pp. 257–260. (Westview Press: Boulder, CO, USA)
Hempy-Mayer K Hempy-Mayer K (2008) Defoliation effects on Bromus Tectorum seed production: implications for grazing. Rangeland Ecology & Management 61, 116–123.
| Defoliation effects on Bromus Tectorum seed production: implications for grazing.Crossref | GoogleScholarGoogle Scholar |
Jantunen J (2003) Vegetation changes in a semi-natural grassland during mowing and grazing periods. Annales Botanici Fennici 40, 255–263.
Johnston MB Johnston MB Johnston MB (2009) Effect of quantity and distribution of rainfalls on Hordeum murinum L. growth and development. Chilean Journal of Agricultural Research 69, 188–197.
| Effect of quantity and distribution of rainfalls on Hordeum murinum L. growth and development.Crossref | GoogleScholarGoogle Scholar |
Kelly JE Kelly JE Kelly JE Kelly JE Kelly JE Kelly JE (2018) Seed contamination in sheep: new investigations into an old problem. Animal Production Science 58, 1538–1544.
| Seed contamination in sheep: new investigations into an old problem.Crossref | GoogleScholarGoogle Scholar |
Kelly JE Kelly JE Kelly JE (2020) The impact of herbicide application and defoliation on barley grass (Hordeum murinum subsp. glaucum) management in mixed pasture legumes. Agronomy 10, 671
| The impact of herbicide application and defoliation on barley grass (Hordeum murinum subsp. glaucum) management in mixed pasture legumes.Crossref | GoogleScholarGoogle Scholar |
Kemp DR, King WM (2001) Plant competition in pastures–implications for management. In ‘Competition and succession in pastures’. (Eds PG Tow, A Lazenby) pp. 85–102. (CAB International: Wallingford, UK)
Llewellyn R, Ronning D, Ouzman J, Walker S, Mayfield A, Clarke M (2016) Impact of weeds on Australian grain production – the cost of weeds to Australian grain growers and the adoption of weed management and tillage practices. March 2016. Report for Grains Research & Development Corporation, CSIRO, Canberra, ACT, Australia.
Lortie CJ, Turkington R (2002) The effect of initial seed density on the structure of a desert annual plant community. Journal of Ecology 90, 435–445.
| The effect of initial seed density on the structure of a desert annual plant community.Crossref | GoogleScholarGoogle Scholar |
Monjardino M, Llewellyn R (2018) Barley grass RIM. Available at https://ahri.uwa.edu.au/research/barley-grass-rim/ [Accessed 18 January 2019]
Moore AD Moore AD Moore AD (1997) GRAZPLAN: decision support systems for Australian grazing enterprises. III. Pasture growth and soil moisture submodels, and the GrassGro DSS. Agricultural Systems 55, 535–582.
| GRAZPLAN: decision support systems for Australian grazing enterprises. III. Pasture growth and soil moisture submodels, and the GrassGro DSS.Crossref | GoogleScholarGoogle Scholar |
Morris HD Morris HD (1958) The effect of nitrogen fertilization and duration of clipping period on forage and grain yields of oats, wheat, and rye. Agronomy Journal 50, 454–457.
| The effect of nitrogen fertilization and duration of clipping period on forage and grain yields of oats, wheat, and rye.Crossref | GoogleScholarGoogle Scholar |
Murray BR (1998) Density-dependent germination and the role of seed leachate. Australian Journal of Ecology 23, 411–418.
| Density-dependent germination and the role of seed leachate.Crossref | GoogleScholarGoogle Scholar |
Myers LF Myers LF (1970) Control of barley grass (Hordeum leporinum) by grazing management in irrigated pastures. Australian Journal of Experimental Agriculture 10, 151–155.
| Control of barley grass (Hordeum leporinum) by grazing management in irrigated pastures.Crossref | GoogleScholarGoogle Scholar |
Pannell DJ (1997) Sensitivity analysis of normative economic models: theoretical framework and practical strategies. Agricultural Economics 16, 139–152.
| Sensitivity analysis of normative economic models: theoretical framework and practical strategies.Crossref | GoogleScholarGoogle Scholar |
Pannell DJ Pannell DJ Pannell DJ Pannell DJ Pannell DJ Pannell DJ (2004) RIM: a bioeconomic model for integrated weed management of Lolium rigidum in Western Australia. Agricultural Systems 79, 305–325.
| RIM: a bioeconomic model for integrated weed management of Lolium rigidum in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Perry DA Perry DA (1983) Studies on death of deteriorated barley seeds in soil. Annals of Applied Biology 102, 183–191.
| Studies on death of deteriorated barley seeds in soil.Crossref | GoogleScholarGoogle Scholar |
Piltz JW, Flinn SJ, Weston LA (2019) Comparative effects of grazing, herbicide or forage conservation on barley grass content in Trifolium subterraneum L. clover-based pasture. Crop & Pasture Science 70, 800–806.
| Comparative effects of grazing, herbicide or forage conservation on barley grass content in Trifolium subterraneum L. clover-based pasture.Crossref | GoogleScholarGoogle Scholar |
Popay AI (1981) Germination of seeds of five annual species of barley grass. Journal of Applied Ecology 18, 547–558.
| Germination of seeds of five annual species of barley grass.Crossref | GoogleScholarGoogle Scholar |
Popay A Popay A (1982) Effect of pasture competition on barley grass seed production. New Zealand Journal of Agricultural Research 25, 143–146.
| Effect of pasture competition on barley grass seed production.Crossref | GoogleScholarGoogle Scholar |
Powles SB (1986) Appearance of a biotype of the weed, Hordeum glaucum Steud., resistant to the herbicide paraquat. Weed Research 26, 167–172.
| Appearance of a biotype of the weed, Hordeum glaucum Steud., resistant to the herbicide paraquat.Crossref | GoogleScholarGoogle Scholar |
Powles SB Powles SB Powles SB (1992) Eradication of paraquat-resistant Hordeum glaucum Steud. by prevention of seed production for 3 years. Weed Research 32, 207–211.
| Eradication of paraquat-resistant Hordeum glaucum Steud. by prevention of seed production for 3 years.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2016) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria)
Robertson S (2012) EverGraze: tactical management regimes report on simulation modelling at Wagga Wagga. Future Farm Industries Cooperative Research Centre, Perth, WA, Australia.
Rossiter RC (1964) The effect of phosphate supply on the growth and botanical composition of annual type pasture. Australian Journal of Agricultural Research 15, 61–76.
| The effect of phosphate supply on the growth and botanical composition of annual type pasture.Crossref | GoogleScholarGoogle Scholar |
Smith D 1968a) The growth of barley grass (Hordeum leporinum) in annual pasture. 4. The effect of some management practices on barley grass content. Australian Journal of Experimental Agriculture 8, 706–711.
| 1968a) The growth of barley grass (Hordeum leporinum) in annual pasture. 4. The effect of some management practices on barley grass content.Crossref | GoogleScholarGoogle Scholar |
Smith D 1968b) The growth of barley grass (Hordeum leporinum) in annual pasture. 1. Germination and establishment in comparison with other annual pasture species. Australian Journal of Experimental Agriculture 8, 478–483.
| 1968b) The growth of barley grass (Hordeum leporinum) in annual pasture. 1. Germination and establishment in comparison with other annual pasture species.Crossref | GoogleScholarGoogle Scholar |
Smith AP Smith AP Smith AP Smith AP Smith AP Smith AP (2017) Modelling of lucerne (Medicago sativa L.) for livestock production in diverse environments. Crop & Pasture Science 68, 74–91.
| Modelling of lucerne (Medicago sativa L.) for livestock production in diverse environments.Crossref | GoogleScholarGoogle Scholar |
Watkinson A, Freckleton RP, Dowling PM (2000) Weed invasions of Australian farming systems: from ecology to economics. In ‘The economics of biological invasions’. (Eds C Perrings, M Williamson, S Dalmazzone) pp. 94–116. (Edward Elgar Publishing: Melbourne, Vic., Australia)
Weiner J (2004) Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics 6, 207–215.
| Allocation, plasticity and allometry in plants.Crossref | GoogleScholarGoogle Scholar |