

Relative performance of white clover (Trifolium repens) cultivars and experimental synthetics under rotational grazing by beef cattle, dairy cattle and sheep
M. Z. Z. Jahufer A * , J. L. Ford B , G. R. Cousins B and D. R. Woodfield B
A * , J. L. Ford B , G. R. Cousins B and D. R. Woodfield B
A AgResearch Ltd, Grasslands Research Centre, Private Bag 11008, Palmerston North, New Zealand.
B PGG Wrightson Seeds Limited, C/- Grasslands Research Centre, Building 88, Private Bag 11008, Palmerston North, New Zealand.
Crop & Pasture Science 72(11) 926-938 https://doi.org/10.1071/CP21084
Submitted: 10 February 2021 Accepted: 18 July 2021 Published: 26 November 2021
© 2021 The Author(s) (or their employer(s)). Published by CSIRO Publishing Open Access CC BY-NC
Abstract
Assessment of the relative performance of white clover (Trifolium repens L.) cultivars, using multi-year and multi-location seasonal growth trials, is key to identification of material with specific and broad adaptation. This paper is based on a multi-year and multi-location study of 56 white clover entries comprising 14 commercial cultivars and 42 experimental synthetic lines evaluated for seasonal growth under rotational grazing across four locations in New Zealand over 4 years. The four locations (and animals grazing) were: Kerikeri (beef cattle), Aorangi (beef cattle), Ruakura (dairy cattle), Lincoln (sheep). Significant (P < 0.05) genotypic variation among the 56 entries, and genotype × year, genotype × location and genotype × season interactions, were estimated. We were able to identify cultivars and experimental synthetics with specific and broad adaptation to the three grazing management types. Cvv. AberDance, Apex, Demand, Prestige, Quartz and Riesling, with leaf size ranging from small to medium–large, showed highly above-average performance under sheep grazing. Synthetic lines 15 and 45 also had highly above-average performance under sheep grazing. Cvv. Legacy and Kopu II showed above-average performance under cattle and dairy grazing. Synthetics 15, 48, 49, 44, 22 and 18 and cv. Quartz had above-average performance under all three grazing managements. Synthetics 27, 33 and 38 had highly above-average performance across all three grazing managements and were superior to all 14 cultivars evaluated. Several of these superior synthetics are being tested across multiple grazing environments. Among the 14 cultivars evaluated, Legacy and Quartz showed superior seasonal growth performance across the three grazing managements. Quartz is being evaluated in several on-farm trials across temperate regions of the world.
Keywords: broad adaptation, forage breeding, genotype-by-environment interaction, grazing management, leaf size, multi-location testing, pattern analysis, vegetative persistence.
Introduction
Pastures based on white clover (Trifolium repens L.) and perennial ryegrass (Lolium perenne L.) form the main feed base for the milk, meat and wool production industries in Australasia. Superior feeding value, high acceptability by stock and fixation of atmospheric nitrogen make white clover a significant perennial forage legume in temperate New Zealand and Australian farming systems. Reviews by Ulyatt (1973) and Thomson (1984) highlight the importance of white clover for improved animal production. The annual contribution of white clover to New Zealand’s economy is more than NZ$3 billion, considering its value to the grazing and seed industries, nitrogen fixation and honey production (Caradus et al. 1996b). In a survey of gross value of production from wool, sheep and beef from pastures with a significant proportion of white clover in the dryland temperate zone of Australia, Jahufer et al. (1996) estimated values for the individual states of AU$202 m (New South Wales), $256 m (Victoria), $18 m (Tasmania), $0.6 m (South Australia) and $6 m (Western Australia).
For white clover to enhance pasture production, stable seasonal herbage yield and plant persistence are two important criteria (Gramshaw et al. 1989). However, white clover performance in Australasia is constrained by a range of abiotic and biotic factors. Factors such as summer moisture stress, plant parasitic nematodes and clover root weevil (Sitona lepidus) (Barbour et al. 1996; Eerens et al. 2001; Knowles et al. 2003; Mercer et al. 2008) limit the genetic potential of white clover. Summer moisture stress is considered a major constraint on vegetative persistence and herbage production in temperate dryland environments of Australia (FitzGerald and Clark 1993; Hutchinson 1993). In New Zealand, moisture stress during summer and autumn can deplete the white clover content in pastures (Brock et al. 2003; Knowles et al. 2003). Intensification of New Zealand farming systems is changing pastoral environments, resulting in less favourable conditions for the traditional clover/ryegrass pastures (Clark et al. 2001). Another significant challenge to these pastures is the increasing climatic variability associated with global warming (Newton and Edwards 2007).
Several commercial white clover cultivars are available to farmers across the cattle, dairy and sheep grazing industries (Caradus et al. 1996a; Woodfield 1999). However, there is a need for continuous genetic improvement of white clover not only to overcome environmental constraints but also to improve production by grazing animals. New livestock breeds with greater genetic potential and nutritional demands require even more productive and reliable perennial pastures (Kemp et al. 2010).
Multi-location evaluation of elite breeding lines, together with commercial cultivars, across key target environments is a vital phase in cultivar development programs. This phase enables the identification of superior breeding material with specific or broad adaptation relative to commercial check cultivars, and provides valuable information on the relative performance of these cultivars across environments. We report a study of 56 white clover entries consisting of 14 commercial cultivars and 42 experimental synthetics, evaluated for seasonal growth across four locations in New Zealand for a period of 4 years. We examine relative performance of the cultivars and experimental synthetics at Kerikeri, Ruakura, Aorangi and Lincoln, under beef cattle, dairy cattle and sheep rotational grazing.
Materials and methods
Entries
In total, 56 white clover entries including 14 commercial cultivars and 42 experimental synthetic lines were evaluated. Here, an experimental synthetic is an F2 developed from an isolated polycross of elite parental genotypes. The commercial cultivars and their leaf sizes (VS, very small; S, small; SM, small–medium; M, medium; ML, medium–large; L, large) were: AberAce (VS), AberDance (M), Bounty (M), Apex (M), Demand (SM), Huia (M), Klondike (L), Kopu II (L), Legacy (L), Prestige (S), Quartz (ML), Riesling (ML), Sustain (ML) and Tribute (ML). Breeding methods and pedigree descriptions of the experimental synthetics are not provided for reasons of intellectual property.
Trial sites
Field trials were established in 2014 at four locations: AgResearch, Lincoln Research Farm in Canterbury; AgResearch, Aorangi Experimental Farm; Ruakura Research Centre; and Plant and Food Research Station, Kerikeri, Northland. Key location characteristics are presented in Table 1.

|
Field trials
All four locations were planted according to row–column experimental designs (John 1987) with three replicates. The trials at Kerikeri, Ruakura and Aorangi were sown in autumn 2014. At all three locations, seed of perennial ryegrass cv. Ceres150 (AR37) was direct drilled into the experimental area at the rate of 20 kg ha−1. This was followed immediately by hand sowing seed of the white clover entries at 5 kg ha−1. Each of the three locations had plots of size 1 m by 2 m. At Lincoln, the trial was planted in early spring 2014. Ceres150 (AR37) perennial ryegrass was direct drilled together with the white clover entries at the same sowing rates as used at the other three locations. The experimental plots at Lincoln were 1.5 m by 4 m in size. Post-planting hand weeding and herbicide spot spraying were used to control broadleaf weeds and white clover from buried seed. Superphosphate fertiliser (150 kg ha−1) was applied across the trials in late autumn in Years 1 and 3.
Grazing and trait measurements
Rotational grazing was implemented at all four locations. At Kerikeri and Aorangi, the trials were grazed by Angus cattle when the estimated sward dry matter (DM) yield reached 2200–2500 kg ha−1. Grazing was conducted down to sward residuals of ∼1200 DM kg ha−1. Low annual nitrogen (N) inputs of 20–50 kg ha−1 were applied. At Ruakura, the trial was grazed by Holstein–Friesian dairy cattle when the estimated sward DM yield was ∼2500–3000 kg DM ha−1. Post-grazing residuals were ∼1500 kg DM ha−1. Under dairy grazing, high annual inputs of N were applied: 50 kg ha−1 five times per year, twice in spring (early and late), once in summer (mid to late), once in mid-autumn, and once in winter (mid to late). At Lincoln, grazing was done by Romney ewes. Under sheep grazing, low annual inputs of N were applied: 15–20 kg ha−1 two or three times per year, twice in spring (early and late) and once in mid-autumn. At each location, an F400 FARMWORKS Electronic Rising Plate Meter (www.farmworkspfs.co.nz) was used to estimate the quantity of herbage and provide a rough guide on when to graze. The plots were grazed when the estimated sward DM yield was at ∼2000–2200 kg ha−1 down to a grazing residual of 1100 kg DM ha−1. At each location, the trial was grazed for 2–3 h, season dependent, to ensure rapid and uniform defoliation. After grazing, the animals were immediately removed from the trial area.
Seasonal growth was scored at each location prior to grazing. The score scale ranged from 1 (poor) to 9 (high) (Ford et al. 2015). Leaf size was also measured before grazing. Leaf size was scored using a scale of 1 (very small) to 5 (very large) (Ford et al. 2015). Leaf size score data from Ruakura were not included in the analysis owing to error.
Data analyses
Variance component analyses
Seasonal growth score data were analysed: (i) on an individual location basis across 4 years, and also across all years, seasons and locations; (ii) within each season across all 4 years and locations; (iii) within each grazing type (beef cattle, dairy cattle, sheep) and combinations of grazing types (beef cattle/sheep, beef cattle/dairy cattle), across all 4 years; and (iv) within Years 2–4 within each individual grazing management type. Leaf size score data were analysed across all seasons, 4 years and three locations (Kerikeri, Aorangi and Lincoln). Variance component analyses were conducted by using the residual maximum likelihood (REML) procedure (Patterson and Thompson 1971, 1975; Harville 1977) in DeltaGen software (Jahufer and Luo 2018; https://deltagen.agresearch.co.nz/app/deltagen). Linear mixed models were used in the analyses, using the REML algorithm. The estimated genotypic means were based on best linear unbiased predictors (BLUPs) (White and Hodge 1989).
The linear mixed model was used for analyses within individual locations across seasons and years:
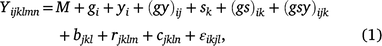
where Yijklmn is the value of an attribute measured from entry i in row m and column n of replicate l nested in season k in year j. Here i = 1, …, ng; j = 1, …, ny; k = 1, …, ns; l = 1, …, nb; m = 1, …, nr; and n = 1, …, nc; where g, y, s, b, r and c are entries, years, seasons, replicates, rows and columns, respectively. M is the overall mean; gi is the random effect of entry i, 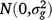 ; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j,
; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j, 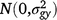 ; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k,
; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k, 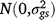 ; (gsy)ijk is the effect of the interaction between entry i, season k and year j,
; (gsy)ijk is the effect of the interaction between entry i, season k and year j, 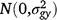 ; bjkl is the random effect of replicate l within season k in year j,
; bjkl is the random effect of replicate l within season k in year j, 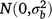 ; rjklm is the random effect of row m within replicate l within season k in year j,
; rjklm is the random effect of row m within replicate l within season k in year j, 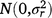 ; cjkln is the random effect of column n within replicate l within season k in year j,
; cjkln is the random effect of column n within replicate l within season k in year j, 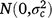 ; and εijklmn is the residual effect of entry i in row m and column n of replicate l during season k in year j,
; and εijklmn is the residual effect of entry i in row m and column n of replicate l during season k in year j, 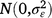 .
.
For the within individual season analysis across locations and years the linear model was similar to Eqn 1, except that sk, season, was replaced by lk, location.
The linear mixed model was used for analysis across all years, seasons and locations:
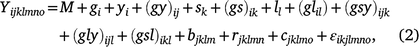
where Yijklmno is the value of an attribute measured from entry i in row n and column o of replicate m nested in location l in season k in year j. Here i = 1, …, ng; j = 1, …, ny; k = 1, …, ns; l = 1, …, nl; m = 1, …, nb; n = 1, …, nr; o = 1, …, nc; where g, y, s, l, b, r and c are entries, years, seasons, locations, replicates, rows and columns, respectively. M is the overall mean; gi is the random effect of entry i, 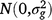 ; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j,
; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j, 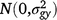 ; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k,
; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k, 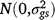 ; ll is the fixed effect of location l; (gl)il is the effect of the interaction between entry i and location l,
; ll is the fixed effect of location l; (gl)il is the effect of the interaction between entry i and location l, 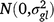 ; (gsy)ijk is the effect of the interaction between entry i, season k and year j,
; (gsy)ijk is the effect of the interaction between entry i, season k and year j, 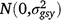 ; (gly)ijl is the effect of the interaction between entry i, location l and year j,
; (gly)ijl is the effect of the interaction between entry i, location l and year j, 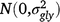 ; (gsl)ikl is the effect of the interaction between entry i, season k and location l,
; (gsl)ikl is the effect of the interaction between entry i, season k and location l, 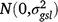 ; bjklm is the random effect of replicate m in location l within season k in year j,
; bjklm is the random effect of replicate m in location l within season k in year j, 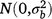 ; rjkln is the random effect of row n within replicate m in location l within season k in year j,
; rjkln is the random effect of row n within replicate m in location l within season k in year j, 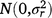 ; cjklo is the random effect of column o within replicate m in location l within season k in year j,
; cjklo is the random effect of column o within replicate m in location l within season k in year j, 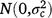 ; and εijklmno is the residual effect of entry i in row n and column o of replicate m in location l during season k in year j,
; and εijklmno is the residual effect of entry i in row n and column o of replicate m in location l during season k in year j, 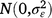 .
.
The linear mixed model was used for analysis across the three grazing management types, all seasons and years:
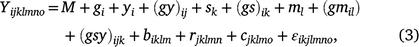
where Yijklmno is the value of an attribute measured from entry i in row n and column o of replicate m nested in grazing management l in season k in year j. Here i = 1, …, ng; j = 1, …, ny; k = 1, …, ns; l = 1, …, nl; m = 1, …, nb; n = 1,…, nr; o = 1, …, nc; where g, y, s, l, b, r and c are entries, years, seasons, grazing management, replicates, rows and columns, respectively. M is the overall mean; gi is the random effect of entry i, 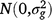 ; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j,
; yj is the fixed effect of year j; (gy)ij is the effect of the interaction between entry i and year j, 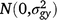 ; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k,
; sk is the fixed effect of season k; (gs)ij is the effect of the interaction between entry i and season k, 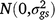 ; ll is the fixed effect of grazing management l; (gl)il is the effect of the interaction between entry i and grazing management location l,
; ll is the fixed effect of grazing management l; (gl)il is the effect of the interaction between entry i and grazing management location l, 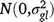 ; (gsy)ijk is the effect of the interaction between entry i, season k and year j,
; (gsy)ijk is the effect of the interaction between entry i, season k and year j, 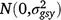 ; bjklm is the random effect of replicate m in grazing management l within season k in year j,
; bjklm is the random effect of replicate m in grazing management l within season k in year j, 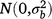 ; rjkln is the random effect of row n within replicate m in grazing management l within season k in year j,
; rjkln is the random effect of row n within replicate m in grazing management l within season k in year j, 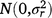 ; cjklo is the random effect of column o within replicate m in grazing management l within season k in year j,
; cjklo is the random effect of column o within replicate m in grazing management l within season k in year j, 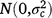 ; and εijklmno is the residual effect of entry i in row n and column o of replicate m in grazing management l during season k in year j,
; and εijklmno is the residual effect of entry i in row n and column o of replicate m in grazing management l during season k in year j, 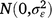 .
.
The linear models used in the analysis of entry performance within the beef cattle, dairy cattle and sheep grazing type managements, and across the beef cattle/sheep and beef cattle/dairy cattle management combinations, across locations and years, were a reduced form of Eqn 3.
Pattern analysis
Pattern analysis, a combination of cluster analysis and principal component analysis, was conducted by using DeltaGen to provide graphical summary of the performance of the 56 white clover entry-by-growth BLUP matrices generated from the variance component analyses: within individual locations; within individual seasons; within individual years; across all years, seasons and locations; grazing management type. Each of the entry-by-growth BLUP matrices, from the different variance component analyses, was summarised by using cluster analysis to generate cultivar groups. This was followed by principal component analysis (ordination) of the same white clover entry-by-growth BLUP matrices to generate biplots. In DeltaGen, each of the entry groups identified from clustering is assigned a different colour and superimposed on the biplot. This resulted in a graphical summary of information within each of the entry-by-growth BLUP matrices.
In DeltaGen, cluster analysis is performed using a hierarchical agglomerative classification procedure with squared Euclidean distance as a measure of dissimilarity (Burr 1968, 1970; Wishart 1969) and the Hartigan clustering algorithm (Hartigan 1975) is used as the grouping strategy. Principal component analysis was conducted according to Jolliffe (2002). Before conducting cluster analysis, the data were standardised to remove scaling effects (Cooper and DeLacy 1994), using the ‘Standardisation’ option in DeltaGen.
Results
Leaf size and mean seasonal growth
There was significant (P < 0.05) genotypic variation among the white clover entries for mean leaf size across all seasons, years and locations Kerikeri, Aorangi and Lincoln (Table 2). Genotype × location interaction for leaf size was also significant (P < 0.05). There was significant (P < 0.05) genotypic variation for mean seasonal growth across 4 years among the white clover entries at each of the locations Kerikeri, Ruakura, Aorangi and Lincoln (Table 2). Genotype × year interactions were significant (P < 0.05) at all locations. Significant (P < 0.05) genotype × season interaction was estimated only at Ruakura and Aorangi. There was significant (P < 0.05) genotype × season × year interaction at Lincoln. These interactions indicated that there was a change in the relative performance among the white clover entries across seasons and years, depending on the location. Analysis of mean seasonal growth of the 56 entries across 4 years and four locations indicated significant (P < 0.05) genotypic variation among them. The interactions of the entries with years, locations and seasons were all significant (P < 0.05).

|
The range of mean leaf size scores, sorted from small to large, based on evaluation of the 56 white clover entries across 4 years and three locations is presented in Fig. 1; the means of only 55 entries are shown, with cv. AberDance excluded owing to error. Cvv. Huia, Prestige, Bounty, AberAce, Demand and Tribute showed leaf sizes at the lower end of the score scale (i.e. small). Cvv. Apex, Sustain, Klondike and Riesling showed small–medium to medium leaf size. Cvv. Kopu II, Quartz and Legacy had leaf sizes within the medium–large range of the score scale. Cv. Legacy had the largest leaf size among the 13 cultivars. There was a range of leaf sizes among the experimental synthetics from small (51, 10, 52, 56, 1, 4) to large (33, 37, 36).

|
Cluster analysis of the 56 white clover entry-by-multi-location growth BLUP matrix generated four entry groups (Fig. 2). Groups 1 and 2 had above-average performance across the four locations Lincoln, Aorangi, Ruakura and Kerikeri. Group 1 had a higher average performance than Group 2, and consisted of 11 experimental synthetics (8, 23, 25, 26, 27, 28, 33, 35, 36, 37, 38) and large-leaf cv. Legacy. Synthetics 27, 35, 37 and Legacy showed broad adaptation across the four locations. In Group 1, whereas synthetics 25 and 36 showed specific adaptation to Lincoln, synthetics 33, 23 and 28 had specific adaptation to Kerikeri.

|
Group 2 consisted of 17 experimental synthetics and two cultivars, Quartz and Kopu II. Quartz showed specific adaptation to the sheep-grazed location, Lincoln. This cultivar also had above-average performance across all four locations. Kopu II showed above-average performance across Kerikeri, Ruakura and Aorangi. Its performance under sheep grazing at Lincoln was below average. Based on the association among the directional vectors, location pairs Kerikeri and Ruakura, Ruakura and Aorangi, and Aorangi and Lincoln showed positive correlation (angle between the vectors <90°). There was a weak positive association between Kerikeri and Lincoln. Note that these relationships among the four locations are based on mean seasonal growth of the 56 white clover entries across the 4 years.
Growth within individual seasons across locations
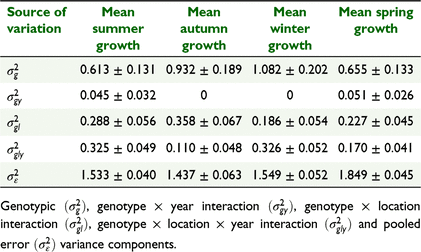
Variance component analysis across locations and years within each season indicated significant (P < 0.05) genotypic variation among the 56 white clover entries for summer, autumn, winter and spring growth (Table 3). There was significant (P < 0.05) genotype × location interaction within each season. There was also significant (P < 0.05) genotype × location × year interaction within the summer, autumn, winter and spring seasons. There was no significant (P > 0.05) genotype × year interaction.
|
Fig. 3 provides a graphical summary of mean individual seasonal growth of the 56 white clover entries across the four locations and 4 years. The directional vectors indicate a positive association among all four seasons (angle between vectors <90°); in particular, spring and autumn showed the 56 entries having a similar average performance across the four locations and 4 years. Cluster analysis of the 56 entry-by-four seasons growth BLUP matrix generated four entry groups. Group 1, which had above-average seasonal growth, consisted of 21 experimental synthetics and cvv. Quartz and Legacy. Some members, including Quartz, showed broad seasonal adaptation. Synthetics 23, 24, 28, 34 and 38 had highly above-average winter growth; and synthetics 20, 29, 33 and 56 showed highly above-average summer growth. Whereas cv. Quartz showed above-average performance across all seasons, cv. Legacy showed highly above-average summer seasonal performance. Cv. Kopu II, a member of Group 2, also showed above-average growth in summer, but had below-average winter growth.

|
Grazing type and seasonal growth
Analysis of mean seasonal growth across all years, seasons and locations within each grazing type and combinations of dairy cattle/sheep and beef cattle/dairy cattle indicated significant (P < 0.05) genotypic variation among the 56 entries (Table 4). There was significant (P < 0.05) genotype × year interaction estimated for all the grazing types and grazing combinations. Although there was significant (P < 0.05) genotype × grazing type (dairy cattle/sheep and beef cattle/dairy cattle) interaction, other contributors to this variance component could also be location-specific, abiotic and biotic effects.

|
Variance component analysis of mean seasonal growth in Years 2–4 within each of the grazing type managements indicated significant (P < 0.05) genotypic variation among the 56 white clover entries (Table 5). For the cattle grazing type management at two locations Kerikeri and Aorangi, there was significant (P < 0.05) genotype × location interaction in Years 2–4.

|
Performance of the 56 white clover entries under beef cattle, dairy cattle and sheep grazing, based on mean seasonal growth in individual Years 2, 3 and 4 of the multi-location trials is graphically summarised in the biplots (Fig. 4a–c). Cluster analysis of the entry-by-mean individual year growth score BLUPs, estimated from the beef cattle grazed data collected across two locations, Kerikeri and Aorangi, generated three entry groups (Fig. 4a). Group 1 with above-average performance across all years had 27 members and included cvv. Quartz and Legacy. Synthetics 23, 26, 29 and 33 showed highly above-average performance across the three years. Quartz showed above-average performance, especially in Year 4. Legacy had above-average in Years 2 and 3 but was average in Year 4. Cv. Kopu II, in Group 2, showed above-average performance in Year 4. There were positive correlations between the years; however, Years 2 and 3 had the closest association.

|
Under dairy cattle grazing (Fig. 4b), cluster analysis revealed six groups. Group 6, the highly above-average group, consisted of 10 experimental synthetics of which 33, 35, 36 and 42, along with cv. Legacy, had highly above-average performance across all years. Group 5, showing above-average performance, consisted of 14 experimental synthetics and cvv. Sustain, Quartz and Kopu II. Of the Group 5 cultivars, Kopu II had slightly above-average performance across Years 2–4, and Quartz, with a higher performance in Years 2 and 3, had lower performance in Year 4. Experimental synthetics 8, 36, 37 and 45 from Group 6 and 9, 20 and 21 from Group 5 showed highly above-average performance in Year 4.
Under sheep grazing (Fig. 4c), three entry groups were observed from cluster analysis. Group 1 consisted of 19 experimental synthetics and seven cultivars: AberDance, Apex, Demand, Legacy, Prestige, Quartz and Riesling. This group comprised entries that had highly above-average performance across all three years, and entries with high performance specific to Years 3 and 4. Synthetics 15, 23, 33 and 48 had highly above-average performance across all years. Cv. Quartz had the highest above-average performance across all years. Cv. AberDance also had highly above-average performance across years but lower than Quartz. Cvv. Apex, Demand, Legacy, Prestige and Riesling showed above- to highly above-average performance in Years 3 and 4. There was a slight negative correlation (angle between the vectors >90°) between Year 2 and Years 3 and 4, whereas Years 3 and 4 were highly positively correlated (angle between the vectors <90°).
The combination of cluster and principal component analysis of the 56 white clover entry-by-seasonal growth across all years BLUP matrix within each grazing management type generated a graphical summary (Fig. 5). There was a strong similarity in performance of the 56 entries under beef cattle and dairy cattle grazing, as indicated by the close association (angle between the vectors <90°) between the two directional vectors Dairy and Cattle (Fig. 5). As indicated by the directional vectors, response of the 56 entries to sheep grazing was different to that under dairy and cattle grazing. Cluster analysis revealed five entry groups. Group 1 consisted of five small to medium–large leaf cultivars, Prestige (S), Demand (SM), AberDance (M), Apex (M) and Riesling (ML), and experimental synthetic 54. This group of entries showed highly above-average performance specific to sheep grazing. Group 5 consisted of eight synthetics and medium–large leaf cv. Quartz. In Group 5, synthetics 18, 22, 39, 44, 48 and 49 had above-average performance under sheep grazing, whereas Quartz and synthetics 15 and 45 showed highly above-average performance. Cv. Quartz and synthetics 15, 48, 49, 44, 22 and 18, also had above-average performance under dairy and cattle grazing. All members of Group 4 had highly above-average performance under both dairy and cattle grazing management. Experimental synthetics 27, 33, and 38 showed highly above-average performance under all three grazing managements. The members of Group 3 showed specific adaptation to both dairy and cattle grazing management. This varied from above-average to highly above-average performance. The large-leaf cv. Kopu II had above-average performance. Cv. Bounty, a member of Group 2, showed average performance under sheep grazing and below-average performance under both cattle and dairy grazing management.

|
Discussion
Analysis of the performance of the 56 white clover entries consisting of 42 experimental synthetics and 14 cultivars, evaluated for seasonal growth across four locations over 4 years, showed significant genotype × environment interaction at different levels: genotype × year interaction 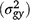 , genotype × location interaction
, genotype × location interaction 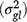 , genotype × season interaction
, genotype × season interaction 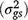 , genotype × location × year interaction
, genotype × location × year interaction 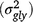 and genotype × season × year interaction
and genotype × season × year interaction 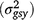 , depending on the season, location and year combinations. There was also significant genotype × location interaction estimated for leaf size. These results clearly emphasise the importance of conducting multi-year–season–location evaluation trials in white clover breeding programs, especially under different grazing management types, as reported in this study: beef cattle, dairy cattle and sheep, under rotational grazing. Trials across multiple locations, years and seasons are especially important when selecting for broad adaptation (Cooper and DeLacy 1994; Cooper and Byth 1996). There have been several reports on genotype × environment interaction for a range of traits in white clover. Jahufer et al. (2013) reported significant genotype × year interaction for mean seasonal yield in white clover. Jahufer et al. (1999) reported significant full-sib family × environment × year interaction for a range of morphological traits including stolon density, stolon branching, number of nodes, rooted node number and summer herbage yield among 80 white clover full-sibs. Significant genotype × environment interactions for a range of white clover morphological traits such as internode length, leaf size, plant height, stolon density and dry matter yield have also been reported (Jahufer et al. 2009, 2016; Annicchiarico and Piano 2000; Finne et al. 2000). Significant line × year interactions for the proportion of clover in sward, clover dry weight and leaf size was estimated from evaluation of elite breeding lines in two trials rotationally grazed, one by sheep and the other by cattle, across 3 years (Caradus 1993).
, depending on the season, location and year combinations. There was also significant genotype × location interaction estimated for leaf size. These results clearly emphasise the importance of conducting multi-year–season–location evaluation trials in white clover breeding programs, especially under different grazing management types, as reported in this study: beef cattle, dairy cattle and sheep, under rotational grazing. Trials across multiple locations, years and seasons are especially important when selecting for broad adaptation (Cooper and DeLacy 1994; Cooper and Byth 1996). There have been several reports on genotype × environment interaction for a range of traits in white clover. Jahufer et al. (2013) reported significant genotype × year interaction for mean seasonal yield in white clover. Jahufer et al. (1999) reported significant full-sib family × environment × year interaction for a range of morphological traits including stolon density, stolon branching, number of nodes, rooted node number and summer herbage yield among 80 white clover full-sibs. Significant genotype × environment interactions for a range of white clover morphological traits such as internode length, leaf size, plant height, stolon density and dry matter yield have also been reported (Jahufer et al. 2009, 2016; Annicchiarico and Piano 2000; Finne et al. 2000). Significant line × year interactions for the proportion of clover in sward, clover dry weight and leaf size was estimated from evaluation of elite breeding lines in two trials rotationally grazed, one by sheep and the other by cattle, across 3 years (Caradus 1993).
Leaf size is commonly used to classify white clover cultivars (Frame and Laidlaw 2005) and is also highly related to grazing system fit (i.e. beef/dairy cattle or sheep). Small leaf types are considered suitable for set-stocked sheep grazing, medium types for use under rotational sheep grazing, and medium–large to very large types for rotational beef cattle grazing (Abberton and Marshall 2005). Leaf size provides an indirect assessment of white clover plant type because of its positive and negative correlation with morphological traits associated with growth and vegetative persistence. For example, leaf size has a positive correlation with plant height and stolon diameter, and a negative correlation with stolon number (Jahufer et al. 1994; Abberton and Marshall 2005). Small leaf types are positively correlated with high stolon densities, associated with vegetative persistence (Caradus and Williams 1981; Piano and Annicchiarico 1995).
In our study, the mean leaf size scores measured from 13 cultivars across years and locations did not generally reflect those reported in the literature (Woodfield et al. 2003; Widdup et al. 2015; Gerard et al. 2017). For example, results from our experiment assigned cvv. Huia (medium leaf size), Bounty (medium leaf size), AberAce (medium leaf size), Demand (small–medium leaf size) and Tribute (medium leaf size) within the small leaf size range. Cv. Kopu II, considered as a large leaf type, scored as medium–large in our trials. This deviation of leaf size may be attributed to the plasticity of white clover under grazing (Caradus 1993; Brock and Hay 1996, 2001). The significant genotype × location and genotype × location × year interactions estimated for leaf size in our study further support these observations. There was a range of leaf sizes among the experimental synthetics, from small to large. Synthetics 51, 10 and 52 were in the small leaf category, and 33, 37 and 36 were in the large leaf category. This information is an important criterion for identifying experimental synthetics suitable for cattle, dairy and sheep grazing management systems.
Pattern analysis provided graphical summarises on performance of the 56 entries based on seasonal growth: (i) across the four locations Kerikeri, Ruakura, Aorangi and Lincoln; (ii) within seasons spring, summer, autumn and winter; (iii) during Years 2–4 within each grazing type; and (iv) across seasons and years within grazing type, beef cattle, dairy cattle and sheep, all under rotational grazing management. The biplot summarising mean seasonal growth of the 42 synthetics and 14 cultivars across the four locations clearly indicated broad adaptation of the synthetics 27, 35 and 37 and cv. Legacy. Cv. Quartz showed above-average specific adaptation at the sheep-grazed location, Lincoln. This cultivar also showed slightly above-average performance at Aorangi, Ruakura and Kerikeri. Cv. Kopu II showed broad adaptation across the beef cattle and dairy cattle grazed locations. Experimental synthetics 8, 27, 35 and 37, and cv. Legacy, showed superior broad adaptation across all three types of grazing management compared with cvv. Kopu II and Quartz.
Several experimental synthetics showed broad adaptation across all four seasons and were superior to cv. Quartz, which had above-average adaptation. Synthetics such as 23, 24, 28, 34 and 38 showed highly above-average winter growth. Synthetics 20, 29, 33 and 56, showed highly above-average summer growth. Compared with cv. Quartz, Kopu II showed low above-average growth in summer, spring and autumn. Kopu II had below-average winter growth. Cv. Legacy had highly above-average summer growth.
The perennial behaviour of white clover, an important selection criterion in cultivar development, is attributed to continuous vegetative growth of stolons (Westbrooks and Tesar 1955; Caradus and Williams 1989). During the first 12 months of growth, the white clover plant loses its primary seedling taproot and depends on nodal roots that support future stolon growth (Westbrooks and Tesar 1955). These significant changes in plant development make Year 1 performance an unreliable predictor of future vegetative persistence, and therefore, it was not included in the assessment of vegetative persistence across the 4 years in our study. Only years 2–4 were considered. Performance of the 56 white clover entries in Year 4 was of particular importance because it provided an assessment of overall vegetative persistence across the 4 years of evaluation within each grazing management type. Several experimental synthetics had above-average performance in Year 4 under the three grazing management types. Experimental synthetic 33 (large leaf size) showed superior performance in Year 4 under all three grazing managements. Synthetics 27 and 38, both with medium–large leaf size, had above-average performance in Year 4 under beef cattle and sheep grazing.
Our investigation enabled identification of cultivars and experimental synthetics with specific and broad adaptation to the three grazing management types. The small to medium–large leaf cultivars, AberDance (M), Apex (M), Demand (SM), Prestige (S) and Riesling (ML), showed highly above-average performance under sheep grazing. Some of these small to medium leaf cultivars with high stolon densities have been developed for sheep grazing (Caradus and Woodfield 1997; Woodfield et al. 2001). Quartz and synthetics 15 and 45 had highly above-average performance under sheep grazing. Synthetics 15, 48, 49, 44, 22 and 18 and Quartz also had above-average performance under all three grazing management types. Cvv. Legacy and Kopu II showed above-average performance under cattle and dairy grazing. There were also several synthetics with highly above-average performance under dairy and cattle grazing: 8, 25, 34, 35, 36, 37, 42, 47. Synthetics 27, 33 and 38 had highly above-average performance across all three grazing managements and were superior to the 14 cultivars evaluated. Some of these superior synthetics are currently being tested across multiple grazing environments to assess their merit.
Considering seasonal growth response of the 14 white clover cultivars evaluated in our study, Quartz and Legacy showed the best performance under all three grazing management types. However, Legacy showed better response to cattle and dairy grazing, whereas Quartz showed superior performance under sheep grazing and evidence of broad adaptation. Legacy and Quartz were superior to the other 12 cultivars evaluated: AberAce, AberDance, Apex, Bounty, Demand, Huia, Klondike, Kopu II, Prestige,Riesling, Sustain, Tribute.
Quartz was developed by combining material from the breeding populations of cvv. Saracen, Trophy and Tribute, with persistent ecotypes selected from dairy farms in the Waikato region of New Zealand that had known history of pasture renewal for >70 years. Cvv. Saracen and Trophy were developed from elite drought-tolerant breeding lines generated from a successful New Zealand/Australia trans-Tasman collaboration (Ayres et al. 2007; Jahufer et al. 2012). The development of cv. Quartz was based on combining a diverse range of genetic material to generate a cultivar with adaptation across New Zealand grazing systems. Results from our multi-location and multi-year field study under beef cattle, dairy cattle and sheep grazing have demonstrated the merit of Quartz for broad adaptation under New Zealand conditions. This cultivar is currently being evaluated in several on-farm trials across temperate regions of the world.
Data availability
All data analysed will be made available as requested. However, information on breeding history and pedigree of the 42 experimental synthetics reported in this manuscript is confidential and will not be made available.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
We acknowledge Grasslands Innovation Limited for the financial support.
Acknowledgements
Special thanks to Angus Heslop, Keith Widdup, Ben Harvey, Doug Ryan, Phil Olyott and Kathryn Panckhurst for their significant contribution to trial planting, establishment, maintenance, and data collection.
References
Abberton MT, Marshall AH (2005) Progress in breeding perennial clovers for temperate agriculture. The Journal of Agricultural Science 143, 117–135.| Progress in breeding perennial clovers for temperate agriculture.Crossref | GoogleScholarGoogle Scholar |
Annicchiarico P, Piano E (2000) Response of white clover genotypes to evaluation environments of dense and spaced planting, and implications for selection. Euphytica 111, 111–120.
| Response of white clover genotypes to evaluation environments of dense and spaced planting, and implications for selection.Crossref | GoogleScholarGoogle Scholar |
Ayres JF, Caradus JR, Murison RD, Lane LA, Woodfield DR (2007) Grasslands Trophy: a new white clover (Trifolium repens L.) cultivar with tolerance of summer moisture stress. Australian Journal of Experimental Agriculture 47, 110–115.
| Grasslands Trophy: a new white clover (Trifolium repens L.) cultivar with tolerance of summer moisture stress.Crossref | GoogleScholarGoogle Scholar |
Barbour M, Caradus JR, Woodfield DR, Silvester WB (1996) Water stress and water use efficiency of ten white clover cultivars. In ‘White clover: New Zealand’s competitive edge’. (Ed. DR Woodfield) pp. 159–162. (New Zealand Grassland Association: Mosgiel, New Zealand)
Brock JL, Hay MJM (1996) A review of the role of grazing management on the growth and performance of white clover cultivars in lowland New Zealand pastures. In ‘White clover: New Zealand’s competitive edge’. (Ed. DR Woodfield) pp. 65–70. (New Zealand Grassland Association: Mosgiel, New Zealand)
Brock JL, Hay MJM (2001) White clover performance in sown pastures: a biological/ecological perspective. Proceedings of the New Zealand Grassland Association 63, 73–83.
| White clover performance in sown pastures: a biological/ecological perspective.Crossref | GoogleScholarGoogle Scholar |
Brock JL, Hyslop MG, Widdup KH (2003) A review of red and white clovers in the dryland environment. In ‘Legumes for dryland pastures’. New Zealand Grassland Research and Practice Series 11, pp. 101–108. (New Zealand Grassland Association: Mosgiel, New Zealand)
Burr EJ (1968) Cluster sorting with mixed character types. I. Standardization of character values. Australian Computer Journal 1, 97–99.
Burr EJ (1970) Cluster sorting with mixed character types. II. Fusion strategies. Australian Computer Journal 2, 98–103.
Caradus JR (1993) White clover breeding line performance under sheep and cattle grazing. In ‘Proceedings of the 10th Australian plant breeding conference’. Gold Coast, Qld. Vol. II. pp. 35–36. (Conference Organising Committee: Canberra)
Caradus JR, Williams WM (1981) Breeding for improved white clover production in New Zealand hill country. In ‘Plant physiology and herbage production’. British Grasslands Society Occasional Symposium No. 13. (Ed. CF Wright) pp. 163–168. (British Grasslands Society: Nottingham, UK)
Caradus JR, Williams WM (1989) Breeding for legume persistence in New Zealand. In ‘Persistence of forage legumes’. (Eds GS Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 523–540. (American Society of Agronomy: Madison, WI, USA)
Caradus JR, Woodfield DR (1997) Review: World checklist of white clover varieties II. New Zealand Journal of Agricultural Research 40, 115–206.
| Review: World checklist of white clover varieties II.Crossref | GoogleScholarGoogle Scholar |
Caradus JR, Hay RJM, Woodfield DR (1996a) The positioning of white clover cultivars in New Zealand. In ‘White clover: New Zealand’s competitive edge’. (Ed. DR Woodfield) pp. 45–49. (New Zealand Grassland Association: Mosgiel, New Zealand)
Caradus JR, Woodfield DR, Stewart AV (1996b) Overview and vision for white clover. In ‘White clover: New Zealand’s competitive edge’. (Ed. DR Woodfield) pp. 1–6. (New Zealand Grassland Association: Mosgiel, New Zealand)
Clark DA, Matthew C, Crush JR (2001) More feed for New Zealand dairy systems. Proceedings of the New Zealand Grassland Association 63, 283–288.
Cooper M, Byth DE (1996) Understanding plant adaptation to achieve systematic applied crop improvement: a fundamental challenge. In ‘Plant adaptation and crop improvement’. (Eds M Cooper, GL Hammer) pp. 5–23. (CAB International: Wallingford, UK)
Cooper M, DeLacy IH (1994) Relationships among analytical methods used to study genotypic variation and genotype-by-environment interaction in plant breeding multi-environment experiments. Theoretical and Applied Genetics 88, 561–572.
| Relationships among analytical methods used to study genotypic variation and genotype-by-environment interaction in plant breeding multi-environment experiments.Crossref | GoogleScholarGoogle Scholar | 24186111PubMed |
Cowie J (1978) Soils and agriculture of Kiranga County, North Island, New Zealand. New Zealand Soil Bureau Bulletin 33. New Zealand Soil Bureau, Lower Hutt, New Zealand.
Eerens JPJ, Cooper BM, Willoughby BE, Woodfield DR (2001) Searching for clover root weevil (Sitona lepidus) resistance/tolerance: a progress report. Proceedings of the New Zealand Grassland Association 63, 177–181.
| Searching for clover root weevil (Sitona lepidus) resistance/tolerance: a progress report.Crossref | GoogleScholarGoogle Scholar |
Finne MA, Rognli OA, Schjelderup I (2000) Genetic variation in a Norwegian germplasm collection of white clover (Trifolium repens L.). Euphytica 112, 33–44.
| Genetic variation in a Norwegian germplasm collection of white clover (Trifolium repens L.).Crossref | GoogleScholarGoogle Scholar |
FitzGerald RD, Clark S (1993) Grazing management for white clover in temperate pastures for sheep and beef cattle. In ‘White clover: a key to increasing milk yields’. (Ed. W Mason). pp. 73–77. (Dairy Research and Development Corporation: East Melbourne, Vic., Australia)
Ford JL, Cousins GR, Jahufer Z, Baird IJ, Woodfield DR, Barrett BA (2015) Grasslands Legacy: a new, large-leaved white clover cultivar with broad adaption. Journal of New Zealand Grasslands 77, 211–218.
| Grasslands Legacy: a new, large-leaved white clover cultivar with broad adaption.Crossref | GoogleScholarGoogle Scholar |
Frame J, Laidlaw S (2005) Prospects for temperate forage legumes. In ‘Grasslands: developments, opportunities and perspectives’. (Eds EG Reynolds, J Frame). pp. 1–28. (Science Publishers: Enfield, CT, USA)
Gerard PJ, Ferguson CM, van Amsterdam S (2017) Comparison of New Zealand perennial clovers for resilience against common pasture pests. New Zealand Plant Protection 70, 241–249.
| Comparison of New Zealand perennial clovers for resilience against common pasture pests.Crossref | GoogleScholarGoogle Scholar |
Gramshaw D, Read JW, Collins WJ, Carter ED (1989) Sown pastures and legume persistence: an Australian overview. In ‘Persistence of forage legumes’. (Eds GS Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 523–540. (American Society of Agronomy: Madison, WI, USA)
Hartigan JA (1975) ‘Clustering algorithms’. (John Wiley & Sons: Hoboken, NJ, USA)
Harville DH (1977) Maximum likelihood approaches to variance component estimation and related problems. Journal of the American Statistical Association 72, 320–338.
| Maximum likelihood approaches to variance component estimation and related problems.Crossref | GoogleScholarGoogle Scholar |
Hewitt AE (1993) ‘New Zealand Soil Classification’, Landcare Research Science Series No. 1. (Manaaki-Whenua Press: Lincoln, New Zealand)
Hutchinson KJ (1993) White clover in the competitive environment of the pasture sward. In ‘White clover: a key to increasing milk yields’. (Ed. W. Mason) pp. 95–99. (Dairy Research and Development Corporation: East Melbourne, Vic., Australia)
Jahufer MZZ, Luo D (2018) DeltaGen: A comprehensive decision support tool for plant breeders. Crop Science 58, 1118–1131.
| DeltaGen: A comprehensive decision support tool for plant breeders.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Cooper M, Brien LA (1994) Genotypic variation for stolon and other morphological attributes of white clover (Trifolium repens L.). Populations and their influence on herbage yield in the summer rainfall region of New South Wales. Australian Journal of Agricultural Research 45, 703–720.
| Genotypic variation for stolon and other morphological attributes of white clover (Trifolium repens L.). Populations and their influence on herbage yield in the summer rainfall region of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Lane LA, Ayres JF (1996) White clover improvement for Australian dryland environments. 1. The white clover collection. Australian Plant Introduction Review 26, 1–15.
Jahufer MZZ, Cooper M, Bray RA, Ayres JF (1999) Evaluation of white clover (Trifolium repens L.) populations for summer moisture stress adaptation in Australia. Australian Journal of Agricultural Research 50, 561–74.
| Evaluation of white clover (Trifolium repens L.) populations for summer moisture stress adaptation in Australia.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Clements R, Durant R, Woodfield DR (2009) Evaluation of white clover (Trifolium repens L.) commercial cultivars and experimental synthetics in south-west Victoria, Australia. New Zealand Journal of Agricultural Research 52, 407–415.
| Evaluation of white clover (Trifolium repens L.) commercial cultivars and experimental synthetics in south-west Victoria, Australia.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Ford JL, Widdup KH, Harris C, Cousins G, Ayres JF, Lane LA, Hofmann RW, Ballizany WL, Mercer CF, Crush RJ, Williams WM, Woodfield DR, Barrett BA (2012) Improving white clover for Australasia. Crop & Pasture Science 63, 739–745.
| Improving white clover for Australasia.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Dunn A, Baird I, Ford JL, Griffiths AG, Jones CS, Woodfield DR, Barrett BA (2013) Genotypic variation for morphological traits in a white clover mapping population evaluated across two environments and three years. Crop Science 53, 460–472.
| Genotypic variation for morphological traits in a white clover mapping population evaluated across two environments and three years.Crossref | GoogleScholarGoogle Scholar |
Jahufer MZZ, Ford JL, Woodfield DR, Barrett BA (2016) Genotypic evaluation of introduced white clover (Trifolium repens L.) germplasm in New Zealand. Crop & Pasture Science 67, 897–906.
| Genotypic evaluation of introduced white clover (Trifolium repens L.) germplasm in New Zealand.Crossref | GoogleScholarGoogle Scholar |
John JA (1987) Row–column designs. In ‘Cyclic designs’. Monographs on Statistics and Applied Probability. (Springer: Boston, MA, USA)
| Crossref |
Jolliffe IT (2002) ‘Principal component analysis. Vol. XXIX’, 2nd edn. Springer Series in Statistics. (Springer-Verlag: New York, NY, USA)
Kemp PD, Kenyon PR, Morris ST (2010) The use of legume and herb forage species to create high performance pastures for sheep and cattle grazing systems. Revista Brasileira de Zootecnia 39, 169–174.
| The use of legume and herb forage species to create high performance pastures for sheep and cattle grazing systems.Crossref | GoogleScholarGoogle Scholar |
Knowles IM, Fraser TJ, Daly MJ (2003) White clover: plant loss in drought and subsequent recovery after rain. In ‘Legumes for dryland pastures. Proceedings of New Zealand Grassland Association symposium’. Grassland Research and Practice Series No. 11. (Ed. DJ Moot) pp. 37–42. (New Zealand Grassland Association: Wellington, New Zealand)
Lowe DJ (2010) Introduction to the landscapes and soils of the Hamilton Basin. In ‘Guidebook for pre-conference North Island, New Zealand ‘Volcanoes to oceans’ field tour (27–30 July). 19th World soils congress, International Union of Soil Sciences, Brisbane’. Soil and Earth Sciences Occasional Publication No. 3. (Eds DJ Lowe, VF Neall, M Hedley, B Clothier, A Mackay) pp. 1.24–1.61. (Massey University: Palmerston North, New Zealand)
Mercer CF, Bell NL, Yeates GW (2008) Plant-parasitic nematodes on pasture in New Zealand. Australasian Plant Pathology 37, 279–288.
| Plant-parasitic nematodes on pasture in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Newton PCD, Edwards GR (2007) Plant breeding for a changing environment. In ‘Agroecosystems in a changing climate’. (Eds PCD Newton, RA Carran, GR Edwards, PA Niklaus) pp. 309–319. (Taylor & Francis: London, UK)
Patterson HD, Thompson R (1971) Recovery of inter-block information when block sizes are unequal. Biometrika 58, 545–554.
| Recovery of inter-block information when block sizes are unequal.Crossref | GoogleScholarGoogle Scholar |
Patterson HD, Thompson R (1975) Maximum likelihood estimation of components of variance. In ‘Proceedings of the 8th International Biometric Conference’. pp. 197–207.
Piano E, Annicchiarico P (1995) Persistence of Ladino white clover ecotypes and its relationship with other agronomic traits. Grass and Forage Science 50, 195–198.
| Persistence of Ladino white clover ecotypes and its relationship with other agronomic traits.Crossref | GoogleScholarGoogle Scholar |
Taylor NH, Pohlen IJ (1968) Classification of New Zealand soils. In ‘Soils of New Zealand’. New Zealand Soil Bureau Bulletin 26(3). (Ed. J Luke) (New Zealand Soil Bureau: Lower Hutt, New Zealand)
Thomson DJ (1984) Forage legumes. British Grassland Society Occasional Symposium 16, 78–92.
Ulyatt MJ (1973) The feeding value of herbage. In ‘Chemistry and biochemistry of herbage. Vol. 3’. (Eds GW Butler, R Bailey) pp. 131–178. (Academic Press: New York, NY, USA)
Westbrooks FE, Tesar MB (1955) Tap root survival of ladino clover. Agronomy Journal 47, 403–410.
White TL, Hodge GR (1989) ‘Predicting breeding values with applications in forest tree improvement.’ Forestry Sciences 33. (Kluwer Academic: Boston, MA, USA)
Widdup KH, Ford JL, Cousins GR, Woodfield DR, Caradus JR, Barrett BA (2015) A comparison of New Zealand and overseas white clover cultivars under grazing in New Zealand. Proceedings of the New Zealand Grassland Association 77, 51–56.
Wishart D (1969) Algorithm for hierarchical classifications. Biometrics 25, 165–170.
Woodfield DR (1999) Genetic improvements in New Zealand forage cultivars. Proceedingsof the New Zealand Grassland Association 61, 3–7.
Woodfield DR, Clifford PTP, Cousins GR, Ford JL, Baird IJ, Miller JE, Woodward SL, Caradus JR (2001) Grasslands Kopu II and Apex: new generation white clovers. Proceedings of the New Zealand Grassland Association 63, 103–108.
Woodfield DR, Clifford PTP, Baird IJ, Cousins GR, Miller JE, Widdup KH, Caradus JR (2003) Grasslands Tribute: a multi-purpose white clover for Australasia. Proceedings of the New Zealand Grassland Association 65, 157–162.