

Effects of nitrogen fertilisation and irrigation on seed yield and yield components of signal grass (Urochloa decumbens)
M. W. Canto A D , E. J. PanceraA Department of Animal Science, State University of Maringá, Maringá, PR 87020-900, Brazil.
B Grazing Ecology Research Group, Federal University of Rio Grande do Sul, Porto Alegre, RS 91501-970, Brazil.
C Department of Agronomy, State University of Maringá, Maringá, PR 87020-900, Brazil.
D Corresponding author. Email: mwcanto@uem.br
Crop and Pasture Science 71(3) 294-303 https://doi.org/10.1071/CP18369
Submitted: 17 August 2018 Accepted: 23 January 2020 Published: 1 April 2020
Journal compilation © CSIRO 2020 Open Access CC BY-NC-ND
Abstract
The effects of nitrogen (N) fertiliser rate and irrigation on seed yield and its components were evaluated for signal grass (Urochloa decumbens (Stapf) R.D. Webster; syn. of Brachiaria decumbens Stapf) cv. Basilisk in a field experiment in Umuarama, Paraná, Brazil. Two water regimes (irrigated and non-irrigated) and four nitrogen (N) fertiliser rates (0, 25, 50 and 75 kg ha–1) were applied to perennial signal grass crops in a split-plot randomised complete block design with three replications. In two consecutive harvests, favourable rainfall resulted in irrigation having limited influence on most measurements, and the combined application of irrigation and N fertiliser did not improve seed yield. Compared with the nil N, the highest N application rate significantly increased seed yield for the first crop (266 vs 498 kg ha–1) and the second crop (104 vs 286 kg ha–1). Nitrogen fertilisation significantly increased number of seed per area, reproductive tiller density and plant biomass at harvest for the first and second crops. Harvest index, 1000-seed weight, reproductive tiller weight, number of spikelets per panicle and number of seeds per panicle were unaffected by N rate. Harvest index ranged from 1.10% to 3.63% and 1000-seed weight from 2.15 to 3.36 g. There were no treatment effects on number of days to flowering or anthesis. Fertilisation with 75 kg N ha–1 for the first crop and 50–75 kg N ha–1 for the second crop maximised signal grass seed yield.
Additional keywords: plant phenology, seed production, seed yield components, tropical grass.
Introduction
Signal grass (Urochloa decumbens (Stapf) R.D. Webster; syn. of Brachiaria decumbens Stapf) is a warm-season (C4) perennial grass introduced to Australia from germplasm collected in Africa (Oram 1990). In Brazil, signal grass is widely planted (Jank et al. 2014) because of its resistance to stress from grazing and water deficit, and its tolerance to acidic soil conditions. However, seed yield of signal grass is often low owing to meiotic abnormalities that impair pollen viability (Ricci et al. 2011) and poor seed retention, which leads to the production of light, immature seeds (Hopkinson et al. 1996).
Brazil is the largest producer of seeds of the genus Urochloa in the world (Jank et al. 2014). The country grows ~12 000 ha of signal grass seed crops, mainly in the Central-West Region and in the states of São Paulo, Minas Gerais and Bahia, where two harvests are possible per year (Andrade et al. 1983; Souza 2013). Despite an average annual rainfall >1000 mm, these regions are frequently affected by rainfall deficits and poor seed yields of signal grass. Inadequate seasonal rainfall and soils with low water-holding capacity, particularly sandy soils with low organic matter content, can reduce plant growth and yield stability. Nitrogen (N) availability is another key factor affecting plant growth and reproduction, even under non-limiting water conditions.
Few studies have compared the seed yield of rainfed and irrigated signal grass crops, especially at different rates of N fertilisation. In a study in Thailand, signal grass was grown from April (close cut) to August (harvest) 1995 and received irrigation only during the dry period (May) (Gobius et al. 2001). An N application rate of 200 kg ha–1 improved seed yield to only 122.6 kg ha–1. The authors observed no differences in inflorescence emergence, plant biomass at harvest, 1000-seed weight and seed purity but measured a small difference in panicle density and number of seeds per panicle among crops fertilised with 50, 100 and 200 kg N ha–1.
Research on the effects of N fertilisation on rainfed seed crops of signal grass is also scarce. Some studies found inconsistent effects of N fertilisation rates on seed yield and yield components (Carmo et al. 1988; Condé and Garcia 1988; Garcia et al. 1989). These experiments were conducted on soils with low fertility, and crop phenology was not investigated. Andrade et al. (1983) examined two signal grass crops grown in the Brazilian Cerrado by applying 50 kg N ha–1 for 3 years, and observed that seed yield increased from 88 to 260 kg ha–1 for the first crop and from 40 to 285 kg ha–1 for the second crop. However, the physiological basis for this response remains to be elucidated.
It is important to understand the combined effects of irrigation and N application on the seed yield of signal grass in order to encourage management practices that result in high productivity over greater areas; however, costs associated with these management methods must be economically justified by increasing seed yields. We hypothesised that the combination of irrigation and N fertilisation can improve the seed yield of consecutive crops of signal grass. Hence, we investigated seed yield of signal grass under different water regimes and N rates in a sandy loam soil of relatively high fertility. We also examined the effect of water regime and fertiliser application on key phenological traits, seed yield components and pre-harvest plant morphology.
Materials and methods
Site characteristics
Field experiments were conducted from October 2009 to May 2010 at the State University of Maringá, Umuarama, Paraná, Brazil (23°44′S, 53°17′W; 480 m a.m.s.l.). The soil at the site is a dystrophic Red Latosol (Santos et al. 2006), or Typic Paleudult (United States Taxonomy; Soil Survey Staff 2014), with a sandy loam texture (79% sand, 3% silt, 18% clay). The climate is humid subtropical with hot summers (Cfa, according to the Köppen climate classification; Alvares et al. 2013). Mean annual precipitation ranges from 1000 to 1500 mm (Wrege et al. 2011), with ~70% occurring during the growing season of signal grass (mid-September to March). Precipitation and daily maximum and minimum temperature data were obtained from a weather station 150 m from the experimental site.
Signal grass was established by sowing in spring 2005 following a no-till maize (Zea mays L.) crop. In August 2009, soil samples were collected from three depths (0–5, 5–10 and 10–20 cm) (Table 1). Soil pH (in CaCl2) was determined as per the SMP method (described by Mielniczuk et al. 1969). Potassium (K) and phosphorus (P) were measured by Mehlich-1. Other soil indices were hydrogen + aluminium (H + Al) (SMP method); calcium (Ca), magnesium (Mg) and Al (KCl method); SO42–-sulfur (monocalcium phosphate); zinc (Mehlich-1); and boron (hot water). Cation-exchange capacity (Ca2+ + Mg2+ + K+ + (H+ + Al3+)) at soil depths of 0–5, 5–10 and 10–20 cm was 7.65, 6.07 and 5.85 cmolc dm–3, respectively. Organic carbon content (Walkley–Black method) at soil depths of 0–5, 5–10 and 10–20 cm was 2.14, 1.33 and 1.07 g dm–3, respectively, which characterises this soil as having medium organic matter content (CQFS 2004). Soil tests indicated that P and K levels in the topsoil were adequate for crop production. Soil inorganic N concentration in the root-zone was not measured.

|
Cultural practices
All plots received a basal dressing of single superphosphate (52.4 kg P ha–1) and potassium chloride (49.8 kg K ha–1) in 2007 and 2008 to improve soil fertility. On 9 October 2009 (in preparation for the first crop) and 5 February 2010 (in preparation for the second crop), the subplots were cut to 10 cm with a mower. A few days later, the plants (seed and straw) were removed. Straw removal enhances seed yield of signal grass (Stür and Humphreys 1988). On 24 October 2009, single superphosphate (78.6 kg P ha–1) and potassium chloride (49.8 kg K ha–1) were applied to replace nutrients lost through forage removal. Fertilisers were applied to the soil surface according to local recommendations (CQFS 2004). On 1 April 2010, an organophosphate pesticide (chlorpyrifos at 1.0 L ha–1, Lorsban 480 BR; Dow AgroSciences, São Paulo) was applied to control spittlebugs.
Experimental design and treatments
Treatments were arranged in a split-plot, randomised complete block design with three replications. Whole-plot treatments were irrigation (rainfall + supplemental irrigation) and no irrigation (rainfall only), and subplot treatments consisted of N fertiliser rates of 0, 25, 50 and 75 kg N ha–1. Whole plots measured 16.0 m by 8.0 m, and subplots 4.0 m by 8.0 m. The same treatments were applied to the same subplots in the first (October 2009–January 2010) and second (February–May 2010) crop cycles.
The distance between irrigated and non-irrigated blocks was 8.0 m. The sprinkler irrigation system consisted of a motor pump and 100-mm PVC pipes with eight spray tubes. Evaporation data from a class A evaporation pan, effective precipitation, and the FAO-56 crop coefficient were used to estimate irrigation water requirements (Allen et al. 1998). Water balance (Thornthwaite and Mather 1955) was calculated for each seed-production cycle. Plants were periodically irrigated until flowering. Irrigation was discontinued after flowering on the basis of previous research (Lambert 1967; Cookson et al. 2000). In cool-season perennial grasses, many of the studies on irrigation (Lambert 1967; Hebblethwaite 1977; Chastain et al. 2014) and precipitation during anthesis (Abel and Boelt 2018) suggested that wet weather and amount and timing of precipitation could depress pollination, as well reduce yield-component development. Nitrogen as granular ammonium nitrate (340 g N kg–1) was spread manually on the soil surface at the appropriate rates for the subplots receiving 25, 50 and 75 kg N ha–1 on 30 October 2009 (21 days after mowing for the first crop) and 25 February 2010 (20 days after mowing for the second crop).
Crop phenology
It is difficult to determine the onset of anthesis in signal grass because plants can vary widely in the number of anthers per spikelet and because anthers on different spikelets can mature at different times (Hopkinson et al. 1996). We measured the time to onset of flowering and anthesis by examining reproductive tillers in the field every week from the first week of December 2009 onwards for the first crop, and from the first week of April 2010 onwards for the second crop. The onset of flowering was defined as the complete emergence of 5–10 panicles from the leaf sheath per m2. Anthesis was defined as when 30% of the reproductive tillers had florets with fully exserted anthers and stigma.
Seeds were harvested 32–38 days after the onset of flowering. The harvest date was chosen on the basis of seed maturity and seed shedding (Castro et al. 1994). The numbers of days from cutting to onset of flowering, from cutting to anthesis, from cutting to harvest (crop duration), from onset of flowering to harvest, and from anthesis to harvest (seed-filling phase) were recorded. Growing degree units (GDU) were calculated by using a base temperature of 15°C, according to the following equation (Villa Nova et al. 2007) (Eqn 1):

where Tmax is the daily maximum temperature and Tmin is the daily minimum temperature (°C).
Yield components
The number of vegetative and reproductive tillers (tillers with panicles fully exserted from the flag leaf sheath) was determined by harvesting two quadrats each 1.0 m by 1.0 m from separate areas near the centre of each subplot. Tillers were hand-clipped at ground level, manually separated into vegetative and reproductive tiller fractions, counted, placed in paper bags, dried for 4 days at 60°C in a forced-air oven, and weighed. Additionally, plants from two randomly selected 0.5 m by 0.5 m quadrats were hand-clipped at ground level from each subplot, dried at 60°C for 4 days, and weighed to obtain plant biomass (kg ha–1), which is the sum of spikelet dry weight, seed dry weight, and shoot dry weight per ha.
Plant height was measured from the soil surface to the tip of the highest panicle in 20 randomly selected reproductive tillers per subplot. Twenty-five panicles were randomly collected from each subplot and used to determine panicle length and the number of racemes, spikelets and seeds (per panicle). Unfilled spikelets were included in the count. After 5 months of storage, panicles were manually threshed to obtain the seeds for further analysis.
Seed quality components, floret site utilisation, seed yield and harvest index
Mature seeds were separated from spikelets and immature seeds by using a South Dakota blower. Clean seeds were examined to confirm their physical purity according to local methods (MAPA 2009). A seed quality analysis was undertaken by the Maringa State University Seed Laboratory. Pure seed is considered if it is either germinated or seed that looked normal. In this analysis, empty spikelets were not considered pure seeds. Hundred-seed weight was determined, converted to 1000-seed weight and adjusted to 12% moisture (Hare et al. (2005). All seed measurements relate to clean seeds.
Number of seeds per m2 was calculated using Eqn 2:

where reproductive tiller density is the number of reproductive tillers per m2.
Signal grass produces two florets per spikelet, one male sterile floret and one hermaphrodite fertile floret (Hopkinson et al. 1996). Thus, each spikelet can originate only one seed. The sterile floret was disregarded in this study, and the number of florets per panicle was considered equal to the number of spikelets per panicle at harvest. Floret site utilisation (FSU) was calculated using Eqns 3, 4 and 5:

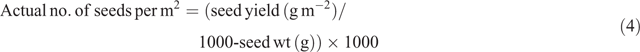

Potential floret density is defined as the sum of empty and filled florets at maturity.
Seed yield (kg ha–1) was not measured directly; rather, it was calculated from yield components according to the equation of Johnson et al. (2010) for Poa pratensis L.:

Harvest index was measured as the ratio of calculated seed yield to plant biomass (kg ha–1) and expressed as a percentage.
Flag leaf area and pre-harvest plant morphological characteristics
The flag leaf length and width (cm) of 15 reproductive tillers from each subplot were measured at harvest. Flag leaf area (cm2) was then calculated using the following equation of Bianco et al. (2000):
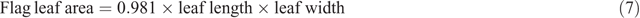
Most of the leaves of signal grass remain green until maturity. The following data were collected at harvest from 25 randomly chosen reproductive tillers from each subplot: number of green, fully expanded leaves (visible ligule) per tiller; number of senescent leaves per tiller; and number of nodes per tiller.
Data analyses
Analyses of variance were completed for the two consecutive crops using Statistica version 7.0 (StatSoft, Tulsa, OK, USA). Significant differences between water regimes were investigated by the F-test. When an interaction was not significant, the water-regime factor was removed from regression analysis. Linear and quadratic models were used to determine the responses of measured or calculated variables to N-application rates and plotted to identify trends. Selection of the best fit model to the data was identified by using the highest coefficient of determination (R2) criterion. In all analyses, differences were considered significant when P < 0.05.
Results
Weather conditions
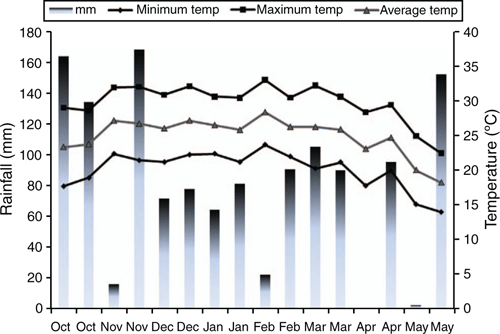
Rainfall and temperature during the growing season of each crop are shown in Fig. 1. Average rainfall was 688 mm for October–January, and 555 mm for February–May. Precipitation distribution and soil-water content were suitable for plant growth throughout the entire study period (October–May) and were not considered to have limited growth in any of the crop cycles (Fig. 2). The two crops grew vigorously and were not badly affected by insects. As a consequence, irrigation did not influence seed yield or most yield components, and no water regime × N rate interaction effect was observed on any of the parameters (P > 0.05). Therefore, in all data presentations, the values for N rates are means of the irrigated and non-irrigated treatments and values for water-regime treatments are means of all rates of N, and the discussion will focus on the effects of N rates.
|

|
Seed yield, number of seeds per area, harvest index, floret site utilisation, potential floret density, 1000-seed weight and physical purity
Seed yield increased linearly with N rate for the first crop (Fig. 3, Table 2) and quadratically for the second crop. A linear response model of seed number per area to N-fertiliser rate provided the best fit for the data from the first crop, whereas a curvilinear response model provided the best fit for the second crop (Fig. 4, Table 2). The relationship between N rate and potential floret density was best described by a quadratic equation for both the first and second crops (Fig. 4). FSU, 1000-seed weight and physical purity were not affected by N rates. Applications of N or irrigation did not consistently influence harvest index (P > 0.05). Harvest index ranged from 2.03% to 3.63% in the first crop and from 1.10% to 2.47% in the second crop.

|

|

|
Irrigation did not cause significant changes in seed yield or number of seeds per area (Table 3). FSU was significantly higher in irrigated plots. Potential floret density was significantly lower in irrigated than non-irrigated plots for the first crop. 1000-seed weight did not differ under different water regimes for the first crop but was slightly lower in irrigated plots for the second crop. Seed physical purity was slightly but significantly lower in non-irrigated than irrigated plots for both crops.

|
Yield components
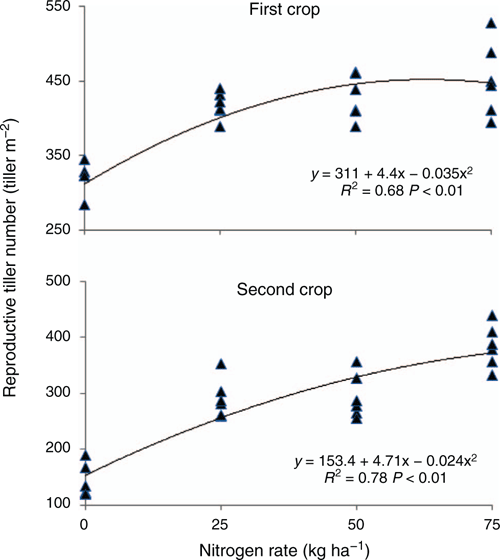
Reproductive tiller density was influenced by N-application rate and ranged from 306 to 453 tillers m–2 for the first crop and from 142 to 384 tillers m–2 for the second crop (Fig. 5, Table 4). The relationships between total tiller number and N rate for the first crop and percentage of reproductive tillers and N rate for the second crop were best described by quadratic equations (Table 5), whereas the response of total tiller number to N rate for the second crop was described by a linear equation. For the first crop, N rate did not affect the percentage of reproductive tillers (data did not fit linear or quadratic models). The response of plant biomass to N rates was best fitted to a quadratic equation for the first crop and a linear equation for the second crop (Fig. 6). Reproductive tiller weight was not affected (P > 0.05) by treatments. Reproductive tiller weight ranged from 1.9 to 2.6 g dry matter (DM) tiller–1 in the first crop and from 1.8 to 2.8 g DM tiller–1 in the second crop.
|

|

|

|
No differences in reproductive tiller density, total tiller number or percentage of reproductive tillers were observed between the two water regimes for either crop (Table 6). Water regime had no effect on plant biomass for the first crop; however, plant biomass was higher in non-irrigated plots for the second crop (Table 6).

|
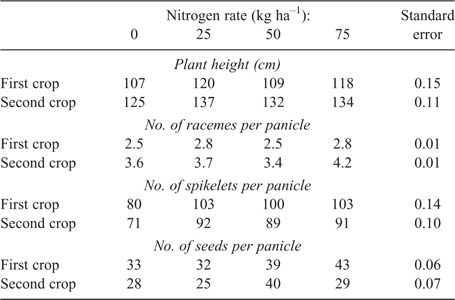
Plant height was not affected by N rate for either crop (Table 7) but was greater in plots without irrigation (Table 8). No significant (P > 0.05) differences were found among treatments for panicle length for either crop. Panicle lengths ranged from 19 to 21 cm in the first crop and from 15 to 17 cm in the second crop. There were no differences between the two water regimes for number of racemes per panicle for the first crop (Table 8); however, irrigation did reduce the number of racemes per panicle from 3.9 to 3.5 for the second crop. Nitrogen rate also had an effect on number of racemes per panicle for the second crop, but data did not fit linear or quadratic models. For the first crop, irrigation decreased the number of spikelets per panicle but did not influence the number of seeds per panicle (Table 8). For the second crop, the number of seeds per panicle was significantly less in non-irrigated plots, and the numbers of spikelets per panicle and seeds per panicle were affected by N rates, but data did not fit linear or quadratic models.
|

|
Crop phenology
Water regime and N rate had no significant (P > 0.05) effect on time to flowering, anthesis or harvest. Flowering, anthesis and harvest dates were, respectively, 11 December, 23 December 2009 and 15 January 2010 in the first crop, and 20 April, 4 May and 29 May 2010 in the second crop. Thus, crop duration, GDU, and number of days from cutting to flowering, cutting to anthesis, onset of flowering to harvest and anthesis to harvest were considered not to vary with irrigation or N treatments (P > 0.05). At the first crop and second crop, respectively, calculated GDU were 668 and 791 for the period cutting to onset of flowering, 809 and 910 for cutting to anthesis, 399 and 210 for onset of flowering to harvest, 258 and 91 for anthesis to harvest, and 1067 and 1001 for crop duration. At the first crop and second crop, numbers of days were 63 and 74 for the period cutting to onset of flowering, 88 and 75 for cutting to anthesis, 39 and 35 for onset of flowering to harvest, 25 and 23 for anthesis to harvest, and 113 and 98 for crop duration.
Flag leaf area and pre-harvest plant morphological characteristics
Nitrogen application and water regime had no significant (P > 0.05) effects on flag leaf area or pre-harvest plant morphological characteristics. Flag leaf area ranged from 1.9 to 3.9 cm2 in the first crop and from 0.6 to 3.3 cm2 in the second crop. At harvest, the numbers of green, fully expanded leaves per tiller, senescent leaves per tiller and nodes per tiller were, respectively, 7.7–10.7, 3.3–5.3 and 4.3–5.3 in the first crop, and 6.7–9.0, 4.7–6.3 and 6.8–8.8 in the second crop.
Discussion
Temperatures during the study period were typical of the growing season for signal grass in the region (October–May), with lower average temperatures during April–May. Rainfall was similar to long-term averages during the entire experimental period, although the second crop received 12% less rainfall (546 mm) than the first crop (619 mm) during the growing season. Little irrigation was applied because rainfall resulted in no obvious deficiencies in soil moisture. Prior to harvest, both plots showed some signs of lodging caused by strong winds and rain.
At first harvest, there was a linear increase in seed yield as N rates increased to 75 kg N ha–1. Compared with the value obtained without fertilisation (0 kg N ha–1), seed yield increased 34% with 25 kg N ha–1, 50% with 50 kg N ha–1 and 88% with 75 kg N ha–1. At second harvest, the highest seed yield values were obtained with 50 kg N ha–1 (293 kg/ha) and 75 kg N ha–1 (287 kg ha–1). The seed yield of the first harvest showed a greater response to N fertilisation than that of the second harvest. These results suggest that the seed yield of signal grass might respond to N rates >75 kg ha–1. Carmo et al. (1988) reported that 300 kg N ha–1 produced the highest seed yields of signal grass (229 kg ha–1), compared with the control (95.7 kg ha–1) and treatments of 35 kg N ha–1 (149.6 kg ha–1), 70 kg N ha–1 (199.5 kg ha–1) and 150 kg N ha–1 (204.2 kg ha–1).
The increase in seed yield in response to N may be related to the increase in plant biomass (Fig. 6), because both parameters are influenced by growth rate, photosynthetic rate and leaf area growth. Removal of crop residues after cutting (at the beginning of October and post-harvest in January) caused significant N exports, which might also have influenced seed yield responses to application of N, particularly in the second crop. The combination of residue removal and N application increased seed yield (Loeppky and Coulman 2002). However, there is evidence that these practices can lead to problems with grass persistence and lower seed yields in subsequent years. Importantly, there is a scarcity of studies evaluating the influence of soil N concentration and effective rooting-zone on seed yield of signal grass, especially considering that this species has a high rooting capacity.
Under our growing conditions, 75 kg N ha–1 applied to the first crop and 50 or 75 kg N ha–1 applied to the second crop appeared optimal for maximising seed yield under favourable rainfall. The N-fertiliser management recommendations may differ for other geographic regions. Most signal grass seed crops in Brazil are successfully produced from seeds that fall to the ground and are collected by using a highly specialised sweeper (Souza 2013). Extrapolation of our results to field conditions where seeds accumulate on the ground and are then harvested by machine-sweeping might be inappropriate.
To the best of our knowledge, there are only three reports dealing with the effects of N rate on seed yield of signal grass in Brazil (Carmo et al. 1988; Condé and Garcia 1988; Garcia et al. 1989). In one of these studies, seed yield was calculated on the basis of yield components (Garcia et al. 1989). In the other two reports, yield was evaluated by directly determining the weight of seeds per area (Carmo et al. 1988; Condé and Garcia 1988). In the present study, seed number per area and potential floret density were greater in first crop than second crop at all N rates (Table 2), possibly because of N removed by crop residues and/or weather conditions in second crop. In this study, the amounts of N removed as plant residues were higher than the amounts of N applied for the second crop (MW Canto, ED Pancera Junior, AB Barth, C Bremm, PU Vier, GM Almeida, ACS Costa, unpubl. data). A previous study of tall fescue (Schedonorus phoenix (Scop.) Holub) seed crops showed that plant responses to irrigation depend on the amount of rainfall during the growing season and can vary throughout the years (Huettig et al. 2013).
Seed yield reflected the changes in plant biomass (Fig. 6), but there was no difference in harvest index among N or irrigation treatments. A low harvest index is expected for undomesticated grass grown for leaf production (forage). Seeds represented 2.8% and 1.8% of the total plant biomass of the first and second crops, respectively. For cool-season grasses, the harvest index generally decreases as N rate increases (Chastain et al. 2014; Koeritz et al. 2015).
Floret site utilisation can be defined as the percentage of florets present at anthesis resulting in viable seeds (Elgersma 1985). In this study, we estimated the percentage of florets and potential floret density on the basis of the number of spikelets per panicle present at harvest, using a method similar to that described by Cookson et al. (2000). Seed number per unit area and potential floret density were significantly increased by N application for both crops because of the positive effect of N on reproductive tiller number (Fig. 5). However, field studies of winter cereals have shown that higher inflorescence densities adversely affect the number and weight of seeds per inflorescence owing to adjustment of these components in response to variable growing conditions. We did not use any method for quantifying the number of seeds that mature early or fall to the ground. Gobius et al. (2001), for instance, used nylon mesh bags to secure all seeds produced by signal grass and reported seed yield values of 81.4 kg ha–1 at 50 kg N ha–1, 100 kg ha–1 at 100 kg N ha–1 and 122.6 kg ha–1 at 200 kg N ha–1. However, those authors noted that, although nylon bags may be useful for collecting mature seeds that are easily shed from the panicle, this procedure can cause lodging, which favours fungal infections and may therefore influence seed-yield development.
Both crops produced seeds with low 1000-seed weight (Table 2) relative to previous studies (Carmo et al. 1988; Gobius et al. 2001). Hopkinson et al. (1996) considered that seed weights <450 mg 100 seeds–1 indicate low percentage of mature seeds in signal grass. Other studies have shown that application of N can influence mean seed weight in signal grass (Carmo et al. 1988) and creeping signal grass (Urochloa humidicola (Rendle) Morrone & Zuloaga) (Catuchi et al. 2017). However, little is known about the physiological basis for these observations.
In general, unfertilised plants (0 kg N ha–1) had lower reproductive tiller density (310 tillers m–2 for the first crop and 150 tillers m–2 for the second crop), produced fewer tillers and had visually lighter green leaves than plants receiving N fertiliser. These are all symptoms that can be attributed to N deficiency. Immature panicles were observed in all plots but were more abundant in plots treated with higher N rates. In our study, tillers bearing incompletely exserted panicles were not counted as reproductive tillers. Reproductive tiller density was higher in the first crop than in the second. These results suggest that weather conditions (Fig. 1) were more favourable for the production of panicles during the first crop cycle. These differences in panicle production may have resulted from differences in weather conditions and an interaction of soil fertility and weather conditions. Such observations underscore the importance of evaluating at least two harvests when studying signal grass. Furthermore, differences in reproductive tiller density between the two crops may be due to differences in harvest time or plant growth environments. Reproductive tiller density obtained in the present study when using 50 and 75 kg N ha–1 was lower than that reported by Condé and Garcia (1988) of 607 reproductive tillers m–2 when using 120 kg N ha–1 in signal grass grown without irrigation.
The highest total tiller numbers were in the 50 and 75 kg N ha–1 treatments in the first crop, and in the 75 kg N ha–1 treatment in the second crop. Reproductive tiller percentage generally followed trends observed for total tiller number.
Plant biomass increased with increasing N application through an increased number of tillers (Table 4), many of which were small, had low leaf area and did not produce panicles. Gobius et al. (2001) concluded that N rate (50–200 kg N ha–1) had no effect on plant biomass at harvest (9569–13 008 kg DM ha–1). In Australia, where signal grass is harvested by means of single-pass harvesters, high levels of biomass can reduce harvesting efficiency (Hopkinson and English 1982). In the second crop, we observed that irrigation led to a small decrease in plant biomass at harvest, which might have been caused by irrigation-induced N loss. Our results for reproductive tiller weight contrast with those of Canto et al. (2012, 2016), who found this variable to be significantly affected by N rate in Mombasa grass (Megathyrsus maximus (Jacq.) B.K.Simon & Jacobs cv. Mombaça).
Plant height was reduced by 15% (first crop) and 12% (second crop) with irrigation. This may be partly due to lodging. In seed crops of signal grass, N application changed panicle structure (1.6 racemes panicle–1 at 0 kg N ha–1 vs 2.1 racemes panicle–1 at 112 kg N ha–1) (Garcia et al. 1989), a finding that is in contrast with our results. We observed that numbers of spikelets per panicle and seeds per panicle were not affected by N application rates up to 75 kg N ha–1; however, N-application rate was lower than in the previous study. Hare et al. (2005), in a study of signal grass grown in north-eastern Thailand, recorded 40.0–43.3 spikelets raceme–1 in consecutive years.
Stür (1986) described that inflorescence exsertion, time of anthesis and seed maturation define the reproductive development of signal grass tillers. Our results indicate these indices of reproductive development were not influenced by differences in N availability in the soil. Growing degree-days were similar for both crops, but the first crop had fewer days to harvest than the second because of warmer growing conditions. The numbers of calendar days from onset of flowering to harvest and from anthesis to harvest were similar for both crops. However, the accumulated GDU for these periods were, respectively, 399 and 258 for the first crop and 210 and 91 for the second crop. This may be due to the cooler temperatures recorded from anthesis to harvest for the second crop (first half of April) (Fig. 1).
The consecutive crops did not differ in flag leaf area and pre-harvest plant morphological characteristics (number of expanded leaves, senescent leaves and nodes at harvest) regardless of N application, indicating that these indices cannot be manipulated through N application.
Conclusions
Two consecutive signal grass seed crops were possible under good rainfall conditions in an area of intermediate elevation in Brazil. Nitrogen application at 75 kg N ha–1 for the first crop and 50 or 70 kg N ha–1 for the second crop resulted in maximum seed yields. The effect of N rate on reproductive tiller density was the principal determinant of seed yield. Plant biomass was also strongly affected. The combined application of irrigation and high N rates had little additional effect on seed yield. These results suggest that in seasons of frequent rainfall in high-yield environments, application of irrigation has limited potential to increase yield of signal grass usefully over the range of N-application rates tested and is unlikely to be economically beneficial for seed production. Nitrogen application had little effect on number of seeds per panicle and no significant effect on flag leaf area or pre-harvest plant morphological characteristics. Nitrogen fertilisation also had no effect on the onset of flowering or anthesis. Our results provide information for the development of N-management strategies for signal grass seed production. Further study with N rates >75 kg ha–1 is needed to test for improved seed yield in signal grass plants. This research was conducted in one soil type and initial soil mineral-N concentrations were not estimated. Further research is needed to identify environmental and soil factors that affect the relationship between seed yield and yield components in signal grass grown under different N rates.
Conflicts of interest
The authors declare no conflicts of interest.
Funding statement
This research did not receive any specific funding.
References
Abel S, Boelt B (2018) Precipitation during anthesis reduces seed set in perennial ryegrass. Grass and Forage Science 73, 239–246.| Precipitation during anthesis reduces seed set in perennial ryegrass.Crossref | GoogleScholarGoogle Scholar |
Allen RGPereira LSRaes DSmith M (1998 ) ‘Crop evapotranspiration: guidelines for computing crop water requirements.’ FAO Irrigation and Drainage Paper 56. (Food and Agriculture Organization of the United Nations: Rome)
Alvares CA, Stape JL, Sentelhas PC, Moraes Gonçalves JL, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorologische Zeitschrif 22, 711–728.
| Köppen’s climate classification map for Brazil.Crossref | GoogleScholarGoogle Scholar |
Andrade RP, Thomas D, Fergunson JE (1983) Seed production of pasture species in a tropical savanna region of Brazil. II. Grasses. Tropical Grasslands 17, 59–64.
Bianco S, Brendolan RA, Alves PLCA, Pitelli RA (2000) Estimativa da área foliar de plantas daninhas: Brachiaria decumbens Stapf. e Brachiaria brizantha (Hochst.) Stapf. Planta Daninha 18, 79–83.
| Estimativa da área foliar de plantas daninhas: Brachiaria decumbens Stapf. e Brachiaria brizantha (Hochst.) Stapf.Crossref | GoogleScholarGoogle Scholar |
Canto MW, Barth Neto A, Pancera Júnior EJ, Gasparino E, Boleta VS (2012) Produção e qualidade de sementes do capim-mombaça em função da adubação nitrogenada. Bragantia 71, 430–437.
| Produção e qualidade de sementes do capim-mombaça em função da adubação nitrogenada.Crossref | GoogleScholarGoogle Scholar |
Canto MW, Almeida GM, da Costa ACS, Barth Neto A, Scaliante Júnior JR, Orlandini CF (2016) Seed production of ‘mombasa’ grass subjected to different closing cut dates and nitrogen rates. Pesquisa Agropecuária Brasileira 51, 766–775.
| Seed production of ‘mombasa’ grass subjected to different closing cut dates and nitrogen rates.Crossref | GoogleScholarGoogle Scholar |
Carmo MA, Nascimento Júnior D, Mantorani EA (1988) Efecto de la fertilización y la época de cosecha en la producción y la calidad de semillas de Brachiaria decumbens. Pasturas Tropicales 10, 19–22.
Castro RD, Vieira MGGC, Carvalho MLM (1994) Influência de métodos e épocas de colheita sobre a produção e qualidade de sementes de Brachiaria decumbens cv. Basilisk. Revista Brasileira de Sementes 16, 6–11.
| Influência de métodos e épocas de colheita sobre a produção e qualidade de sementes de Brachiaria decumbens cv. Basilisk.Crossref | GoogleScholarGoogle Scholar |
Catuchi TA, Soratto RP, Francisquini Júnior A, Aranda EA, Guidorizzi FVC, Tiritan CS (2017) Nitrogen management, nitrogen use efficiency, and seed yield and quality of creeping signalgrass. Crop Science 57, 2865–2874.
| Nitrogen management, nitrogen use efficiency, and seed yield and quality of creeping signalgrass.Crossref | GoogleScholarGoogle Scholar |
Chastain TG, Garbacik CJ, Young WC (2014) Spring-applied nitrogen and trinexapac-ethyl effects on seed yield in perennial ryegrass and tall fescue. Agronomy Journal 106, 628–633.
| Spring-applied nitrogen and trinexapac-ethyl effects on seed yield in perennial ryegrass and tall fescue.Crossref | GoogleScholarGoogle Scholar |
Condé AR, Garcia J (1988) Influência de níveis e épocas de aplicação de nitrogênio sobre o rendimento, qualidade, e componentes da produção de sementes do capim-braquiária. Revista Brasileira de Sementes 10, 63–71.
| Influência de níveis e épocas de aplicação de nitrogênio sobre o rendimento, qualidade, e componentes da produção de sementes do capim-braquiária.Crossref | GoogleScholarGoogle Scholar |
Cookson WR, Rowarth JS, Cameron KC (2000) The response of a perennial ryegrass (Lolium perenne L.) seed crop to nitrogen fertilizer application in the absence of moisture stress. Grass and Forage Science 55, 314–325.
| The response of a perennial ryegrass (Lolium perenne L.) seed crop to nitrogen fertilizer application in the absence of moisture stress.Crossref | GoogleScholarGoogle Scholar |
CQFS (2004) ‘Manual de adubação e calagem para os Estados do Rio Grande do Sul e de Santa Catarina.’ (Comissão de Química e Fertilidade do Solo RS/SC, Sociedade Brasileira de Ciência do Solo: Porto Alegre, RS, USA)
Elgersma E (1985) Floret site utilization in grasses: definitions, breeding perspectives and methodology. Journal of Applied Seed Production 3, 50–54.
Garcia R, Cani PC, Obeid JC, da Silva RF (1989) Influência do nitrogênio, cortes e épocas de colheita sobre a produção de sementes do capim-braquiária (Brachiaria decumbens Stapf.). Revista Brasileira de Zootecnia 18, 482–490.
Gobius NR, Phaikaev C, Pholsen P, Rodchompoo O, Susena W (2001) Seed yield and its components of Brachiaria decumbens cv. Basilisk, Digitaria milanjiana cv. Jarra and Andropogon gayanus cv. Kent in north-east Thailand under different rates of nitrogen application. Tropical Grasslands 35, 26–33.
Hare MD, Tatsapong P, Lunpha A, Wongpichet K (2005) Brachiaria species in north-east Thailand: dry matter yields and seed production. Tropical Grasslands 29, 99–106.
Hebblethwaite PD (1977) Irrigation and nitrogen studies in S.23 ryegrass grown for seed. 1. Growth, development, seed yield components and seed yield. The Journal of Agricultural Science 88, 605–614.
| Irrigation and nitrogen studies in S.23 ryegrass grown for seed. 1. Growth, development, seed yield components and seed yield.Crossref | GoogleScholarGoogle Scholar |
Hopkinson JM, English BH (1982) Harvest efficiency in seed crops of gatton panic (Panicum maximum) and signal grass (Brachiaria decumbens). Tropical Grasslands 16, 201–205.
Hopkinson JM, Souza FHD, Diulgheroff S, Ortiz A, Sanchez M (1996) Reproductive physiology, seed production, and seed quality of Brachiaria. In ‘Brachiaria: biology, agronomy, and improvement’. (Eds JW Miles, BL Maass, CB do Valle, V Kumble) pp. 124–140. (CIAT: Cali, Colombia)
Huettig KD, Chastain TG, Garbacick CJ, Young WC, Wysocki DJ (2013) Spring irrigation of tall fescue for seed production. Field Crops Research 144, 297–304.
| Spring irrigation of tall fescue for seed production.Crossref | GoogleScholarGoogle Scholar |
Jank L, Barrios SC, do Valle CB, Simeão RM, Alves GF (2014) The value of improved pastures to Brazilian beef production. Crop & Pasture Science 65, 1132–1137.
| The value of improved pastures to Brazilian beef production.Crossref | GoogleScholarGoogle Scholar |
Johnson RC, Johnson WJ, Bertoli FB, Golob CT (2010) Seed yield, development, and variation in diverse Poa pratensis accessions. Crop Science 50, 337–344.
| Seed yield, development, and variation in diverse Poa pratensis accessions.Crossref | GoogleScholarGoogle Scholar |
Koeritz EJ, Watkins E, Ehlke NJ (2015) Seeding rate, row spacing, and nitrogen rate effects on perennial ryegrass seed production. Crop Science 55, 2319–2333.
| Seeding rate, row spacing, and nitrogen rate effects on perennial ryegrass seed production.Crossref | GoogleScholarGoogle Scholar |
Lambert DA (1967) The effects of nitrogen and irrigation on Timothy (Phleum pratense) grown for production of seed. I. Vegetative growth. The Journal of Agricultural Science 69, 225–230.
| The effects of nitrogen and irrigation on Timothy (Phleum pratense) grown for production of seed. I. Vegetative growth.Crossref | GoogleScholarGoogle Scholar |
Loeppky HA, Coulman BE (2002) Crop residue removal and nitrogen fertilization affects seed production. Agronomy Journal 94, 450–454.
| Crop residue removal and nitrogen fertilization affects seed production.Crossref | GoogleScholarGoogle Scholar |
MAPA (2009) ‘Regras para análises de sementes.’ (Ministério da Agricultura Pecuária e Abastecimento, MAPA/SDA/ACS: Brasília, DF, Brazil)
Mielniczuk J, Ludwick AE, Bohnen H (1969) Recomendações de adubo e calcário para os solos e culturas do Rio Grande do Sul. Boletim Técnico 2. Faculdade de Agronomia, UFRGS, Porto Alegre, RS, Brazil.
Oram RN (1990) ‘Register of Australian herbage plant cultivars.’ (CSIRO: Melbourne)
Ricci GCL, Souza-Kaneshima AM, Felismino MF, Mendes-Bonato AB, Pagliarini MS, do Valle CB (2011) Chromossome numbers and meiotic analysis in the pre-breeding of Brachiaria decumbens (Poaceae). Journal of Genetics 90, 289–294.
| Chromossome numbers and meiotic analysis in the pre-breeding of Brachiaria decumbens (Poaceae).Crossref | GoogleScholarGoogle Scholar |
Santos HG, dos Jacomine PKT, dos Anjos LHC, de Oliveira VA, de Oliveira, JB, Coelho MR, Lumbreras JF, Cunha TJF (2006) ‘Sistema Brasileiro de Classificação de Solos.’ (Embrapa Solos: Rio de Janeiro)
Soil Survey Staff (2014) ‘Keys to Soil Taxonomy.’ (USDA Natural Resources Conservation Service: Washington, DC)
Souza FHD (2013) Produção de sementes para pastagens tropicais e subtropicais. In ‘Forragicultura. Ciência, tecnologia e gestão dos recursos forrageiros’. (Eds RA Reis, TF Bernardes, GR Siqueira) pp. 367–380. (Gráfica Multipress: Jaboticabal, SP, USA)
Stür WW (1986) Reproductive development of the apex of Brachiaria decumbens Stapf. Annals of Botany 58, 569–575.
| Reproductive development of the apex of Brachiaria decumbens Stapf.Crossref | GoogleScholarGoogle Scholar |
Stür WW, Humphreys LR (1988) Defoliation and burning effects on the tillering of Brachiara decumbens. Journal of Applied Ecology 25, 273–277.
| Defoliation and burning effects on the tillering of Brachiara decumbens.Crossref | GoogleScholarGoogle Scholar |
Thornthwaite CW, Mather JR (1955) ‘The water balance. Publications in climatology.’ (Drexel Institute of Technology: Philadelphia, PA, USA)
Villa Nova NA, Tonato F, Pedreira CGS, Medeiros HR (2007) Método alternativo para cálculo da temperatura base de gramíneas forrageiras. Ciência Rural 37, 545–549.
| Método alternativo para cálculo da temperatura base de gramíneas forrageiras.Crossref | GoogleScholarGoogle Scholar |
Wrege MS, Steimetz S, Reisser Júnior C, Almeida IR (2011) ‘Atlas climático da região sul do Brasil. Estados do Paraná, Santa Catarina e Rio Grande do Sul.’ (Embrapa: Pelotas, RS, USA)