

Evaluation of performance and stability of new sources for tolerance to post-emergence herbicides in lentil (Lens culinaris ssp. culinaris Medik.)
Rind Balech A * , Fouad Maalouf A * , Somanagouda B. Patil B , Kamal Hejjaoui B , Lynn Abou Khater A , Karthika Rajendran C , Diego Rubiales D and Shiv Kumar B
A * , Fouad Maalouf A * , Somanagouda B. Patil B , Kamal Hejjaoui B , Lynn Abou Khater A , Karthika Rajendran C , Diego Rubiales D and Shiv Kumar B
A International Center for Agricultural Research in the Dry Areas (ICARDA), Terbol, Lebanon.
B ICARDA, Rabat, Morocco.
C VIT School of Agricultural Innovations and Advanced Learning (VAIAL), Vellore Institute of Technology, Vellore 632014, Tamil Nadu, India.
D Institute for Sustainable Agriculture, Consejo Superior de Investigaciones Científicascic (CSIC), Córdoba, Spain.
Crop & Pasture Science 73(11) 1264-1278 https://doi.org/10.1071/CP21810
Submitted: 27 December 2021 Accepted: 4 April 2022 Published: 25 May 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution 4.0 International License (CC BY)
Abstract
Context: Yield losses in lentil (Lens culinaris ssp. culinaris Medik.) caused by weeds are estimated at 20–80%. In the absence of effective broad-spectrum herbicides, the menace of weed has emerged as a serious yield constraint in lentil.
Aims: Identification of tolerance to two broad-spectrum post-emergence herbicides, imazethapyr and metribuzin.
Methods: 221 lentil accessions were screened at the experimental research station of the International Center for Agricultural Research in the Dry Areas, Terbol (Lebanon) over four crop seasons (2014/15–2018/19). During the 2014/15 season, a preliminary screening experiment was conducted with 221 lentil accessions in an alpha lattice design with two replications, where accessions were treated at the pre-flowering stage at 150% of the recommended dose of imazethapyr (112.5 g active ingredient/ha) or metribuzin (315 g active ingredient/ha). Based on the preliminary results, 38 accessions were selected for further evaluation. In subsequent testing, the selected accessions were screened at 100% and 150% of the recommended doses of imazethapyr or metribuzin.
Key results: Herbicide treatments delayed the flowering time and maturation of tested lentil accessions. The plant height of selected tolerant accessions was reduced by 15% and 8% in the plots treated by imazethapyr and metribuzin, respectively. Reduction index (RIsy) was an effective tool to select for herbicide tolerance. Stability analysis indicated that two accessions (IG4400 and IG5722) performed under high rainfall environments while two other accessions (IG323 and IG4605) were adapted to low rainfall environments.
Conclusions: Four accessions (IG323, IG5722, IG4400, IG4605) were identified as independently tolerant to metribuzin and imazethapyr.
Implications: The identified herbicide-tolerant accessions can be used to develop herbicide-tolerant cultivars.
Key words: genetic variability, herbicide tolerance, imazethapyr, Lens culinaris, lentil, metribuzin, post-emergence herbicide, weed control.
Introduction
Lentil (Lens culinaris ssp. culinaris Medik.) is an important cool-season food legume crop worldwide. It has been incorporated for many decades in the culinary traditions of several countries especially in the Mediterranean, West Asia, Sub-Saharan Africa, and South Asia regions for being highly rich in protein (26%), prebiotics and micronutrients (Kumar et al. 2014). It is considered as a key option for sustainable intensification and diversification of cereal-based cropping systems due to its positive effect on cereal crops, adaptation to local conditions, ability to fix atmospheric nitrogen, and capability to reduce carbon footprints and water use (Joshi et al. 2017; Ouji and Mouelhi 2017). The latest triennium average suggests that the global production of lentil is 6.28 million tons from 5.40 million ha area with an average productivity of 1163 kg/ha (FAOSTAT 2021). The top ten lentil producing countries, namely Canada, India, Australia, Turkey, United States of America, Nepal, Kazakhstan, Bangladesh, Russian Federation, and China, collectively produced more than 92% of global output (FAOSTAT 2021). With rising interest in plant-based protein among health-conscious people, the gap between demand and supply of protein-rich lentils is increasing (Rubiales et al. 2021). To bridge this gap, there is a dire need to increase the productivity per unit area by adopting improved varieties and crop management practices.
Lentil is a poor weed competitor due to its shallow roots, poor early vigour, and slow vegetative growth. Its open growth habit easily stimulates the emergence and development of a plethora of weeds at early crop growth stages (Smitchger et al. 2012) especially in cool-season environments. The major annual broadleaf weeds competing with lentil are Centaurea balsamita, Ranunculus arvensis, Cephalaria syriaca, Lactuca serriola, Sonchus oleraceus, Sinapis arvensis and Setaria viridis (Wall and McMullan 1994; Erman et al. 2004; Merriam et al. 2021). The estimated yield losses caused by these annual weeds vary from 20 to 80% and may reach 100% in highly infested fields (Erman et al. 2004; Tepe et al. 2005) depending on the environmental conditions, and density and diversity of weed species (Yadav et al. 2007). On the other hand, the parasitic weeds affecting lentil production are broomrapes (mainly Orobanche crenata, and O. aegyptiaca) and dodders (mainly Cuscuta campestris), which can cause severe yield damages of up to 95%, especially in North Africa and Western and Central Asia (Rubiales and Fernández-Aparicio 2012).
Several weed management practices such as manual weeding, late sowing, higher plant densities, soil sterilisation, fertilisation and irrigation scheduling are suggested to control weeds in lentil fields (Brand et al. 2007). Still, most of these strategies turn out to be costly with low efficiency (Yenish 2007). The pre-emergence application of broad-spectrum herbicides such as metribuzin and imazethapyr is regarded as one of the most effective and economical methods to control weeds in lentil fields because of their ability to suppress weed growth and prevent yield losses (Elkoca et al. 2005). These pre-emergence herbicides control weeds at the early stage of crop growth, but weeds germinating after crop emergence become a menace to crop production (Gaur et al. 2013). Indeed, lentil cultivars are highly sensitive to these herbicides when used as post-emergence treatment. Metribuzin (C8H14N4OS), which belongs to the triazinone family is a pre- and post-emergence herbicide used to control both broadleaf and grass weeds in crops like soybeans (Soltani et al. 2005). Imazethapyr, an imidazolinone herbicide, can be used as a pre- and post-emergence herbicide to effectively control a wide range of weeds in legume crops, especially lentils (Hanson and Thill 2001; Teja et al. 2017). Therefore, selection of germplasm tolerant to post-emergence herbicides would be one of the major strategies to control weeds in lentils. Selection for combined resistance to both herbicides with different modes of action would allow herbicide rotation to retard the selection of herbicide resistance in weeds.
Several studies were conducted in lentils to identify tolerance to herbicides (Hanson and Thill 2001; Fedoruk et al. 2011; Yadav et al. 2013). In Australia, metribuzin tolerance in lentils has been identified (Mcmurray 2019) as a means of enabling the control of broadleaf weeds. In West Asia and North Africa there is a need to develop herbicide-tolerant lentils to control weeds such as Orobanche crenata and Cuscuta spp. and other annual broadleaves. Therefore, the present study was undertaken in the region to identify promising lentil accessions tolerant to both metribuzin and imazethapyr and to assess the efficiency of adopted herbicide tolerance scores. The second objective was to evaluate the performance and stability of selected accessions under diverse environments.
Materials and methods
Materials and experiments
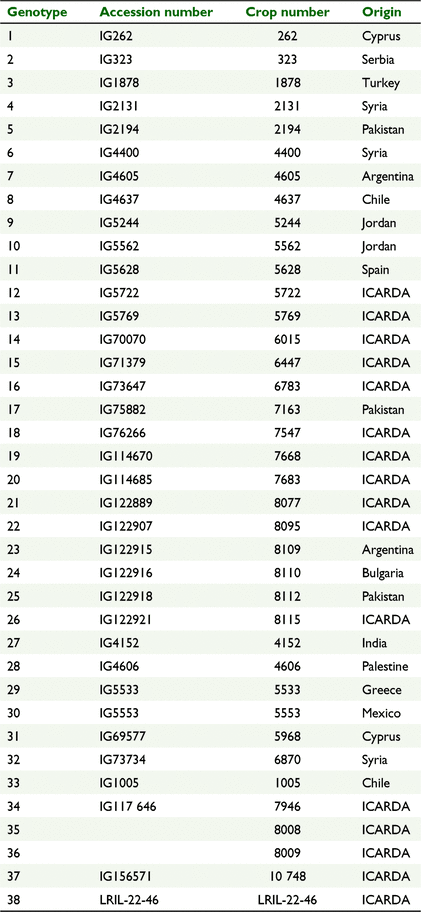
221 accessions of cultivated lentils, among them 105 landraces collected from 38 countries and 116 breeding lines developed at the International Center for Agricultural Research in the Dry Areas (ICARDA, Supplementary materials Table S1), were evaluated for their response to imazethapyr and metribuzin applied at the post-emergence stage under field conditions. The experiments were conducted at ICARDA experimental research station, Terbol, Lebanon (33.81°N, 35.98°E) at 890 metres above sea level. Terbol is characterised by cool winters and high rainfall as typical of its continental to semi-arid climate, with clay soil. The average precipitation during the crop seasons was 537 mm and the average temperature fluctuated between −1°C and 31°C (Fig. 1).

|
In this study, we conducted four experiments: two preliminary experiments (Experiment 1 and 2) and two validation experiments (Experiment 3 and 4), where imazethapyr and metribuzin were applied at the pre-flowering stage (5–6th node stage, 10–15 cm plant height). Standard agronomic practices were applied as following: soil fertilisation by adding nitrogen, phosphorus and potassium fertiliser (NPK 15-15-15) at 250 kg/ha; weeds were controlled by a pre-emergence application of pendimethalin at 1200 g active ingredient (a.i.)/ha followed by manual weeding to control weeds; sitona was controlled by spraying lambda-cyhalothrin at 40 g a.i./ha; thrips were controlled by a combination of thiamethoxam and acetamiprid at 200 g a.i./ha each and fungal diseases were controlled by a combination of azoxystrobin and difenoconazole respectively at 72.8 and 45.6 g a.i./ha.
The experiments were planted in late November and harvested in late May in rotation with cereals; durum wheat [Triticum turgidum L. subsp. durum (Desf.) van Slageren] or bread wheat (Triticum aestivum L.).
The plot size of the four experiments was a single row of 1 m length with 0.3 m distance between rows. The details of the experiments are as follows:
Experiment 1
Experiment 1 comprised 221 accessions and two repetitive tolerant checks (IG4400 and IG4605; previously identified at ICARDA in preliminary screening); which were conducted in a strip design with two treatments and two replicates during 2014/15. The two treatments imazethapyr at 112.5 g a.i./ha (1.5TI) and metribuzin at 315 g a.i./ha (1.5TM) were applied at 150% of the recommended dose as per the label recommendation of the two herbicides metribuzin (Sencor: Bayer) and imazethapyr (Pursuit: BASF).
Experiment 2
Experiment 2 comprised 38 accessions selected from Experiment 1, including 34 tolerant and four susceptible accessions (Table 1); it was conducted in an alpha design with two replicates during 2015/16. Two herbicide treatments, imazethapyr 112.5 g a.i./ha (1.5TI) and metribuzin 315 g a.i./ha (1.5TM), were applied at 150% of the recommended doses.
|
Experiment 3
The same set of 38 accessions (Table 1) was evaluated for their performance against imazethapyr (1TI: 75 g a.i./ha) and metribuzin (1TM: 210 g a.i./ha) and compared with control (C) during 2016/17.
Experiment 4
The same set of 38 accessions (Table 1) was again assessed for their performance against five treatments, namely imazethapyr (1TI: 75 g a.i./ha) and (1.5TI: 112.5 g a.i./ha), metribuzin (1TM: 210 g a.i./ha) and (1.5TM: 315 g a.i./ha) and control (C) during 2018/19.
Recorded observations
Based on the lentil ontology (Kumar and Rajendran 2016), the following observations were recorded:
Herbicide damage score (HDS)
HDS was recorded following a 1–5 scale after 2 weeks (HDS1) and 5 weeks (HDS2) of herbicide application to assess the ability of accessions to recover from the herbicide treatments (Gaur et al. 2013). The detailed description of HDS is as following:
-
HDS = 1: No damage occurred, no symptoms of phytotoxicity shown and the plants are in excellent shape with a similar appearance to the control plants.
-
HDS = 2: Slight damage observed by a light inhibition of growth with a marginal yellowing of some leaves; these plants continue normal vegetative growth to flowering and podding stages.
-
HDS = 3: Moderately damaged accessions showing a clear difference with the untreated plot by the appearance of necrosis on leaves and a lower vegetative growth with a clear deformation on the newly formed apical leaves and a rate of mortality below 25%. Plants at this stage were able to proceed to the flowering and podding stages.
-
HDS = 4: severely damaged accessions where plants have a poor vegetative growth caused by a severe chlorosis, narrowing and burning of leaves. These plants stop the development of new leaves totally and the mortality rate varies between 25% and 75%. The flowering stage is heavily affected since the flowers are burned.
-
HDS = 5: Severe damage of the crop with mortality above 75% per plot.
Crop phenology
Observations were taken on days to 50% flowering (DFLR) and 95% maturity (DMAT) on a plot basis.
Agronomic and yield traits
At maturity, three plants were taken randomly to record observations on plant height (PLHT), biological yield/plant (BY), number of pods/plant (NPP), number of seeds/plant (NSP), and seed yield/plant (SY) and the average of three plants was used for statistical analysis.
Reduction indexes
Reduction index (RItrait) was estimated to measure the performance of selected tolerant accessions, as follows (Sharma et al. 2018):

where  is the trait value of an evaluated accession under herbicide treatments, and
is the trait value of an evaluated accession under herbicide treatments, and 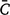 is the value of the same accession under controlled conditions without any herbicide treatments. This reduction index was calculated for plant height (PLHT), biological yield per plant (BY) and seed yield per plant (SY).
is the value of the same accession under controlled conditions without any herbicide treatments. This reduction index was calculated for plant height (PLHT), biological yield per plant (BY) and seed yield per plant (SY).
Statistical analysis
The statistical row-column model was applied to detect differences among accessions (A), herbicide treatments (T) and their interaction (A×T) for phenological and agronomic traits using the Genstat statistical software (Goedhart and Thissen 2010). Differences in the effects of accessions and herbicide treatments were assessed using P values. The best-unbiased values of each accession and treatment were estimated by applying thestatistical software. Ordinal regression analysis was performed between HDS1 and HDS2 and the reduction index to assess the efficiency of the herbicide tolerance score.
Multiple experiments analysis over the years was conducted using the method of residual maximum likelihood (REML) where A, T and A × T were fitted in the fixed model while years, replicates and blocks were fitted in the random model. In addition, each herbicide treatment in every season was considered an independent environment to assess the stability of 38 accessions selected for herbicide tolerance in the preliminary studies. Genetics, genetics × environment (GGE) biplot of multi-environment trial (MET) analysis of these accessions were conducted using the best linear unbiased predictions (BLUPs) to evaluate their replicability over the seasons under diverse herbicides. A line was drawn to connect each treatment to the biplot origin to visualise the relationship between the herbicide treatments, called vectors. The angle between two vectors was used to approximate the correlation between the two herbicide treatments (Yan and Tinker 2006; Kaya and Turkoz 2016). The smaller the angle between two vectors, the higher is the correlation between the two environments. Finally, the biplot showed the mega-environments by drawing an ellipse around similar environments which share the same sector.
Results
Herbicide damage score
The HDS1 score of lentil accessions ranged between 2 and 5 for imazethapyr at 112.5 g a.i./ha (1.5TI) during 2014/15, showing wide range of variation among herbicide tolerance. Among 221 accessions tested, 21 accessions scored two with slight damage on leaves with marginal yellowness, 123 accessions scored three with moderate damage with leaf necrosis, 68 accessions with score four were severely damaged with 25–75% mortality, and nine accessions scored five with total mortality. The HDS2 score, taken after 5 weeks of herbicide treatment, indicated accentuated damage in all accessions. Based on the HDS2 score, ten accessions with marginal leaf yellowness recorded scores of two, 92 with moderate levels of damage scored three, 107 accessions with severe damage scored four and finally, 12 accessions with total crop failure scored five. For metribuzin at 315 g a.i./ha treatment (1.5TM), HDS1 showed wide variation with seven accessions scoring two with minimum damage (marginal leaf burning), 85 scoring three with moderate damage (leaf necrosis and lower vegetative growth), 127 scoring four with high damage (severe leaf burning) and two accessions scoring five with total mortality for more than 75% of plants in the treated plot. The HDS2 score, taken after 5 weeks of herbicide treatment, showed recovery from the herbicide damage with the formation of new leaves. The HDS2 score showed that only one accession scored one with no visible damage, 31 scored two with slight damage, 114 scored three with moderate damage, 73 scored four with a mortality rate between 25 and 75% and two accessions scored five with a mortality rate above 75%.
Based on the herbicide damage score in the preliminary screening trials, 38 accessions were selected for further evaluation to confirm their tolerance. Validation trials conducted during 2018/19 (Experiment 4) showed that lentil accessions recovered from the herbicide damage within 5 weeks after the application of imazethapyr at 75 g a.i./ha whereas the damage was accentuated when treated with imazethapyr at 112.5 g a.i./ha (1.5TI) (Fig. 2). On the other hand, the results of Experiment 3 conducted during 2016/17 showed that the damage was accentuated 5 weeks after treatment with imazethapyr at 75 g a.i./ha. For metribuzin, lentil accessions showed recovery from the herbicide damage after 5 weeks of metribuzin treatments during 2016/17 and 2018/19 (Fig. 2).

|
Crop phenology
Combined analysis of variance showed that for days to 50% flowering (DFLR) and 95% maturity (DMAT) data, P < 0.001 among accessions (A), herbicide treatment (T) and years (Y) was detected (Table 2). Analysis of variance for these traits at each experiment among accessions and herbicide treatments also showed that P was less than 0.001 over the years except for DFLR among herbicide treatments (T) during 2015/16 (Experiment 2) (Table 3). Moreover, P < 0.001 obtained for the A × T interactions over the years except for DFLR during 2014/15 (Experiment 1).

|

|
It was observed that the pre-flowering phase was prolonged in plots treated with imazethapyr than in metribuzin. Experiment 1 showed that flowering date in lentil accessions was delayed by an average of 4.9 days in the imazethapyr treatment at 112.5 g a.i./ha (1.5TI), when compared to metribuzin treatment at 315 g a.i./ha (1.5TM). In addition, during 2016/17, the average flowering time in lentil accessions was delayed by 7.6 days for imazethapyr at 75 g a.i./ha treatment. In contrast, for metribuzin at 210 g a.i./ha, there was no delay when compared with the control (Table 4). The same results were observed in the 2018/19 season.

|
The number of days to maturity of lentil accessions was prolonged in plots treated with imazethapyr (1TI), but not in plots treated with metribuzin (1TM) during 2016/17 and 2018/19. Moreover, when increasing the dose of both herbicides, no delays in either flowering and maturity time were observed during 2018/19 (Table 4).
Agronomic traits
Combined analysis of variance showed that for plant height, P was less than 0.001 for plant height, among accessions (A) and herbicide treatments (T) (Table 2). In each experiment, we observed that P was less than 0.001 for plant height among lentil accessions (A) and herbicide treatments (T) over the years except for the herbicide treatments (T) during 2014/15 and 2015/16 (Table 3). The average plant height of lentil accessions was shorter under herbicide treatments than in untreated control plots except when compared with metribuzin treatments (1TM and 1.5TM) in 2018/19 (Table 4).
Yield attributes
Combined analysis of variance showed that for biological yield (BY), P < 0.01 was observed among accessions (A) and herbicide treatments (T) (Table 2). At the level of each experiment, we observed that P was inferior than 0.001 for biological yield among lentil accessions (A) and herbicide treatments (T) except for treatments (T) during 2014/15 (Table 3).
During 2016/17, the average biological yield of the untreated control plots (control) was higher than the average biological yield of imazethapyr at 75 g a.i./ha (1TI) and metribuzin at 210 g a.i./ha (1TM). However, during 2018/19 (Experiment 4), the average biological yield of the untreated control plots was higher than the average biological yield of lentils treated with imazethapyr with any dose, but showed no difference with either dosage of metribuzin at 315 g a.i./ha (1.5TM) (Table 4).
Combined analysis of variance showed that P was less than 0.01 for seed yield (SY), number of pods per plant (NPP) and number of seeds per plant (NSP) among accessions (A) and herbicide treatments (T) (Table 2). In each experiment, P was inferior than 0.001 for seed yield, number of pods per plant and number of seeds per plantamong lentil accessions (A) and herbicide treatments (T) over the years, except for the herbicide treatments (T) of number of pods per plant and number of seeds per plant during 2016/17 and of seed yield and number of pods per plant during 2018/19 (Table 3). The Accessions × Treatment (A × T) interaction showed P value above 0.05 indicating that there is no interaction between accessions and herbicide treatments applied (Table 3). All yield attributes (SY, NPP and NSP) were higher in untreated control plots than the herbicide-treated plots (Table 4). However, increased dosages of herbicides did not further affect the seed yield, number of pods per plant and number of seeds per plant as exemplified in Experiment 4 during 2018/19 (Table 4).
Reduction index
Combined analysis of variance showed that P was less than 0.01 for reduction index of plant height (RIPLHT), biological yield per plant (RIBY) and yield per plant (RISY), among accessions (A) and herbicide treatments (T) (Table 2). At the level of each experiment, the analysis of variance for the reduction index of plant height (RIPLHT), biological yield per plant (RIBY) and yield per plant (RISY), P < 0.005 among accessions (A), herbicide treatments (T) and A × T interaction during all the cropping seasons except among the herbicide treatments (T) of RIPLHT during 2016/17 (Experiment 3) and among the herbicide treatments (T) and A × T interaction of RIBY during 2018/19 (Experiment 4) (Table 3).
The ordinal regression analysis of Experiment 3 conducted during 2016/17 showed that for the reduction index of plant height (RIPLHT), biological yield (RIBY) and seed yield (RISY), P was less then 0.05 with the herbicide damage score (HDS2) except for RISY with imazethapyr at 112.5 g a.i./ha (1.5TI) and for RIBY and RISY with metribuzin at 315 g a.i./ha (1.5TM) (Table 5). The average RIPLHT, RIBY and RISY increased from 27.8 to 39.8%, from 46.5 to 98.3% and from 78.6 to 99.6% when the herbicide damage score (HDS2) increased after the treatment of imazethapyr with 75 g a.i./ha during 2016/17. The same observation was made for metribuzin. The results of Experiment 4 conducted during 2018/19 indicated no ordinal regression between HDS2 score and reduction indexes for plant height, biological yield (BY) and seed yield (Table 5).

|
Selection of tolerant accessions
Correlation analysis between HDS2 and reduction index of plant height (RIPLHT), biological yield (RIBY) and seed yield (RISY) showed no correlation between HDS2 of four herbicide treatments (1TI, 1.5TI, 1TM and 1.5TM) and the estimated reduction indexes of three traits (Table 5). The HDS score was helpful in preliminary screening; however, there is a need to rely on the yield and reduction index for validation. In addition, some accessions showed phenological recovery and vegetative growth after treatment, but none was capable of full recovery because of unfavourable environmental conditions.
Based on the estimated RISY, four accessions confirmed their tolerance to both herbicides (IG323, IG5722, IG4400, IG4605) (Table 6). The selected accessions (IG323, IG5722, IG4605, IG4400) did not show a delay in flowering and maturity under different metribuzin treatments (1TM: 210 g a.i./ha and 1.5TM: 315 g a.i./ha). Interestingly, IG4605 flowered earlier than the control even under imazethapyr treatments (1TI) and (1.5TI). Overall, the phenology of tolerant accessions was not affected when treated with any of the herbicides.

|
Replicability analysis
The tested accessions under validation trials performed differently among seasons and herbicide treatments as P was less than 0.001 of Accession × Year × Treatment, and Accession × Year and Treatment × Year (Table 2). GGE biplot analysis was conducted for grain yield to assess the replicability of lentil accessions selected previously in the preliminary studies of herbicide tolerance. The biplot accounted for >70% of the variation in grain yield in relation to genotypes and their interactions with environment (Fig. 3). The biplot was divided into eight sectors and four mega-environments. Two mega-environments were represented by one season-treatment each: Mega-environment 1 (E1: season 2016/17 of imazethapyr at 75 g a.i./ha) and Mega-environment 2 (E5: season 2018/19 of metribuzin at 210 g a.i./ha) and the other two representing more than one season-treatment and Mega-environment 3 (E2: season 2016/17 of metribuzin at 210 g a.i./ha and E3: season 2016/17 control untreated with herbicide) and Mega-environment 4 (E4: season 2018/19 of imazethapyr at 75 g a.i./ha, E6: 2018/19 season of metribuzin at 315 g a.i./ha and E7: 2018/19 season control untreated with herbicide). The two major mega-environments (3 and 4) aligned with the weather conditions during 2016/17 and 2018/19, respectively (Fig. 3).

|
Four lentil accessions, namely IG5628, IG5769, IG114670 and IG4152 were located close to the origin, indicating their wide adaptation and highest stability with similar performance under normal growth conditions without herbicide treatments and with either metribuzin and imazethapyr in Mega-environment 1; (E1); and Mega-environment 3; (E2, E3) (Fig. 3). This observation shows the replicability of these accessions in seasons with low rainfall conditions, represented by the environmental conditions of 2016/17, highlighted by a total precipitation equivalent to 458 mm (Fig. 1a).
Five accessions, namely IG2194, IG4637, IG73647, IG1005 and ILL8008 were located in Mega-environment 4 with no herbicide treatments (control) and were adapted to environments treated by imazethapyr and metribuzin, indicated their wide adaptability with similar performance under the different herbicide treatments. E4, E6 and E7 are 2018/19 environments highlighted by a total precipitation equivalent to 709 mm (Fig. 1a). This shows that the environmental conditions had a higher effect on the accessions than the treatments applied. Fig. 3 also shows that IG4400 is the most tolerant under Mega-environment 4, thus under high rainfall conditions.
The four selected tolerant genotypes showed adaptability to different seasons and herbicide treatments. IG 4400 was located in Mega-environment 4 with no herbicide treatments (control) and was adapted to environments treated by imazethapyr and metribuzin, indicated its adaptability to high rainfall conditions. IG4605 was located in Mega-environment 3 (E2, E3) with no herbicide treatments (control) and was adapted to environments treated by imazethapyr and metribuzin, indicating their adaptability to low rainfall conditions. IG323 is not stable and not adapted to any mega-environment. Still, the yield data (SY) showed that IG323 had low yield under low rainfall conditions and high yield under high rainfall conditions. IG 5722 is located near the centre, and in the sector of E4 and E5 (E4: season 2018/19 of imazethapyr at 75 g a.i./ha, E5: season 2018/19 of metribuzin at 210 g a.i./ha). Thus, it is a tolerant accession under a high rainfall environment.
Discussion
Weeds are considered a major constraint affecting the production of lentil and the application of herbicides is considered one of the most efficient techniques to control weeds and avoid yield losses in many crops (Garcia De Arevalo et al. 1992). In lentil, pre-emergence herbicides are available for use to control weeds efficiently at the early growing stage but not in the North and East Africa regions. Therefore, the selection of lentil accessions tolerant to herbicides is essential to integrate lentil into the cropping system. Our results showed a wide range of genetic variability for herbicide tolerance in lentils which allow introgression of the tolerance to widely adapted cultivars. This observation was reported in earlier studies in lentil (Sharma et al. 2016), chickpea (Cicer arietinum L.) (Taran et al. 2010; Gaur et al. 2013; Chaturvedi et al. 2014), faba bean (Vicia faba L.) (Abou-Khater et al. 2021), and field pea (Pisum sativum L.) (Hanson and Thill 2001).
Crop response after herbicide treatments
The herbicide damage in legume crops can be accentuated in susceptible accessions while tolerant ones can recover after being affected in the first 2 weeks of treatment. This has been reported in various studies in lentil (Sharma et al. 2018), chickpea (Goud et al. 2013), soybean (Belfry et al. 2015) and faba bean (Abou-Khater et al. 2021). Our experiments showed similar results as, after 5 weeks of herbicide treatments with imazethapyr, the tolerant accessions recovered from the damage while susceptible ones showed accentuated damage. This is expected since lentil is sensitive at less than 5% of the recommended dose of Imidazolinone herbicides (Stork 1995). On the contrary, tolerant accessions could recover from the damage after five weeks even when an increased dose of metribuzin (1.5 ×) was applied in our experiments. The recovery of vegetative growth can be attributed to the metabolic degradative ability of each accession to metabolise the herbicidal toxic components and detoxify the plants (Shoup et al. 2003).
Herbicide damage score was found to be associated with reduction indexes in the experiment conducted in 2017/18, which was a moderately rainy growing season. This indicates the efficiency of the HDS for screening for herbicide tolerance in lentils and aligns with the results obtained in faba bean (Abou-Khater et al. 2021). However, this observation was not replicated during 2018/19 in our experiments; this is because of heavy rain during the winter season and abnormally high temperatures during pod set (April) and grain filling (May), which affected the growth habit of the plants. For that reason, the reduction index did not correlate with HDS in 2018/19.
In conclusion, the herbicide damage scores (HDS) give a general overview of the reaction of genotypes against a tested herbicide. In the case of screening a large number of genotypes, the HDS can indicate highly susceptible lines, which can be excluded from further testing. In validation trials with a limited number of test entries, reduction indexes might be more reliable for validating the reaction of tolerant lines. Therefore, the validation of tolerant accessions in our study was based on the reduction index for yield (RISY). The same selection method was also adopted in faba bean (Abou-Khater et al. 2021). However, Taran et al. (2013), Gaur et al. (2013) in chickpea and Sharma et al. (2018) in lentils found a high correlation between herbicide tolerance and morpho-physiological traits, yield and yield components. Therefore, they relied on the herbicide damage score to rank the tolerance of the tested genotypes. Our study shows that visual assessment of the plant through the HDS after herbicide application is proved to be a rapid and easy method for identifying tolerance to imazethapyr and metribuzin at preliminary trial but not sufficient and reliable in validating highly tolerant lines.
Effect of herbicides on crop phenology
In all the four experiments conducted, flowering of lentil accessions was delayed under herbicide treatments. Consequently, the pre-flowering phase in treated plots was prolonged. Similar results were also obtained in previous studies in lentils (Sharma et al. 2016, 2018), chickpea (Taran et al. 2010; Gaur et al. 2013; Chaturvedi et al. 2014) and faba bean (Abou-Khater et al. 2021). This might be explained by the fact that the recovery of accessions from herbicide treatments involved secondary plant growth with delayed flowering time and pod setting.
Effect of herbicides on agronomic and yield traits
Application of imazethapyr and metribuzin reduced plant height and biological yield. Similar observations were made earlier on lentil (Sharma et al. 2016, 2018), faba bean (Abou-Khater et al. 2021) and chickpea (Taran et al. 2010; Goud et al. 2013). In addition, yield components reduced in all the conducted experiments. These observations are in agreement with the results of Gaur et al. (2013) and Goud et al. (2013). Similarly, in lentils, Sharma et al. (2016, 2018) reported a reduction in yield and yield attributes in the herbicide-treated accessions when compared to untreated control.
Replicability
The GGE biplot permits us to determine specific and wide adaptability of different accessions to mega-environments. Our study indicated two major mega-environments as each of them consistently represents various trials (Yan et al. 2007), where four selected accessions (IG323, IG4400, IG4605 and IG 5722) showed stability to environments. However, two accessions IG4400 and IG5722 were found to be specifically adapted to herbicide treatments under high rainfall conditions and the other two (IG323 and IG4605) were found in highly specific environments characterised by herbicide treatments and low rainfall. This is the first report of the stability of these herbicide-tolerant accessions. The accessions were screened at Terbol station assuming that this station represents ideal environments for screening for herbicide tolerance as observed in faba bean (Abou-Khater et al. 2021). Further investigation is also ongoing to validate the observation made on faba bean by Abou-Khater et al. (2021) and to identify suitable environments for screening for herbicide tolerance in lentil. The selected accessions should be crossed to widen their adaptability to different environments.
Conclusion
This study suggests the presence of tolerance to post-emergence herbicide treatments in lentils. An herbicide-tolerant variety is an integral part of the integrated weed management package for the most effective and economical approach to for weed management. A large genetic variability for herbicide tolerance was observed in lentil germplasm in our study. Sixteen tolerant genotypes of lentil without any effect of herbicide treatment on phenology were identified. Further investigation is required to study the genetics of herbicide tolerance in these genotypes before their efficient use as a donor in the lentil breeding program.
Supplementary material
Supplementary material is available online.
Data availability
The data is available and can be shared by corresponding authors upon request.
Conflicts of interest
The authors declare that there is no conflicts of interest.
Declaration of funding
This research was funded by the CGIAR Research Program (CRP) grain legume program from 2012–16 and by the Arab Fund for Economic and Social Development (AFESD) for the implementation of decentralised center around in five Arab countries.
Author contributions
Rind Balech conducted the field experiments at Terbol station during 2014/15, 2015/16, 2016/17 and 2018/19 and wrote the first draft. Karthika Rajendran and Somanagouda B. Patil reviewed the paper. Kamal Hejjaoui and Lynn Abou Khater contributed to the implementation of the trials and to data management. Diego Rubiales is Rind’s PhD director; he revised the paper. Shiv Kumar contributed to fund raising and reviewing the paper. Fouad Maalouf is Rind’s PhD second supervisor; he supervised the implementation of all experiments and made a major contribution to the paper writing and editing.
Acknowledgements
We would like to take this opportunity to thank Marie Wehbe, Ghazi Khatib for the hard work that has been done through the whole cycles of experimentations that were conducted.
References
Abou-Khater L, Maalouf F, Patil SB, Balech R, Rubiales D, Kumar S (2021) Identification of tolerance to metribuzin and imazethapyr herbicides in faba bean. Crop Science 61, 2593–2611.| Identification of tolerance to metribuzin and imazethapyr herbicides in faba bean.Crossref | GoogleScholarGoogle Scholar |
Belfry KD, McNaughton KE, Sikkema PH (2015) Weed control in soybean using pyroxasulfone and sulfentrazone. Canadian Journal of Plant Science 95, 1199–1204.
| Weed control in soybean using pyroxasulfone and sulfentrazone.Crossref | GoogleScholarGoogle Scholar |
Brand J, Yaduraju NT, Shivakumar BG, Mcmurray L (2007) Weed management. In ‘Lentil: an ancient crop for modern times’. (Eds S Yadav, D McNeil, P Stevenson) pp. 159–172. (Springer: Dordrecht, Netherlands)
Chaturvedi SK, Muraleedhar A, Gaur PM, Neelu M, Singh K, Nadarajan N (2014) Genetic variations for herbicide tolerance (Imazethapyr) in chickpea (Cicer arietinum). Indian Journal of Agricultural Sciences 84, 968–970.
Elkoca E, Kantar F, Zengin H (2005) Weed control in lentil (Lens culinaris) in eastern Turkey. New Zealand Journal of Crop and Horticultural Science 33, 223–231.
| Weed control in lentil (Lens culinaris) in eastern Turkey.Crossref | GoogleScholarGoogle Scholar |
Erman M, Tepe I, Yazlik A, Levent R, Ipek K (2004) Effect of weed control treatments on weeds, seed yield, yield components and nodulation in winter lentil. Weed Research 44, 305–312.
| Effect of weed control treatments on weeds, seed yield, yield components and nodulation in winter lentil.Crossref | GoogleScholarGoogle Scholar |
Fedoruk LK, Johnson EN, Shirtliffe SJ (2011) The critical period of weed control for lentil in western Canada. Weed Science 59, 517–526.
| The critical period of weed control for lentil in western Canada.Crossref | GoogleScholarGoogle Scholar |
FAOSTAT (2021) Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. FAO, Rome, Italy. Available at https://www.fao.org/faostat/en/#data
Garcia De Arevalo RC, Lusarreta CA, Neyra CB, Sanchez MA, Algarra PJH (1992) Chemical control of annual weeds in field beans (Vicia faba) in central Spain. Weed Science 40, 96–100.
| Chemical control of annual weeds in field beans (Vicia faba) in central Spain.Crossref | GoogleScholarGoogle Scholar |
Gaur P, Jukanti A, Samineni S, Chaturvedi S, Singh S, Tripathi S, Singh I, Singh G, Das T, Aski M, Mishra N, Nadarajan N, Gowda C (2013) Large genetic variability in chickpea for tolerance to herbicides imazethapyr and metribuzin. Agronomy 3, 524–536.
| Large genetic variability in chickpea for tolerance to herbicides imazethapyr and metribuzin.Crossref | GoogleScholarGoogle Scholar |
Goedhart PW, Thissen JTNM (2010) ‘Biometris GenStat procedure library manual.’ 13th edn. (No. 18.08. 10). (Wageningen Universiteit: Wageningen, Netherlands)
Goud VV, Murade NB, Khakre MS, Patil AAN (2013) Efficacy of imazethapyr and quizalofop-ethil herbicides on growth and yield of chickpea. The Bioscan 8, 1015–1018.
Hanson BD, Thill DC (2001) Effects of imazethapyr and pendimethalin on lentil (Lens culinaris), pea (Pisum sativum), and a subsequent winter wheat (Triticum aestivum) crop. Weed Technology 15, 190–194.
| Effects of imazethapyr and pendimethalin on lentil (Lens culinaris), pea (Pisum sativum), and a subsequent winter wheat (Triticum aestivum) crop.Crossref | GoogleScholarGoogle Scholar |
Joshi M, Timilsena Y, Adhikari B (2017) Global production, processing and utilization of lentil: a review. Journal of Integrative Agriculture 16, 2898–2913.
| Global production, processing and utilization of lentil: a review.Crossref | GoogleScholarGoogle Scholar |
Kaya Y, Turkoz M (2016) Evaluation of genotype by environment interaction for grain yield in durum wheat using non-parametric stability statistics. Turkish Journal of Field Crops 21, 51–59.
| Evaluation of genotype by environment interaction for grain yield in durum wheat using non-parametric stability statistics.Crossref | GoogleScholarGoogle Scholar |
Kumar S, Rajendran K (2016) Lentil ontology. Available at https://cropontology.org/term/CO_339:ROOT
Kumar S, Hamwieh A, Manickavelu A, Kumar J, Sharma TR, Baum M (2014) Advances in lentil genomics. In ‘Legumes in omics era’. (Eds S Gupta, N Nadarajan, DS Gupta) pp. 111-130. (Springer Science+Business Media: New York, NY, USA)
| Crossref |
Mcmurray LS (2019) Improving metribuzin tolerance in lentil (Lens culinaris). PhD Thesis, University of Adelaide, School of Agriculture, Food and Wine, Adelaide, SA, Australia.
Merriam A, Malone J, Gill G, Preston C (2021) Can rotations improve management of herbicide-resistant annual sowthistle (Sonchus oleraceus) and prickly lettuce (Lactuca serriola) in lentil production systems of southern Australia? Weed Technology 35, 532–538.
| Can rotations improve management of herbicide-resistant annual sowthistle (Sonchus oleraceus) and prickly lettuce (Lactuca serriola) in lentil production systems of southern Australia?Crossref | GoogleScholarGoogle Scholar |
Ouji A, Mouelhi M (2017) Influence of sowing dates on yield and yield components of lentil under semi-arid region of Tunisia. Journal of New Sciences 38, 2077–2082.
Rubiales D, Fernández-Aparicio M (2012) Innovations in parasitic weeds management in legume crops. A review. Agronomy for Sustainable Development 32, 433–449.
| Innovations in parasitic weeds management in legume crops. A review.Crossref | GoogleScholarGoogle Scholar |
Rubiales D, Annicchiarico P, Vaz Patto MC, Julier B (2021) Legume breeding for the agroecological transition of global agri-food systems: a european perspective. Frontiers in Plant Science 12, 782574
| Legume breeding for the agroecological transition of global agri-food systems: a european perspective.Crossref | GoogleScholarGoogle Scholar | 34868184PubMed |
Sharma SR, Singh S, Aggarwal N, Kaur J (2016) Effect of post-emergence herbicide metribuzin application on morpho-physiological traits, yield and yield components in lentil (Lens culinaris Medik.). In ‘Proceedings international conference on pulses’. (International Center for Agricultural Research in the Dry Areas (ICARDA): Amman, Jordan)
Sharma SR, Singh S, Aggarwal N, Kaur J, Gill RK, Kushwah A, Patil SB, Kumar S (2018) Genetic variation for tolerance to post-emergence herbicide, imazethapyr in lentil (Lens culinaris Medik.). Archives of Agronomy and Soil Science 64, 1818–1830.
| Genetic variation for tolerance to post-emergence herbicide, imazethapyr in lentil (Lens culinaris Medik.).Crossref | GoogleScholarGoogle Scholar |
Shoup DE, Al-Khatib K, Peterson DE (2003) Common waterhemp (Amaranthus rudis) resistance to protoporphyrinogen oxidase-inhibiting herbicides. Weed Science 51, 145–150.
| Common waterhemp (Amaranthus rudis) resistance to protoporphyrinogen oxidase-inhibiting herbicides.Crossref | GoogleScholarGoogle Scholar |
Smitchger JA, Burke IC, Yenish JP (2012) The critical period of weed control in lentil (Lens culinaris) in the Pacific Northwest. Weed Science 60, 81–85.
| The critical period of weed control in lentil (Lens culinaris) in the Pacific Northwest.Crossref | GoogleScholarGoogle Scholar |
Soltani N, Deen B, Bowley S, Sikkema PH (2005) Effects of pre-emergence applications of flufenacet plus metribuzin on weeds and soybean (Glycine max). Crop Protection 24, 507–511.
| Effects of pre-emergence applications of flufenacet plus metribuzin on weeds and soybean (Glycine max).Crossref | GoogleScholarGoogle Scholar |
Stork PR (1995) Field leaching and degradation of soil applied herbicides in a gradationally textured alkaline soil: chlorsulfuron and triasulfuron. Australian Journal of Agricultural Research 46, 1445–1458.
| Field leaching and degradation of soil applied herbicides in a gradationally textured alkaline soil: chlorsulfuron and triasulfuron.Crossref | GoogleScholarGoogle Scholar |
Taran B, Warkentin TD, Vandenberg A, Holm FA (2010) Variation in chickpea germplasm for tolerance to imazethapyr and imazamox herbicides. Canadian Journal of Plant Science 90, 139–142.
| Variation in chickpea germplasm for tolerance to imazethapyr and imazamox herbicides.Crossref | GoogleScholarGoogle Scholar |
Taran B, Holm F, Banniza S (2013) Response of chickpea cultivars to pre- and post-emergence herbicide applications. Canadian Journal of Plant Science 93, 279–286.
| Response of chickpea cultivars to pre- and post-emergence herbicide applications.Crossref | GoogleScholarGoogle Scholar |
Teja KC, Duary B, Dash S, Mallick RB (2017) Post-emergence application of imazethapyr for weed management in lentil. SATSA Mukhapatra - Annual Technical Issue 21, 183–188.
Tepe I, Erman M, Yazlik A, Levent R, Ipek K (2005) Comparison of some winter lentil cultivars in weed–crop competition. Crop Protection 24, 585–589.
| Comparison of some winter lentil cultivars in weed–crop competition.Crossref | GoogleScholarGoogle Scholar |
Wall DA, McMullan PM (1994) Effectiveness of several new selective herbicides in lentils (Lens culinaris). Crop Protection 13, 553–557.
| Effectiveness of several new selective herbicides in lentils (Lens culinaris).Crossref | GoogleScholarGoogle Scholar |
Yadav SS, Rizvi AH, Manohar M, Verma AK, Shrestha R, Chen C, Bejiga G, Chen W, Yadav M, Bahl PN (2007) Lentil growers and production systems around the world. In ‘Lentil: an ancient crop for modern times’. (Eds SS Yadav, D McNeil, PC Stevenson). pp. 415–442. (Springer: Dordrecht, Netherlands)
| Crossref |
Yadav R, Singh R, Yadav K (2013) Weed management in lentil. Indian Journal of Weed Science 45, 113–115.
Yan W, Tinker NA (2006) Biplot analysis of multi-environment trial data: principles and applications. Canadian Journal of Plant Science 86, 623–645.
| Biplot analysis of multi-environment trial data: principles and applications.Crossref | GoogleScholarGoogle Scholar |
Yan W, Kang MS, Ma B, Woods S, Cornelius PL (2007) GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Science 47, 643–653.
| GGE biplot vs. AMMI analysis of genotype-by-environment data.Crossref | GoogleScholarGoogle Scholar |
Yenish JP (2007) Weed management in chickpea. In ‘Chickpea breeding and management’. (Eds S Shyam, S Yadav, W Chen) pp. 233–245. (Cromwell Press. Ltd: London, UK)
| Crossref |