

Salinity tolerance and avoidance mechanisms at germination among messina (Melilotus siculus) accessions
Robert P. Jeffery A C , Megan H. Ryan A , Natasha L. Ayers A B and Phillip G. H. Nichols A
A C , Megan H. Ryan A , Natasha L. Ayers A B and Phillip G. H. Nichols A
A School of Agriculture and Environment and Institute of Agriculture, University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia.
B Current address: AgriStart, 1/14 Burler Drive, Vasse, WA 6280, Australia.
C Corresponding author. Email: robert.peter.jeffery@gmail.com
Crop and Pasture Science - https://doi.org/10.1071/CP20427
Submitted: 10 November 2020 Accepted: 17 February 2021 Published online: 21 June 2021
Journal Compilation © CSIRO 2021 Open Access CC BY
Abstract
Messina (Melilotus siculus) cv. Neptune, an annual pasture legume native to the Mediterranean Basin, has recently been released for saltland pastures in southern Australia following demonstration of biomass production and persistence superior to other commercial pasture legumes in saline environments prone to winter waterlogging. Self-regenerating annual pasture legumes also require seed adaptations for both tolerating and avoiding salinity at germination in these environments. This study examined diversity within Neptune and 20 other messina accessions for salt tolerance at germination, recovery of germinability from temporary salt stress, and timing and extent of hardseed softening, compared with balansa clover (Trifolium michelianum) cv. Frontier, burr medic (Medicago polymorpha) cv. Scimitar and white melilot (Melilotus albus) cv. Jota. Germination rates after 14 days at 300 mM NaCl relative to 0 mM NaCl were ≥99% for Neptune and 18 other messina accessions, 66% for Scimitar, 21% for Jota and 11% for Frontier. No genotype germinated at 600 mM NaCl; however, when transferred to 0 mM NaCl after 14 days at 600 mM, all genotypes except Scimitar and Jota recovered partial germination, ranging from 13% to 93% of controls (0 mM NaCl for 28 days). The softening rate of hard (impermeable) seeds in the field varied among genotypes, with deferral of hardseed softening until late autumn–early winter, when rainfall is more likely, indicating greater persistence. The months in which the hardseed level first became significantly lower than the initial level (in freshly harvested seeds) after placement on the soil surface in December were: March for Frontier; April for Scimitar, Jota and Neptune; and March–July for all messina accessions. This study confirmed that messina has high salt tolerance and several avoidance mechanisms at germination that contribute to its adaptation to saline soils in southern Australia. Several messina accessions were superior to Neptune for individual traits which could be exploited for plant breeding. These results also have implications for saltland pastures in other regions of the world with Mediterranean-type climates.
Keywords: genetic resources, germination, hardseededness, Melilotus, pasture legumes, salinity tolerance.
Introduction
Dryland salinity is a major threat to the productivity of Australian agriculture (Rogers et al. 2005; Bennett et al. 2009). In 2000, an estimated 5.7 Mha of Australian agricultural land was affected by dryland salinity, equating to an annual production loss of AU$120 million (NLWRA 2001). By 2050, this area is predicted to reach 17 Mha and cost up to $400 million annually (Government of Western Australia 2000). Mediterranean-type climates, such as occur in much of southern Australia, are characterised by dry hot summers and cool winters during which up to 65% of the annual rainfall is received, and a growing season of 4–9 months (Porqueddu et al. 2016). In these environments, the development and commercial release of salt-tolerant plant species adapted to saline environments is crucial to ensure the sustainability of agricultural productivity (Howieson et al. 2000; Rogers et al. 2005; Hughes et al. 2008).
Inhibition of germination under saline conditions results from either osmotic effects of salts that lower the water potential or toxic effects of salts that damage cell membranes and cause leakages, or both effects combined (Ungar 1962). In southern Australia, NaCl, the predominant salt in >90% of soils, is generally responsible for the toxic effects of salinity on plant function (NLWRA 2001). Many saline soils are also prone to winter waterlogging in southern Australia (McFarlane and Williamson 2002), with the salt tolerance of most plants severely compromised under waterlogged conditions (Barrett-Lennard and Shabala 2013). This represents a significant limitation to the productivity of saltland pastures.
Since the 1890s, self-regenerating annual pasture legumes native to the Mediterranean region have been vital components of agriculture in southern Australian, utilised for livestock grazing and fodder production and for their ability to fix atmospheric nitrogen (N2) (Robson 1990; Howieson et al. 2000; Nichols et al. 2012). The main species sown have been subterranean clover (Trifolium subterraneum L.) and annual medics (Medicago spp.), with other legumes also introduced since the mid-1980s (Smetham 2003; Loi et al. 2005; Nichols et al. 2007, 2012). These legumes are also sown in other regions with Mediterranean or quasi-Mediterranean climates, including the Mediterranean Basin itself, the southern Cape of South Africa, central Chile and Argentina, California and southern United States of America, and parts of New Zealand (Porqueddu et al. 2016).
However, subterranean clover is highly sensitive to salt (West and Taylor 1981), whereas the commonly sown annual medic species, particularly burr medic (Medicago polymorpha L.), have moderate salt tolerance but susceptibility to waterlogging (Bennett et al. 2009; Rogers et al. 2011; Kotula et al. 2019). Among other annual legumes previously sown in saline areas, balansa clover (T. michelianum L.) has high waterlogging tolerance but tolerates only mild salinity (Rogers and Noble 1991; Kotula et al. 2019), and white melilot (Melilotus albus Medik.) is susceptible to waterlogging (Evans and Kearney 2003; Rogers et al. 2008). No other commercial pasture legume species had tolerance to both stresses (Nichols et al. 2008). This led to efforts to find new annual pasture legumes adapted to saline areas prone to winter waterlogging (Dear et al. 2008).
Following a review by Rogers et al. (2005) of salt-tolerant annual legumes, glasshouse experiments were initiated to identify species with salt and waterlogging tolerance (Rogers et al. 2008, 2011; Teakle et al. 2011, 2012; Striker et al. 2015; Striker and Colmer 2016). Concurrent field studies were conducted across southern Australia to identify annual legumes able to persist on saline soils prone to waterlogging (Nichols et al. 2008). These studies identified messina (Melilotus siculus (Turra) Vitman ex. B.D.Jacks), an aerial-seeding annual legume adapted to saline marshes in the Mediterranean Basin (Marañon et al. 1989), as the most tolerant species to the dual stresses of salinity (electrical conductivity (ECe) ≤30 dS/m in the top 10 cm of soil during summer) and winter waterlogging. This coincided with the identification and subsequent release of a salt-tolerant Sinorhizobium medicae strain able to nodulate messina (Bonython et al. 2011; Ballard et al. 2014). Subsequent field trials on saline, waterlogged soils with 21 messina accessions collected from the wild identified one accession (SA40002), later released as cv. Neptune, as the most productive and persistent genotype (Nichols et al. 2019). These trials also confirmed the superior agronomic performance of messina over balansa clover, burr medic and white melilot under these conditions.
Apart from high salt tolerance during the vegetative phase, messina also possesses adaptations for both tolerating and avoiding salinity at germination (Nichols et al. 2009). This is particularly important for self-regenerating annual pasture legumes, which must germinate and establish seedlings each year. Soil salinity levels in the top 10 cm are highest in southern Australia over summer and generally decline after the first germinating rains of the growing season, due to salts being leached down the profile with further rainfall (McFarlane and Williamson 2002; Nichols et al. 2008). The extent and timing of these rainfall events are important for defining the duration and level of surface salinity experienced by germinating seedlings. Thus, if rainfall is sufficient to leach salts from the soil surface, germination can occur in soil with relatively low salinity and only the most salt-sensitive species will fail to establish. If, however, these rains are sufficient for germination but insufficient to leach salts, high seedling losses will occur for all but the most salt-tolerant species. Thus, an annual pasture legume adapted to saline environments must have high salinity tolerance as a seedling and/or mechanisms to avoid germination at times of high salinity.
The ability of imbibed seeds to tolerate salinity at germination, when plants are most susceptible to salt, is an important adaptation for self-regenerating annual legumes in saline environments (Rogers et al. 2005; Nichols et al. 2009; Rogers et al. 2011). Nichols et al. (2009) showed that messina suffered no loss of germination at 300 mM NaCl, relative to the non-saline control, compared with only 23% germination for burr medic and 18% for balansa clover. Rogers et al. (2011) showed variation among 15 messina accessions, including Neptune, for ability to germinate at 320 mM NaCl, suggesting the potential to select messina genotypes with better salinity tolerance than Neptune.
The role of hardseededness (seed coat impermeability) in spreading germination of annual pasture legumes over several seasons is well understood for non-saline environments (Taylor 2005). Newly ripened seeds of most species contain a high proportion of hard (impermeable) seeds. Some of these ‘soften’ (become permeable) over the summer–autumn period and are available to re-establish the species, whereas residual hard seeds form a seedbank and soften over subsequent summers. The timing of seed softening is critical for seedling survival. In seasons with germination-inducing rainfall followed by a drought period that kills seedlings (termed a ‘false break’), species that defer hardseed softening until late autumn or early winter, when rainfall is more likely, tend to be more persistent (Norman et al. 1998, 2005; 2006; Loi et al. 2005). Such false breaks occur in 61–72% of years in southern Australia (Chapman and Asseng 2001).
Nichols et al. (2009) demonstrated that hardseededness and delayed seed softening are important traits in annual legumes adapted to saline land, by spreading the risks of germination over seasons and avoiding germination at times of high salinity. The impermeable seed coat shields seeds from the toxic effects of salt ions, and delayed hardseed softening acts to defer germination until salts are more likely to have been flushed from the soil surface. Nichols et al. (2009) showed that messina had later seed softening than balansa clover, burr medic and subterranean clover. However, the timing of seed softening has not been investigated for Neptune and the variation among messina genotypes is unknown.
Another adaptive mechanism to saline environments is the ability of permeable seeds to recover germinability in non-saline solution following exposure to highly saline solution, such as occurs with germination after sporadic rainfall events at the start of the growing season. Nichols et al. (2009) showed messina seeds recovered 31% germinability upon transfer to non-saline solution following exposure to 600 mM NaCl for 21 days, compared with 0% for both burr medic and subterranean clover. Teakle et al. (2010) reported similar findings among accessions of Lotus tenuis Waldst. & Kit. ex Willd. This adaptive mechanism has not been examined for individual messina genotypes, including Neptune.
This paper examines diversity within Neptune and 20 other messina accessions for germination and seedling traits important for adaptation to saline soils, compared with cultivars of balansa clover, burr medic and white melilot. The studied traits include tolerance to salinity at germination, the ability to recover germination in non-saline conditions following temporary exposure to salinity, and timing of hardseed softening over the summer–autumn period following seedset. We hypothesised that Neptune would demonstrate performance for these traits superior to the other species. We also hypothesised that diversity would exist for these traits within messina. Identification of messina accessions with traits superior to Neptune could lead to their inclusion in breeding programs to develop cultivars with even greater adaptation to saline environments.
Materials and methods
Plant material
Cultivar Neptune (formerly known as accession SA40002) and 20 other messina accessions were compared with three commercial control species: balansa clover cv. Frontier, burr medic cv. Scimitar and white melilot cv. Jota (Table 1). All germplasm was originally sourced from the Australian Pastures Genebank, operated by the South Australian Research and Development Institute. The term ‘genotype’ is used to describe all cultivars and accessions in the study.

|
Plants were grown to produce seeds for this study at the Department of Primary Industries and Regional Development (DPIRD) South Perth campus (31.98°S, 115.88°E). Sowing date was 31 May 2010. Two replicates of each genotype were sown in a randomised block design. Plots consisted of single rows 3.5 m long, sown to 2.5 g seed and 180 kg/ha of Super Potash 3 : 1 fertiliser (6.8% P, 12.4% K, 7.9% S, 15.0% Ca; CSBP Fertilisers). Before sowing, seeds were scarified with compressed air in a brass cylinder with fine sandpaper to overcome hardseededness (seed coat dormancy). Messina seeds were inoculated with the Sinorhizobium medicae strain specially developed for messina, SRDI 554 (Bonython et al. 2011); Scimitar and Jota seeds were inoculated with the commercial Group AM S. medicae strain, WSM 1115; and Frontier seeds were inoculated with the commercial Group C Rhizobium leguminosarum bv. trifolii strain WSM 1325.
Plots were hand-weeded and irrigated by overhead sprinkler when required. Plants were checked every 2–3 days, and time to first flowering was recorded as the number of days from sowing until plots had at least 50% of plants with at least one open flower. Watering ceased on 10 November, when all messina plots were well into seedset and the latest flowering entry, Jota, had commenced flowering. Plants were harvested by hand on 3 December, except for Jota, which was harvested on 18 December. Freshly harvested pods were dried at 40°C for 3 days. A sample of dried pods equivalent to ~5000 seeds was then weighed out from each plot for Expt 1. These were carefully rubbed between a cork block and a corrugated rubber mat to remove seeds from pods without scarification of the seed coat, according to the procedure of Nichols et al. (2009). The remaining pods were mechanically scarified with a thresher for use in Expt 2.
Experiment 1:timing and extent of hardseed softening
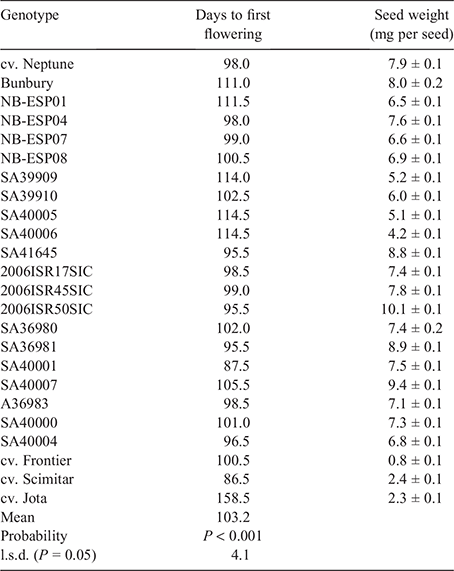
Softening of hard seeds was measured in the field over 530 days in 24 genotypes with nine sampling times and three replicates. Unscarified seeds of each genotype were counted into 27 lots of 100 seeds by using a Contador electronic seed counter (Pfeuffer, Germany). The weights of six samples of 100 seeds were recorded for each genotype and used to calculate the mean seed weight (mg per seed) (Table 2).
|
Fibreglass fly-wire (1.5 mm mesh) was cut and sealed to make 24 individual strips, each consisting of 24 pockets (80 mm by 50 mm), into which 100 seeds of individual genotypes were placed. The genotype locations within each strip were randomised. The mesh strips were pinned to the soil surface in full sunlight at the DPIRD South Perth campus on 23 December 2010. Three mesh strips, each representing a replicate, were removed from the field every 28 days and germination tests performed on the seeds that remained in each pocket. Sampling dates in 2011 (no. of days in the field) were 20 January (28 days), 17 February (56 days), 17 March (84 days), 14 April (112 days), 12 May (140 days), 9 June (168 days) and 7 July (196 days). Three strips were kept in the field for a further year and sampled on 5 June 2012 (530 days). The extra seed lot was used to measure initial hardseed levels of freshly harvested seeds on 23 December 2010.
Germination tests were undertaken within 5 days of removal from the field. Seeds were placed in Petri dishes (90 mm diameter) lined with two Whatman No. 1 filter papers and moistened with a 50 : 50 tap-water/deionised water solution. Petri dishes were incubated at 15°C for 14 days, with germinated seeds counted and removed every 3 days. Imbibed seeds were considered to be those that had swollen, and germination was defined as the emergence of a radicle at least 5 mm in length. Decayed seeds, defined as imbibed seeds able to be compressed with forceps and covered in fungal growth, were removed to avoid contamination following counting. Unimbibed seeds that could not be compressed with forceps were classified as hard seeds. The hardseed percentage was calculated as the number of hard seeds remaining after 14 days as a proportion of the total number of seeds originally placed in the flywire pocket.
Experiment 2: salt tolerance at germination and recovery from temporary salinity stress
This experiment examined the relative salt tolerance of germinating seeds and the ability of imbibed seeds temporarily exposed to high levels of salinity to recover germination in non-saline conditions. The experimental design consisted of the 24 genotypes used in Expt 1 with five salinity treatments and four replicates (n = 50 scarified seeds); Messina accession 2006ISR50SIC had only three replicates because of insufficient seed quantities. Germination conditions were the same as for Expt 1, except that Petri dishes were sealed with Parafilm to prevent evaporation. Salinity treatments consisted of: (i) 0, 300 or 600 mM NaCl solutions, all containing a basal 5 mM CaSO4; and (ii) two additional ‘recovery’ treatments involving the transfer of seeds from either 300 or 600 mM NaCl solution after 14 days to a non-saline (0 mM NaCl) solution for a further 14 days.
After the initial 14 days of treatment, imbibed seeds that had not germinated were transferred to new Petri dishes containing solutions of either 0 mM NaCl (‘recovery’ treatment) or the same NaCl concentration (300 or 600 mM) and then incubated for a further 14 days. Germination percentages were calculated for all treatments as the proportion of imbibed seeds after the first 14 days in the original solution, and after a further 14 days in the second solution. The same definitions of imbibed, germinated and decayed (dead) seeds as in Expt 1 were used, but the number of decayed seeds removed was subtracted from the total sample size.
For the two ‘recovery treatments’, the percentage germination recovered after 14 days at 0 mM NaCl of seeds transferred from 300 or 600 mM NaCl was calculated as a percentage of the germination in the non-saline control (0 mM NaCl) after 28 days.
Statistical analyses
Genotypic differences in the timing of hardseed softening and the percentage germination of imbibed seeds after 14 days in 300 or 600 mM NaCl relative to 0 mM NaCl were examined using a two-way analysis of variance (ANOVA) with α = 0.05. The sample dates for significant softening, defined as a significant (P < 0.05) reduction in hardseed levels compared with those in freshly harvested seed, were detected using one-way ANOVA and least significant differences (l.s.d.). Germination recovery data were analysed with a general ANOVA after checking for normality and homogeneity of residuals. No transformations were required, based on normal and residual plots. The l.s.d. values at P = 0.05 were calculated. Statistical analyses were conducted using Genstat 12th Edition (VSN International).
Results
Experiment 1: field softening of hard seeds
Flowering times for messina accessions varied from 88 to 115 days after sowing (DAS), with Neptune flowering at 98 DAS (Table 2). The earliest and latest flowering genotypes were Scimitar (87 DAS) and Jota (159 DAS), respectively; Frontier flowered at 101 DAS (Table 2). The mean seed weight was 7.9 mg for Neptune and ranged from 4.2 to 10.1 mg for all 20 messina accessions. Seeds were comparatively smaller for Scimitar (2.4 mg), Jota (2.3 mg) and especially Frontier (0.8 mg) (Table 2).
Seed softening patterns are presented with messina genotypes grouped according to country of origin (Fig. 1). All genotypes had hardseed levels of >80% initially (freshly harvested, day 0 in Fig. 1), except for messina accessions SA40006, SA39909 and SA40005, with 64%, 65% and 78%, respectively. The proportion of hard seeds remaining after 196 days (8 months) in the field differed (P < 0.001) among genotypes (Fig. 1). Softening events, defined here as the date (no. of days in the field) at which the hardseed level first became significantly lower (P < 0.05) than the initial level, also differed (P < 0.001) among genotypes. Softening occurred later and to a lesser extent for messina genotypes from Spain, Russia, Tunisia and Italy than Neptune. The earliest softening event occurred by 17 March (84 days) for messina accessions SA36981, 2006ISR17SIC and NB-ESP07 and Frontier; by 14 April (112 days) for Neptune, 13 accessions, Scimitar and Jota; by 12 May (140 days) for accessions SA39909, Bunbury and NB-ESP08; and by 7 July (196 days) for accession SA40007 (Fig. 1).

|
The date by which at least 33% of initial hard seeds had softened was 17 March (84 days) for Frontier; 14 April (112 days) for Scimitar, Neptune and accessions SA36981, 2006ISR17SIC and 2006ISR45SIC; and 12 May (140 days) for Jota and accessions SA36983, SA39910, SA40000, SA40001, SA40005, SA40006, SA41645 and 2006ISR50SIC (Fig. 1). The latest date by which at least 33% of initial hard seeds had softened was 9 June (168 days) for messina accessions SA36980, SA39909 and NB-ESP01. However, seeds of six accessions (SA40004, SA40007, Bunbury, NB-ESP04, NB-ESP07 and NB-ESP08) did not soften to this degree (Fig. 1).
The date by which the absolute maximum softening of hard seeds had occurred was 12 May (140 days) for messina accession SA41645 and Scimitar; 9 June (168 days) for Neptune, 12 accessions and Frontier; and 7 July (196 days) for seven accessions and Jota.
Considering hardseed levels on 7 July (196 days in the field), accession SA40007 was the hardest genotype (85%), followed by four accessions (NB-ESP04, NB-ESP08, SA40004, NB-ESP07) and Scimitar, all of which had >70% hard seeds. Neptune (39%) and the 14 other accessions (except SA40006) had intermediate hardseed levels (range of 27–68%). The three genotypes with the lowest final hardseed levels were accession SA40006 (14%), Frontier (25%) and Jota (5%). These levels equate to the proportion of seeds remaining in the soil as dormant seeds for germination in subsequent years. Of interest was the slight increase in hardseed level in several genotypes between the June and July sampling times.
The residual hardseed levels of the last lot of seeds, removed on 5 June 2012 (after 530 days in the field), are shown in Fig. 2. Neptune had 7.9% of its original seeds retained as hard seeds, and Scimitar 4.7%. Only five messina accessions had >21% of seeds retained as hard seeds (33–58% for NBESP04, NBESP08, SA40004, SA40007 and SA41645). No hard seeds remained for Frontier and Jota.

|
Experiment 2
Salt tolerance at germination
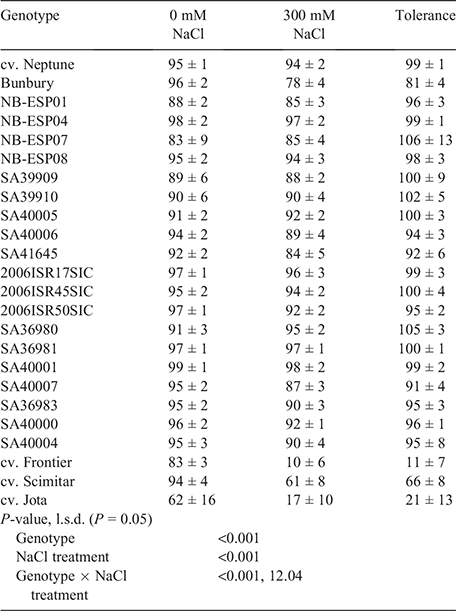
The germination of imbibed seeds at 0 mM NaCl for 14 days was >90% for all genotypes except accessions SA39909 (89%), NB-ESP01 (88%) and NB-ESP07 (83%), Frontier (83%) and Jota (62%) (Table 3). The effect of 300 mM NaCl on germination after 14 days differed (P < 0.001) among genotypes. Neptune and 10 accessions were tolerant of 300 mM NaCl after 14 days, with no difference in germination relative to 0 mM NaCl; germination at 300 mM NaCl relative to the non-saline control was 99% for Neptune, >90% for 19 accessions, 81% for the accession Bunbury and 66% for Scimitar, but only 21% for Jota and 11% for Frontier (Table 3). No seeds of any genotype germinated after 14 days at 600 mM NaCl.
|
Recovery of germination in non-saline conditions following temporary exposure to salinity stress
The percentage of total seeds that imbibed after 28 days was >96% for all genotypes across all salinity levels. Germination percentages of imbibed seeds did not differ between 0 and 300 mM NaCl treatments after 28 days for Neptune and all 20 accessions (Table 4). By contrast, germination percentage was greatly reduced after 28 days at 300 mM NaCl relative to the non-saline control for Frontier (22%), Jota (23%) and Scimitar (78%).
|
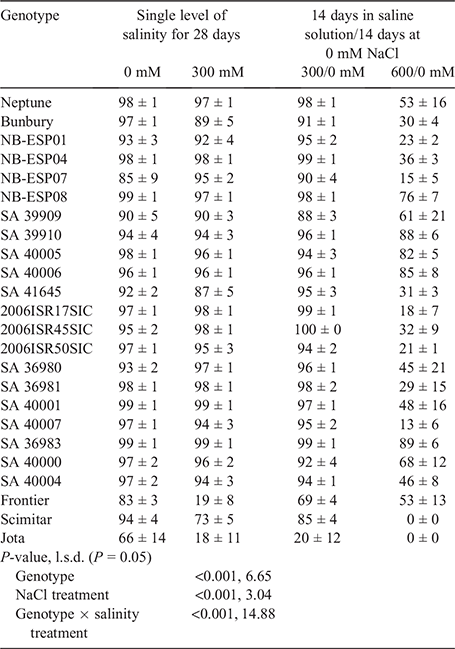
The percentage germination of imbibed seeds when transferred to 0 mM NaCl for a further 14 days following 14 days at 300 mM NaCl was 20% for Jota, 69% for Frontier and 85% for Scimitar, whereas it was 98% for Neptune and ≥88% for all accessions (Table 4). After 14 days at 0 mM NaCl following 14 days at 300 mM NaCl, there were no differences in germination relative to 28 days at 0 mM NaCl for Neptune and 17 accessions; the other three accessions (SA40000, SA40005 and Bunbury) had lower (P < 0.05) germination, but these reductions (<6%) were relatively small. On the other hand, seeds of Jota recovered only 26% germination in 0 mM NaCl after 14 days at 300 mM NaCl relative to 28 days at 0 mM NaCl; 83% and 91% of seeds recovered germination in Frontier and Scimitar, respectively.
When transferred to 0 mM NaCl following 14 days at 600 mM NaCl, three accessions (SA36983, SA39910 and SA40006) recovered full germinability (not significantly different from non-saline controls at 28 days) (Table 4; values displayed as percentage of germination of non-saline controls in Fig. 3). Partial germination, whereby germination was lower (P < 0.05) than at 0 mM NaCl for 28 days, was recovered by Neptune (54%) and the other 17 accessions (14–84%) (Fig. 3). Three of these accessions (SA40000, SA40005 and NB-ESP08) recovered >70% germination, while the percentage germination recovered in the remaining 14 accessions varied from 15% to 70% of the non-saline controls (Fig. 3). Of the control species, Frontier also recovered partial germination (64% of non-saline control) when transferred to 0 mM NaCl after 14 days at 600 mM NaCl; however, Scimitar and Jota had no recovery of germination (Fig. 3).

|
Discussion
This study confirms that messina is better adapted than existing options for saltland pastures in southern Australia. It has a high tolerance of salinity at germination, able to recover germination from temporary exposure to very high salinity levels, and seed softening patterns that allow the majority of seeds to germinate when salt concentrations at the soil surface are likely to be lowest. Neptune and all 20 messina accessions had superior tolerance to germination at 300 mM NaCl, better recovery of germinability following transfer from 600 to 0 mM NaCl, and delayed seed softening compared with Frontier balansa clover, Scimitar burr medic and Jota white melilot, which are the three ‘next best’ commercially available annual pasture legume species for saltland pastures (DPIRD 2019; Nichols et al. 2019). This study supports observations by Nichols et al. (2009) from a composite of three messina accessions when compared with Frontier and Scimitar. It also supports the findings of Rogers et al. (2011), who observed variation among Neptune and 15 other messina accessions in their ability to germinate at 240 and 320 mM NaCl.
The ability to germinate at 300 mM equates to an ECe of ~30 dS/m, depending on soil texture (Barrett-Lennard et al. 2013). This is consistent with the field results of Nichols et al. (2008, 2019), who found that messina was able to persist over several seasons on soils with surface (0–10 cm) ECe levels that reached a maximum of 30 dS/m over summer.
Despite the superior persistence and agronomic performance of Neptune in the field compared with other messina accessions (Nichols et al. 2019), in the present study it did not show the highest expression for any of the mechanisms of salt tolerance and avoidance among the messina accessions examined. This implies the potential to utilise plant breeding to develop cultivars better adapted than Neptune to germination in saline soils with seasonally high surface salinity. It also suggests that there are other factors influencing adaptation not measured in this study. These results have implications for the development of cultivars better suited to saline soils prone to waterlogging, because tolerance to waterlogging in the absence of salinity (Rogers et al. 2011) and presence of salinity in glasshouse experiments varied among messina accessions, some of which had better tolerance that Neptune (Striker et al. 2015). The development of new messina cultivars with improved adaptions for germination on saline soils could dramatically increase the profitability of saltland pastures in southern Australia and other regions of the world with Mediterranean-type climates.
Timing and extent of hardseed softening and its relevance for saline soils in Mediterranean type climates
The temporal patterns and extent of hardseed softening found in this study are consistent with the findings of Nichols et al. (2009), who reported that, by June (after 168 days in the field), a mixture of three messina accessions (SA36980, SA36982 and SA36983) had a hardseed level of 62%; this is similar to the results for SA36980 (42%) and SA36983 (51%) in the present study, although the first significant softening event occurred a month earlier than in the former study. The general trends in timing, rate and extent of hardseed softening for Frontier and Scimitar in this study were similar to those of previous studies by Nichols et al. (2009) and Loi et al. (2005); no comparable softening data exist for Jota.
The significance of the timing and rate of seed softening as a salinity avoidance mechanism is related to the timing of the break of season, defined by the first germination-inducing rain events, which generally occur from April to early May in southern Australia (Chapman and Asseng 2001). In these regions, soil surface (0–10 cm) salinity concentrations tend to be highest over summer and at the break of season; they then generally decrease following rain events sufficient to flush salt from the soil surface (McFarlane and Williamson 2002; Nichols et al. 2008). For instance, surface salinity (0–10 cm) at a field site at Darkan, Western Australia, was highest (ECe >30 dS/m) from mid-January to mid-May (summer–autumn) and lowest (ECe <8 dS/m) from mid-July to mid-September (winter) (Nichols et al. 2010). The impermeable testa (seed coat) of hard seeds prevents the uptake of Na+ and, presumably, Cl– ions, essentially shielding seeds from ion toxicity in saline conditions (Nichols et al. 2009). Seed softening affects the timing of germination and the proportion of seeds that are able to germinate following any given rainfall event (Taylor 2005). Delayed seed softening is an important salinity avoidance mechanism enabling germination to occur when reliable rainfall events and reductions in soil surface salinity are most likely, that is, after the break of season (Nichols et al. 2009). Thus, softening rate can also contribute to distributing the risk of germination over several periods within a season or even over multiple seasons (Norman et al. 2005, 2006).
Differences in the timing and extent of hardseed softening over 196 days among Neptune and the 20 individual messina accessions have implications for determining the best adapted messina genotypes for avoidance of seasonally high surface salinity at germination in specific environments. For instance, in relation to timing of hardseed softening, accessions SA40007, SA39909, Bunbury and NB-ESP04 would be best suited to germinating in seasonally saline soils in southern Australia because they soften later than all other genotypes (by May), restricting germination until continuing winter rainfall events are more likely and salts have been flushed from the soil surface (Nichols et al. 2008). By contrast, Neptune, Scimitar, Jota and 13 messina accessions first softened by mid-April, whereas Frontier and four accessions softened the earliest (by March). This means that the proportion of seeds able to germinate in response to a specific summer rainfall event before April, when the risk of seedling losses from extremely saline conditions or drought is highest in southern Australia, would be less for all messina genotypes than for Frontier (Taylor 2005).
There was also variation in the extent of seed softening among messina accessions, which has consequences for their persistence across seasons (Norman et al. 2005; Taylor 2005). By July, after one summer–autumn period of softening, 15 messina accessions had higher hardseed levels (41–84%) than Neptune (39%), which translates to a greater proportion of residual hard seeds remaining dormant in the soil for germination in following years. This enables self-regeneration in the absence of significant seed production the previous year (Norman et al. 2005; Nichols et al. 2009), particularly important in risky environments for germination and plant growth, such as those subject to salinity. Frontier and Jota, with low levels of residual hard seeds carried over in the seedbank, are at much greater risk of pasture failure should seedset be severely impacted. Five messina accessions still had >33% of their original seed retained as hard seeds after two summer–autumn periods. However, such levels may be too high for reliable regeneration, because messina is likely to be grown in permanent pastures on saline areas, requiring germination of new seedlings each year (Nichols et al. 2019). This contrasts with annual legumes for rotation with crops, which require high levels of hard seeds for reliable self-regeneration after the cropping phase (Howieson et al. 2000; Loi et al. 2005; Taylor 2005; Nichols et al. 2012). Overall, the variation in timing and extent of hardseed softening suggests the potential to develop new cultivars better adapted than Neptune to saline environments in southern Australia.
The slight increase in hardseed levels observed in some genotypes between the June and July sampling times has been observed in other annual legume seed-softening studies (Nichols et al. 2009; PGH Nichols, DM Peck, A Stefanski, BJ Wintle and RJ Simpson, unpubl. data). The reason and ecological significance are unknown, but the phenomenon is worthy of further investigation.
Salt tolerance at germination
This study concurs with Rogers et al. (2011) that variation in salt tolerance at germination exists among messina accessions. However, germination after 28 days at 300 mM NaCl in this study was much higher for all messina accessions than at 320 mM in the study of Rogers et al. (2011), whereas Neptune, SA40000, SA40006 and SA40007 had similar germination at 240 mM NaCl. Rogers et al. (2011) also reported that both Frontier and Scimitar had no germination at 320 mM NaCl, in contrast to 19% and 73%, respectively, at 300 mM NaCl in the present study. This suggests that there may be a threshold salinity level between 300 and 320 mM NaCl at which messina and Scimitar become more susceptible to salinity. Differences could also be related to the 15–20°C incubation temperatures used in this study versus the 17–26°C regime used by Rogers et al. (2011); the inhibitory effects of salinity on germination can be magnified at higher temperatures (Guan et al. 2009). The high tolerance to 300 mM NaCl at germination of Neptune and all 20 messina accessions examined is an important finding, because it suggests that selection can be focused on other useful agronomic traits to develop new messina cultivars.
Germination recovery upon transfer from extremely saline to non-saline conditions
The ability to recover germination following a decrease in high salt concentrations at the soil surface is important for allowing imbibed seeds at the break of season to survive and germinate once salts are leached by follow-up winter rainfall (Redmann 1974; Nichols et al. 2009). However, the ability to recover germination is an advantage only in situations where follow-up rains occur before the soil dries out and the seed perishes (Taylor 2005). The finding that Neptune and all other messina accessions recovered partial germination upon transfer from 600 mM NaCl to non-saline solution confirms the results of Nichols et al. (2009) for a mixture of three accessions. Also important was the finding of significant variation for recovery, with two accessions (SA36983 and SA39910) having >90% recovery and five other accessions having >68% recovery, compared with Neptune with 54% recovery. Of interest was the finding that Frontier balansa clover recovered 63% of germination after transfer from 600 mM NaCl, also confirming previous results of Nichols et al. (2009), whereas Scimitar and Jota did not recover any germination.
The large range among messina accessions in ability to recover germination after exposure to high levels of salinity has not been demonstrated before and has implications for breeding programs to select messina cultivars with greater ability to recover than Neptune. Despite the important implications of these findings, the actual mechanism of recovery is poorly understood and requires further investigation to improve understanding of its adaptive importance for saline soils.
Conclusions
This study has confirmed that messina has high salt tolerance, and several avoidance mechanisms at germination that are generally lacking in balansa clover, burr medic and white melilot; these help to explain its superior growth and persistence on saline soils in southern Australia. Also of note is the variation for these traits among messina accessions, with several being superior for individual traits to the current cultivar, Neptune. This has implications for plant breeding for the development of new messina cultivars with even greater adaptation to saline soils. These results have applicability beyond southern Australian agriculture and could benefit the profitability of saltland pastures in other regions with Mediterranean-type climates around the world.
Conflicts of interest
The authors declare no conflict of interest.
Acknowledgements
We express gratitude to Mr Bradley Wintle, Mr Richard Snowball and other members of DPIRD for their technical assistance and use of facilities, and to Marcia Vistisen for her advice on seed testing procedures. This research was conducted by the senior author as part of an Honours project at the University of Western Australia and did not receive any specific funding.
References
Ballard RA, Charman N, Bonython AL, Nichols PGH, Craig AD (2014) SRDI 544: a new inoculant strain of Sinorhizobium medicae for use on messina (Melilotus siculus): an annual legume for salt-land pastures. In ‘Proceedings of the 17th Australian Nitrogen Fixation Conference’. Adelaide, S. Aust. (Eds VVSR Gupta, M Unkovich, BN Kaiser) pp. 92–94. (Australian Society of Nitrogen Fixation: Adelaide, SA)Barrett-Lennard EG, Shabala SN (2013) The waterlogging/salinity interaction in higher plants revisited: focusing on the hypoxia-induced disturbance to K+ homeostasis. Functional Plant Biology 40, 872–882.
| The waterlogging/salinity interaction in higher plants revisited: focusing on the hypoxia-induced disturbance to K+ homeostasis.Crossref | GoogleScholarGoogle Scholar | 32481157PubMed |
Barrett-Lennard EG, Bennett SJ, Altman M (2013) Survival and growth of perennial halophytes on saltland in a Mediterranean environment is affected by depth to watertable in summer as well as subsoil salinity. Crop & Pasture Science 64, 123–136.
| Survival and growth of perennial halophytes on saltland in a Mediterranean environment is affected by depth to watertable in summer as well as subsoil salinity.Crossref | GoogleScholarGoogle Scholar |
Bennett SJ, Barrett-Lennard EG, Colmer TD (2009) Salinity and waterlogging as constraints to saltland pasture production: a review. Agriculture, Ecosystems & Environment 129, 349–360.
| Salinity and waterlogging as constraints to saltland pasture production: a review.Crossref | GoogleScholarGoogle Scholar |
Bonython AL, Ballard RA, Charman N, Nichols PGH, Craig AD (2011) New strains of rhizobia that nodulate regenerating messina (Melilotus siculus) plants in saline soils. Crop & Pasture Science 62, 427–436.
| New strains of rhizobia that nodulate regenerating messina (Melilotus siculus) plants in saline soils.Crossref | GoogleScholarGoogle Scholar |
Chapman R, Asseng S (2001) An analysis of the frequency and timing of false break events in the Mediterranean region of Western Australia. Australian Journal of Agricultural Research 52, 367–376.
| An analysis of the frequency and timing of false break events in the Mediterranean region of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Dear BS, Reed KFM, Craig AD (2008) Outcomes of the search for new perennial and salt tolerant pasture plants for southern Australia. Animal Production Science 48, 578–588.
| Outcomes of the search for new perennial and salt tolerant pasture plants for southern Australia.Crossref | GoogleScholarGoogle Scholar |
DPIRD (2019) Neptune messina: a new pasture legume for saline soils prone to waterlogging. Agriculture and Food, Department of Primary Industries and Regional Development, Perth, W. Aust. Available at: https://www.agric.wa.gov.au/neptune (accessed 10 August 2020)
Evans PM, Kearney GA (2003) Melilotus albus (Medik.) is productive and regenerates well on saline soils of neutral to alkaline reaction in the high rainfall zone of south-western Victoria. Australian Journal of Experimental Agriculture 43, 349–355.
| Melilotus albus (Medik.) is productive and regenerates well on saline soils of neutral to alkaline reaction in the high rainfall zone of south-western Victoria.Crossref | GoogleScholarGoogle Scholar |
Government of Western Australia (2000) Natural resource management in Western Australia: State Salinity Strategy 2000. Prepared by the State Salinity Council, Perth, W. Aust.
Guan B, Zhou D, Zhang H, Tian Y, Japhet W, Wang P (2009) Germination responses of Medicago ruthenica seeds to salinity, alkalinity, and temperature. Journal of Arid Environments 73, 135–138.
| Germination responses of Medicago ruthenica seeds to salinity, alkalinity, and temperature.Crossref | GoogleScholarGoogle Scholar |
Howieson JG, O’Hara GW, Carr SJ (2000) Changing roles for legumes in Mediterranean agriculture: developments from an Australian perspective. Field Crops Research 65, 107–122.
| Changing roles for legumes in Mediterranean agriculture: developments from an Australian perspective.Crossref | GoogleScholarGoogle Scholar |
Hughes SJ, Snowball R, Reed KFM, Cohen B, Gajda K, Williams AR, Groeneweg SL (2008) The systematic collection and characterisation of herbaceous forage species for recharge and discharge environments in southern Australia. Australian Journal of Experimental Agriculture 48, 397–408.
| The systematic collection and characterisation of herbaceous forage species for recharge and discharge environments in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Kotula L, Kwa HY, Nichols PGH, Colmer TD (2019) Tolerance and recovery of the annual pasture legumes Melilotus siculus, Trifolium michelianum and Medicago polymorpha to soil salinity, soil waterlogging and the combination of these stresses. Plant and Soil 444, 267–280.
| Tolerance and recovery of the annual pasture legumes Melilotus siculus, Trifolium michelianum and Medicago polymorpha to soil salinity, soil waterlogging and the combination of these stresses.Crossref | GoogleScholarGoogle Scholar |
Loi A, Howieson JG, Nutt BJ, Carr SJ (2005) A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Australian Journal of Experimental Agriculture 45, 289–299.
| A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems.Crossref | GoogleScholarGoogle Scholar |
Marañon T, Garcia LV, Troncoso A (1989) Salinity and germination of annual Melilotus from the Guadalquivir Delta (SW Spain). Plant and Soil 119, 223–228.
| Salinity and germination of annual Melilotus from the Guadalquivir Delta (SW Spain).Crossref | GoogleScholarGoogle Scholar |
McFarlane DJ, Williamson DR (2002) An overview of water logging and salinity in southwestern Australia as related to the ‘Ucarro’ experimental catchment. Agricultural Water Management 53, 5–29.
| An overview of water logging and salinity in southwestern Australia as related to the ‘Ucarro’ experimental catchment.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Loi A, Nutt BJ, Evans PM, Craig AD, Pengelly BC, Dear BS, Lloyd DL, Revell CK, Nair RM, Ewing MA, Howieson JG, Auricht GA, Howie JH, Sandral GA, Carr SJ, de Koning CT, Hackney BF, Crocker GJ, Snowball R, Hughes SJ, Hall EJ, Foster KJ, Skinner PW, Barbetti MJ, You MP (2007) New annual and short-lived perennial pasture legumes for Australian agriculture: 15 years of revolution. Field Crops Research 104, 10–23.
| New annual and short-lived perennial pasture legumes for Australian agriculture: 15 years of revolution.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Craig AD, Rogers ME, Albertsen TO, Miller SM, McClements DR, Hughes SJ, D’Antuono MF, Dear BS (2008) Production and persistence of annual pasture legumes at five saline sites in southern Australia. Australian Journal of Experimental Agriculture 48, 518–535.
| Production and persistence of annual pasture legumes at five saline sites in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Malik AI, Stockdale M, Colmer TD (2009) Salt tolerance and avoidance mechanisms at germination of annual pasture legumes: importance for adaptation to saline environments. Plant and Soil 315, 241–255.
| Salt tolerance and avoidance mechanisms at germination of annual pasture legumes: importance for adaptation to saline environments.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Barrett-Lennard EG, Bennett S (2010) Pasture legumes and grasses for saltland. Farmnote 432, June 2010. Department of Agriculture and Food, Western Australia, Perth.
Nichols PGH, Revell CK, Humphries AW, Howie JH, Hall EJ, Sandral GA, Ghamkhar K, Harris CA (2012) Temperate pasture legumes in Australia—their history, current use, and future prospects. Crop & Pasture Science 63, 691–725.
| Temperate pasture legumes in Australia—their history, current use, and future prospects.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Ballard RA, Pearce AL, Wintle BJ, Craig AD (2019) ‘Neptune’, the world’s first messina (Melilotus siculus) cultivar: an annual pasture legume for saline soils prone to winter waterlogging. In ‘Cells to Satellites. Proceedings of the 19th Agronomy Australia Conference’. 25–29 August 2019. Wagga Wagga, NSW. (Ed. J Pratley) http://www.agronomyaustraliaproceedings.org/
NLWRA (2001) Australian dryland salinity assessment 2000. Extent, impacts, processes, monitoring and management options. National Land and Water Resources Audit, Canberra, ACT.
Norman HC, Cocks PS, Smith FP, Nutt BJ (1998) Reproductive strategies in Mediterranean annual clovers: germination and hardseededness. Australian Journal of Agricultural Research 49, 973–982.
| Reproductive strategies in Mediterranean annual clovers: germination and hardseededness.Crossref | GoogleScholarGoogle Scholar |
Norman HC, Cocks PS, Galwey NW (2005) Annual clovers (Trifolium spp.) have different reproductive strategies to achieve persistence in Mediterranean-type climates. Australian Journal of Agricultural Research 56, 33–43.
| Annual clovers (Trifolium spp.) have different reproductive strategies to achieve persistence in Mediterranean-type climates.Crossref | GoogleScholarGoogle Scholar |
Norman HC, Smith FP, Nichols PGH, Si P, Galwey NW (2006) Variation in seed softening patterns and impact of seed production environment on hardseededness in early maturing genotypes of subterranean clover. Australian Journal of Agricultural Research 57, 65–74.
| Variation in seed softening patterns and impact of seed production environment on hardseededness in early maturing genotypes of subterranean clover.Crossref | GoogleScholarGoogle Scholar |
Porqueddu C, Ates S, Louhaichi M, Kyriazopoulos AP, Moreno G, del Pozo A, Ovalle C, Ewing MA, Nichols PGH (2016) Grasslands in ‘Old World’ and ‘New World’ Mediterranean-climate zones: past trends, current status and future research priorities. Grass and Forage Science 71, 1–35.
| Grasslands in ‘Old World’ and ‘New World’ Mediterranean-climate zones: past trends, current status and future research priorities.Crossref | GoogleScholarGoogle Scholar |
Redmann RE (1974) Osmotic and specific ion effects on the germination of alfalfa. Canadian Journal of Botany 52, 803–808.
| Osmotic and specific ion effects on the germination of alfalfa.Crossref | GoogleScholarGoogle Scholar |
Robson AD (1990) The role of self-regenerating pasture in rotation with cereals in Mediterranean areas. In ‘The role of legumes in the farming systems of the Mediterranean areas’. Developments in Plant and Soil Sciences Vol. 38. (Eds AE Osman, MH Ibrahim, MA Jones) pp. 217–236. (Springer)
Rogers ME, Noble CL (1991) The effect of NaCl on the establishment and growth of balansa clover (Trifolium michelianium Savi. var. balansae Boiss.). Australian Journal of Agricultural Research 42, 847–857.
| The effect of NaCl on the establishment and growth of balansa clover (Trifolium michelianium Savi. var. balansae Boiss.).Crossref | GoogleScholarGoogle Scholar |
Rogers ME, Craig AD, Munns RE, Colmer TD, Nichols PGH, Malcolm CV, Barrett-Lennard E, Brown AJ, Semple WS, Evans PM, Cowley K, Hughes SJ, Snowball R, Bennett SJ, Sweeney GC, Dear BS, Ewing MA (2005) The potential for developing fodder plants for the salt-affected areas of southern and eastern Australia: an overview. Australian Journal of Experimental Agriculture 45, 301–329.
| The potential for developing fodder plants for the salt-affected areas of southern and eastern Australia: an overview.Crossref | GoogleScholarGoogle Scholar |
Rogers ME, Colmer TD, Frost K, Henry D, Cornwall D, Hulm E, Deretic J, Hughes SR, Craig AD (2008) Diversity in the genus Melilotus for tolerance to salinity and waterlogging. Plant and Soil 304, 89–101.
| Diversity in the genus Melilotus for tolerance to salinity and waterlogging.Crossref | GoogleScholarGoogle Scholar |
Rogers ME, Colmer TD, Nichols PGH, Hughes SJ, Frost K, Cornwall D, Chandra S, Miller SM, Craig AD (2011) Salinity and waterlogging tolerance amongst accessions of messina (Melilotus siculus). Crop & Pasture Science 62, 225–235.
| Salinity and waterlogging tolerance amongst accessions of messina (Melilotus siculus).Crossref | GoogleScholarGoogle Scholar |
Smetham ML (2003) A review of subterranean clover (Trifolium subterraneum L.): its ecology, and use as a pasture legume in Australasia. Advances in Agronomy 79, 303–350.
| A review of subterranean clover (Trifolium subterraneum L.): its ecology, and use as a pasture legume in Australasia.Crossref | GoogleScholarGoogle Scholar |
Striker GG, Colmer TD (2016) Flooding tolerance of forage legumes. Journal of Experimental Botany 68, 1851–1872.
| Flooding tolerance of forage legumes.Crossref | GoogleScholarGoogle Scholar |
Striker GG, Teakle NL, Colmer TD, Barrett-Lennard EG (2015) Growth responses of Melilotus siculus accessions to combined salinity and root-zone hypoxia are correlated with differences in tissue ion concentrations and not differences in root aeration. Environmental and Experimental Botany 109, 89–98.
| Growth responses of Melilotus siculus accessions to combined salinity and root-zone hypoxia are correlated with differences in tissue ion concentrations and not differences in root aeration.Crossref | GoogleScholarGoogle Scholar |
Taylor GB (2005) Hardseededness in Mediterranean annual pasture legumes in Australia: a review. Australian Journal of Agricultural Research 56, 645–661.
| Hardseededness in Mediterranean annual pasture legumes in Australia: a review.Crossref | GoogleScholarGoogle Scholar |
Teakle NL, Snell A, Real D, Barrett-Lennard EG, Colmer TD (2010) Variation in salinity tolerance, early shoot mass and shoot ion concentrations within Lotus tenuis: towards a perennial pasture legume for saline land. Crop & Pasture Science 61, 379–388.
| Variation in salinity tolerance, early shoot mass and shoot ion concentrations within Lotus tenuis: towards a perennial pasture legume for saline land.Crossref | GoogleScholarGoogle Scholar |
Teakle NL, Armstrong J, Barrett-Lennard EG, Colmer TD (2011) Aerenchymatous phellem in hypocotyl and roots enables O2 transport in Melilotus siculus. New Phytologist 190, 340–350.
| Aerenchymatous phellem in hypocotyl and roots enables O2 transport in Melilotus siculus.Crossref | GoogleScholarGoogle Scholar |
Teakle NL, Bowman S, Barrett-Lennard EG, Real D, Colmer TD (2012) Comparisons of annual pasture legumes in growth, ion regulation and root porosity demonstrate that Melilotus siculus has exceptional tolerance to combinations of salinity and waterlogging. Environmental and Experimental Botany 77, 175–184.
| Comparisons of annual pasture legumes in growth, ion regulation and root porosity demonstrate that Melilotus siculus has exceptional tolerance to combinations of salinity and waterlogging.Crossref | GoogleScholarGoogle Scholar |
Ungar IA (1962) Influence of salinity on seed germination in succulent halophytes. Ecology 43, 763–764.
| Influence of salinity on seed germination in succulent halophytes.Crossref | GoogleScholarGoogle Scholar |
West DW, Taylor JA (1981) Germination and growth of cultivars of Trifolium subterraneum L. in the presence of sodium chloride salinity. Plant and Soil 62, 221–230.
| Germination and growth of cultivars of Trifolium subterraneum L. in the presence of sodium chloride salinity.Crossref | GoogleScholarGoogle Scholar |