

Tagasaste silvopastures in steep-hill country. 2. Effect of increasing proximity to tagasaste on growth and survival of companion pasture species
Katherine Tozer A * , Emma Noakes B , Grant Douglas C , Rose Greenfield A and Catherine Cameron A
A * , Emma Noakes B , Grant Douglas C , Rose Greenfield A and Catherine Cameron A
A AgResearch, Private Bag 3123, Hamilton 3240, New Zealand.
B AgResearch, Private Bag 11008, Palmerston North 4410, New Zealand.
C GBDScience, Church Street, Palmerston North 4410, New Zealand.
Abstract
Tagasaste (Cytisus proliferus), a fast-growing leguminous tree, has potential to supplement pasture production in steep-hill country and to increase pasture resilience.
In the companion paper, we quantified tagasaste production characteristics. Here, we determine the effect of proximity of 10-year-old tagasaste trees on productivity of eight pasture species including grasses (perennial ryegrass, Lolium perenne; cocksfoot, Dactylis glomerata; prairie grass, Bromus willdenowii; microlaena, Microlaena stipoides), perennial legumes (white clover, Trifolium repens; red clover, T. pratense; lotus, Lotus pedunculatus), and an annual legume (subterranean clover, T. subterraneum).
A site was established in the East Coast region of the North Island of New Zealand on steep-hill country (>20° slope). Herbage production, nutritive value and survival of pasture species established as spaced transplants were measured over 3 years.
Cocksfoot had high survival, herbage production and metabolisable energy content but was negatively affected by proximity to tagasaste. Microlaena was not significantly affected by proximity to tagasaste; however, it was much less productive and had lower nutritive values than the other grasses. Only 40% of perennial ryegrass transplants survived 3 years, and survival of perennial legumes was negligible. Subterranean clover was able to set seed in the open and in shade.
Cocksfoot was the most productive grass species, and microlaena was least affected by proximity to tagasaste. Given the poor persistence of perennial clovers, annual clovers may be better suited to a tagasaste silvopasture on steep, dry hillsides.
Mixtures of cocksfoot and subterranean clover may be well suited to summer-dry hillsides, between and under trees in a tagasaste silvopasture. Microlaena may provide some forage and can maintain groundcover despite shade.
Keywords: feed value, forages, forage trees, herbage production, hill country, nutritive values, pasture productivity, tree lucerne, tree-pasture systems.
Introduction
Tagasaste (Cytisus proliferus L. f. var. palmensis Christ) is a fast-growing, leguminous tree that can provide palatable forage of high nutritive value for grazing livestock (Snook 1986; Francisco-Ortega et al. 1991; Dann and Trimmer 2003). It grows in drought-prone sites in well-drained soils but does not tolerate waterlogging. Research into tagasaste establishment and production in annual- and perennial-based pastures has previously been conducted on flat or gently sloping land in Mediterranean, temperate and tropical regions (Oldham and Moore 1989; Townsend and Radcliffe 1990; Douglas et al. 1996; Assefa et al. 2012); however, no data were available on tagasaste productivity when grown on steep-hill country (>20° slope). In the first paper of this two-part series, we reported on tagasaste edible dry matter (DM) production, mineral content and nutritive value when grown in perennial pastures on steep-hill country (Tozer et al. 2023).
Furthermore, there is a lack of understanding of which perennial pasture species are most productive and persistent in temperate hill-country pastures where spaced tagasaste trees have been established for browse forage (hereafter referred to as a ‘tagasaste silvopasture’). Previous studies have demonstrated that shading can reduce the nutritive value of hill-country pastures, although effects were often confounded with changes in botanical composition when comparing open pasture and pasture under the tree canopy (e.g. Guevara-Escobar et al. 2007; Benavides et al. 2009). Studies have also focused on sampling of whole pasture rather than individual species. Research is therefore required to understand the effects of tree shading on the nutritive value of individual pasture species.
Species selected for this study were those considered most suitable for the environment of the experimental hill-country site, based on climatic factors and published research. Where average annual rainfall exceeds 700 mm, species such as perennial ryegrass (Lolium perenne L.) and prairie grass (Bromus willdenowii Kunth), a short-lived perennial, can provide high-quality feed in temperate hill-country pastures (e.g. Suckling 1959; Fulkerson et al. 2000; Waller and Sale 2001). Cocksfoot (Dactylis glomerata L.) provides a productive alternative for drier regions (Lolicato and Rumball 1994). Microlaena (also called weeping grass or rice grass; Microlaena stipoides (Labill.) R.Br.) is indigenous to New Zealand, Australia and the biogeographical region of Malesia (Edgar and Connor 2000; Mitchell et al. 2016). It is recommended as an option for low-input pastures in Australia, although it responds well to applications of nitrogen (Chivers and Aldous 2005).
Companion legumes for these perennial pasture grasses include white clover (Trifolium repens L.), red clover (T. pratense L.) and lotus/greater lotus (Lotus pedunculatus Cav., syn. L. uliginosus) (Orr and Wedderburn 1996; Díaz et al. 2005). Lotus provides an alternative to white clover for acidic, low-fertility soils (Hopkins et al. 1996). In drier areas, the annual legume subterranean clover (Trifolium subterraneum L.) is also widely used (Moss et al. 2022).
We conducted this study to determine how the growth, nutritive value and survival of these pasture species were affected by proximity to 10-year-old tagasaste trees in hill country in the East Coast region of the North Island of New Zealand. We focused on the relative performance of the perennial grass, perennial legume and annual legume species separately, to inform the selection of species for a grass–legume mix. It was hypothesised that increasing proximity to tagasaste would increase shading and decrease productivity (growth, survival and nutritive value) of all pasture species, but that some species would be more severely affected than others by increasing proximity.
Methods
Site establishment and management
Details of location, climate, soils, micro-climatic variables (e.g. soil moisture, soil surface temperature, solar radiation), characteristics of the tagasaste trees (e.g. height, canopy spread), annual DM production and botanical composition of resident pasture, and site management are presented in Part 1 of this two-part series (Tozer et al. 2023). Briefly, the field site was on a typical beef and sheep hill-country farm in the Eastern Coast region of the North Island of New Zealand (39°01′11″S, 177°34′17.69″E). The area has a temperate maritime climate with average maximum temperatures (°C) in the mid-20s in summer (December–February) and average minimum temperature of ~6°C in winter (June–August), when most of the rain falls. The 10-year average annual rainfall (2005–14) is 1510 mm. Summer droughts occurred in 2019–20 and 2020–21. The soils are Orthic raw soils according to the New Zealand Soil Classification system (Hewitt 2010).
The field site was on a north-west-facing hillside, with an average slope of 30°, and comprised a 10-year-old tagasaste silvopasture with 156 trees ha−1. When measured in November 2018, tagasaste trees had an average (± s.e.m., n = 7) height of 4.3 ± 0.2 m, canopy width of 6.1 ± 0.4 m, trunk diameter (at the first branch) of 24.3 ± 0.9 cm and root-collar diameter of 25.7 ± 4.1 cm, with an average of 7.1 ± 0.9 branches >2.5 cm diameter.
Experimental design and treatments
The experimental design has been detailed in Part 1. Briefly, the study was established as a split-plot, randomised complete block design with seven replicates of three shade treatments (heavy shade, light shade, open pasture) positioned across the hill within the tagasaste plantation. Shade treatments were the main plots. Heavy shade plots were established within 1 m of a tagasaste trunk; light shade plots 1–2 m from a tagasaste trunk; and open pasture plots were positioned between trees and not directly under the canopy of a tagasaste tree. However, the open pasture treatment could not be regarded as uninfluenced by adjacent tagasaste trees, above or below the ground.
Within each main plot, nine pasture species treatments and an unsown, bare-ground treatment were randomly allocated to subplots (30 cm by 30 cm), which were arranged in two adjacent rows of five perpendicular to the hillslope. In the first year, the nine pasture species treatments comprised three perennial grass species, three perennial legume species and three subterranean clover cultivars. In the second year, a poorly performing subterranean clover cultivar was replaced with a prairie grass treatment. The bare-ground treatment enabled us to determine the composition of the seedbank and is reported in the companion paper. Pasture species were selected based on their use in New Zealand hill country and their ability to tolerate moderate levels of shading and drought (Table 1).
| Common name | Cultivar(s) | Botanical name | Characteristics considered beneficial for use in hill country in a tagasaste silvopasture | |
|---|---|---|---|---|
| Perennial ryegrass | Grasslands Samson | Lolium perenne L. | Widely sown species (grass control) (Waller and Sale 2001) | |
| Cocksfoot | Savvy | Dactylis glomerata L. | Shade and drought tolerant (Devkota et al. 1998; Sanada et al. 2010) | |
| Microlaena | Wakefield | Microlaena stipoides (Labill.) R.Br. | Shade and drought tolerant (Firth et al. 2002; Mitchell et al. 2016) | |
| Prairie grassA | Atom | Bromus willdenowii Kunth | Shade tolerant and productive under lax grazing (Burtt-Davy 1916; Stewart 1996) | |
| White clover | Grasslands Nomad | Trifolium repens L. | Widely sown species (legume control) | |
| Red clover | Grasslands Relish | T. pratense L. | Shade tolerant (Van Sambeek et al. 2007; St. John 2008) | |
| Subterranean clover | Antas, Denmark | T. subterraneum L. | Drought tolerant (Devkota et al. 1997; Widdup and Pennell 2000; Smetham 2003) | |
| Lotus/greater lotus | Grasslands Trojan | Lotus pedunculatus Cav. (syn. L. uliginosus) | Shade tolerant (Devkota et al. 1997) |
APrairie grass was established in the second year.
On 14 March 2018, 50 bare seeds of each pasture species were sown into Hillson root trainers (3.8 cm by 3.6 cm by 12 cm) containing a commercial potting mix. The root trainers were then placed in a glasshouse with an average day temperature of 18.5°C and night temperature of 11.9°C. Seedlings were watered with Thrive All Purpose Soluble Fertiliser (1 g L−1 applied to 28 plants; Yates, Wyee, NSW, Australia) at 2-week intervals from April to August 2018, when they were transplanted into the field site. Because microlaena has a low germination rate (<25%), plants were purchased from a commercial retailer, subdivided to match the size of other test grass species (~3 tillers plant−1) and transplanted into Hillson root trainers.
At the field site, branches and debris were removed from the plot locations, and Roundup (Bayer CropScience, Melbourne, Vic., Australia) was applied to existing vegetation in each plot and its surrounds (1.8 m by 1.8 m area) at a rate of 10 mL Roundup L−1 water, using a knapsack sprayer, in early August 2018.
On 22 and 23 August, 2 weeks after herbicide application, the soil was cultivated with a shovel to a depth of 15 cm, the seedlings were transplanted into the centre of each subplot, and the soil was firmly pressed around each plant. Each transplant was irrigated with ~700 mL water and identified with a plastic label. On 23 August after transplanting, slug bait was applied to each subplot (25 g, Eden Gardener Snail Bait containing metaldehyde at 5 g kg−1).
On 26 September and 9 October 2018, any dead transplants were replaced and slug bait re-applied at the same rate. The number of tillers (or stolons) on each plant was counted on 26 September 2018. Signs of a viral disease were apparent on Campeda subterranean clover (red colouration of leaves) and became more severe over successive months. Campeda was eliminated from the dataset because there were concerns that any effects of shade treatments would be confounded with disease effects. There was no sign of disease in the other cultivars (Antas and Denmark).
In September 2018, ~5 weeks after seedlings were transplanted, the number of tillers (or stolons) per plant was similar for all three shade treatments (average across species 11 plant−1). Of the grasses, number of tillers per plant was highest for perennial ryegrass (29 tillers), intermediate for microlaena (21 tillers) and lowest for cocksfoot (14 tillers) (± 1.3, standard error of difference; P < 0.001). There was no difference among white clover, red clover and lotus for number of stolons per plant (average 9 stolons), or between Antas and Denmark subterranean clover (average 6 stolons) (P > 0.05).
Antas and Denmark subterranean clover were re-established in 2019 to provide 2 years of consecutive data. Because it was not possible to obtain data on Campeda in the first year, the biennial species prairie grass was substituted as an alternative pasture species given its suitability for hill country (Table 1). Antas and Denmark subterranean clover and Atom prairie grass seeds were sown in seed-raising mix in Hillson root trainers on 17 April 2019 as described above. In July 2019, subterranean clover seedlings (Antas 5.8 ± 0.6 stolons plant−1 and Denmark 5.4 ± 0.5 stolons plant−1) were transplanted into the same subplots as in 2018, and prairie grass seedlings (2.0 ± 0.3 tillers plant−1, n = 8) were transplanted into the former Campeda treatment subplots. Slug bait was applied as above.
Livestock were excluded from the tagasaste silvopasture during establishment between August and October 2018. The plantation was grazed approximately every 6 weeks between October 2018 and June 2021 with Coopworth ewes and lambs or hoggets, at an average stocking rate of 254 ± 31 stock units ha−1. Within 24 h before grazing, plots were covered with a plastic mesh sheet (with 1-cm diameter cells) pinned to the ground. This prevented stock from grazing and trampling the test plants while still allowing sunlight to reach the plants and allowing stock access to the surrounding pasture. The mesh sheet was removed within 48 h after stock were removed from the plantation. Further details on grazing management are provided in the companion paper.
Measurements
Measurements were taken at intervals of 4–7 weeks from 22 August 2018 to 9 June 2021. There were four measurements of the pasture species in 2018 (August–December), seven in 2019, eight in 2020 and four in 2021 (January–June). Measurement frequency declined over summer when pasture growth was slower, and there were fewer visits than envisaged throughout autumn 2020 owing to COVID-19-related travel restrictions. Measurements were completed within 48 h.
Pasture species survival, herbage production, herbage quality, canopy cover (plant area), plant damage and soil moisture content were assessed on each measurement occasion as follows.
To measure survival, any plants with green tissue were recorded as surviving and those with no green tissue were recorded as dead. Plants classified as dead occasionally had green tissue present on subsequent measurement dates, in which case they were reclassified as surviving.
For assessment of herbage production and quality, herbage was removed at the top of the pseudostem (typically ~5 cm above ground level) by using scissors. Retaining the pseudostem is thought to enable satisfactory replenishment of carbohydrate reserves for several pasture species (e.g. perennial ryegrass, cocksfoot, prairie grass) and minimise defoliation stress (Turner et al. 2007). The herbage from each plant was quickly frozen in dry ice. Samples were stored at −20°C, freeze-dried (FD80 freeze dryer; Cuddon, Blenheim, NZ), weighed, and ground to a powder of particle size <1 mm (Cyclone sample mill; UDY, Fort Collins, CO, USA). Ground samples were analysed by Hill Laboratories (Hamilton, NZ) using near-infrared spectroscopy (Corson et al. 1999) to obtain estimates of metabolisable energy (ME), crude protein, soluble sugars, neutral detergent fibre (NDF) and ash. Dry weights were summed for each species to obtain seasonal and total annual herbage production.
Canopy cover of each plant was visually assessed in a 30 cm by 30 cm quadrat divided into 5 cm by 5cm cells. The total number of cells occupied by each plant was multiplied by the cell area. A cell was considered empty if <50% filled with vegetation, and occupied if >50% filled with vegetation (Hayes et al. 2021).
Plant damage was quantified visually, assigning a score of 1 to a plant that was vigorous and healthy with no disease/invertebrate damage; a score of 2 to a plant with disease/invertebrate damage and up to 50% of tillers/stolons dying; and a score of 3 to a plant with >50% of tillers/stolons dying and the whole plant with disease/invertebrate damage.
Subterranean clover seedling emergence from the seedbank was quantified in autumn 2019. All subterranean clover seedlings that emerged from the Antas and Denmark subterranean clover treatments were counted and removed. These data were collected to determine whether the Antas and Denmark transplants established in 2018 were able to produce viable seed.
Soil moisture content was measured as described in Tozer et al. (2023). In summary, soil moisture content was measured at a depth of 0–12 cm using a portable time domain reflectometry instrument (Hydrosense II; Campbell Scientific, Logan, UT, USA), calibrated for the field site using gravimetric soil moisture values.
Statistical analyses
All data other than survival were analysed by split-plot analysis of variance (ANOVA) using GenStat 21st edition (GenStat 2021). Treatments used in the analyses were three shade levels (heavy, light, open), species (or cultivars in the case of the annual legume), and the shade × species interaction. Pasture species were classed as perennial grasses, perennial legumes and annual legumes and were analysed separately because of high mortality rates in perennial and annual legumes and because there was no interest in comparing grass species with legume cultivars. Survival data were analysed as a generalised linear mixed model with binomial distribution, with shade and species/cultivar as fixed effects, and site and shade within site as random effects. A repeated-measures split-plot ANOVA was performed for herbage quality variables for grasses in the second and third year and damage score in the first year. In all cases, there were significant interactions between time and fixed treatments (P < 0.001). Other repeated-measures analyses failed to converge owing to sparse data and inconsistent sampling (e.g. some replicates were eliminated from the analyses if plants had died). Consequently, times were analysed individually. This demonstrated how differences between means changed over time without formally measuring those patterns. Residuals from the analyses were checked for normality. Herbage production was square-root transformed to equalise the variance to meet the normality assumptions of the analysis. Mean separation was assessed by Fisher’s protected least significant difference.
Results
Survival
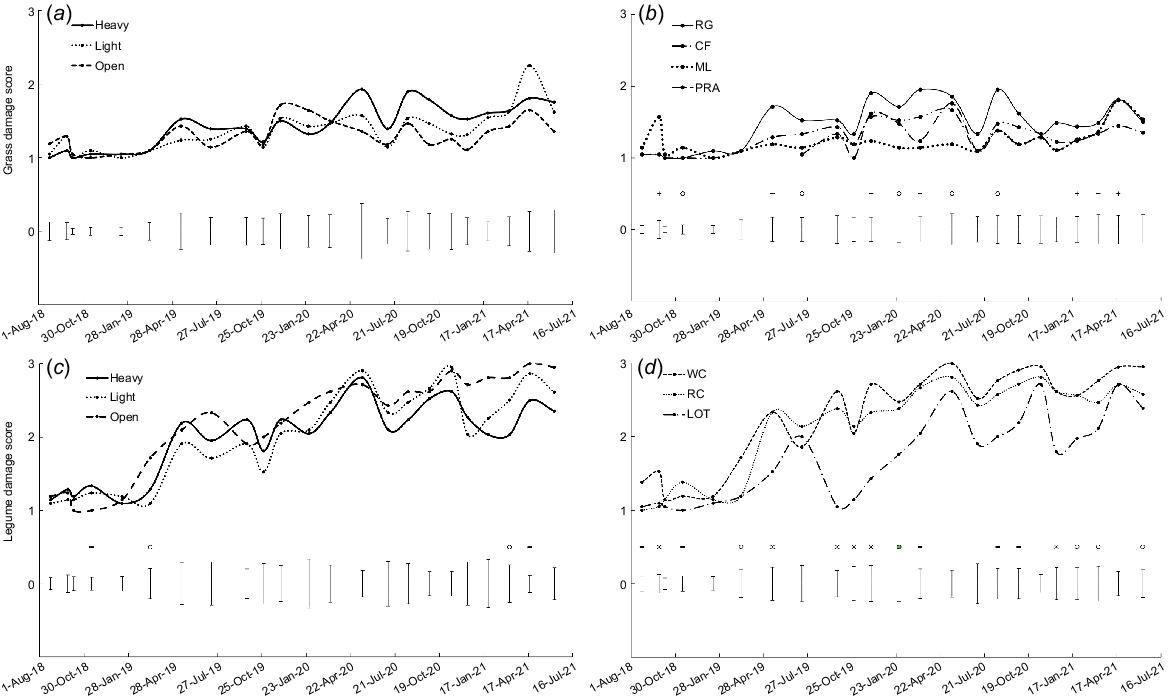
Survival was lower in light shade than in heavy shade and open pasture in April 2021 (P < 0.05, Fig. 1a). Survival of grass transplants remained high in the first year (>90%), but by the end of the study only 20% of transplants survived in light shade and an average of 85% in the heavy shade and open pasture treatments.
Transplant survival (as evidenced by the presence of green tissue) for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. On some occasions, plants appeared to be dead (i.e. no green tissue present) but later recovered. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.
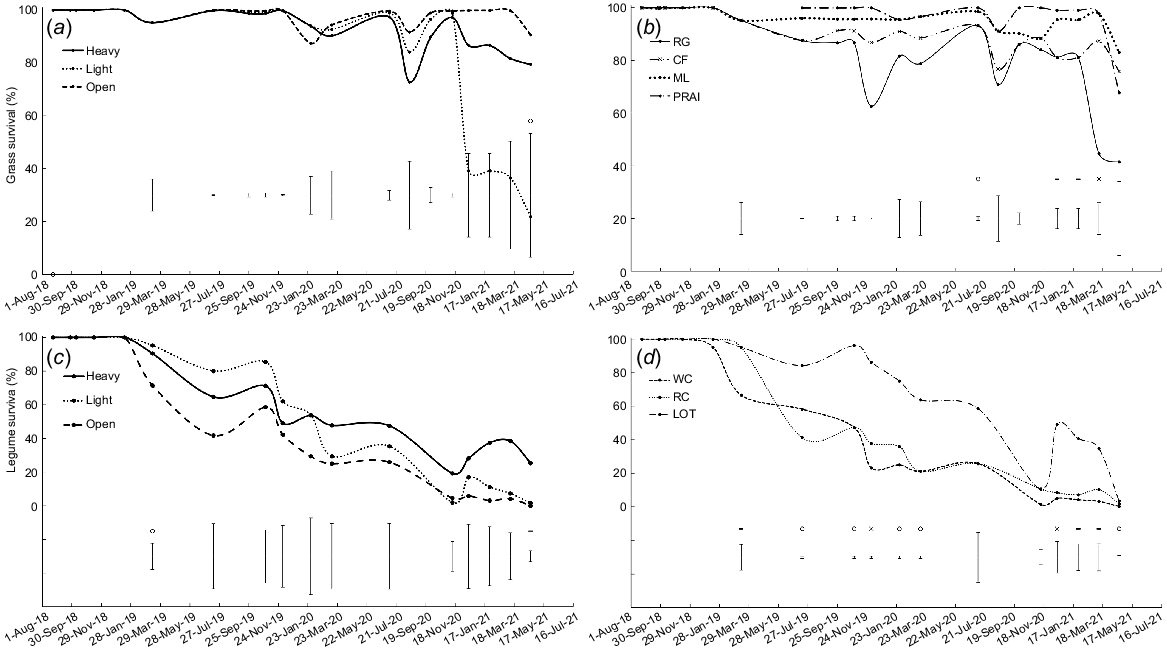
In July 2020, survival was higher for microlaena and prairie grass than for perennial ryegrass and cocksfoot (P < 0.05, Fig. 1b). From midsummer 2020 onwards, survival was generally highest for microlaena and prairie grass and lowest for perennial ryegrass, with cocksfoot intermediate (P < 0.01 in December, January and March; P = 0.051 in April). Approximately 80% of microlaena but only 40% of perennial ryegrass transplants were alive in April 2021 (Fig. 1b).
In March 2019, legume survival was higher in the light and heavy shade treatments than in open pasture (P < 0.05, Fig. 1c). By April 2021, survival was higher in heavy shade (25%) than in light shade (2%) and open pasture, where no legumes survived (P < 0.01, Fig. 1c).
There were significant differences among legume species in survival on 10 occasions, when survival was generally higher for lotus than for white clover and red clover (Fig. 1d, P < 0.05). By April 2021, only 4% of lotus, 2% of red clover and <1% of white clover transplants were alive.
Herbage production
Shading (heavy and/or light shade) reduced total and seasonal herbage production of the grasses during most seasons and all years compared with open pasture (P < 0.05, Table 2). Herbage production in open pasture was often >3-fold greater than in heavy shade, with production in light shade intermediate but not always significantly different from either heavy shade or open pasture.
| Season | Perennial grasses (g DM plant−1) | Perennial legumes (g DM plant−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heavy | Light | Open | s.e.d. | P-value | Heavy | Light | Open | s.e.d. | P-value | |||
| 2018–19 | Spring | 5.3a | 11.5b | 13.9b | 2.55 | 0.016 | 4.1a | 8.5a | 16.0b | 2.24 | <0.001 | |
| Summer | 12.9a | 35.5b | 40.8b | 10.68 | 0.049 | 8.7a | 21.2a | 42.4b | 8.18 | <0.001 | ||
| Autumn | 2.9 | 6.5 | 6.8 | 1.80 | 0.084 | 0.5 | 0.9 | 2.3 | 1.69 | n.s. | ||
| Total annualA | 20.9a | 53.4b | 61.3b | 14.71 | 0.038B | 13.3a | 30.6b | 62.4c | 9.01 | <0.001B | ||
| 2019–20 | Winter | 0.7a | 1.2a | 5.9b | 1.16 | 0.004 | 0.2 | 0.2 | 0.4 | 0.17 | n.s. | |
| Spring | 4.9a | 8.4a | 31.9b | 6.60 | 0.006 | 7.7 | 7.6 | 8.8 | 1.70 | n.s. | ||
| Summer | 8.8a | 15.3ab | 28.0b | 6.33 | 0.041 | 9.5 | 13.8 | 15.5 | 4.03 | n.s. | ||
| Autumn | 1.3a | 4.6ab | 11.9b | 3.78 | 0.025 | 0.7 | 1.0 | 1.9 | 1.08 | n.s. | ||
| Total annual | 15.7a | 29.6a | 77.7b | 16.80 | 0.013 | 18.1 | 22.6 | 26.6 | 5.72 | n.s. | ||
| 2020–21 | Winter | 2.7a | 4.8a | 14.4b | 3.81 | 0.028 | 0.5 | 1.1 | 1.6 | 0.85 | n.s. | |
| Spring | 6.4a | 10.6a | 25.0b | 5.83 | 0.022 | 1.7 | 1.3 | 3.0 | 1.86 | n.s. | ||
| Summer | 6.1a | 7.9ab | 16.2b | 4.51 | 0.043 | 4.5 | 3.3 | 2.9 | 1.85 | n.s. | ||
| Autumn | 7.2 | 7.0 | 14.2 | 3.87 | n.s. | 2.2 | 1.5 | 0.8 | 0.79 | n.s. | ||
| Total annual | 22.3a | 30.2a | 69.7b | 16.85 | 0.024 | 8.9 | 7.1 | 8.2 | 4.80 | n.s. | ||
Data were analysed separately for the grass species and legume species.
s.e.d., standard error of the difference; n.s., not significant (P > 0.05).
Within rows and pasture type (grasses or legumes), means followed by the same letter (or no letter) are not significantly different (P > 0.05). Raw means and s.e.d.s are presented, with the P-value and mean separation from square-root transformed data analysis.
ATotal annual herbage production was for only three seasons in the first year.
BSignificant shade × species treatment interaction (shade × species combination means are provided in Tables 4 and 5).
Cocksfoot had the greatest seasonal and total annual herbage production of the perennial grasses from summer 2018–19 onwards for all seasons and years (Table 3). Cocksfoot was more productive than microlaena in the first year, and its DM production was ~3-fold greater than that of perennial ryegrass and microlaena in the second year and 2-fold greater in the third year. Perennial ryegrass was also more productive than microlaena in 2018–19 (P < 0.001), whereas from winter 2019 onward, DM production was similar for the two species (P > 0.05, Table 3). Total annual herbage production of (2-year-old) prairie grass was greater than that of (3-year-old) perennial ryegrass transplants in the final year of the study (P < 0.05, Table 3).
| Season | Perennial grasses (g DM plant−1) | Perennial legumes (g DM plant−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RG | CF | ML | PRA | s.e.d. | P-value | WC | RC | LOT | s.e.d. | P-value | |||
| 2018–19 | Spring | 16c | 13b | 2a | n/a | 1.4 | <0.001 | 5a | 12b | 11b | 1.4 | <0.001 | |
| Summer | 29b | 49c | 11a | n/a | 6.0 | <0.001 | 8a | 31b | 34b | 6.5 | <0.001 | ||
| Autumn | 4a | 9b | 4a | n/a | 1.2 | <0.001 | 0a | 1a | 4b | 0.7 | <0.001 | ||
| Total annual | 49b | 71c | 17a | n/a | 7.3 | <0.001A | 14a | 44b | 49b | 7.6 | <0.001A | ||
| 2019–20 | Winter | 3b | 6c | 1a | <1a | 1.0 | <0.001 | <1 | <1 | <1 | 0.19 | n.s. | |
| Spring | 12a | 31b | 6a | 12a | 4.8 | <0.001 | 4a | 6a | 14b | 3.0 | 0.010 | ||
| Summer | 8a | 28c | 13ab | 21bc | 4.9 | <0.001 | 8 | 12 | 20 | 5.6 | n.s. | ||
| Autumn | 2a | 12c | 4a | 5ab | 2.1 | <0.001 | 1 | 1 | 2 | 1.2 | n.s. | ||
| Total annual | 25a | 78b | 24a | 37a | 11.5 | <0.001 | 13a | 19ab | 36b | 8.8 | 0.039 | ||
| 2020–21 | Winter | 5a | 12b | 5a | 7ab | 2.3 | 0.033 | <1 | <1 | 2 | 1.0 | n.s. | |
| Spring | 9a | 19b | 8a | 19b | 3.2 | <0.001 | 1 | <1 | 4 | 1.9 | n.s. | ||
| Summer | 6a | 15b | 8a | 11ab | 2.6 | 0.008 | 3 | 3 | 5 | 2.0 | n.s. | ||
| Autumn | 6a | 14b | 8ab | 11b | 3.0 | 0.022 | 1 | 2 | 2 | 0.9 | n.s. | ||
| Total annual | 26a | 59b | 30a | 48b | 9.3 | 0.002 | 6 | 6 | 12 | 5.0 | n.s. | ||
Data are averaged over shade treatments for perennial grasses and for perennial legumes.
s.e.d., standard error of the difference; n.s., not significant (P > 0.05); n/a, not applicable.
Within rows and pasture type (grasses or legumes), means followed by the same letter (or no letter) are not significantly different (P > 0.05). Raw means and s.e.d.s are presented, with the P-value and mean separation from square-root transformed data analysis.
ASignificant shade × species treatment interaction (shade × species combination means are provided in Tables 4 and 5).
There was a significant shade × grass species interaction in the first year (P < 0.05, Table 4) which demonstrated that herbage production of perennial ryegrass and cocksfoot was strongly suppressed by heavy shade, whereas microlaena was unaffected by shading, although as noted, it had significantly lower production than cocksfoot and ryegrass.
| Parameter | Season/year | Heavy | Light | Open | s.e.d. | Interaction P-value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RG | CF | ML | PRA | RG | CF | ML | PRA | RG | CF | ML | PRA | |||||
| Herbage production (g DM plant−1) | 2018–19 (sqrt) | 4.6a | 5.6a | 2.6 | 7.3b | 9.2b | 3.7 | 7.9b | 8.8b | 4.8 | 1.16 | 0.035 | ||||
| Canopy cover (cm2 plant−1) | Average 2018–19 | 35a | 59a | 20 | 59b | 99b | 26 | 52b | 70a | 31 | 7.4 | 0.011 | ||||
| Average 2019–20 | 46 | 104a | 44 | 54 | 75 | 134a | 50 | 85 | 75 | 189b | 72 | 70 | 23.1 | 0.037 | ||
| Plant damage score (1–3) | Autumn 2021 | 2.2b | 1.9 | 1.1 | 1.5 | 2.6b | 1.4 | 1.6 | 1.4 | 1.5a | 1.4 | 1.3 | 1.6 | 0.33 | 0.025 | |
| ME (MJ kg DM−1) | Summer 2018–19 | 9.3ab | 9.2 | 9.9b | 8.8a | 9.6 | 9.3b | 9.8b | 9.3 | 8.4a | 0.39 | 0.015 | ||||
| Soluble sugars (% of DM) | Winter 2019 | 5a | 5 | 6 | 6a | 6a | 5 | 6 | 8ab | 12b | 6 | 7 | 10b | 1.2 | 0.044 | |
| Autumn 2021 | 8a | 6 | 8 | 9a | 10b | 6 | 8 | 8a | 12c | 5 | 9 | 11b | 0.8 | 0.036 | ||
| Annual average 2020–21 | 9a | 6 | 7 | 8a | 9a | 6 | 8 | 9a | 11b | 6 | 8 | 11b | 0.6 | 0.023 | ||
| NDF (% of DM) | Average 2018–19 | 47b | 52b | 49 | n/a | 48b | 50ab | 49 | n/a | 43a | 48a | 50 | n/a | 1.1 | 0.011 | |
| Summer 2019–20 | 45 | 52ab | 50a | 56 | 47 | 51a | 53a | 54 | 47 | 55b | 57b | 54 | 1.8 | 0.004 | ||
ME, metabolisable energy; NDF, neutral detergent fibre; RG, perennial ryegrass; CF, cocksfoot; ML, microlaena; PRA, prairie grass; s.e.d., standard error of difference; n/a, not applicable.
Within rows, means followed by the same letter (or no letter) are not significantly different (P > 0.05) when comparing the same species across the three shade treatments. Prairie grass was not present in 2018–19.
Herbage production in spring and summer, and total annual herbage production, were greater in open pasture than in heavy and light shade in 2018–19 (P < 0.05, Table 2).
Herbage production in spring 2018, summer 2018–2019 and autumn 2019, and total annual herbage production in 2018–19, were greater for lotus and, often, red clover than for white clover (P < 0.001, Table 3). Total annual herbage production in 2019–20 was also greater for lotus than white clover, with red clover intermediate but not significantly different from either of the other species (P < 0.05, Table 3).
There was a significant shade × legume species interaction for total annual herbage production (Table 5). Compared with open pasture, total annual herbage production of white clover was not affected by shading, whereas herbage production of red clover and lotus was suppressed by light and heavy shade (P < 0.01, Table 5). From the second year onwards, legumes performed poorly in all treatments and there were no significant interactions.
| Parameter | Season/year | Heavy | Light | Open | s.e.d. | Interaction P-value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WC | RC | LOT | WC | RC | LOT | WC | RC | LOT | |||||
| Herbage production (g DM plant−1) | 2018–19 (sqrt) | 2.3 | 3.4a | 3.8a | 4.1 | 4.8a | 6.0b | 3.7 | 9.0b | 8.9c | 0.87 | 0.003 | |
| Canopy cover (cm2 plant−1) | Autumn 2019 | 21 | 14 | 43a | 30 | 15 | 61ab | <1 | 32 | 79b | 14.7 | 0.039 | |
| Plant damage score (1–3) | Average 2018–19 | 1.5ab | 1.5b | 1.2 | 1.3a | 1.4ab | 1.2 | 1.7b | 1.2a | 1.2 | 0.11 | 0.002 | |
| ME (MJ kg DM−1) | Autumn 2019 | 11.3b | 10.2 | 9.8a | 11.1ab | 11.1 | 10.1ab | 10.2a | 10.2 | 11.0b | 0.49 | 0.043 | |
| Soluble sugars (% of DM) | Autumn 2019 | 6 | 7 | 1a | 6 | 8 | 2a | 5 | 8 | 6b | 0.8 | 0.004 | |
| NDF (% of DM) | Average 2018–19 | 32 | 38 | 38b | 34 | 35 | 37b | 33 | 35 | 31a | 1.7 | 0.031 | |
| Ash (% of DM) | Autumn 2019 | 15b | 12a | 13 | 13ab | 12a | 11 | 10a | 17b | 10 | 1.9 | 0.045 | |
ME, metabolisable energy; NDF, neutral detergent fibre; WC, white clover; RC, red clover; LOT, lotus; s.e.d., standard error of difference.
Within rows, means followed by the same letter (or no letter) are not significantly different (P > 0.05) when comparing the same species across the three shade treatments.
Nutritive values
In January 2020, ME content was lower in open pasture than in heavy shade, with light shade intermediate (P < 0.05, Fig. 2a). There were differences among perennial grasses on 16 occasions (P < 0.05) with an overall trend of ME content being highest for perennial ryegrass, similar and sometimes lower for cocksfoot, and lowest for microlaena and prairie grass (Fig. 2b).
Metabolisable energy (ME) content for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.

In summer 2018–19, compared with open pasture, light shading reduced the ME content of perennial ryegrass and increased that of microlaena, but there was no effect of shade treatment on cocksfoot ME content (shade × grass species interaction P < 0.05, Table 4).
There were differences among shade treatments on three occasions (P < 0.05), although there was no consistent trend (Fig. 2c). ME content was generally higher in white clover and red clover than in lotus, with differences being significant on 13 occasions (P < 0.05, Fig. 2d).
In autumn 2019, compared with open pasture, heavy shading increased the ME content of white clover and reduced that of lotus, but there was no effect of shade treatment on red clover ME content (shade × legume species interaction P < 0.05, Table 5).
Crude protein content was higher in heavy shade than in open pasture, and intermediate in light shade, with significant differences occurring on eight occasions (P < 0.05, Fig. 3a). There were significant differences among grass species on 12 occasions, but trends were inconsistent (P < 0.05, Fig. 3b).
Crude protein content (% of DM) for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.
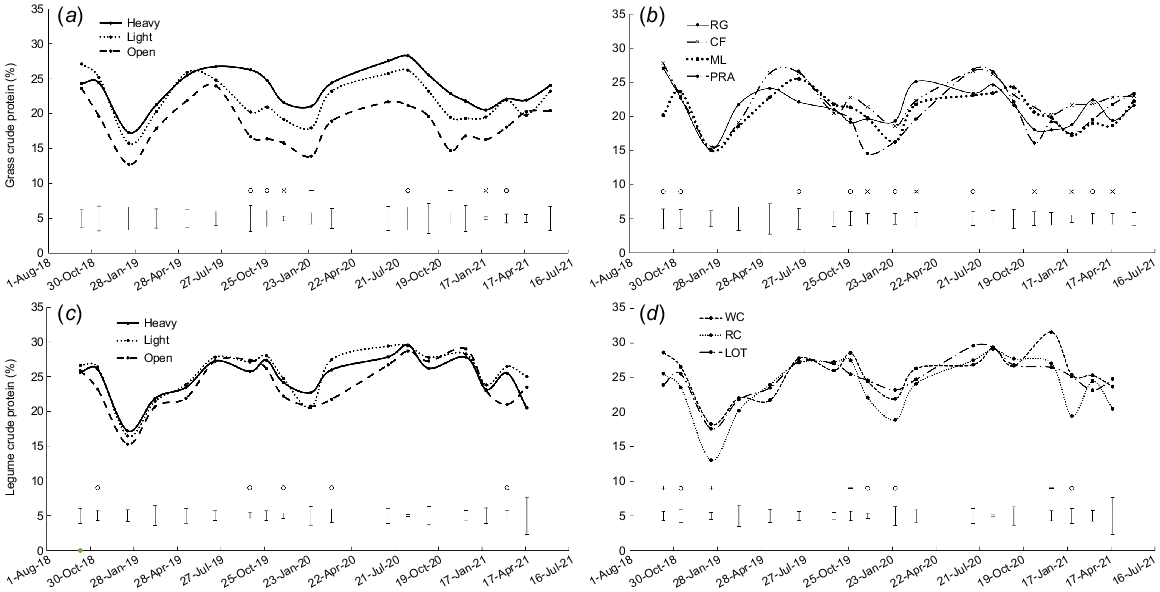
There were differences in crude protein content among the shade treatments on five occasions and among legume species on eight occasions (P < 0.05, Fig. 3c, d). Crude protein content was generally higher in heavy shade and/or light shade than in open pasture and higher in white clover and/or lotus than red clover.
There were differences in soluble sugar content among shade treatments on five occasions (P < 0.05, Fig. 4a) and among grass species on 15 occasions (P < 0.05, Fig. 4b). Soluble sugar content was higher in open pasture than in heavy shade, with light shade intermediate (Fig. 4a). Soluble sugar content was often lowest for prairie grass and fluctuated for the other species (Fig. 4b).
Soluble sugar content (% of DM) for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. o, P < 0.05; -, P < 0.01; x, P < 0.001.
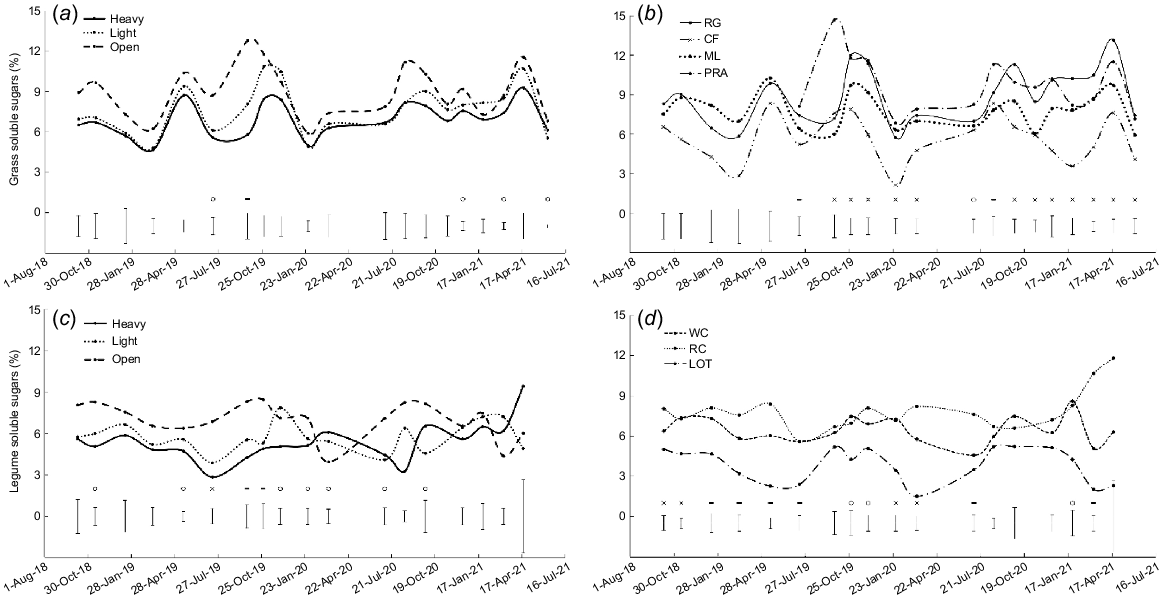
In winter 2019 and autumn 2021, and on average for 2020–21, shading reduced the soluble sugar content in perennial ryegrass and prairie grass but there was no effect of shade treatment in cocksfoot or microlaena (shade × grass species interaction P < 0.05, Table 4).
There were differences in soluble sugar content among shade treatments on 10 occasions (P < 0.05, Fig. 4c) and among legume species on 13 occasions (P < 0.05, Fig. 4d). Generally, the soluble sugar content was greater in open pasture than heavy and light shade. It was also higher in red clover than in lotus, with white clover intermediate (Fig. 4d).
In autumn 2019, there was no effect of shade treatment on white clover or red clover but shading severely reduced the soluble sugar content in lotus (average of 1% in heavy and light shade vs 6% in open pasture; shade × legume species interaction P < 0.01, Table 5).
There was no effect of shade treatment on NDF content (P > 0.05, Fig. 5a). There were differences among grass species on 19 occasions, with perennial ryegrass generally having a lower NDF content than the other three grass species (P < 0.05, Fig. 5b).
Neutral detergent fibre content (NDF, % of DM) for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.
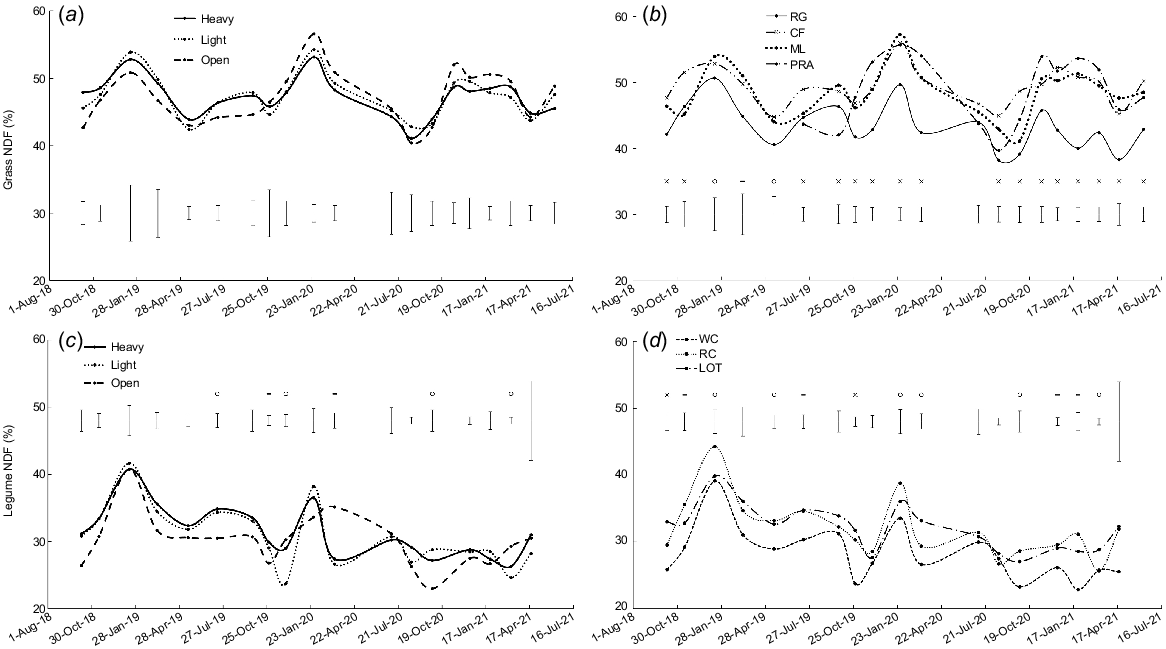
In spring 2018 (data not shown), and for the average annual values for 2018–19 (Table 4), compared with open pasture, shading (heavy and/or light shade) increased the NDF content of perennial ryegrass and cocksfoot but there was no effect of shade treatment on microlaena (shade × grass species interaction, P < 0.05). In summer 2019–20, the trends were reversed: shading reduced the NDF content of cocksfoot and microlaena but there was no effect of shade treatment on perennial ryegrass or prairie grass (shade × grass species interaction, P < 0.01, Table 4).
Compared with open pasture, shading increased the NDF content in winter or spring (July 2019, October 2019, September 2020; P < 0.05). The reverse occurred when sampled in summer or autumn, when NDF was lower in heavy and/or light shade than open pasture (December 2019, March 2020, March 2021; P < 0.05, Fig. 5c). There were differences between perennial legumes on 12 occasions, with a general trend of higher NDF content in red clover and lotus than white clover (P < 0.05, Fig. 5d).
In 2018–19, compared with open pasture, shading increased the average annual NDF content in lotus but there was no effect of shade treatment on white clover or red clover (shade × legume species interaction P < 0.05, Table 5).
Ash content was greater in heavy shade than open pasture on four occasions, with the ash content in light shade often intermediate (P < 0.05, Fig. 6a). There were differences between grass species on 16 occasions, when the ash content was most often highest for microlaena, intermediate for perennial ryegrass and cocksfoot, and lowest for prairie grass (P < 0.05, Fig. 6b). There was no shade × grass species interaction for ash content (P > 0.05).
Ash content (% of DM) for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.

Ash content of legumes generally followed a similar trend when shaded to that of the grasses, with a higher content in heavy shade than open pasture on four occasions (P < 0.05, Fig. 6c). Ash content was greater in white clover than in red clover and lotus on 10 occasions (P < 0.05, Fig. 6d).
In autumn 2019, compared with open pasture, heavy shade increased the ash content of white clover, and heavy and light shade reduced the ash content of red clover, but there was no effect of shade treatment on the ash content of lotus (shade × legume species interaction P < 0.05, Table 5).
Canopy cover
Canopy cover was greater in open pasture than heavy shade on four occasions and greater in open pasture than light shade on two occasions (P < 0.05, Fig. 7a).
Canopy cover per plant for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.

There were significant differences in canopy cover among grass species for all measurement dates (P < 0.05, Fig. 7b). By January 2019, canopy cover was nearly always higher for cocksfoot than perennial ryegrass and microlaena, with prairie grass generally intermediate (P < 0.01, Fig. 7b).
There were shade × grass species interactions in spring 2018 (P < 0.01, data not shown) and summer 2018–19 (P < 0.01, data not shown) and on average for 2018–19 (P < 0.05, Table 4). There was a consistent trend of greater canopy cover for perennial ryegrass and cocksfoot in light shade than heavy shade, with open pasture being intermediate and not always significantly different from either, whereas for microlaena, canopy cover was not affected by shade treatment (P < 0.05, Table 4). By spring 2019, there was no longer an advantage of light shade; canopy cover of cocksfoot was greater in open pasture than in light shade and heavy shade but there was no effect of shade treatment on any other grass species (P < 0.05, data not shown). The same trend was observed for the average annual canopy cover in 2019–20 (P < 0.05, Table 4).
Canopy cover was greater in open pasture than in one or both shade treatments on five occasions (P < 0.05, Fig. 7c). There were significant differences in canopy cover among perennial legumes on 13 occasions (Fig. 7d, P < 0.05). Canopy cover was generally greater for lotus than for white clover and red clover up to January 2020 (P < 0.05), after which time, trends were less consistent. In January and March 2021, canopy cover was greater for white clover than red clover and lotus (P < 0.05), but by June 2021, there was no difference in canopy cover among the three species (average 26 cm2, P > 0.05, Fig. 7d).
In autumn 2019, canopy cover of lotus was lower in heavy shade than open pasture, whereas white clover and red clover were not affected by shade treatment (shade × legume interaction P < 0.05, Table 5).
Plant damage
There was no effect of shade on plant damage on any occasion (P > 0.05, Fig. 8a).
Plant damage caused by invertebrates or disease for: (a) the three shade treatments averaged over grass species; (b) perennial ryegrass (RG), cocksfoot (CF), microlaena (ML) and prairie grass (PRA) averaged over shade treatments; (c) the three shade treatments averaged over legume species; and (d) white clover (WC), red clover (RC) and lotus (LOT) averaged over shade treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01; x, P < 0.001.
There were significant differences in plant damage among grass species on 12 occasions (P < 0.05, Fig. 8b) On eight of these occasions, plant damage was greatest in ryegrass and least for microlaena, with the other two species intermediate (P < 0.05, Fig. 8b). Damage levels remained <2 (up to 50% of tillers/stolons dying) for all grass species for the study duration.
In autumn 2021, perennial ryegrass sustained more damage in both light shade and heavy shade than in open pasture, but levels of damage were similar across shade treatments for cocksfoot, microlaena and prairie grass (shade × grass species interaction P < 0.05, Table 4).
There was an effect of shade on plant damage in legumes on four occasions (P < 0.05, Fig. 8c), with damage being greater in open pasture than in one or both shade treatments on three of the four occasions. There were significant differences in plant damage among legume species on 16 occasions (P < 0.05, Fig. 8d). White clover sustained the most damage and lotus the least, with red clover intermediate but not always significantly different from either of the other species (P < 0.05, Fig. 8d). The damage score was >2 for white clover and red clover for most of the second and third years, whereas the score fluctuated for lotus.
In 2018–19, compared with open pasture, light shade reduced damage to white clover and heavy shade increased damage to red clover, but lotus was unaffected by shade treatment (shade × legume species interaction P < 0.01, Table 5).
Subterranean clover growth and survival
There was no difference between cultivars on any of the measurement occasions (P > 0.05, Fig. 9a).

There was no effect of shade treatment on subterranean clover seedling populations that emerged from the seedbank beneath the Denmark or Antas planting positions in autumn 2019, which averaged 32 seedlings m−2 in March 2019 and 52 seedlings m−2 in May 2019. In addition, there was no difference in seedling emergence between Antas and Denmark subterranean clover in autumn 2019 (P > 0.05, data not shown).
Total dry weight and average annual canopy cover of subterranean clover were greater in open pasture than in heavy or light shade in 2019–20 (P < 0.001, Table 6). Canopy cover was also greater for Denmark than for Antas in September and October 2018 (P < 0.001, Fig. 9b). There was no difference between the cultivars for damage score on any of the measurement dates (Fig. 9c). There was a trend towards greater damage in heavy and light shade than in open pasture in 2019–20, although differences were not significant (P = 0.053, Table 6).
| Measurement | Shade treatments | Cultivar treatments | Interaction P-value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heavy | Light | Open | s.e.d. | P-value | ANT | DMK | s.e.d. | P-value | ||||
| 2018–19 | Total dry weight (g DM plant−1) | 2.0 | 5.4 | 5.8 | 1.88 | n.s. | 3.5 | 5.3 | 1.58 | n.s. | n.s. | |
| Average canopy cover (cm2 plant−1) | 1.1 | 1.4 | 1.5 | 0.24 | n.s. | 1.2 | 1.5 | 0.25 | n.s. | n.s. | ||
| Average damage score (1–3) | 2.2 | 2.1 | 2.2 | 0.08 | n.s. | 2.1 | 2.2 | 0.06 | n.s. | n.s. | ||
| 2019–20 | Total DM (g DM plant−1) | 3.6a | 6.6a | 25.7b | 3.55 | <0.001 | 12.0 | 12.0 | 3.65 | n.s. | n.s. | |
| Average canopy cover (cm2 plant−1) | 2.9a | 2.8a | 8.1b | 0.71 | <0.001 | 4.9 | 4.3 | 0.72 | n.s. | n.s. | ||
| Average damage score (1–3) | 2.2b | 2.2b | 2.0a | 0.08 | 0.053 | 2.1 | 2.1 | 0.06 | n.s. | n.s. | ||
Means of shade treatments are averaged over cultivars and means of cultivars are averaged over shade treatments.
s.e.d., standard error of the difference; n.s., not significant.
Within rows and shade or cultivar treatments, means followed by the same letter (or no letter) are not significantly different (P > 0.05). Interaction P-value is for the shade treatment × cultivar interaction.
There was no difference between cultivars, and no shade × cultivar interaction, for total dry weight, average canopy cover or average damage score in either 2018–19 or 2019–20 (P > 0.05, Table 6). In the first and second years, there was no effect of shade or cultivar on the average content of ME, crude protein, NDF or ash (data not presented, P > 0.05). Soluble sugar content was lower in heavy and light shade than in open pasture (average of 9.8% vs 31.2%, P < 0.01) in the first year and greater for Denmark than Antas in the second year (10.7% vs 8.8%, P < 0.01).
There were no shade × cultivar interactions for any of the nutritive value measurements in either year (P > 0.05).
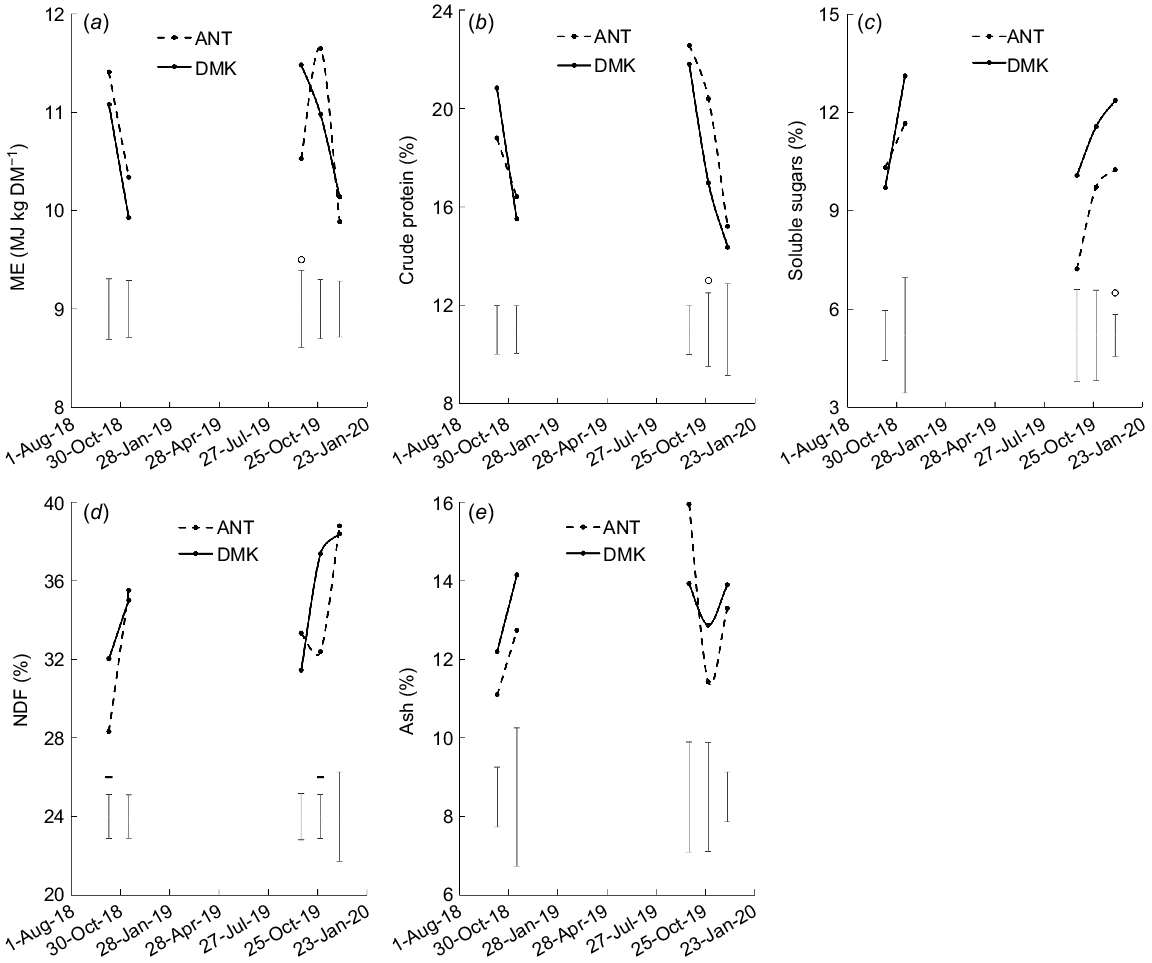
Denmark had higher contents than Antas of ME in September 2019 (P < 0.05, Fig. 10a), soluble sugars in December 2019 (P < 0.01, Fig. 10c), and NDF in October 2018 and 2019 (P < 0.01, Fig. 10d), and a lower content of crude protein in October 2019 (P < 0.05, Fig. 10b). There was no difference between Antas and Denmark in ash content (P > 0.05, Fig. 10e).
(a) Metabolisable energy (ME), (b) crude protein (% of DM), (c) soluble sugars (% of DM), (d) neutral detergent fibre (NDF, % of DM), and (e) ash (% of DM) of subterranean clover cvv. Antas (ANT) and Denmark (DMK) averaged over the heavy shade, light shade and open pasture treatments. Error bars represent the standard error of the difference for each measurement date. o, P < 0.05; -, P < 0.01.
Soil moisture
There was no difference in average soil moisture content among the grasses or among annual or perennial legumes other than in January 2019 when soil moisture content was greater for microlaena plots than cocksfoot plots (9% vs 7%, P < 0.05). There were no shade × species interactions (P > 0.05).
Discussion
Performance of grass species
There were large differences among shade treatments in levels of solar and photosynthetic radiation throughout the study, but fewer differences among the shade treatments in soil moisture or soil nutrient content (Tozer et al. 2023). This suggests that the effects of increasing proximity to tagasaste on plant growth and survival were strongly associated with increased shading, although competition for other resources cannot be ruled out. On this basis, we have focused on species performance in relation to shade, although other factors such as moisture stress are discussed to a lesser extent.
There was minimal impact of proximity to tagasaste on perennial grass survival, except when survival decreased in the light shade treatment towards the end of the study. However, increasing proximity to tagasaste severely reduced herbage production, canopy cover and the content of soluble sugars, increased the contents of crude protein and ash, and had negligible effect on plant damage, ME and NDF. As hypothesised, there were differences in how the species responded to the shade treatments as demonstrated by the interactions.
Cocksfoot was the most productive of the grass species and has been recommended for dry hill country where perennial ryegrass fails to persist (Lolicato and Rumball 1994). Survival was high, with >75% of transplants surviving for the duration of the study. Although its production was suppressed under shade more than some of the other species such as prairie grass, it was still the most productive species in terms of both quantity and quality, with a high content of ME and crude protein when shaded.
Peeters (2004) noted that cocksfoot ‘tolerates shade very well, for instance in old orchards’ (p. 129). It was abundant in the resident pasture at the field site. Others have also found cocksfoot to be tolerant of shade under controlled conditions, although effects are cultivar-dependent (Lin et al. 1998). By contrast, Grime et al. (2007) found in their survey of British flora that cocksfoot was much more common in unshaded environments, disturbed habitats, wastelands and grasslands than in woods and plantations and shady environments. Douglas et al. (2006) recommended cocksfoot for use in lightly shaded and lightly grazed poplar (Populus spp.) silvopastures, where it established and grew well. The ability of cocksfoot to tolerate shade may be moderated by the defoliation regime; cocksfoot swards can deteriorate under heavy grazing and are sensitive to trampling (Peeters 2004). Possibly, the defoliation regime for the spaced plants in our study was not optimal for cocksfoot and reduced its productivity, particularly when heavily shaded by tagasaste.
Perennial ryegrass can grow in a wide range of microsites in New Zealand hill country owing to its large genotypic and phenotypic variation (Wedderburn and Pengelly 1991). Perennial ryegrass was not well adapted to the field site, as demonstrated by the strong decrease in survival by the end of the measurement period, when only 40% of transplants survived, and its absence in the resident pasture.
Perennial ryegrass growth was suppressed when near the tagasaste trees in the heavy and light shade treatments. This was demonstrated by a reduction in perennial ryegrass herbage production, canopy cover, ME content and soluble sugar content, and an increase in plant damage and NDF content with shading. In their floristic survey of Britain, Grime et al. (2007) found that perennial ryegrass was absent in shaded environments such as woodlands, plantations and hedgerows and negligible where the soil pH was <5. Peeters (2004) also noted that perennial ryegrass is not tolerant of shade, is sensitive to drought and is best adapted to mild and wet climates. At this field site, there was shade, low soil pH (averaging 5.3), drought for three consecutive summers (comparing the average monthly rainfall with the long-term monthly rainfall in fig. 1 of Tozer et al. 2023), and consequently low soil moisture content, which decreased to ~15% during summer. If rainfall had approached the long-term average, the outcomes for ryegrass may have been considerably different.
Although prairie grass was assessed for only 2 years, its survival had decreased to 70% by the end of the study and was similar to that of the 3-year-old cocksfoot. Webby et al. (1990) also found that prairie grass failed to persist for more than three growing seasons in summer-wet hill country of the Upper North Island of New Zealand. Prairie grass was more tolerant of shading than perennial ryegrass and cocksfoot. Its herbage production, canopy cover, plant damage and NDF content were unaffected by proximity to tagasaste, although the content of soluble sugars declined. Prairie grass is heat- and drought-tolerant (Peeters 2004) and can be productive over summer in hill country of the North Island of New Zealand, given appropriate grazing management (Sithamparanathan 1979). It is also more tolerant of shade than perennial ryegrass owing to its more upright growth habit (Langer 1970).
Microlaena was the grass most tolerant of shading, as demonstrated by the lack of significant shade-treatment effects on herbage production, canopy cover, plant damage, and contents of ME, soluble sugar, NDF and ash. In pastures in the Northern Tablelands of New South Wales, Australia, microlaena is prevalent in swards under shade trees frequented by resting livestock, near the edges of sheep camps and around tree stumps (Whalley et al. 1978; Magcalemacandog and Whalley 1991), indicating that it is shade-tolerant and can withstand intensive trampling. It also forms pure swards under trees in New Zealand (Smith 2005). Microlaena has a rhizome a few centimetres below ground, which can enable it to recover after close grazing and drought (Mitchell 2013; Mitchell et al. 2016). The ability of microlaena to produce low levels of forage in heavy shade and withstand trampling confers some advantages over cocksfoot for dry, shady areas, although its lack of herbage production and low nutritive value compared with perennial ryegrass and cocksfoot limit its usefulness as a pasture species. Microlaena is present in North Island hill-country pastures such as those where this study was conducted (e.g. Smale et al. 1997) and was the most abundant species in both the shade and open resident pasture treatments. If the environment is not suitable for perennial ryegrass, cocksfoot or prairie grass, such as in dry areas subject to heavy shade, microlaena could enable the maintenance of groundcover, despite summer droughts and stock trampling. Microlaena lines have been selected for grazing in Australia, but it is expensive to purchase, it cannot be established using standard drilling methods, and its establishment is sporadic over an extended period, which leaves it vulnerable to weed ingress (Whalley and Jones 1997). The most realistic option is to manage existing swards of microlaena in New Zealand hill country to maintain groundcover where more desirable pasture species are unable to survive.
The prevalence of microlaena in the resident pasture as described in Tozer et al. (2023), and its ability to produce a similar yield in the open and in light shade, would also explain why the yield of resident pasture was similar in open pasture and light shade. If tagasaste had been more widely spaced so that there was less shading, and the resident pasture had been dominated by other species such as perennial ryegrass, there may have been a much greater decline in pasture production with increasing proximity to tagasaste.
Performance of perennial legume species
Legume survival was low regardless of the shade treatment; legumes were much less resilient than the grasses and were highly susceptible to disease and insect damage. Poor survival of test legume species in this study is consistent with the absence of legumes in the resident pasture.
The lack of persistence of legumes in pastures has been attributed to selective grazing by livestock and competition with grasses (Haynes 1980; Hoveland 1989). In this study, stock were excluded from the plots and the plants were spaced to reduce competition with neighbouring pasture species. Other factors such as moisture-deficit stress, heat stress, shading, pest invertebrates and disease pressures were the more likely causes leading to their demise (Hoveland 1989; Benavides et al. 2009). Although it is not known which invertebrate pests were present, clover root weevil (Sitona lepidus Gyllenh) is abundant in New Zealand and causes severe damage to pasture legumes (Zydenbos et al. 2011).
Shading reduced legume herbage production severely in the first year, after which production was low for all legumes regardless of the shade treatment. However, there were differences in how the legumes responded to shade.
During the first 18 months, lotus was the most productive of the legumes with the highest survival, but its productivity was severely suppressed by shading compared with red clover and white clover. This was demonstrated by a reduction in herbage production, canopy cover, and contents of ME and soluble sugars with increased proximity to tagasaste, and an increase in the content of NDF. Lotus is best suited to damp, acidic, low-fertility soils with infrequent defoliation (Sheath 1980; White 1995; Grime et al. 2007). Although lotus was found to be more tolerant of shading than white clover or subterranean clover in a pot study when moisture was not limiting (Devkota et al. 1997), its absence from woodlands and plantations in a survey of British flora (Grime et al. 2007) suggests that its shade tolerance is low. The combination of frequent defoliation and low soil moisture content, particularly under the tagasaste canopy, is a likely reason for its lack of persistence in this study.
White clover was largely unaffected by proximity to tagasaste. This may reflect that it was the most unproductive of the perennial legumes with high levels of plant damage, such that shading had little additional impact. Given the multiple summer droughts that occurred and low soil moisture content, low productivity of white clover is to be expected. White clover is typically associated with higher rainfall, summer-wet pastures in New Zealand, and its persistence is compromised by drought (Knowles et al. 2003). Grime et al. (2007) also found that white clover was not tolerant of drought, and was not present in woodland or plantation habitats in Britain. Severe shading (by ~80% of full sunlight) reduced white clover productivity by up to 93% in mixtures of perennial ryegrass and white clover under controlled conditions simulating different shading patterns of trees (Ehret et al. 2015). White clover was also found to be less tolerant than lotus of shading (Devkota et al. 1997). Possibly, shade in our study ameliorated drought stresses experienced by white clover, because plant damage was lower under light shade than in open pasture in the first year, and the contents of ME and ash were higher under heavy shade than open pasture in autumn 2019.
Red clover produced more herbage than white clover. Suckling (1960) also found red clover to be more productive than white clover in summer-moist hill country in the Central North Island. However, its productivity was reduced by increased proximity to tagasaste and there was an increase in plant damage and a reduction in the ash content as shading increased. The ability of red clover to tolerate shading depends on temperature and moisture-deficit stress (Gist and Mott 1957), both of which occurred during the study. Grime et al. (2007) found that it was abundant in moist environments, absent from woodlands and not tolerant of shade.
Subterranean clover test cultivar performance
Subterranean clover cultivars vary in the extent to which they can tolerate shade (Mauromicale et al. 2010). In our study, the two cultivars tested were similarly affected by shade, which reduced their productivity (i.e. herbage production and canopy cover) and increased visual signs of damage in the second year. Mauromicale et al. (2010) found that cultivars which established and grew rapidly were more able to tolerate shade than those with a slower growth pattern. The two cultivars in our study had similar growth patterns, so it is unlikely that differences in shade tolerance would be detected based on their growth habit.
Species mixtures for a tagasaste silvopasture
A tagasaste silvopasture may require periods of infrequent and lax grazing to limit overgrazing of the more slowly regenerating tagasaste. This may enhance the persistence of legumes in a tagasaste silvopasture. White clover, red clover and lotus can benefit from infrequent and lax defoliation during their reproductive period and during drought to enable seed production and protect the carbohydrate reserves. Subterranean clover must also be grazed lightly to enable the seed production necessary for its continued regeneration (Sheath 1980; Brock et al. 2003; Stevens et al. 2020; Moss et al. 2022). Given the lack of persistence of legumes in our study, we suggest that a less frequent and/or less severe defoliation regime may be required to improve legume persistence.
Perennial ryegrass, cocksfoot and prairie grass can also benefit from grazing exclusion during their reproductive period in late spring–summer. This enables reseeding and stimulates regrowth tillering when conditions are suitable for growth in autumn (Hume et al. 1990; Dowling et al. 1996; Tozer et al. 2021). In our study, all species were defoliated at the same time. However, each species has different functional traits that necessitate different defoliation management strategies to maximise their production and persistence (Duru et al. 2005). Grazing frequency and intensity would therefore need to be tailored for the sown pasture species. Further research is required to develop grazing guidelines for a tagasaste silvopasture to ensure persistence and productivity of both the pasture and woody components.
Based on our results, there are several possibilities for understorey mixtures that require further investigation. Cocksfoot–subterranean clover mixtures have been regularly sown in dryland environments in New Zealand where summer droughts are recurrent and the inclusion of subterranean clover can lead to significant increases in pasture yield (Ates et al. 2010). Subterranean clover can be oversown (e.g. White et al. 1972). Hard grazing or herbicide application prior to oversowing is critical to reduce competition with resident pasture species and allow establishment (Tozer and Douglas 2016). Alternatively, hill country pastures can also be managed to increase the content of subterranean clover when drilling is not possible (Olykan et al. 2019). Microlaena can also grow well with cocksfoot (Magcalemacandog and Whalley 1991) and could occupy the shadier areas in a tagasaste silvopasture to which cocksfoot is less suited. The intended use of the mixtures is also to be considered. In this study, we have focused on characteristics such as herbage production and nutritive values for improving livestock production. If other ecosystem services are required (e.g. pollination or erosion control), other characteristics such as flowering duration and root length should be considered (Hanisch et al. 2020).
An alternative strategy could be to introduce lotus when establishing a tagasaste silvopasture, which is fenced to exclude livestock for several years during the tagasaste establishment period. This may enable lotus to establish and ensure its persistence, even in drought-prone areas. In the central northern area of the North Island of New Zealand, lotus comprised >50% of total DM in Years 3–5 on oversown, drought-prone, low-fertility landslips that had been fenced for the first 2 years to exclude livestock (Lambert et al. 1993). To avoid the high levels of lotus mortality that occurred in our study, grazing post-establishment would need to be infrequent (e.g. three or four times each year) and competition with grasses would also need to be managed (Charlton and Brock 1980).
Conclusions
Cocksfoot shows potential for sowing on summer-dry hillsides between and under trees in a tagasaste silvopasture. In close proximity to tagasaste, where growth of all species is strongly suppressed, microlaena may provide limited, low-quality forage, but may assist in maintaining pasture cover, which has environmental benefits.
Lotus could be oversown when establishing a tagasaste plantation in which grazing is excluded during the first few years. However, it may be difficult for perennial legumes to persist beyond several years in a tagasaste silvopasture subject to dry summers with high temperatures, especially when growing near tagasaste. In these environments, subterranean clover may be a more useful companion legume for cocksfoot.
Spaced-plant studies are useful for screening of species, but larger scale studies are required before establishment and management guidelines for a tagasaste silvopasture can be developed. We recommend that: (1) research is undertaken on establishing cocksfoot–subterranean clover mixes in tagasaste silvopastures; (2) strategies be developed to increase microlaena abundance in the shadier areas; (3) lotus establishment strategies are developed for summer-dry hill country in new plantings of tagasaste from which stock are excluded; (4) the research is expanded to different environments; and (5) the value proposition for such a system in hill country is developed.
Acknowledgements
The leadership and involvement of experienced East Coast farmers has been invaluable. Thanks to Agricom for providing the cocksfoot, perennial ryegrass, white clover, red clover and lotus, Lusietta seeds for providing the subterranean clover cultivars, and Oratai Nurseries for providing the microlaena. Thanks to Tony Craven, Caitlyn Dawbin, Craig Traill, Elizabeth North, Tracy Dale, Wai Aparau for assistance with field work and laboratory analyses. Thanks also to Peter Manson (Hawke’s Bay Regional Council) for his expert advice throughout the study, Ian Tarbotton for providing strategic direction, and Jim Crush (AgResearch), and Cara Brosnahan and Mhairi Sutherland (Beef + Lamb New Zealand) for their constructive comments on the manuscript.
References
Assefa G, Peters KJ, Kijora C, Minta M (2012) Field performance of tagasaste (Chamaecytisus palmensis) under different harvesting management in a tropical highland area of Ethiopia. Ethiopian Journal of Agricultural Sciences 22, 143-158.
| Google Scholar |
Ates S, Tongel MO, Moot DJ (2010) Annual herbage production increased 40% when subterranean clover was over-drilled into grass dominant dryland pastures. Proceedings of the New Zealand Grassland Association 72, 3-10.
| Crossref | Google Scholar |
Benavides R, Douglas GB, Osoro K (2009) Silvopastoralism in New Zealand: review of effects of evergreen and deciduous trees on pasture dynamics. Agroforestry Systems 76, 327-350.
| Crossref | Google Scholar |
Brock JL, Hyslop MG, Widdup KH (2003) A review of red and white clovers in the dryland environment. NZGA: Research and Practice Series 11, 101-107.
| Crossref | Google Scholar |
Burtt-Davy J (1916) Economic grasses. The Agricultural Journal of South Africa 3, 15-16.
| Google Scholar |
Charlton JFL, Brock JL (1980) Establishment of Lotus pedunculatus and Trifolium repens in newly developed hill country. New Zealand Journal of Experimental Agriculture 8, 243-248.
| Crossref | Google Scholar |
Chivers IH, Aldous DE (2005) Reproductive and vegetative responses of different accessions of Microlaena stipoides (Labill.) R.Br. to nitrogen applications and supplementary irrigation in southern Australia. The Rangeland Journal 27, 151-157.
| Crossref | Google Scholar |
Corson DC, Waghorn GC, Ulyatt MJ, Lee J (1999) NIRS: Forage analysis and livestock feeding. Proceedings of the New Zealand Grassland Association 61, 127-132.
| Crossref | Google Scholar |
Devkota NR, Kemp PD, Hodgson J (1997) Screening pasture species for shade tolerance. Proceedings of the Agronomy Society of New Zealand 27, 119-128.
| Google Scholar |
Devkota NR, Kemp PD, Valentine I, Hodgson J (1998) Performance of perennial ryegrass and cocksfoot cultivars under tree shade. Proceedings of the Agronomy Society of New Zealand 28, 129-135.
| Google Scholar |
Douglas GB, Bulloch BT, Foote AG (1996) Cutting management of willows (Salix spp.) and leguminous shrubs for forage during summer. New Zealand Journal of Agricultural Research 39, 175-184.
| Crossref | Google Scholar |
Douglas GB, Walcroft AS, Hurst SE, Potter JF, Foote AG, Fung LE, Edwards WRN, van den Dijssel C (2006) Interactions between widely spaced young poplars (Populus spp.) and introduced pasture mixtures. Agroforestry Systems 66, 165-178.
| Crossref | Google Scholar |
Dowling PM, Kemp DR, Michalk DL, Klein TA, Millar GD (1996) Perennial grass response to seasonal rests in naturalised pastures of central New South Wales. The Rangeland Journal 18, 309-326.
| Crossref | Google Scholar |
Duru M, Tallowin JRB, Cruz P (2005) Functional diversity in low-input grassland farming systems: characterisation, effect and management. Agronomy Research 3, 125-138.
| Google Scholar |
Ehret M, Graß R, Wachendorf M (2015) The effect of shade and shade material on white clover/perennial ryegrass mixtures for temperate agroforestry systems. Agroforestry Systems 89, 557-570.
| Crossref | Google Scholar |
Firth DJ, Jones RM, McFadyen LM, Cook BG, Whalley RDB (2002) Selection of pasture species for groundcover suited to shade in mature macadamia orchards in subtropical Australia. Tropical Grasslands 36, 1-12.
| Google Scholar |
Francisco-Ortega J, Fernandez-Galvan M, Santos-Guerra A (1991) A literature survey (1696–1991) on the fodder shrubs tagasaste and escobon (Chamaecytisus proliferus (L.fil) Link sensu lato) (Fabaceae: Genisteae). New Zealand Journal of Agricultural Research 34, 471-488.
| Crossref | Google Scholar |
Fulkerson WJ, Fennell JFM, Slack K (2000) Production and forage quality of prairie grass (Bromus willdenowii) in comparison to perennial ryegrass (Lolium perenne) and tall fescue (Festuca arundinacea) in subtropical dairy pastures. Australian Journal of Experimental Agriculture 40, 1059-1068.
| Crossref | Google Scholar |
Gist GR, Mott GO (1957) Some effects of light intensity, temperature, and soil moisture on the growth of alfalfa, red clover and birdsfoot trefoil seedlings. Agronomy Journal 49, 33-36.
| Crossref | Google Scholar |
Guevara-Escobar A, Kemp PD, Mackay AD, Hodgson J (2007) Pasture production and composition under poplar in a hill environment in New Zealand. Agroforestry Systems 69, 199-213.
| Crossref | Google Scholar |
Hanisch M, Schweiger O, Cord AF, Volk M, Knapp S (2020) Plant functional traits shape multiple ecosystem services, their trade-offs and synergies in grasslands. Journal of Applied Ecology 57, 1535-1550.
| Crossref | Google Scholar |
Hayes RC, Newell MT, Pembleton KG, Peoples MB, Li GD (2021) Sowing configuration affects competition and persistence of lucerne (Medicago sativa) in mixed pasture swards. Crop & Pasture Science 72, 707-722.
| Crossref | Google Scholar |
Haynes RJ (1980) Competitive aspects of the grass-legume association. Advances in Agronomy 33, 227-261.
| Crossref | Google Scholar |
Hopkins A, Martyn TM, Johnson RH, Sheldrick RD, Lavender RH (1996) Forage production by two Lotus species as influenced by companion grass species. Grass and Forage Science 51, 343-349.
| Crossref | Google Scholar |
Hoveland CS (1989) Legume persistence under grazing in stressful environments of the United States. In ‘Persistence of forage legumes’. (Eds GC Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 375–385. (American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA)
Hume DE, Falloon RE, Hickson RE (1990) Productivity and persistence of prairie grass (Bromus willdenowii Kunth) 2. Effects of natural reseeding. New Zealand Journal of Agricultural Research 33, 395-403.
| Crossref | Google Scholar |
Knowles I, Fraser T, Daly M (2003) White clover: loss in drought and subsequent recovery. NZGA: Research and Practice Series 11, 37-41.
| Crossref | Google Scholar |
Lambert MG, Trustrum NA, Costall DA, Foote AG (1993) Revegetation of erosion scars in Wairarapa hill country. Proceedings of the New Zealand Grassland Association 55, 177-181.
| Crossref | Google Scholar |
Langer RHM (1970) Growth of prairie grass (Bromus unioloides H.B.K.) in different temperatures and light intensities. In ‘Proceedings of the XI International Grassland Congress’. Surfers Paradise, Qld, Australia. (Ed. NJT Norman) pp. 502–506. (University of Queensland Press: St. Lucia, Qld, Australia)
Lin CH, McGraw RL, George MF, Garrett HE (1998) Shade effects on forage crops with potential in temperate agroforestry practices. Agroforestry Systems 44, 109-119.
| Crossref | Google Scholar |
Lolicato S, Rumball W (1994) Past and present improvement of cocksfoot (Dactylis glomerata L.) in Australia and New Zealand. New Zealand Journal of Agricultural Research 37, 379-390.
| Crossref | Google Scholar |
Magcalemacandog DB, Whalley RDB (1991) Distribution of Microlaena stipoides and its association with introduced perennial grasses in a permanent pasture on the Northern Tablelands of New South Wales. Australian Journal of Botany 39, 295-303.
| Crossref | Google Scholar |
Mauromicale G, Occhipinti A, Mauro RP (2010) Selection of shade-adapted subterranean clover species for cover cropping in orchards. Agronomy for Sustainable Development 30, 473-480.
| Crossref | Google Scholar |
Mitchell ML, Virgona JM, Jacobs JL, Kemp DR (2016) Summer drought survival and recovery in Microlaena stipoides. The Rangeland Journal 38, 501-510.
| Crossref | Google Scholar |
Moss WM, Nichols PGH, Foster KJ, Ryan MH, Erskine W, Guzzomi AL (2022) A century of subclover: lessons for sustainable intensification from a historical review of innovations in subterranean clover seed production. Advances in Agronomy 171, 305-339.
| Crossref | Google Scholar |
Oldham CM, Moore PM (1989) Tagasaste Chamaecytisus palmensis an evergreen fodder tree, in grazing systems of Mediterranean type climates; 1. Feed value for reproduction when grazed by merino ewes at joining. In ‘Proceedings of the XVI International Grasslands Congress’. Nice, France. (Ed. R Desroches) pp. 1251–1252. (Association Française pour la Production Fourragère)
Olykan ST, Lucas RJ, Nicholson DJ, Doscher C, Moot DJ (2019) Maximising the subterranean clover content on a summer-dry Wairarapa hill-country farm through grazing management. Journal of New Zealand Grasslands 81, 91-100.
| Crossref | Google Scholar |
Orr SJ, Wedderburn ME (1996) Assessing the persistence of some pasture legumes in hill country. Proceedings of the New Zealand Grassland Association 58, 259-264.
| Crossref | Google Scholar |
Sheath GW (1980) Effects of season and defoliation on the growth habit of Lotus pedunculatus Cav. cv. ‘Grasslands Maku’. New Zealand Journal of Agricultural Research 23, 191-200.
| Crossref | Google Scholar |
Sithamparanathan J (1979) I. Temperate grasses. New Zealand Journal of Experimental Agriculture 7, 157-162.
| Crossref | Google Scholar |
Smale MC, McLeod M, Smale PN (1997) Vegetation and soil recovery on shallow landslide scars in tertiary hill country, East Cape region, New Zealand. New Zealand Journal of Ecology 21, 31-41.
| Google Scholar |
Smetham ML (2003) Subterranean clover (Trifolium subterraneum): its history and current and future research in New Zealand. NZGA: Research and Practice Series 11, 61-72.
| Crossref | Google Scholar |
Smith PH (2005) The suitability of a native grass species, Microlaena stipoides, for use as an amenity turfgrass in the Auckland region. In ‘Greening the city: bringing biodiversity back into the urban environment. Proceedings of Convention on Biological Diversity’. 21–24 October 2003, Christchurch, New Zealand. (Ed. M Dawson) (Royal New Zealand Institute of Horticulture)
Stevens DR, Garden JP, Garden N, Casey MJ (2020) Can Lotus pedunculatus over-sowing in low-fertility tussock country increase farm resilience? Journal of New Zealand Grasslands 82, 171-181.
| Crossref | Google Scholar |
Stewart AV (1996) Potential value of some Bromus species of the section Ceratochloa. New Zealand Journal of Agricultural Research 39, 611-618.
| Crossref | Google Scholar |
Suckling FET (1959) Pasture management trials on unploughable hill country at te Awa. New Zealand Journal of Agricultural Research 2, 488-543.
| Crossref | Google Scholar |
Suckling FET (1960) Productivity of pasture species on Hill Country. New Zealand Journal of Agricultural Research 3, 579-591.
| Crossref | Google Scholar |
Townsend RJ, Radcliffe JE (1990) Tagasaste forage production systems. New Zealand Journal of Agricultural Research 33, 627-634.
| Crossref | Google Scholar |
Tozer K, Douglas G (2016) Pasture establishment on non-cultivable hill country: a review of the New Zealand literature. NZGA: Research and Practice Series 16, 213-224.
| Crossref | Google Scholar |
Tozer K, Müller K, Craven A, Cameron C (2021) Grazing strategies for resilience of ryegrass (Lolium perenne) dominant pastures in hill country. Crop & Pasture Science 72, 947-968.
| Crossref | Google Scholar |
Tozer K, Douglas G, Noakes E, Greenfield R, Cameron C (2023) Tagasaste silvopastures in steep-hill country. 1. Tagasaste edible dry-matter production and nutritive value. Crop & Pasture Science
| Crossref | Google Scholar |
Turner LR, Donaghy DJ, Lane PA, Rawnsley RP (2007) Distribution of water-soluble carbohydrate reserves in the stubble of prairie grass and orchardgrass plants. Agronomy Journal 99, 591-594.
| Crossref | Google Scholar |
Van Sambeek JW, Navarrete-Tindall NE, Garrett HE, Lin CH, McGraw ML, Wallace DC (2007) Ranking the shade tolerance of forty-five candidate groundcovers for agroforestry plantings. The Temperate Agroforester 15(4), 1-10.
| Google Scholar |
Waller RA, Sale PWG (2001) Persistence and productivity of perennial ryegrass in sheep pastures in south-western Victoria: a review. Australian Journal of Experimental Agriculture 41, 117-144.
| Crossref | Google Scholar |
Webby RW, Sheath GW, Boom CJ (1990) Performance of new pasture cultivars in a hill country finishing system. Proceedings of the New Zealand Grassland Association 51, 151-156.
| Crossref | Google Scholar |
Wedderburn ME, Pengelly WJ (1991) Resident ryegrass in hill country pastures. Proceedings of the New Zealand Grassland Association 53, 91-95.
| Crossref | Google Scholar |
Whalley RDB, Robinson GG, Taylor JA (1978) General effects of management and grazing by domestic livestock on the rangelands of the Northern Tablelands of New South Wales. The Rangeland Journal 1, 174-190.
| Crossref | Google Scholar |
White JGH (1995) A review of legume introduction in tussock grasslands with particular reference to species tolerant of low nutrient inputs. Proceedings of the New Zealand Agronomy Society 25, 77-85.
| Google Scholar |
White JGH, Meijer G, Langer RHM (1972) Oversowing grasses on sunny and shady faces. Proceedings of the New Zealand Grassland Association 34, 139-146.
| Crossref | Google Scholar |
Widdup K, Pennell C (2000) Suitability of new subterranean clovers in the Canterbury region. Proceedings of the New Zealand Grassland Association 62, 161-165.
| Crossref | Google Scholar |
Zydenbos SM, Barratt BIP, Bell NL, Ferguson CM, Gerard PJ, McNeill MR, Phillips CB, Townsend RJ, Jackson TA (2011) The impact of invertebrate pests on pasture persistence and their interrelationship with biotic and abiotic factors. Pasture Persistence – Grassland Research and Practice Series 15, 109-118.
| Crossref | Google Scholar |