

Silicon-mediated improvement of biomass yield and physio-biochemical attributes in heat-stressed spinach (Spinacia oleracea)
Quratulain Khosa A , Qamar uz Zaman A * , Tingting An B , Kamran Ashraf C , Asim Abbasi D , Saba Nazir A , Riffat Naz A and Yinglong Chen E *
A * , Tingting An B , Kamran Ashraf C , Asim Abbasi D , Saba Nazir A , Riffat Naz A and Yinglong Chen E *
A Department of Environmental Sciences, The University of Lahore, Lahore 54590, Pakistan.
B Institute of Soil and Water Conservation, Chinese Academy of Sciences, and Northwest A&F University, Yangling 712100, China.
C Department of Food Sciences, Government College University Faisalabad, Sahiwal Campus, Sahiwal 57000, Pakistan.
D Department of Environmental Sciences, Kohsar University, Murree, Pakistan.
E The UWA Institute of Agriculture, and School of Agriculture and Environment, The University of Western Australia, Perth, WA 6009, Australia.
Handling Editor: Enrico Francia
Crop & Pasture Science 74(3) 230-243 https://doi.org/10.1071/CP22192
Submitted: 3 June 2022 Accepted: 1 August 2022 Published: 6 September 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Defensive action of plants against biotic and abiotic stresses has been augmented by silicon (Si). Spinach (Spinacia oleracea L.) is a nutritious leafy vegetable that is a cold-tolerant but heat-sensitive crop.
Aims and methods: The ability of exogenous application of Si (0, 2, 4 and 6 mmol L−1 in the form of K2SiO3) to alleviate heat stress in spinach cv. Desi Palak was investigated. After an acclimatisation period, plants were grown with or without heat stress for 15 days, followed by Si treatment for 25 days. Plant growth and physiology were assessed at 65 days after sowing.
Key results: Heat stress significantly inhibited plant growth, water status and photosynthesis, soluble sugar and protein contents, and osmolyte status in spinach leaves, but increased electrolyte leakage, activities of antioxidant enzymes, and proline content. Application of Si alleviated heat stress by enhancing water status, photosynthetic pigments, soluble proteins and essential minerals, and by reducing damage of the plasma membrane. The accumulation of osmolytes counterbalance the osmotic stress imposed by heat.
Conclusions: Silicon plays a vital role in alleviating effects of heat stress by improving water status and photosynthetic rate, and accruing osmoprotectants other than proline.
Implications: Exogenous application of Si is an efficient strategy to boost tolerance of spinach plants to heat stress, having significant impact on growth and productivity of spinach at higher temperatures.
Keywords: antioxidants, heat stress, photosynthetic pigments, physio-biochemical, plasma membrane, silicon, spinach, water relations.
Introduction
Food security is essential to a growing global population, but it is jeopardised by climate change and global warming. Various studies have shown that crop production is endangered by increases in temperature at the regional level (Abdelrahman et al. 2017). By the end of the 21st Century, the average temperature will have been raised by 1.8–4.0°C (Bita and Gerats 2013; Alizadeh et al. 2022).
Heat stress significantly affects plant activities including seed germination, development, photosynthesis and reproduction, resulting in serious impacts on plant growth and, ultimately, yield of useful products (Hasanuzzaman et al. 2011). In order to survive under stressed conditions, plants have various morphological, physiological and molecular responses (Lobanov et al. 2008; Janská et al. 2010; Govindaraj et al. 2018). Specific field management options (selection of cultivar, date of sowing, method of sowing and irrigation scheduling) can be practiced to improve production under stressed conditions. However, heat stress severely limits the productivity of crop plants; for example, it limits wheat global productivity by >6% for each degree increase in temperature (Asseng et al. 2015). Despite the negative effects on overall crop production, increasing temperature has some beneficial effects on productivity in colder regions (Challinor et al. 2014; Zandalinas et al. 2021).
Spinach (Spinacia oleracea L.), a green nutritious leafy vegetable, is a cold-tolerant, heat-sensitive crop (Zhao et al. 2018; Giordano et al. 2021). Heat stress significantly affects its growth and development, causing a considerable decline in quality and yield. Studies have been done on the heat-shock response of spinach, both with whole plants and with detached leaf tissue (Moradpour et al. 2021). After being exposed to heat shock (35–50°C) for 30 min, CO2 assimilation decreases and pigment proteins in thylakoid membranes aggregate, slowing the plant’s ability to photosynthesise (Tang et al. 2007; Shabbir et al. 2022). The first heat-shock proteins in spinach leaf tissue are induced when the temperature reaches 28°C, and a full range of heat-shock proteins are produced at 36°C. If a spinach genotype had a high germination percentage at high temperature (e.g. 35°C), it could be tolerant to heat at the germination stage (Chitwood et al. 2016; Zaman et al. 2022).
Cultural practices such as irrigation, crop residues and mulching in combination with genetic modifications of crop plants are considered important tools for managing different abiotic stresses (Wahid 2007; Zaman et al. 2022). Exogenous application of mineral nutrients helps plants to combat heat stress by increasing their heat-tolerance capacity (Sarwar et al. 2019). In general, silicon (Si) supplementation to soil improves seed germination, root and shoot development, photosynthesis, nutrient uptake, nitrogen fixation, secondary metabolism and different yield attributes of crops (Mostofa et al. 2021). Furthermore, plants supplemented with Si have shown enhanced tolerance to various abiotic stresses including temperature extremes, salinity, drought, radiation and heavy metal toxicity (Khan et al. 2020b; Younis et al. 2020). Silicon applications to tomato, cucumber, rice, strawberry, date palm, barley, poinsettia and salvia effectively reduced the deleterious effects of heat stress by improving various growth-related attributes (Muneer et al. 2017; Khan et al. 2020a). In terms of plant growth, Si an essential nutrient involved in various key functions (i.e. membrane stability, cell wall stabilisation, enhancement of key enzyme activities and phytohormones interaction) under abiotic stress (Naz et al. 2022; Shabbir et al. 2022).
Use of mineral fertilisation is considered a novel approach to mitigate deleterious effects of heat stress in spinach. Moreover, balanced Si fertilisation is essential for normal physiological functioning and maintaining structural stability of plants (Waraich et al. 2012; Shabbir et al. 2022; Zaman et al. 2022). Therefore, understanding the response of spinach to heat stress when receiving nutrient amendments will be of value to spinach growers. In this regard, the present study was designed to assess the efficiency of exogenously applied Si at varying levels on plant growth, biomass, physio-biochemical parameters, antioxidant activity and osmolyte status under heat stress in spinach plants.
Materials and methods
Experimental design and treatments
A pot experiment with spinach plants was conducted in a naturally lit glasshouse at the Department of Environmental Sciences, The University of Lahore, Pakistan. A completely randomised design was used consisting of two factors with three replications per treatment: heat stress application (normal growth conditions, or heat stress); and Si application (0, 2, 4 and 6 mmol L−1). Heat stress was applied at 15 days after sowing (DAS) by placing the pots in a transparent, polyethylene-sheet tunnel, and Si was applied in the form of K2SiO3.
Experiment establishment and management
Homogenous seeds of spinach cv. Desi Palak were sterilised with 0.1% (w/v) sodium dodecyl solution and then washed with deionised water. Ten seeds were sown into each plastic pot (22.5 cm top diameter, 16.5 cm base diameter, 18 cm depth) filled with 7 kg sterilised soil. At 10 DAS, five healthy plants were maintained per pot. All pots were kept in open space under normal environmental conditions until application of the stress treatments. Tap water was used as source of irrigation to pot soil capacity on daily basis. A 50% Hoagland solution of moderate strength as a source of essential nutrients was applied weekly at a rate of 1 L per pot. The optimum temperature range for spinach growth is considered to be 25–30°C (Atherton and Farooque 1983) and its growth is suppressed at 35°C (Leskovar et al. 1999; Katzman et al. 2001).
After an acclimatisation period of 15 days, heat stress was imposed. A plastic tunnel made of black-tinted transparent polyethylene sheet was constructed over the pots by using bamboo sticks. Tiny holes were made in the sheeting to minimise the humidity. The pots in control treatment were placed under normal conditions (Shahid et al. 2017). Temperature and humidity were recorded with a digital temperature and humidity probe (Digital Multimeter 50302; Novanna Measurement Systems, Bury St Edmunds, UK). During heat stress, the temperature of control and heat-stressed pots was recorded twice a day and averaged (Both et al. 2015). A considerable increase in temperature was observed in heat-stressed pots compared with control pots (Fig. 1). After 15 days of heat stress, foliar application of Si (0, 2, 4, 6 mmol L−1) in the form of potassium silicate (K2SiO3) salt in distilled water was applied, using 500 mL of the solution in each of two sprays at a 10-day interval (Naz et al. 2022). After 25 days of exposure to Si (65 DAS), data for morphological, biochemical and physiological attributes were recorded.

|
Growth attributes
Following harvest at 65 DAS, plants were separated into leaves and roots for measurement of growth parameters. Number of leaves was counted, and plant height, leaf length and width were measured (in cm) using a ruler. Leaf area (cm2) was measured using the LI-3100 Area Meter (LI-COR Biosciences, Lincoln, NE, USA). Plants were then washed with distilled water to remove adhered soil particles and air-dried. Roots and leaves were oven-dried at 70°C for 48 h for estimation of dry weights of root and leaves separately.
Gas exchange attributes
At 65 DAS, photosynthetic rate (A), stomatal conductance (gs) and transpiration rate (E) were measured on fully expanded uppermost leaves by using a portable infrared gas analyser (ADC Bioscientific, Hoddesdon, UK) at light saturation intensity between 09:00 and 12:00 on a sunny day, as described by Emanuil et al. (2020).
Biochemical attributes
Electrolyte leakage
Small pieces of leaves (at 65 DAS) were dipped in deionised water and electrolyte leakage (EL) level was measured. The first reading of EL was taken after incubation of the sample at 32°C for 2 h, and the second reading after incubation at 121°C for 20 min (Dionisio-Sese and Tobita 1998). The following formula was used to calculate the EL level of samples:
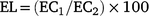
Chlorophyll content
A crushed sample of plant leaf (~5 g) was added to a test tube containing 85% acetone (v/v) and placed under dark conditions for 24 h for pigment extraction. The sample was then centrifuged for 10 min at 4000g and 4°C. A spectrophotometer (Halo DB-20/DB-20S; Dynamica Scientific, Newport Pagnell, UK) at wavelengths of 470, 647 and 664.5 nm was used to measure the amount of chlorophyll in the supernatant, following the methods described by Lichtenthaler (1987).
Enzymatic antioxidants
Fresh spinach leaves (1.0 g) were extracted in 50 mM phosphate buffer (pH ~7.8) and the homogenate was centrifuged at 15 000g for 10 min; the supernatant was used for assaying enzyme activity. The activity of peroxidase (POD) was measured according to the method described by Velikova et al. (2000), catalase (CAT) activity following the method presented by Aebi (1974), and superoxide dismutase (SOD) activity following the procedure presented by Beauchamp and Fridovich (1971).
Water-related attributes
The method of Turner and Kramer (1980) was used for determination of relative water content (RWC), and the following formula was used for the calculation:

where FW is fresh weight, TW is turgid weight, and DW is dry weight.
The water potential (Ψw) obtained by fresh leaves was measured by the use of a Pressure Bomb (Soilmoisture Equipment, Santa Barbara, CA, USA). Leaf samples, which were already used for RWC, were frozen, thawed, squeezed and centrifuged (5000g). An osmometer (Digital Osmometer; Wescor, Logan, UT, USA) was used to obtain osmotic potential.
Determination of osmolytes
Fresh leaf (0.5 g) was taken and ground in buffer (pH 7.2); a protease inhibitor cocktail in 1 μM phosphate buffered saline was added to make a homogenous mixture. The saline buffer comprised deionised water, 1.37 mM NaCl, 2 mM KH2PO4, 2.7 mM KCl and 10 mM Na2HPO4, with the pH adjusted by adding HCl. The solution was autoclaved and then centrifuged (12 000g) for ~5 min to separate the supernatant. Total soluble proteins were detected via Bradford assay (Bradford 1976), and samples were stored. Various dilutions of bovine serum albumin (10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 μg μL−1) were taken to construct standard curves. Deionised water (400 μL) with dye stock was added and the prepared sample tubes were incubated for 30 min at room temperature and then vortexed. A UV 4000 UV-Vis spectrophotometer (Dynamica Company, London, UK) was used to read the absorbance level of the sample.
The method of Maehly and Chance (1954) was followed for proline determination. Sulfosalicylic acid (3% w/v) was mixed with 0.5-g samples of fresh leaf and then filtered; the filtered samples were placed in test tubes and treated with glacial acetic acid and ninhydrin (2.5%). Samples were heated in test tubes in a water bath (100°C) for 60 min, after which toluene was added to the test tubes for the separation of chromophores. A UV-Vis spectrophotometer was used to record the optical density (520 nm). The method defined by Giannakoula et al. (2008) use to detect soluble sugar content.
Statistical analyses
Data were analysed using Fisher’s analysis of variance (ANOVA) for significance testing. Tukey’s honestly significant difference test at P = 0.05 was applied for comparison of means where the ANOVA indicated significant differences. Regression and correlation analyses were computed by using the Minitab-19 statistical software. All statistical computations were performed with Statistix software ver. 10 (Analytical Software, Tallahassee, FL, USA).
Results
Growth and biomass attributes
Heat stress, foliar-applied Si and their interaction significantly (P ≤ 0.01) affected growth and biomass attributes of spinach plants. Heat stress decreased plant height (20.4%), number of leaves (22.8%), leaf length (22.1%), leaf width (13.2%), leaf area (29.8%), root dry weight (11.1%) and leaf dry weight (25.92%) compared with the unstressed control (Fig. 2). Under normal and heat-stressed conditions, respectively, maximum plant height (27.1 and 21.4 cm), number of leaves (8.67 and 7.33), leaf length (16.8 and 13.5 cm), leaf width (3.6 and 3.1 cm), leaf area (55.5 and 42.5 cm2), root fresh weight (3.72 and 3.36 g), root dry weight (0.39 and 0.35 g), leaf fresh weight (11.93 and 10.46 g) and leaf dry weight (1.15 and 0.96 g) were observed with foliar application of 4 mmol L−1 of Si solution (Fig. 2).

|
Photosynthetic attributes
Heat stress, Si application and their interaction significantly (P ≤ 0.01) affected photosynthetic attributes of spinach plants. Decreases in transpiration rate (9.45%), photosynthetic rate (24.4%) and stomatal conductance (10.5%) were observed with heat treatment relative to the unstressed control. However, maximum increases in transpiration rate (58.7%), photosynthetic rate (158%) and stomatal conductance (64.1%) were observed with foliar application of 4 mmol L−1 of Si solution compared with the nil-Si control under heat-stress conditions (Fig. 3).

|
Enzymatic antioxidants and physio-biochemical attributes
Heat stress, foliar application of Si and their interaction showed significant (P ≤ 0.01) impact on biochemical (Fig. 4) and enzymatic (Fig. 5) attributes of spinach. Under heat stress, decreases in chlorophyll contents (21.1%) and carotenoid contents (22.1%), and increases in SOD (11.2%), POD (12.08%), CAT (18.8%) and electrolyte leakage (49.9%) were observed compared with the unstressed control. Maximum improvements in chlorophyll and carotenoid contents and decreases in SOD (56.67%), POD (45.7%), CAT (58.4%) and electrolyte leakage (33.5%) were observed with foliar application of 4 mmol L−1 of Si compared with the nil-Si control under heat-stress conditions (Figs 4 and 5).

|

|
Water-related attributes
Foliar-applied Si, heat stress and their interaction showed significant (P ≤ 0.01) impact on water-related attributes of spinach. Under heat stress, decreases in RWC (11.81%), water potential (25.66%) and osmotic potential (21.67%) were observed compared with the unstressed control. Under normal and heat-stressed conditions, respectively, maximum RWC (91.67% and 77.22%), water potential (−1.86 and −2.25 MPa), and osmotic potential (−2.24 and −2.63 MPa) were observed with foliar application of 4 mmol L−1 of Si (Fig. 6).

|
Osmolyte attributes
Heat stress and foliar-applied Si significantly (P ≤ 0.01) affected soluble sugar, soluble protein and proline contents of spinach plants. Heat stress significantly decreased the soluble sugar (20.92%) and soluble protein (17.71%) contents, and increased proline content (16.52%). Maximum soluble sugar and protein contents of 17.46 and 35.84 mg g−1 FW under control conditions, and 14.68 and 31.05 mg g−1 FW under heat-stressed conditions, were observed with foliar application of 4 mmol L−1 of Si (Fig. 7).

|
Correlation matrix
Growth, enzymatic, water-related and biochemical attributes were subjected to correlation analysis (Table 1). All of the enzymatic activities (SOD, POD and CAT) were negatively correlated with chlorophyll content, leaf dry weight, root dry weight and RWC. Significant positive correlations of enzymatic activities were noted with electrolyte leakage, proline content, osmotic potential and water potential, and among those four parameters. Electrolyte leakage correlated negatively with leaf dry weight, root dry weight and RWC. Chlorophyll content was positively correlated with leaf dry weight, root dry weight and RWC.

|
Regression analysis
Growth, physiological, water-related and biochemical attributes were also tested using regression analysis (Fig. 8a–f). The R2 values of regressions were: leaf dry weight and RWC, 73.36% (Fig. 8a); electrolyte leakage and osmotic potential, 79% (Fig. 8b); chlorophyll content and leaf dry weight association, 94.84% (Fig. 8c); stomatal conductance and water potential, 72.35% (Fig. 8d); soluble sugar and proline contents, only 45.90% (Fig. 8e); and photosynthetic rate and leaf area, 88.38% (Fig. 8f).

|
Discussion
Under stressful conditions such as drought, salt and high temperature, plants may alter their morphological, molecular, physiological and biochemical processes as adaptation strategies (Wahid 2007; Chaudhry and Sidhu 2022). The present study showed that heat stress significantly decreases the growth and biomass of spinach plants. High temperature causes loss of cell water content, ultimately reducing cell size and growth (Ali et al. 2021; Hassan et al. 2021; Okereke et al. 2021). Under extreme heat stress, plants can show programmed cell death in specific cells or tissues; on the other hand, moderate heat stress for extended periods causes gradual death. Both types of injury or death can lead to the shedding or shortening of leaves, reduction in fresh and dry biomass, or even death of the entire plant (Abdelrahman et al. 2017; Das et al. 2017; Fahad et al. 2017; Hussain et al. 2019). The results presented here concur with those of previous reports by Mahdieh et al. (2015), Abbas et al. (2017), and An et al. (2022), where the application of exogenous Si alleviated the negative effects of abiotic stress and restored plant growth. In the present investigation, we found improved leaf area, better growth, and greater fresh and dry weights in response to application of Si (Fig. 9). This suggests the involvement of Si in cell division and expansion, which might lead to internodal elongation and influence the height of the plant (Soundararajan et al. 2014; Hussain et al. 2019; Younis et al. 2020). The literature reveals that Si is considered a plant-growth-regulator-like compound that is necessary for cell division and expansion (Hwang et al. 2007; Khan et al. 2020a; An et al. 2022), and has a defensive role against a wide range of environmental stresses (Tripathi et al. 2013; Shalaby et al. 2021).

|
In all green plants, the most fundamental physiological process is photosynthesis, and all of its components are sensitive to stress conditions; any type of stress at any stage of life affects overall photosynthetic efficiency of green plants (Alabdallah et al. 2021). The present findings reveal that heat stress significantly decreases the photosynthetic attributes of spinach plants compared with control temperatures, which might be due to the decreased stomatal conductivity and transpiration rate (Mahdavi et al. 2021). In tomato, high stomatal conductivity and transpiration rate under heat stress improve leaf cooling, providing better protection for chlorophyll and maintaining relatively high photosynthetic rate (Haque et al. 2021). Under abiotic stress environments, photosynthetic pigment degradation occurs; for example, under salt stress conditions, chlorophyll pigments break down (Iqbal et al. 2021). The improvement of spinach physiological attributes might be the result of increased total chlorophyll content with optimum Si application under heat-stressed conditions (Fig. 9). Breakage in the walls of chloroplasts could be delayed with Si application, by which photosynthetic and transpiration rate and efficiency can be improved (Xie et al. 2014; Das et al. 2021). Effects of foliar-applied Si in improving photosynthetic potential and efficiency by opening angle of leaves, keeping the leaf erect, and decreasing self-shading have been reported in rice, barely, wheat and sugarcane (Soratto et al. 2012; Othmani et al. 2021).
Chlorophyll is very sensitive to heat stress (Murkowski 2001; Song et al. 2018). High-temperature-induced decline in chlorophyll and carotenoid contents has been reported in field and vegetable crops (Jeon et al. 2006). The same trend was found in spinach plants grown under a high-temperature regime in our study. Leakage of electrolytes, which are indicators of stress sensitivity, was higher in heat-treated spinach plants than the control. The increase in lipid peroxidation might be a result of the overproduction and accumulation of reactive oxygen species (ROS), which then causes membrane peroxidation, protein degradation and DNA damage, severely inhibiting growth (Awasthi et al. 2015; Zhang et al. 2017). In one study, Si enhanced chlorophyll and carotenoid contents in a barley cultivar under high-temperature regimes, possibly by protecting the chlorophyll pigments from oxidative damage through strengthening of the level of carotenoids (Heile et al. 2021; Zaman et al. 2021). Exogenously applied Si improved plant defence systems to detoxify ROS induced under heat stress, which in turn helped to increased chlorophyll and carotenoid contents by limiting electrolyte leakage (Heile et al. 2021; Zaman et al. 2021).
Tolerant plants show a tendency to protection against the damaging effects of ROS through the synthesis of various enzymatic and nonenzymatic ROS-scavenging and detoxification systems (Jing et al. 2020). Activities also differ depending upon tolerance or susceptibility of different crop varieties, their growth stages and growing season (Li et al. 2020). The antioxidants CAT, SOD and POD are known for the dismutation of hydrogen peroxide to water and molecular oxygen in cells, as well as the elimination of stress-induced ROS directly or indirectly via the production of ascorbate and glutathione (Kim et al. 2014). In this study, we found that the activities of ROS-eliminating enzymes differed significantly in spinach plants under heat-stressed and control conditions. Plants receiving foliar-applied Si showed enhanced enzymatic activity during heat stress compared with normal temperature conditions, concurring with previous findings (Soundararajan et al. 2015; Heile et al. 2021; Zaman et al. 2021).
In addition to biomass, heat stress caused a significant reduction in water-related attributes of spinach plants (Sinha et al. 2021). Our results are in agreement with previous reports in rice (Fahad et al. 2016) and wheat (Hameed et al. 2012). The decrease in leaf water content and water potential might affect plant metabolism and decrease plant growth and biomass (Sun et al. 2020). The application of Si alleviated heat stress significantly by improving the water status of spinach plants. Naz et al. (2022) found that, at a balanced Si concentration, RWC was almost constant under abiotic stress, which was probably due to an advanced ability for osmotic adjustment. Moreover, Si application helps in removing restrictions and improves turgor pressure in the cells to maintain water balance. Maximum RWC in the treatment with Si at 4 mmol L−1 under heat stress might be due to the protective effect of Si in stressed plants by maintaining membrane integrity, because one of the primary effects of heat stress is a disruption of membrane integrity. Similar findings have been reported for wheat, sorghum and maize plants, suggesting that the positive impact of Si application under drought stress may be associated with reduced transpiration (Wang et al. 2021).
Plants affected by heat stress show the accumulation of compatible solutes such soluble sugars and proline (Rivero et al. 2014; Zhao et al. 2018). In the present study, enhanced proline content was observed under heat stress compared with control temperatures. Increased accumulation of proline due to heat stress has been reported by other researchers, who concluded that proline helps to stabilise membranes, subcellular structures and cellular redox potential by destroying the radicals (Hussain et al. 2018; Wang et al. 2018; Naz et al. 2022). Liu et al. (2011) observed a significant increase in starch content under heat-shock treatment >30°C. The increase in osmolyte status, soluble protein and soluble sugars and decrease in proline content in response to Si application may be due to the fact that Si has a pivotal role in binding amino acids to form specific proteins (Soundararajan et al. 2014; Rady et al. 2019; Zaman et al. 2022), and Si is actively engaged in formation of DNA and functioning of mRNA (Abbas et al. 2015; AlKahtani et al. 2021). In addition, foliar-applied Si reduced the accumulation of proline content in the heat-stressed seedlings, which is associated with improved growth of spinach plants under heat stress (Fig. 9). This result suggests that foliar-applied Si could provide protection to cells by keeping the accumulation of proline to an optimum level, and Si probably employed other osmoprotectants for stress mitigation, such that a high level of proline accumulation was not required. The major outcomes of this research are that an appropriate level of Si not only improves the morpho-physiological parameters but also decreases electrolyte leakage. This finding addresses issues of health and nutritional challenges and of improving farm productivity in regions facing extreme heat waves. However, further research is required to test more genotypes for thermo-tolerance and assess under field conditions.
Conclusion
Heat stress significantly inhibited the growth, physiological, water relations and osmolyte status of spinach plants. Silicon supplementation improved plant growth, physio-biochemical, photosynthetic, and tissue water parameters under heat stress compared with nil-Si treatment. Application of Si enhanced plant growth, most likely through decreased electrolyte leakage. Application of Si also increased enzymatic antioxidants. Application of Si at 4 mmol L−1 performed best in alleviating heat stress in spinach plants. Moreover, exogenous application of Si is an environmentally friendly approach for growing spinach under heat-stressed conditions. Future research activities focusing on root architecture traits, molecular forms of Si and heat-stress interactions, economic benefits and diet diversity in addition to vital nutrients will be essential to developing agricultural strategies aiming at improving crop yield under abiotic stress.
Data availability
The dataset of this article can be requested from the corresponding author upon suitable request.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This research received no external funding.
References
Abbas T, Balal RM, Shahid MA, Pervez MA, Ayyub CM, Aqueel MA, Javaid MM (2015) Silicon-induced alleviation of NaCl toxicity in okra (Abelmoschus esculentus) is associated with enhanced photosynthesis, osmoprotectants and antioxidant metabolism. Acta Physiologiae Plantarum 37, 6| Silicon-induced alleviation of NaCl toxicity in okra (Abelmoschus esculentus) is associated with enhanced photosynthesis, osmoprotectants and antioxidant metabolism.Crossref | GoogleScholarGoogle Scholar |
Abbas T, Sattar A, Ijaz M, Aatif M, Khalid S, Sher A (2017) Exogenous silicon application alleviates salt stress in okra. Horticulture, Environment, and Biotechnology 58, 342–349.
| Exogenous silicon application alleviates salt stress in okra.Crossref | GoogleScholarGoogle Scholar |
Abdelrahman M, El-Sayed M, Jogaiah S, Burritt DJ, Tran L-SP (2017) The “STAY-GREEN” trait and phytohormone signaling networks in plants under heat stress. Plant Cell Reports 36, 1009–1025.
| The “STAY-GREEN” trait and phytohormone signaling networks in plants under heat stress.Crossref | GoogleScholarGoogle Scholar |
Aebi H (1974) Catalase. In ‘Methods of enzymatic analysis.’ pp. 673–684. (Academic Press: Cambridge, MA, USA)
Alabdallah NM, Hasan MM, Hammami I, Alghamdi AI, Alshehri D, Alatawi HA (2021) Green synthesized metal oxide nanoparticles mediate growth regulation and physiology of crop plants under drought stress. Plants 10, 1730
| Green synthesized metal oxide nanoparticles mediate growth regulation and physiology of crop plants under drought stress.Crossref | GoogleScholarGoogle Scholar |
Ali MM, Waleed Shafique M, Gull S, Afzal Naveed W, Javed T, Yousef AF, Mauro RP (2021) Alleviation of heat stress in tomato by exogenous application of sulfur. Horticulturae 7, 21
| Alleviation of heat stress in tomato by exogenous application of sulfur.Crossref | GoogleScholarGoogle Scholar |
Alizadeh MR, Abatzoglou JT, Adamowski JF, Prestemon JP, Chittoori B, Akbari Asanjan A, Sadegh M (2022) Increasing heat-stress inequality in a warming climate. Earth’s Future 10, e2021EF002488
| Increasing heat-stress inequality in a warming climate.Crossref | GoogleScholarGoogle Scholar |
AlKahtani MDF, Hafez YM, Attia K, Rashwan E, Husnain LA, AlGwaiz HIM, Abdelaal KAA (2021) Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. Antioxidants 10, 398
| Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet.Crossref | GoogleScholarGoogle Scholar |
An T, Gao Y, Kuang Q, Wu Y, Zaman Q, Zhang Y, Xu B, Chen Y (2022) Effect of silicon on morpho-physiological attributes, yield and cadmium accumulation in two maize genotypes with contrasting root system size and health risk assessment. Plant and Soil
| Effect of silicon on morpho-physiological attributes, yield and cadmium accumulation in two maize genotypes with contrasting root system size and health risk assessment.Crossref | GoogleScholarGoogle Scholar |
Asseng S, Ewert F, Martre P, et al. (2015) Rising temperatures reduce global wheat production. Nature Climate Change 5, 143–147.
| Rising temperatures reduce global wheat production.Crossref | GoogleScholarGoogle Scholar |
Atherton JG, Farooque AM (1983) High temperature and germination in spinach. I. The role of the pericarp. Scientia Horticulturae 19, 25–32.
| High temperature and germination in spinach. I. The role of the pericarp.Crossref | GoogleScholarGoogle Scholar |
Awasthi R, Bhandari K, Nayyar H (2015) Temperature stress and redox homeostasis in agricultural crops. Frontiers in Environmental Science 3, 11
| Temperature stress and redox homeostasis in agricultural crops.Crossref | GoogleScholarGoogle Scholar |
Beauchamp C, Fridovich I (1971) Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry 44, 276–287.
| Superoxide dismutase: improved assays and an assay applicable to acrylamide gels.Crossref | GoogleScholarGoogle Scholar |
Bita CE, Gerats T (2013) Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Frontiers in Plant Science 4, 273
| Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops.Crossref | GoogleScholarGoogle Scholar |
Both AJ, Benjamin L, Franklin J, Holroyd G, Incoll LD, Lefsrud MG, Pitkin G (2015) Guidelines for measuring and reporting environmental parameters for experiments in greenhouses. Plant Methods 11, 43
| Guidelines for measuring and reporting environmental parameters for experiments in greenhouses.Crossref | GoogleScholarGoogle Scholar |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72, 248–254.
| A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Crossref | GoogleScholarGoogle Scholar |
Challinor AJ, Watson J, Lobell DB, Howden SM, Smith DR, Chhetri N (2014) A meta-analysis of crop yield under climate change and adaptation. Nature Climate Change 4, 287–291.
| A meta-analysis of crop yield under climate change and adaptation.Crossref | GoogleScholarGoogle Scholar |
Chaudhry S, Sidhu GPS (2022) Climate change regulated abiotic stress mechanisms in plants: a comprehensive review. Plant Cell Reports 41, 1–31.
| Climate change regulated abiotic stress mechanisms in plants: a comprehensive review.Crossref | GoogleScholarGoogle Scholar |
Chitwood J, Shi A, Evans M, Rom C, Gbur EE, Motes D, Chen P, Hensley D (2016) Effect of temperature on seed germination in spinach (Spinacia oleracea). HortScience 51, 1475–1478.
| Effect of temperature on seed germination in spinach (Spinacia oleracea).Crossref | GoogleScholarGoogle Scholar |
Das A, Rushton PJ, Rohila JS (2017) Metabolomic profiling of soybeans (Glycine max L.) reveals the importance of sugar and nitrogen metabolism under drought and heat stress. Plants 6, 21
| Metabolomic profiling of soybeans (Glycine max L.) reveals the importance of sugar and nitrogen metabolism under drought and heat stress.Crossref | GoogleScholarGoogle Scholar |
Das D, Ullah H, Tisarum R, Cha-um S, Datta A (2021) Morpho-physiological responses of tropical rice to potassium and silicon fertilization under water-deficit stress. Journal of Soil Science and Plant Nutrition
| Morpho-physiological responses of tropical rice to potassium and silicon fertilization under water-deficit stress.Crossref | GoogleScholarGoogle Scholar |
Dionisio-Sese ML, Tobita S (1998) Antioxidant responses of rice seedlings to salinity stress. Plant Sciences 135, 1–9.
| Antioxidant responses of rice seedlings to salinity stress.Crossref | GoogleScholarGoogle Scholar |
Emanuil N, Akram MS, Ali S, El-Esawi MA, Iqbal M, Alyemeni MN (2020) Peptone-Induced physio-biochemical modulations reduce cadmium toxicity and accumulation in spinach (Spinacia oleracea L.). Plants 9, 1806
| Peptone-Induced physio-biochemical modulations reduce cadmium toxicity and accumulation in spinach (Spinacia oleracea L.).Crossref | GoogleScholarGoogle Scholar |
Fahad S, Hussain S, Saud S, Hassan S, Tanveer M, Ihsan MZ, Huang J (2016) A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice. Plant Physiology and Biochemistry 103, 191–198.
| A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice.Crossref | GoogleScholarGoogle Scholar |
Fahad S, Bajwa AA, Nazir U, Anjum SA, Farooq A, Zohaib A, Huang J (2017) Crop production under drought and heat stress: plant responses and management options. Frontiers in Plant Science 8, 1147
| Crop production under drought and heat stress: plant responses and management options.Crossref | GoogleScholarGoogle Scholar |
Giannakoula A, Moustakas M, Mylona P, Papadakis I, Yupsanis T (2008) Aluminum tolerance in maize is correlated with increased levels of mineral nutrients, carbohydrates and proline, and decreased levels of lipid peroxidation and Al accumulation. Journal of Plant Physiology 165, 385–396.
| Aluminum tolerance in maize is correlated with increased levels of mineral nutrients, carbohydrates and proline, and decreased levels of lipid peroxidation and Al accumulation.Crossref | GoogleScholarGoogle Scholar |
Giordano M, Petropoulos SA, Rouphael Y (2021) Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 11, 463
| Response and defence mechanisms of vegetable crops against drought, heat and salinity stress.Crossref | GoogleScholarGoogle Scholar |
Govindaraj M, Pattanashetti SK, Patne N, Kanatti AA (2018) Breeding cultivars for heat stress tolerance in staple food crops. In ‘Next generation plant breeding’. (Ed. YÖ Çiftçi) pp. 45–74. (InTechOpen: London, UK)
Hameed A, Goher M, Iqbal N (2012) Heat stress-induced cell death, changes in antioxidants, lipid peroxidation, and protease activity in wheat leaves. Journal of Plant Growth Regulation 31, 283–291.
| Heat stress-induced cell death, changes in antioxidants, lipid peroxidation, and protease activity in wheat leaves.Crossref | GoogleScholarGoogle Scholar |
Haque MS, Husna MT, Uddin MN, Hossain MA, Sarwar AKMG, Ali OM, Abdel Latef AAH, Hossain A (2021) Heat stress at early reproductive stage differentially alters several physiological and biochemical traits of three tomato cultivars. Horticulturae 7, 330
| Heat stress at early reproductive stage differentially alters several physiological and biochemical traits of three tomato cultivars.Crossref | GoogleScholarGoogle Scholar |
Hasanuzzaman M, Hossain MA, Fujita M (2011) Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnology Reports 5, 353–365.
| Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings.Crossref | GoogleScholarGoogle Scholar |
Hassan MU, Chattha MU, Khan I, Chattha MB, Barbanti L, Aamer M, Aslam MT (2021) Heat stress in cultivated plants: nature, impact, mechanisms, and mitigation strategies: a review. Plant Biosystems 155, 211–234.
| Heat stress in cultivated plants: nature, impact, mechanisms, and mitigation strategies: a review.Crossref | GoogleScholarGoogle Scholar |
Heile AO, Zaman Qu, Aslam Z, Hussain A, Aslam M, Saleem MH, Abualreesh MH, Alatawi A, Ali S (2021) Alleviation of cadmium phytotoxicity using silicon fertilization in wheat by altering antioxidant metabolism and osmotic adjustment. Sustainability 13, 11317
| Alleviation of cadmium phytotoxicity using silicon fertilization in wheat by altering antioxidant metabolism and osmotic adjustment.Crossref | GoogleScholarGoogle Scholar |
Hussain M, Farooq S, Hasan W, Ul-Allah S, Tanveer M, Farooq M, Nawaz A (2018) Drought stress in sunflower: physiological effects and its management through breeding and agronomic alternatives. Agricultural Water Management 201, 152–166.
| Drought stress in sunflower: physiological effects and its management through breeding and agronomic alternatives.Crossref | GoogleScholarGoogle Scholar |
Hussain I, Parveen A, Rasheed R, Ashraf MA, Ibrahim M, Riaz S, Afzaal Z, Iqbal M (2019) Exogenous silicon modulates growth, physio-chemicals and antioxidants in barley (Hordeum vulgare L.) exposed to different temperature regimes. Silicon 11, 2753–2762.
| Exogenous silicon modulates growth, physio-chemicals and antioxidants in barley (Hordeum vulgare L.) exposed to different temperature regimes.Crossref | GoogleScholarGoogle Scholar |
Hwang S-J, Hamayun M, Kim H-Y, Na C-I, Kim K-U, Shin D-H, Lee I-J (2007) Effect of nitrogen and silicon nutrition on bioactive gibberellin and growth of rice under field conditions. Journal of Crop Science and Biotechnology 10, 281–286.
Iqbal P, Ghani MA, Ali B, Shahid M, Iqbal Q, Ziaf K, Ahmad J (2021) Exogenous application of glutamic acid promotes cucumber (Cucumis sativus L.) growth under salt stress conditions. Emirates Journal of Food and Agriculture 33, 407–416.
| Exogenous application of glutamic acid promotes cucumber (Cucumis sativus L.) growth under salt stress conditions.Crossref | GoogleScholarGoogle Scholar |
Janská A, Maršík P, Zelenková S, Ovesná J (2010) Cold stress and acclimation: what is important for metabolic adjustment? Plant Biology 12, 395–405.
| Cold stress and acclimation: what is important for metabolic adjustment?Crossref | GoogleScholarGoogle Scholar |
Jeon M-W, Ali MB, Hahn E-J, Paek K-Y (2006) Photosynthetic pigments, morphology and leaf gas exchange during ex vitro acclimatization of micropropagated CAM Doritaenopsis plantlets under relative humidity and air temperature. Environmental and Experimental Botany 55, 183–194.
| Photosynthetic pigments, morphology and leaf gas exchange during ex vitro acclimatization of micropropagated CAM Doritaenopsis plantlets under relative humidity and air temperature.Crossref | GoogleScholarGoogle Scholar |
Jing J, Guo S, Li Y, Li W (2020) The alleviating effect of exogenous polyamines on heat stress susceptibility of different heat resistant wheat (Triticum aestivum L.) varieties. Scientific Reports 10, 7467
| The alleviating effect of exogenous polyamines on heat stress susceptibility of different heat resistant wheat (Triticum aestivum L.) varieties.Crossref | GoogleScholarGoogle Scholar |
Katzman LS, Taylor AG, Langhans RW (2001) Seed enhancements to improve spinach germination. HortScience 36, 979–981.
| Seed enhancements to improve spinach germination.Crossref | GoogleScholarGoogle Scholar |
Khan A, Bilal S, Khan AL, Imran M, Shahzad R, Al-Harrasi A, Lee IJ (2020a) Silicon and gibberellins: synergistic function in harnessing ABA signaling and heat stress tolerance in date palm (Phoenix dactylifera L.). Plants 9, 620
| Silicon and gibberellins: synergistic function in harnessing ABA signaling and heat stress tolerance in date palm (Phoenix dactylifera L.).Crossref | GoogleScholarGoogle Scholar |
Khan MN, Siddiqui MH, AlSolami MA, Alamri S, Hu Y, Ali HM, Al-Amri AA, Alsubaie QD, Al-Munqedhi BMA, Al-Ghamdi A (2020b) Crosstalk of hydrogen sulfide and nitric oxide requires calcium to mitigate impaired photosynthesis under cadmium stress by activating defense mechanisms in Vigna radiata. Plant Physiology and Biochemistry 156, 278–290.
| Crosstalk of hydrogen sulfide and nitric oxide requires calcium to mitigate impaired photosynthesis under cadmium stress by activating defense mechanisms in Vigna radiata.Crossref | GoogleScholarGoogle Scholar |
Kim YH, Khan AL, Waqas M, Shim JK, Kim DH, Lee KY, Lee IJ (2014) Silicon application to rice root zone influenced the phytohormonal and antioxidant responses under salinity stress. Journal of Plant Growth Regulation 33, 137–149.
| Silicon application to rice root zone influenced the phytohormonal and antioxidant responses under salinity stress.Crossref | GoogleScholarGoogle Scholar |
Leskovar DI, Esensee V, Belefant-Miller H (1999) Pericarp, leachate, and carbohydrate involvement in thermoinhibition of germinating spinach seeds. Journal of the American Society for Horticultural Science 124, 301–306.
| Pericarp, leachate, and carbohydrate involvement in thermoinhibition of germinating spinach seeds.Crossref | GoogleScholarGoogle Scholar |
Li C, Han Y, Hao J, Qin X, Liu C, Fan S (2020) Effects of exogenous spermidine on antioxidants and glyoxalase system of lettuce seedlings under high temperature. Plant Signaling & Behavior 15, 1824697
| Effects of exogenous spermidine on antioxidants and glyoxalase system of lettuce seedlings under high temperature.Crossref | GoogleScholarGoogle Scholar |
Lichtenthaler HK (1987) Chlorophylls and caroteniods: pigments of photosynthetic biomembranes. Methods in Enzymology 148, 350–382.
| Chlorophylls and caroteniods: pigments of photosynthetic biomembranes.Crossref | GoogleScholarGoogle Scholar |
Liu P, Guo W, Jiang Z, Pu H, Feng C, Zhu X, Peng Y, Kuang A, Little CR (2011) Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.). The Journal of Agricultural Science 149, 159–169.
| Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.).Crossref | GoogleScholarGoogle Scholar |
Lobanov AV, Hatfield DL, Gladyshev VN (2008) Reduced reliance on the trace element selenium during evolution of mammals. Genome Biology 9, R62
| Reduced reliance on the trace element selenium during evolution of mammals.Crossref | GoogleScholarGoogle Scholar |
Maehly AC, Chance B (1954) The assay of catalases and peroxidases. Methods of Biochemical Analysis 1, 357–424.
| The assay of catalases and peroxidases.Crossref | GoogleScholarGoogle Scholar |
Mahdavi S, Arzani A, Maibody SAMM, Mehrabi AA (2021) Photosynthetic and yield performance of wheat (Triticum aestivum L.) under sowing in hot environment. Acta Physiologiae Plantarum 43, 106
| Photosynthetic and yield performance of wheat (Triticum aestivum L.) under sowing in hot environment.Crossref | GoogleScholarGoogle Scholar |
Mahdieh M, Habibollahi N, Amirjani MR, Abnosi MH, Ghorbanpour M (2015) Exogenous silicon nutrition ameliorates salt-induced stress by improving growth and efficiency of PSII in Oryza sativa L. cultivars. Journal of Soil Science and Plant Nutrition 15, 1050–1060.
| Exogenous silicon nutrition ameliorates salt-induced stress by improving growth and efficiency of PSII in Oryza sativa L. cultivars.Crossref | GoogleScholarGoogle Scholar |
Moradpour M, Abdullah SNA, Namasivayam P (2021) The impact of heat stress on morpho-physiological response and expression of specific genes in the heat stress-responsive transcriptional regulatory network in Brassica oleracea. Plants 10, 1064
| The impact of heat stress on morpho-physiological response and expression of specific genes in the heat stress-responsive transcriptional regulatory network in Brassica oleracea.Crossref | GoogleScholarGoogle Scholar |
Mostofa MG, Rahman MM, Ansary MMU, Keya SS, Abdelrahman M, Miah MG, Phan Tran L-S (2021) Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Critical Reviews in Biotechnology 41, 918–934.
| Silicon in mitigation of abiotic stress-induced oxidative damage in plants.Crossref | GoogleScholarGoogle Scholar |
Muneer S, Park YG, Kim S, Jeong BR (2017) Foliar or subirrigation silicon supply mitigates high temperature stress in strawberry by maintaining photosynthetic and stress-responsive proteins. Journal of Plant Growth Regulation 36, 836–845.
| Foliar or subirrigation silicon supply mitigates high temperature stress in strawberry by maintaining photosynthetic and stress-responsive proteins.Crossref | GoogleScholarGoogle Scholar |
Murkowski A (2001) Heat stress and spermidine: effect on chlorophyll fluorescence in tomato plants. Biologia Plantarum 44, 53–57.
| Heat stress and spermidine: effect on chlorophyll fluorescence in tomato plants.Crossref | GoogleScholarGoogle Scholar |
Naz R, Zaman Qu, Nazir S, Komal N, Chen Y, Ashraf K, Al-Huqail AA, Alfagham A, Siddiqui MH, Ali HM, Khan F, Sultan K, Khosa Q (2022) Silicon fertilization counteracts salinity-induced damages associated with changes in physio-biochemical modulations in spinach. PLoS ONE 17, e0267939
| Silicon fertilization counteracts salinity-induced damages associated with changes in physio-biochemical modulations in spinach.Crossref | GoogleScholarGoogle Scholar |
Okereke CN, Liu B, Kaurilind E, Niinemets Ü (2021) Heat stress resistance drives coordination of emissions of suites of volatiles after severe heat stress and during recovery in five tropical crops. Environmental and Experimental Botany 184, 104375
| Heat stress resistance drives coordination of emissions of suites of volatiles after severe heat stress and during recovery in five tropical crops.Crossref | GoogleScholarGoogle Scholar |
Othmani A, Ayed S, Bezzin O, Farooq M, Ayed-Slama O, Slim-Amara H, Ben Younes M (2021) Effect of silicon supply methods on durum wheat (Triticum durum Desf.) response to drought stress. Silicon 13, 3047–3057.
| Effect of silicon supply methods on durum wheat (Triticum durum Desf.) response to drought stress.Crossref | GoogleScholarGoogle Scholar |
Rady MM, Elrys AS, Abo El-Maati MF, Desoky E-SM (2019) Interplaying roles of silicon and proline effectively improve salt and cadmium stress tolerance in Phaseolus vulgaris plant. Plant Physiology and Biochemistry 139, 558–568.
| Interplaying roles of silicon and proline effectively improve salt and cadmium stress tolerance in Phaseolus vulgaris plant.Crossref | GoogleScholarGoogle Scholar |
Rivero RM, Mestre TC, Mittler R, Rubio F, Garcia-Sanchez F, Martinez V (2014) The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant, Cell & Environment 37, 1059–1073.
| The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants.Crossref | GoogleScholarGoogle Scholar |
Sarwar M, Saleem MF, Ullah N, Ali S, Rizwan M, Shahid MR, Alyemeni MN, Alamri SA, Ahmad P (2019) Role of mineral nutrition in alleviation of heat stress in cotton plants grown in glasshouse and field conditions. Scientific Reports 9, 13022
| Role of mineral nutrition in alleviation of heat stress in cotton plants grown in glasshouse and field conditions.Crossref | GoogleScholarGoogle Scholar |
Shabbir R, Javed T, Hussain S, Ahmar S, Naz M, Zafar H, Pandey S, Chauhan J, Siddiqui MH, Pinghua C (2022) Calcium homeostasis and potential roles to combat environmental stresses in plants. South African Journal of Botany 148, 683–693.
| Calcium homeostasis and potential roles to combat environmental stresses in plants.Crossref | GoogleScholarGoogle Scholar |
Shahid M, Saleem MF, Anjum SA, Shahid M, Afzal I (2017) Effect of terminal heat stress on proline, secondary metabolites and yield components of wheat (Triticum aestivum L.) genotypes. The Philippine Agricultural Scientist 100, 278–286.
Shalaby TA, Abd-Alkarim E, El-Aidy F, Hamed E-S, Sharaf-Eldin M, Taha N, El-Ramady H, Bayoumi Y, dos Reis AR (2021) Nano-selenium, silicon and H2O2 boost growth and productivity of cucumber under combined salinity and heat stress. Ecotoxicology and Environmental Safety 212, 111962
| Nano-selenium, silicon and H2O2 boost growth and productivity of cucumber under combined salinity and heat stress.Crossref | GoogleScholarGoogle Scholar |
Sinha T, Kumar S, Singh AK (2021) Physio-biochemical and yield response of chickpea genotypes under salinity and high temperature stress. International Journal of Environment and Climate Change 11, 136–147.
| Physio-biochemical and yield response of chickpea genotypes under salinity and high temperature stress.Crossref | GoogleScholarGoogle Scholar |
Song L, Guanter L, Guan K, You L, Huete A, Ju W, Zhang Y (2018) Satellite sun-induced chlorophyll fluorescence detects early response of winter wheat to heat stress in the Indian Indo-Gangetic Plains. Global Change Biology 24, 4023–4037.
| Satellite sun-induced chlorophyll fluorescence detects early response of winter wheat to heat stress in the Indian Indo-Gangetic Plains.Crossref | GoogleScholarGoogle Scholar |
Soratto RP, Crusciol CAC, Castro GSA, Costa CHMd, Ferrari Neto J (2012) Leaf application of silicic acid to white oat and wheat. Revista Brasileira de Ciência do Solo 36, 1538–1544.
| Leaf application of silicic acid to white oat and wheat.Crossref | GoogleScholarGoogle Scholar |
Soundararajan P, Sivanesan I, Jana S, Jeong BR (2014) Influence of silicon supplementation on the growth and tolerance to high temperature in Salvia splendens. Horticulture, Environment, and Biotechnology 55, 271–279.
| Influence of silicon supplementation on the growth and tolerance to high temperature in Salvia splendens.Crossref | GoogleScholarGoogle Scholar |
Soundararajan P, Manivannan A, Park YG, Muneer S, Jeong BR (2015) Silicon alleviates salt stress by modulating antioxidant enzyme activities in Dianthus caryophyllus ‘Tula’. Horticulture, Environment, and Biotechnology 56, 233–239.
| Silicon alleviates salt stress by modulating antioxidant enzyme activities in Dianthus caryophyllus ‘Tula’.Crossref | GoogleScholarGoogle Scholar |
Sun YL, Wang YH, Deng LF, Shi X, Bai XF (2020) Moderate soil salinity alleviates the impacts of drought on growth and water status of plants. Russian Journal of Plant Physiology 67, 153–161.
| Moderate soil salinity alleviates the impacts of drought on growth and water status of plants.Crossref | GoogleScholarGoogle Scholar |
Tang Y, Wen X, Lu Q, Yang Z, Cheng Z, Lu C (2007) Heat stress induces an aggregation of the light-harvesting complex of photosystem II in spinach plants. Plant Physiology 143, 629–638.
| Heat stress induces an aggregation of the light-harvesting complex of photosystem II in spinach plants.Crossref | GoogleScholarGoogle Scholar |
Tripathi P, Tripathi RD, Singh RP, Dwivedi S, Goutam D, Shri M, Trivedi PK, Chakrabarty D (2013) Silicon mediates arsenic tolerance in rice (Oryza sativa L.) through lowering of arsenic uptake and improved antioxidant defence system. Ecological Engineering 52, 96–103.
| Silicon mediates arsenic tolerance in rice (Oryza sativa L.) through lowering of arsenic uptake and improved antioxidant defence system.Crossref | GoogleScholarGoogle Scholar |
Turner NC, Kramer PJ (1980) ‘Adaptation of plants to water and high temperature stress.’ (Wiley Publishers: Hoboken, NJ, USA)
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sciences 151, 59–66.
| Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines.Crossref | GoogleScholarGoogle Scholar |
Wahid A (2007) Physiological implications of metabolite biosynthesis for net assimilation and heat-stress tolerance of sugarcane (Saccharum officinarum) sprouts. Journal of Plant Research 120, 219–228.
| Physiological implications of metabolite biosynthesis for net assimilation and heat-stress tolerance of sugarcane (Saccharum officinarum) sprouts.Crossref | GoogleScholarGoogle Scholar |
Wang J, Yuan B, Xu Y, Huang B (2018) Differential responses of amino acids and soluble proteins to heat stress associated with genetic variations in heat tolerance for hard fescue. Journal of the American Society for Horticultural Science 143, 45–55.
| Differential responses of amino acids and soluble proteins to heat stress associated with genetic variations in heat tolerance for hard fescue.Crossref | GoogleScholarGoogle Scholar |
Wang M, Wang R, Mur LAJ, Ruan J, Shen Q, Guo S (2021) Functions of silicon in plant drought stress responses. Horticulture Research 8, 254
| Functions of silicon in plant drought stress responses.Crossref | GoogleScholarGoogle Scholar |
Waraich EA, Ahmad R, Halim A, Aziz T (2012) Alleviation of temperature stress by nutrient management in crop plants: a review. Journal of Soil Science and Plant Nutrition 12, 221–244.
| Alleviation of temperature stress by nutrient management in crop plants: a review.Crossref | GoogleScholarGoogle Scholar |
Xie Z, Song F, Xu H, Shao H, Song R (2014) Effects of silicon on photosynthetic characteristics of maize (Zea mays L.) on alluvial soil. The Scientific World Journal 2014, 718716
| Effects of silicon on photosynthetic characteristics of maize (Zea mays L.) on alluvial soil.Crossref | GoogleScholarGoogle Scholar |
Younis AA, Khattab H, Emam MM (2020) Impacts of silicon and silicon nanoparticles on leaf ultrastructure and TaPIP1 and TaNIP2 gene expressions in heat stressed wheat seedlings. Biologia Plantarum 64, 343–352.
| Impacts of silicon and silicon nanoparticles on leaf ultrastructure and TaPIP1 and TaNIP2 gene expressions in heat stressed wheat seedlings.Crossref | GoogleScholarGoogle Scholar |
Zaman Qu, Rashid M, Nawaz R, Hussain A, Ashraf K, Latif M, Heile AO, Chen Y (2021) Silicon fertilization: a step towards cadmium-free fragrant rice. Plants 10, 2440
| Silicon fertilization: a step towards cadmium-free fragrant rice.Crossref | GoogleScholarGoogle Scholar |
Zaman Qu, Abbasi A, Tabassum S, Ashraf K, Ahmad Z, Siddiqui MH, Alamri S, Maqsood S, Sultan K (2022) Calcium induced growth, physio-biochemical, antioxidants, osmolytes adjustments and phytoconstituents status in spinach under heat stress. South African Journal of Botany 149, 701–711.
| Calcium induced growth, physio-biochemical, antioxidants, osmolytes adjustments and phytoconstituents status in spinach under heat stress.Crossref | GoogleScholarGoogle Scholar |
Zandalinas SI, Fritschi FB, Mittler R (2021) Global warming, climate change, and environmental pollution: recipe for a multifactorial stress combination disaster. Trends in Plant Sciences 26, 588–599.
| Global warming, climate change, and environmental pollution: recipe for a multifactorial stress combination disaster.Crossref | GoogleScholarGoogle Scholar |
Zhang B, Wijesundara NM, Abbey L, Rupasinghe HPV (2017) Growing medium amendments effect on growth, secondary metabolites and anti-streptococcal activity of two species of Plectranthus. Journal of Applied Research on Medicinal and Aromatic Plants 5, 53–59.
| Growing medium amendments effect on growth, secondary metabolites and anti-streptococcal activity of two species of Plectranthus.Crossref | GoogleScholarGoogle Scholar |
Zhao Q, Chen W, Bian J, Xie H, Li Y, Xu C, Ma J, Guo S, Chen J, Cai X, Wang X, Wang Q, She Y, Chen S, Zhou Z, Dai S (2018) Proteomics and phosphoproteomics of heat stress-responsive mechanisms in spinach. Frontiers in Plant Science 9, 800
| Proteomics and phosphoproteomics of heat stress-responsive mechanisms in spinach.Crossref | GoogleScholarGoogle Scholar |