

Characterisation of Sardinian germplasm of the perennial pasture grass Phalaris aquatica
Richard A. Culvenor A D , Stuart Kemp B and Kevin F. M. Reed C
A D , Stuart Kemp B and Kevin F. M. Reed C
A CSIRO Agriculture and Food, GPO Box 1700, Canberra, ACT 2601, Australia.
B PastureWise Pty Ltd, 1485 Bamganie Road, Cargerie, Vic. 3334, Australia.
C Reed Pasture Science, Brighton East, Vic. 3187, Australia.
D Corresponding author. Email: richard.culvenor@csiro.au
Crop and Pasture Science 71(9) 850-863 https://doi.org/10.1071/CP20216
Submitted: 29 June 2020 Accepted: 27 August 2020 Published: 29 September 2020
Journal Compilation © CSIRO 2020 Open Access CC BY-NC
Abstract
Germplasm of the perennial pasture grass Phalaris aquatica L., from Sardinia, Italy, is a potentially valuable source for grass breeders owing to climatic similarities to regions where P. aquatica is used, a relatively high incidence of acidic soils, and exposure to prolonged grazing pressure. At field sites in south-eastern Australia, Sardinian accessions were compared as spaced plants and drill-rows with accessions from southern Europe and north-western Africa and with commercial cultivars. They were also evaluated in grazed swards at three sites over 4 years under conditions that challenge persistence, including heavy grazing pressure, acid soils and drought. Morphologically, the accessions were comparatively dense, fine and short, with similarities to southern European accessions and cultivars of the cv. Australian type. However, they were earlier heading and more summer-dormant, particularly those from southern Sardinia. In drill-rows, Sardinian accessions were later heading and less productive in winter than accessions from Morocco. In swards, Sardinian accessions had lower seedling vigour and winter growth potential than modern winter-active cultivars. However, they were dense and persistent under high grazing pressure, and some accessions survived better than all cultivars on an acid, low-fertility soil. Developing cultivars that are superior to the cv. Australian type with acceptable seed production and alkaloid levels presents a challenge to breeders. However Sardinian germplasm offers a range of maturity times combined with higher levels of summer dormancy and a grazing-tolerant morphology, attributes that may expand the area of adaptation of the species into the hotter and more drought-prone margin.
Keywords: acid soils, aluminium, drought, grazing tolerance, Mediterranean, permanent pasture, persistence, summer dormancy.
Introduction
The Mediterranean Basin has been a rich source of pasture grass and legume germplasm for regions of the world with mild winters and drought-prone summers, such as southern Australia (Donald 1970; Cunningham et al. 1997; Nichols et al. 2012; Porqueddu et al. 2016). The Mediterranean perennial grass Phalaris aquatica L. (phalaris) was first recognised as a valuable forage species through a single introduction into Australia now known as cv. Australian, possibly from Italy (Oram et al. 2009). Phalaris is widely used in Australia and to a lesser extent in Argentina, Uruguay, New Zealand, USA, South Africa and regions of the Mediterranean. Following agronomic and selection work within the original introduction (Trumble 1933; Trumble and Cashmore 1934), germplasm-collection expeditions to the Mediterranean region in the 1950s, such as those of Donald and Miles in 1951 and Neal-Smith in 1954 (Neal-Smith 1955), led to the recognition by Australian pasture breeders of valuable variation in the species. A breeding strategy was adopted that formed broad-based populations from which so-called ‘winter-active’ cultivars were bred, with attributes such as wider adaptation, more vigorous seedlings and higher cool-season productivity than cv. Australian (McWilliam and Latter 1970; Oram et al. 2009). Accessions used in these broad-based populations were widely distributed in geographic origin: Morocco, Algeria, Portugal, Spain, Italy (mainland), Greece and Israel (Oram et al. 2009). Collection localities were predominantly neutral to alkaline in soil pH and frequently in areas protected from grazing or in cultivated areas (Neal-Smith 1955).
One region overlooked by earlier expeditions was the island of Sardinia, Italy. Sardinia is of particular interest to Australian pasture breeders because of its general climatic similarity to the medium- to high-rainfall areas of southern Australia, its higher incidence of acidic soils than in the Mediterranean region more generally, and a long history of continuous, close grazing by sheep, similar to that applied to many Australian pastures (Nichols and Francis 1993). For example, Sardinia has been a major source of accessions and cultivars of subterranean clover (Trifolium subterraneum L.), including several highly valued, late-maturing cultivars (Nichols et al. 2012, 2013). Subterranean clover is a species that prefers acidic rather than alkaline soils (Francis and Gillespie 1981) and is the most common companion legume with P. aquatica in Australia.
Soil acidity and grazing pressure are two factors that reduce the persistence of P. aquatica pastures in southern Australia, particularly when they occur together (Culvenor and Simpson 2014). Acidic soils are widespread in medium- to high-rainfall areas in south-eastern Australia where P. aquatica is climatically well adapted. Breeding for improved acid-soil tolerance has been a priority for P. aquatica in Australia since the 1980s, and two cultivars, Landmaster and Advanced AT, resulted from this (Culvenor and Simpson 2014). Whereas the original cultivar, cv. Australian, is a densely tillered, relatively prostrate cultivar that is highly persistent under commercial grazing pressures, the more erect, large-leaved winter-active cultivars have been less persistent, requiring considerable breeding effort to address (Culvenor and Simpson 2014, 2016).
Phalaris aquatica is widely distributed in Sardinia (Baldini 1993). Given prolonged exposure to both acid soils and high grazing pressure, Sardinian germplasm of P. aquatica may prove valuable for future breeding in southern Australia and similar environments elsewhere. For this reason, collection of P. aquatica germplasm from Sardinia was a priority for a joint Australian and USA perennial-forage collection mission conducted in 1994 in collaboration with institutions in Sardinia, Tunisia and Morocco (Cunningham et al. 1997). A second collection was made in 2000 by G Sandral, NSW Department of Primary Industries, in an expedition focused on perennials for cropping systems in low-rainfall environments. The purpose of the present study was to characterise germplasm from both collections. Phenological and morphological characteristics of spaced plants were compared with those of commercial cultivars and some earlier accessions held by CSIRO. Agronomic performance of the accessions from the 1994 expedition was also examined at multiple sites in south-eastern Australia.
Materials and methods
Germplasm
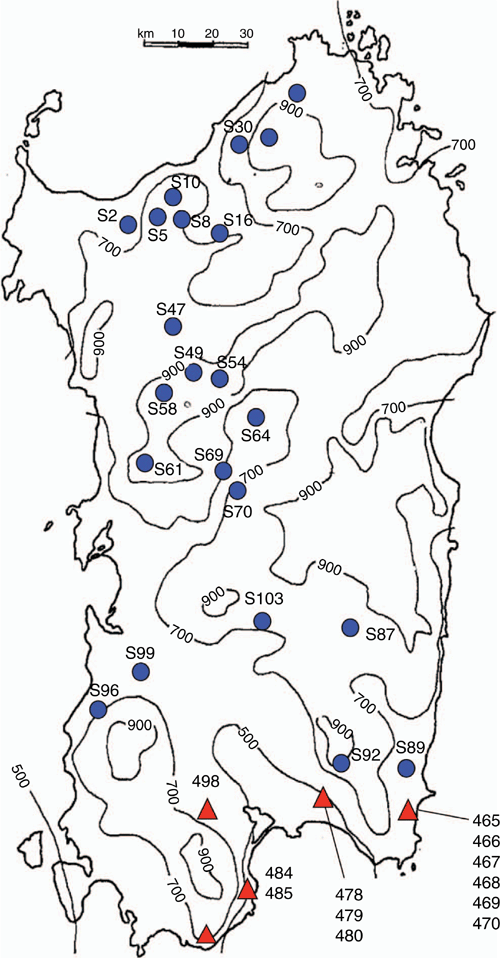
Accessions from the 1994 (Cunningham) and 2000 (Sandral) collections were examined in this study. The 1994 collection emphasised locations in Sardinia with acidic soils (Cunningham et al. 1997). These were largely in central and northern Sardinia, with medium to high rainfall, at altitudes of 540–1100 m (Fig. 1, Table 1). Cunningham et al. (1997) record 24 accessions of P. aquatica as being collected. Of these, 20 were obtained after isolated seed increases were conducted by Agriculture Victoria. The 2000 collection targeted lower altitude and lower rainfall (500–700 mm) areas in the southernmost areas of Sardinia (Fig. 1, Table 1). Of 18 accessions collected, seed of 13 accessions was available for this study. Collection localities in the 1994 expedition were subjected to more grazing pressure than those in the 2000 expedition (Table 1).
|

|
Spaced-plant experiment at Canberra
Sardinian accessions with seed available, 16 from the 1994 collection and 13 from the 2000 collection, were compared with seven accessions from southern Europe (Italy, Greece, Portugal and Spain) and one accession from Algeria residing in the early CSIRO collection, and one accession from Morocco collected by Cunningham et al. (1997). The CSIRO accessions from southern Europe had been selected as being of agronomic interest during screening in Australia in the 1960s. Both accessions from north-western (NW) Africa were selected because they were among the earliest heading and most summer-dormant entries in the study of Culvenor and Boschma (2005); they were included in this study to set a benchmark for earliness and summer dormancy. Two cultivars of the ‘Australian’ type, cvv. Australian and Australian II, and four cultivars of the winter-active type, Holdfast, Holdfast GT, Landmaster and Advanced AT, were also included (Table 1). The Australian-type cultivars are more densely tillered and prostrate with lower winter growth rates than the winter-active cultivars.
Accessions were characterised in a spaced-plant experiment conducted at the CSIRO Ginninderra Experiment Station, Hall, Australian Capital Territory (35°10′58″S, 149°03′31″E; elevation 600 m a.m.s.l.). The site has 690 mm average annual rainfall (AAR) with a slight autumn trough and spring peak. There were 44 entries in the experiment, each represented by a row plot of five plants spaced 0.5 m apart and replicated four times in a randomised block design with a row spacing of 1 m (Table 1). Seedlings were raised in a glasshouse and transplanted to the fertilised field site in late April 2016 and watered by hand to achieve establishment. Rainfall was well above average during winter and early spring 2016, and no further irrigation was required in that year. Growing-season rainfall was below average in 2017, and the experiment received irrigation (25 mm by sprinkler) in July to prevent growth restriction by drought. The experiment was also irrigated (20 mm) on 31 January 2018 as a supplement to a natural rainfall event the day before, allowing observation of summer growth potential.
The experiment was mown to 10–12 cm early in summer 2016 and 2017 to prevent fall of seed, and mown again to 5 cm in June 2017 at the start of the main growth period. Soil tests indicated no need to apply phosphorus (P) fertiliser in 2017 (Colwell P 52 mg/kg). Urea (46% N) was applied in July 2017 at a rate of 100 kg/ha just before irrigation.
Phenological and morphological measurements were made to characterise the germplasm. Seedling size was scored on a scale of 1 (lowest) to 10 (highest) in late August 2016; head emergence date in October–November 2016 and 2017 (at least two heads emerging per plant); reproductive plant height in December 2016 and November 2017; length and width of a typical youngest fully expanded leaf collected in early September 2016 and 2017; average tiller angle from the soil surface estimated visually in early September 2016 and 2017; tiller density scored on a scale of 1 (very sparse) to 7 (very dense) in September 2017; second-year autumn growth scored on a scale of 1 (lowest) to 10 (highest) in May 2017; second-year winter growth scored on a scale of 1 (lowest) to 10 (highest) in September 2017; and area of plant base estimated from measurements made in January 2018. Summer dormancy is an important adaptive mechanism for survival of summer drought in Mediterranean environments (Volaire and Norton 2006). Summer dormancy was assessed as new tillering activity after summer rainfall and the amount of new growth later in summer. Summer tillering activity was visually scored in January 2017 and February 2018 as the number of new tillers shooting from the base of the plant per number of old stems after summer rainfall (first summer) or rainfall plus irrigation (second summer) on a 0–4 scale (adapted from Oram 1984): 0, no new tillers; 1/4, one or two new tillers; 1/2, a few new tillers; 1, half as many new tillers as old stems; 2, number of new tillers » number of old stems; 3, number of new tillers > number of old stems; 4, number of new tillers >> number of old stems. The amount of summer regrowth was visually scored on a 0–7 scale in March 2017 and February 2018.
Agronomic evaluation in drill-rows
Accessions in the Cunningham collection from Sardinia, Morocco and Tunisia were evaluated as part of a commercial breeding program at Meredith, central Victoria (37°49 10 S, 143° 59 15 E; elevation 360 m a.m.s.l.; AAR 690 mm with a late-winter–spring maximum), during 2013–15. All years had below-average annual rainfall (580, 460 and 480 mm, respectively). Results for accessions from Tunisia are not presented here owing to varying levels of contamination from other phalaris species. Rows 2 m long and 0.5 m apart were sown by hand at a rate of ~100 seeds/m in early October 2013.
Rows were scored visually for a range of attributes, only some of which are presented here. Seedling vigour was scored on a scale of 1 (lowest) to 5 (highest) on 10 December 2013; and seasonal row dry matter (DM) yield on a scale of 1 (lowest) to 9 or 10 (highest) during 2014 for autumn (27 May), winter (4 August), spring (24 September) and summer (30 January 2015). DM scores were not calibrated. Heading date was assessed in October and November 2014 and the date of 50% heading in the row was recorded.
Agronomic evaluation in swards
Sardinian accessions from the 1994 collection were evaluated in small sward plots from 1999 to 2003 at three sites in south-eastern Australia established to test the persistence of phalaris germplasm:
-
Rye Park on the Southern Tablelands of New South Wales (34.57°S, 148.93°E), uniform rainfall distribution, AAR 670 mm; soil strongly acidic (pH in 1 : 5 soil : 0.01 m CaCl2 4.0–4.3 to a depth of 0.4 m) and of moderate fertility, aluminium (Al) extracted in 1 : 5 soil : 0.01 m CaCl2 6–14 µg/g to a depth of 0.4 m, Red Sodosol (Australian Soil Classification (ASC), Isbell 2002; Entisol in USDA Soil Taxonomy (ST), Hughes et al. 2018); hard continuous grazing by sheep.
-
Sutton on the Southern Tablelands of New South Wales (35.12°S, 149.28°E), uniform rainfall distribution, AAR 650 mm; soil strongly acidic (pH in CaCl2 4.0–4.2 to a depth of 0.4 m) and of low fertility, CaCl2-extractable Al 14–22 µg/g to a depth of 0.4 m, Australian soil classification Yellow Kurosol (ASC; Ultisol in ST); occasional hard grazing by sheep
-
Bulart in Western Victoria (37.39°S, 141.76°E), winter-dominant rainfall distribution, AAR 637 mm; soil moderately acidic (pH in CaCl2 4.6–5.3 to a depth of 0.4 m), CaCl2-extractable Al 1–5 µg/g to a depth of 0.4 m, Yellow Sodosol (ASC; Entisol in ST); hard rotational grazing by sheep.
Sites on the Southern Tablelands of New South Wales were typical of areas used for wool production where P. aquatica has sometimes persisted poorly. The western Victorian site was in a higher production environment suitable for wool and meat production with a Mediterranean-type climate. Detailed climatic and soil descriptions for Rye Park and Bulart are presented in Culvenor et al. (2007), and for the Sutton site in Culvenor et al. (2004). All sites, but particularly the New South Wales sites, experienced a severe drought in 2002; for example, April–November growing-season rainfall at Rye Park was 250 mm (long-term average 460 mm).
Site management for Rye Park and Bulart is presented in Culvenor et al. (2005, 2009), and for the Sutton site in Culvenor et al. (2004). Briefly, 20 Sardinian accessions and eight control cultivars or breeding lines were sown in 1-m2 plots replicated three times in a row–column design. Controls were two Australian-type cultivars (Australian, Australian II), four winter-active cultivars (Sirosa, Holdfast, Landmaster, Atlas PG), and two winter-active breeding lines (LD97 selected for low summer dormancy, AT98 selected for acid-soil tolerance). The Rye Park and Sutton sites were sown with a seed drill in mid-May 1999 and the Bulart site by hand in June 1999 at a rate of 3 kg/ha, adjusted for germination percentage. The Rye Park site was grazed heavily for short periods up to September 2000, then subjected to continuous grazing mostly at a stocking rate ~12 sheep/ha until June 2003 except for short periods during winter 2001 and early spring 2002. The Sutton site was grazed heavily by mobs of sheep six times in 2000, three times in 2001 and once in the 2002 drought year. The Bulart site was grazed rotationally (~2 weeks on, 4 weeks off) from December 1999 to June 2000, then grazed rotationally (~2 weeks on, 6 weeks off) from August 2000 to June 2003 at an average stocking rate varying from 21 to 30 sheep/ha for the whole period.
Seedling density and DM at Rye Park and Sutton were measured by counting and cutting seedlings in three lengths of 0.2 m placed randomly along rows on 23 September 1999 and weighing the herbage dried at 70°C. At Bulart, all seedlings in a plot were counted on 23 September 1999 and total plot DM was estimated visually on a scale of 1 (lowest) to 9 (highest) calibrated by cutting 22 plots (R2 = 0.80).
Phalaris density was measured as presence of plant base (basal frequency) in cells of wire grids placed over the plots during winter or early spring in each year from 1999 to 2003 at Rye Park and Sutton and during winter in 2002 and 2003 at Bulart. Grids at Rye Park and Sutton contained 54 cells, each 0.15 m by 0.10 m, placed so as to divide each of the six drill-rows into nine 0.10-m segments. A grid of 80 cells, each 0.10 m by 0.10 m, was used at Bulart.
Sward herbage DM at Rye Park was estimated visually on a 1–9 scale calibrated by cutting nine plots (R2 = 0.84) on 9 September 2000 after 4 months of exclusion from grazing, and on a 1–10 scale calibrated by cutting 10 plots (R2 = 0.79) on 25 October 2001 after 2 months of exclusion from grazing. Sward herbage DM at Sutton was estimated visually on a 1–9 scale calibrated by cutting nine plots (R2 = 0.87) in September 2000.
Statistical analyses
All data in the spaced-plant experiment were subjected to analysis of variance (ANOVA) for a randomised block design using Genstat 18 (VSN International, Hemel Hempstead, UK). Analyses with 2 years of data included year, entry, and entry × year effects. Least significant differences at P = 0.05 were calculated. Differences in characteristics between groups of entries formed a priori were analysed by mixed models with group as a fixed effect and entries within groups and replicates as random effects for single-year data. For characters with 2 years of data, group, year, and group × year were fixed effects and entry, entry × year, and replicate within year were random effects. A priori groups were Sardinian 1994 collection, Sardinian 2000 collection, south European accessions, NW African accessions, cultivars Australian-type, and cultivars winter-active type. A principal coordinate analysis (PCA) was performed by using R version 3.6.1 (The R Foundation, Vienna, Austria) on 11 of the 12 characters measured, and a biplot of the first two components was produced using the R package ‘factoextra’ (Kassambara and Mundt 2017). Tiller angle was deleted for clarity in the biplot because it overlaid another character, leaf length, and because its inclusion reduced the variance explained in the PCA. The analysis was restricted to the first three components explaining 90.1% of the variance after inspection of scree plots (Cattell 1966) but only the first two components which explained 85.9% of the variance are presented for simplicity.
Data from the drill-row evaluation were subjected to randomised block ANOVA to determine significant differences among entries. Differences and their significance in the mean values for the Sardinian and Moroccan groups of accessions were analysed by mixed models with origin as a fixed effect and accession and replicate as random effects.
Data from the sward evaluation trials were initially subjected to spatial analysis with random row and column effects modelled as AR1 × AR1 (Gilmour et al. 1997). However, these effects were usually non-significant or marginally significant, so subsequent analyses were conducted as randomised blocks analyses. Comparable basal frequency data for all three sites were available only for 2002 and 2003. Across-sites analyses indicated a highly significant (P < 0.001) entry effect and a much less significant (P < 0.05) site × entry interaction. Comparable yield data for all three sites were available only for seedling DM, for which a highly significant (P < 0.001) entry effect and a non-significant (P > 0.05) site × entry interaction were observed. Otherwise, data were analysed as randomised blocks ANOVA separately for each site to determine significant differences among entries.
Results
Spaced-plant experiment
Variation among entries was significant (P < 0.001) for all phenological and morphological characters measured on spaced plants. For those measured in each of 2 years, the F-ratios for the entry main effect greatly exceeded those for the entry × year interactions even where these were significant at P = 0.01 or 0.001 (Table 2). Because of this, combined with high correlations across years for all characters (Table 2), only means across years are considered here.

|
A biplot of the first two components of a PCA of 11 phenological and morphological characters is shown in Fig. 2. PC3 (not presented here, 4.2% of total variance) was related most closely to area of plant base, and to a lesser extent summer tillering and regrowth. PC1 (66% of total variance) was weighted positively for characters related to DM and plant-size attributes and negatively to tiller density. PC2 (20% of total variance) was weighted most strongly to heading date, base area and summer tillering activity. The biplot clearly separated the modern winter-active cultivars from all other entries on the PC1 axis owing to their larger size, longer and broader leaves, sparser tillering and taller stature. By comparison, both collections of Sardinian accessions were smaller with finer and shorter leaves and a high tiller density. The two collections were separated but with some overlap on the PC2 axis. Accessions from southern Sardinia (2000 collection) were earlier and slightly less summer-active, grouping near an accession from inland Algeria (Entry 38, CPI19280). Accessions from higher altitude and rainfall localities in central and northern Sardinia (1994 collection) tended to be later, more summer-active and more densely tillered. They grouped nearer to the Australian-type cultivars and several accessions from southern Europe. An accession from southern Morocco (Entry 37, M225) was an outlier in the biplot, being very early and low in summer activity with long leaves and sparse tillering. CPI15220 from Spain was also an outlier in the southern European group on the PC2 axis, being earlier and more summer-dormant. It was also relatively tall for this group.

|
Means and ranges of entry groups formed a priori for all attributes are presented in Table 3 and data for selected characters for all entries in Figs 3 and 4. As a group, accessions from the 2000 Sardinian collection were significantly earlier in heading than the 1994 collection group and the southern European accession group, but later than the NW African group (Table 3). The 1994 collection was similar in heading date to the relatively late Australian-type cultivars but nevertheless displayed considerable variation (Fig. 3). The 2000 collection group was similar in heading date to the winter-active group of cultivars. Both groups of Sardinian accessions were lower (P < 0.05) in summer tillering activity and growth than all the commercial cultivars and southern European accessions as a group, and were not significantly (P > 0.05) different from each other. Again, there was considerable variation within each group (Fig. 3). The 1994 Sardinian accession group was higher in summer tillering activity than the NW African group, and the 2000 collection was not different from the NW African group in both summer tillering and growth (Table 3).

|

|

|
Both groups of Sardinian accessions had relatively small seedlings on average, and both were lower in autumn growth than the Australian-type and winter-active cultivar groups (Table 3). All accession groups were similar in winter growth activity to the Australian-type cultivar group except the 2000 Sardinian group, which was lower. All were much lower in winter growth than the winter-active cultivar group (Table 3, Fig. 4). Two accessions in the 1994 Sardinian group were higher in winter growth than other accessions in this group and similar to a high-yielding southern European accession (CPI19344 from Portugal) and the Moroccan accession M225 (Fig. 4). All accession groups and the Australian-type cultivar group had markedly shorter plants and leaves and were more prostrate (lower tiller angle) than the winter-active cultivar group. Both Sardinian groups had narrower leaves than all other groups and were similar in tiller density to the Australian-type cultivars (Table 3, Fig. 4). The 1994 Sardinian group in particular was densely tillered, and significantly (P < 0.05) more so than the 2000 group. There was large variation in base area among Sardinian accessions; however, all but three were lower in base area than cv. Australian (Table 3).
Agronomic evaluation
Drill-rows at Meredith, central Victoria
Performance of the Sardinian and Moroccan accessions from the 1994 collection and of several control cultivars in spaced drill-rows is shown in Fig. 5. The range of heading dates for the Sardinian accessions was similar to that of current cultivars. Compared with Moroccan accessions, the Sardinian accessions, as a group, were significantly (P < 0.001) later in heading, less productive in winter, and more productive in spring and summer but with overlap in these characters (Fig. 5). Distributions of Sardinian and Moroccan accession means were similar for seedling vigour and autumn growth. Although the Sardinian accessions were, in general, less productive in autumn and winter than the current winter-active cultivars, the relative difference appeared smaller than in the spaced-plant experiment, particularly in autumn (Fig. 5). Several Moroccan accessions were similar in winter productivity to the winter-active cultivars when grown as drill-rows, but only one Sardinian accession (S2) approached this level. All cultivars except Holdfast GT were more productive in response to summer rainfall than the group of Sardinian accessions (Fig. 5). The Australian-type cultivar, Australian II, was later in heading than most Sardinian accessions, similar in seedling, autumn and winter growth to the mean of the Sardinian group, and more summer-active (Fig. 5). Spring growth by the Sardinian accessions spanned the range between Australian II and the winter-active cultivars.

|
Multi-site sward plots
Seedling density was considered satisfactory with a mean of 69 seedlings/m2 across the three sites. There was a significant (P < 0.001) difference among entries due to lower establishment by two of the control cultivars (26 seedlings/m2 for Australian, 40 seedlings/m2 for Atlas PG), probably associated with the use of old seed. Basal frequency range was 65–81% among the accessions and 75–79% among the cultivated entries, except for the entries with low seedling numbers (data not shown). The site × entry interaction was not significant (P > 0.05) for seedling density or basal frequency in the establishment year (1999) but was significant (P < 0.05) for basal frequency in 2002 and 2003.
Sardinian accessions were characterised by their high basal frequency at all sites at the end of the study (fifth year, 2003) (Fig. 6). On the acid, moderately fertile soil under continuous stocking at Rye Park, many accessions were significantly higher in frequency than most cultivars despite their initial frequency being similar (Fig. 6a). Eight accessions were higher in basal frequency than cv. Australian, known for its higher persistence under grazing, although this comparison was complicated by the low initial frequency for Australian. Fifteen accessions were higher in frequency than the highest winter-active cultivar. On the very low fertility, acidic soil at Sutton, at least half of the accessions were relatively high in basal frequency in 2003 (Fig. 6b). However, high error variance due to high within-site variation meant that only one accession was higher than all cultivars. Cultivars with low establishment density, Australian and Atlas PG, had high final frequencies relative to their low initial frequency. Plants that did establish may have benefited in survival from this low density. In the higher production, dry-summer environment at Bulart, all entries except for cv. Holdfast were reasonably high in frequency, with the majority of Sardinian accessions being high and similar to cvv. Australian and Australian II (Fig. 6c).

|
Seedling DM on a per-plant basis for the Sardinian accessions across the three sites was similar to the Australian-type cultivars and generally lower than the commercial winter-active cultivars (Fig. 7a). The entry × site interaction was not significant (P > 0.05) for seedling DM. At Rye Park, sward DM for the accessions in September of the second year (2000), following a growth period from mid-May, was similar to, or occasionally less than, that of the Australian-type cultivars and less than that of the winter-active cultivars except for the breeding line LD97 (Fig. 7b). However, in October of the third year (2001), sward yield for the accessions after a period of exclusion from grazing equalled or exceeded that of most cultivars owing to the high tiller density of the accessions (Fig. 7b). Under more difficult soil conditions at Sutton, sward yield by the accessions in September of the second year was similar to that of the Australian-type and winter-active cultivars and breeding lines (Fig. 7c). Two accessions were particularly high in sward yield relative to other entries and similar to the breeding line, AT98, which was bred specifically for tolerance of high-Al acid soils.

|
Discussion
Accessions of P. aquatica from Sardinia were evaluated for basic morphological and phenological variation compared with accessions from southern Europe and NW Africa and commercial cultivars used in Australia. Sardinian accessions displayed similarity to accessions from southern Europe and the Australian-type commercial cultivars; those from warmer areas of southern Sardinia were somewhat earlier in heading and less responsive to summer rainfall (more summer-dormant) than those from cooler and moister areas of Sardinia. Morphologically, the Sardinian accessions were dense, fine, short and prostrate compared with modern winter-active cultivars. Agronomically, an outstanding feature of accessions in the 1994 collection was their high density and persistence when evaluated under typical commercial production conditions in south-eastern Australia. These included conditions that challenge survival of P. aquatica such as heavy grazing pressure, drought and acid soils of low inherent fertility. Yield potential was generally similar to the Australian-type cultivars and lower than present winter-active cultivars at similar densities.
The PCA based on characteristics measured in the spaced-plant experiment suggested that the 1994 collection from central and northern Sardinia, which includes high-elevation areas, mapped most closely with the sample of accessions from southern mainland Europe in terms of heading date, plant morphology and winter growth rate. CPI15220 from Spain was an exception among the southern European group, being relatively tall and early heading. This accession had been previously noted as morphologically like Moroccan types of P. aquatica (Anon. 1967). There was a tendency for accessions in the 1994 collection to be more summer-dormant (lower summer tillering activity and regrowth) than the southern European group, although with considerable overlap. Sardinian accessions from the 1994 collection were generally more summer-active in the drill-row evaluation study than Moroccan accessions collected during the same expedition. Accessions in the 2000 collection from southern Sardinia were similar to the 1994 collection in morphology and winter growth rate but were more summer-dormant and often earlier in maturity time, probably reflecting adaptation to a warmer, drier environment at low altitude in southern Sardinia. They grouped near an accession from Algeria (CPI19280) in the PCA, but were clearly separated from the Moroccan accession M225 because of the contrasting morphology and winter growth rate of the Moroccan accession.
Similarity in aspects of the environment of Sardinia to that of southern Australia was an important motive for undertaking the collections. Compared with commercial cultivars already well adapted to southern Australia, the accessions from the 1994 collection varied in range of heading date from that of the Australian-type cultivars (later) to that of the winter-active cultivars (earlier), whereas the accessions from the 2000 collection were very similar in heading date to the winter-active cultivars in both the spaced-plant and row evaluations. They were also generally later than Moroccan accessions in the row evaluation. Morphologically, they were much more like the Australian-type cultivars than the winter-active cultivars in being short, fine-leaved, densely tillered and relatively prostrate. Sardinian accessions were finer-leaved than the Australian-type cultivars.
Maturity time is an important feature of general adaptation (Cooper and McWilliam 1966; Fjellheim et al. 2014), and Sardinian P. aquatica appears similar to Australian-adapted material in this regard and therefore likely to be adapted to the south-eastern Australian climate. The Sardinian material offers an Australian-type cultivar morphology adapted to heavy grazing with the unique combination of relatively high summer dormancy and a range of maturity times up to very late maturity.
Agronomic evaluation under realistic planting and management conditions is important if new wild material is to be considered for commercial development or incorporated into existing breeding programs. Only the 1994 collection accessions were evaluated under these conditions. Winter is often a time of feed shortage in southern Australia, and winter feed is of high economic value (Ludemann and Smith 2016). The accessions were generally less productive during autumn and particularly during winter than current winter-active cultivars in second-year drill-rows and swards, and their productivity was similar to or less than (2000 collection) that of the Australian-type cultivars. However, there was variation among accessions in both rows and swards, and some of the more productive accessions could be selected for development. Yield during spring in rows was relatively high, falling between the Australian-type cultivar Australian II and some of the winter-active cultivars, and generally higher than Moroccan accessions. Seedling vigour was markedly lower than most winter-active cultivars in the sward evaluation, similar to the Australian-type cultivars. This is unsurprising given the intense selection for seedling vigour in the early breeding of winter-active populations (McWilliam and Latter 1970).
Two of the more productive Sardinian accessions were evaluated by Reed et al. (2008) over 4 years in a Mediterranean-type climate of western Victoria at a locality with 680 mm AAR. Seedling vigour was in the low–intermediate range. Herbage yield over the 4 years was only 5% lower than cv. Australian and 9% lower than the winter-active cv. Holdfast, owing mainly to lower autumn and summer growth.
Improvement of persistence in P. aquatica, and perennial grasses more generally, under various stresses has been a focus for both breeding (Culvenor and Simpson 2014) and management (Kemp et al. 2000). The high density and persistence of the Sardinian cultivars are therefore of commercial interest. All three sites in the sward evaluation received high grazing pressure and experienced periods of drought. As a group, the Sardinian accessions were denser and more persistent after 3 years of grazing than the winter-active cultivars and sometimes cv. Australian, which is renowned for persistence under grazing in high-rainfall areas of southern Australia. Some accessions were also more persistent than the cultivars on a low-fertility soil highly acidic to depth (Sutton site), conditions often resulting in low persistence by P. aquatica (Culvenor et al. 2011). Interestingly, one of the denser accessions at this site was tested for Al tolerance in nutrient solution and found to be relatively sensitive (Requis and Culvenor 2004), and its greater persistence at the Sutton site may have been related to higher grazing tolerance.
Maintenance of high density is desirable for longevity of the sown pasture, soil protection, exclusion of weeds, and other factors related to sustainability, but it does not necessarily lead to the highest levels of animal productivity if combined with low growth rates. Culvenor et al. (1996) observed that cv. Australian maintained a higher density and at times had a higher yield of phalaris herbage than the winter-active cultivars in mixed swards under grazing. However, total pasture DM was higher on the winter-active pastures in autumn–winter, associated with higher phalaris DM per unit basal area and a higher annual grass component. The result was higher wool production from the winter-active cultivars. Combining the high persistence of Sardinian germplasm with selection for higher growth rates, particularly in autumn and winter, would be desirable. Whether growth rates equal to or higher than the Australian-type cultivars can be achieved while maintaining the dense, prostrate morphology of the Sardinian accessions will be a breeding challenge.
Other breeding challenges if the Sardinian accessions are to be exploited are seed production and alkaloids. Efficient seed production is important for a successful commercial cultivar, and modern cultivars of P. aquatica used in Australia have been bred for this. As noted by Oram and Culvenor (1994), natural populations of P. aquatica may not be suitable in this regard because they shed seed very readily and may best be utilised in crosses with lines already selected for seed-production traits such as a densely packed head morphology (McWilliam 1963) or intact rachilla seed retention (McWilliam and Gibbon 1981). The second challenge concerns the level of alkaloids, which are believed to cause toxicity conditions occasionally in livestock grazing P. aquatica, ranging from acute- and chronic-onset ‘staggers’ to very acute ‘sudden death’ (Alden et al. 2014). Although the relationship between toxicity and alkaloid level is not clear (e.g. Oram 1970), cultivars released from the CSIRO program in Australia were nevertheless selected to have lower levels of tryptamine-related alkaloids than cv. Australian, which is relatively high in these alkaloids (Culvenor et al. 2005). Accessions from the Cunningham expedition ranged in levels of tryptamine-related alkaloids from high to moderately low, similar to several winter-active cultivars (Culvenor et al. 2005). They were particularly high in N-methyltyramine, which at one time was hypothesised as a possible cause of sudden death (Anderton et al. 1994) but this has since shown not to be likely (Bourke et al. 2006).
Conclusion
Wild accessions of P. aquatica from the island of Sardinia are likely to be suited to the climate, soils and agricultural management conditions of the permanent-pasture zone of southern Australia and similar environments elsewhere. They offer a range of maturity times combined with higher levels of summer dormancy and a grazing-tolerant morphology which may facilitate expansion of the area of adaptation into hot, drought-prone margins. The ability to provide dense, persistent groundcover could also be of value for soil protection and weed control. Morphologically and in terms of their productivity potential and persistence under grazing, they are much more like the cv. Australian-related group of cultivars than modern winter-active cultivars. Developing cultivars superior to the present cv. Australian type, along with good seed production and acceptable alkaloid levels, will be a breeding challenge. However, variation among the accessions observed in this study will assist in this task.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Acknowledgements
We gratefully acknowledge the collection of original seed by Peter Cunningham, Agriculture Victoria, and Graeme Sandral, NSW Department of Primary Industries. Seed of the Cunningham collection accessions for the sward evaluation was supplied by Bronwen Clark, Agriculture Victoria, and for the row evaluation by USDA-GRIN. We thank Graeme Sandral for supplying original seed for seed increase by CSIRO used in the spaced-plant evaluation. We thank Phillip Veness, Scott McDonald, Terry Mullens and Charlie Veness for technical support, and Rebecca Stutz for conducting the principal coordinates analysis of the spaced-plant trial data. We also thank the landowners on whose properties the sward evaluations were conducted. The spaced-plant experiment was conducted within the Meat & Livestock Australia Phalaris Prebreeding project. The sward evaluation was conducted within a project funded by Australian woolgrowers and the Australian government through Australian Wool Innovation Ltd. Kevin Reed was employed by DPI Hamilton, Victoria, at the time of the multi-site sward study.
References
Alden R, Hackney B, Weston LA, Quinn JC (2014) Phalaris toxicosis in Australian livestock production systems: prevalence, aetiology and toxicology. Journal of Toxins 1, 7Anderton N, Cockrum PA, Walker DW, Edgar JA (1994) Identification of a toxin suspected of causing sudden death in livestock grazing phalaris pastures. In ‘Plant-associated toxins. Agricultural, phytochemical and ecological aspects’. Ch. 49. (Eds SM Colegate, PR Dorling) pp. 269–274. (CAB International: Wallingford, UK)
Anon. (1967) New horizons for phalaris. Rural Research in CSIRO 59, 2–10.
Arrigoni PV (1968) Fitoclimatologia della Sardegna. Webbia 23, 1–100.
| Fitoclimatologia della Sardegna.Crossref | GoogleScholarGoogle Scholar |
Baldini RM (1993) The genus Phalaris L. (Gramineae) in Italy. Webbia 47, 1–53.
| The genus Phalaris L. (Gramineae) in Italy.Crossref | GoogleScholarGoogle Scholar |
Bourke CA, Colegate SM, Culvenor RA (2006) Evidence that N-methyltyramine does not cause Phalaris aquatica-related sudden death in ruminants. Australian Veterinary Journal 84, 426–427.
| Evidence that N-methyltyramine does not cause Phalaris aquatica-related sudden death in ruminants.Crossref | GoogleScholarGoogle Scholar | 17156323PubMed |
Cattell RB (1966) The screen test for the number of factors. Multivariate Behavioral Research 1, 245–276.
| The screen test for the number of factors.Crossref | GoogleScholarGoogle Scholar | 26828106PubMed |
Cooper JP, McWilliam JR (1966) Climatic variation in forage grasses. II. Germination, flowering and leaf development in Mediterranean populations of Phalaris tuberosa. Journal of Applied Ecology 3, 191–212.
| Climatic variation in forage grasses. II. Germination, flowering and leaf development in Mediterranean populations of Phalaris tuberosa.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Boschma SP (2005) Evaluation of phalaris (Phalaris aquatica L.) germplasm for persistence under grazing on the North-West Slopes, New South Wales. Australian Journal of Agricultural Research 56, 731–741.
| Evaluation of phalaris (Phalaris aquatica L.) germplasm for persistence under grazing on the North-West Slopes, New South Wales.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Simpson RJ (2014) Persistence traits in perennial pasture grasses: the case of phalaris (Phalaris aquatica L.). Crop & Pasture Science 65, 1165–1176.
| Persistence traits in perennial pasture grasses: the case of phalaris (Phalaris aquatica L.).Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Simpson RJ (2016) Interaction of plant genotype and management in the persistence of a perennial grass exposed to grazing and soil fertility stresses. Grass and Forage Science 71, 540–558.
| Interaction of plant genotype and management in the persistence of a perennial grass exposed to grazing and soil fertility stresses.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Wood JT, Oram RN (1996) Comparison of winter-active phalaris with the Australian cultivar under rotational grazing. 2. Dry matter on offer, pasture composition and animal production. Australian Journal of Experimental Agriculture 36, 287–297.
| Comparison of winter-active phalaris with the Australian cultivar under rotational grazing. 2. Dry matter on offer, pasture composition and animal production.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Wood JT, Avery AL, Dempsey W, McDonald SE, Ronnfeldt G, Veness PE (2004) Multi-site evaluation on acid soils of a Phalaris aquatica × P. arundinacea × P. aquatica back cross population bred for acid soil tolerance. Australian Journal of Agricultural Research 55, 681–692.
| Multi-site evaluation on acid soils of a Phalaris aquatica × P. arundinacea × P. aquatica back cross population bred for acid soil tolerance.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Reed KFM, McDonald SE (2005) Comparative levels of dimethyltryptamine- and tyramine-related alkaloid toxins in Australian cultivars and some wild populations of Phalaris aquatica. Australian Journal of Agricultural Research 56, 1395–1403.
| Comparative levels of dimethyltryptamine- and tyramine-related alkaloid toxins in Australian cultivars and some wild populations of Phalaris aquatica.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Boschma SP, Reed KFM (2007) Persistence of winter-active phalaris breeding populations, cultivars and other temperate grasses in diverse environments of south-eastern Australia. Australian Journal of Experimental Agriculture 47, 136–148.
| Persistence of winter-active phalaris breeding populations, cultivars and other temperate grasses in diverse environments of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Boschma SP, Reed KFM (2009) Response to selection for grazing tolerance in winter-active populations of phalaris (Phalaris aquatica L.). 1. Persistence under grazing in three environments. Crop & Pasture Science 60, 1097–1106.
| Response to selection for grazing tolerance in winter-active populations of phalaris (Phalaris aquatica L.). 1. Persistence under grazing in three environments.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, McDonald SE, Veness PE, Watson D, Dempsey W (2011) The effect of improved aluminium tolerance on establishment of the perennial grass, phalaris, on strongly acid soils in the field and its relation to seasonal rainfall. Crop & Pasture Science 62, 413–426.
| The effect of improved aluminium tolerance on establishment of the perennial grass, phalaris, on strongly acid soils in the field and its relation to seasonal rainfall.Crossref | GoogleScholarGoogle Scholar |
Cunningham PJ, Graves WL, Chakroun M, Mezni MY, Saidi S, Bounejmate M, Porqueddu C, Reed KFM (1997) Novel perennial forage germplasm from North Africa and Sardinia. Australian Plant Introduction Review 27, 13–46.
Donald CM (1970). Temperate pasture species. In ‘Australian grasslands’. (Ed. R Milton Moore) pp. 303–320. (Australian National University Press: Canberra, ACT)
Fjellheim S, Boden S, Trevaskis B (2014) The role of seasonal flowering responses in adaptation of grasses to temperate climates. Frontiers in Plant Science 5, 431
| The role of seasonal flowering responses in adaptation of grasses to temperate climates.Crossref | GoogleScholarGoogle Scholar | 25221560PubMed |
Francis CM, Gillespie DJ (1981) Ecology and distribution of subterranean clover and Medicago species in Sardinia. Australian Plant Introduction Review 13, 15–25.
Gilmour AR, Cullis BR, Verbyla AP (1997) Accounting for natural and extraneous variation in the analysis of field experiments. Journal of Agricultural, Biological, & Environmental Statistics 2, 269–293.
| Accounting for natural and extraneous variation in the analysis of field experiments.Crossref | GoogleScholarGoogle Scholar |
Hughes P, McBratney AB, Minasny B, Huang J, Micheli E, Hempel J, Jones E (2018) Comparisons between USDA Soil Taxonomy and the Australian Soil Classification system. II. Comparison of order, suborder and great group taxa. Geoderma 322, 48–55.
| Comparisons between USDA Soil Taxonomy and the Australian Soil Classification system. II. Comparison of order, suborder and great group taxa.Crossref | GoogleScholarGoogle Scholar |
Isbell RF (2002) ‘The Australian Soil Classification.’ Revised edn (CSIRO Publishing: Melbourne)
Kassambara A, Mundt F (2017) factoextra: Extract and visualize the results of multivariate data analyses. R package version 1.0.5. The R Foundation, Vienna. Available at: https://cran.r-project.org/web/packages/factoextra/index.html (accessed 8 September 2020).
Kemp DR, Michalk DL, Virgona JM (2000) Towards more sustainable pastures: lessons learnt. Australian Journal of Experimental Agriculture 40, 343–356.
| Towards more sustainable pastures: lessons learnt.Crossref | GoogleScholarGoogle Scholar |
Ludemann CI, Smith KF (2016) A comparison of methods to assess the likely on‐farm value for meat production systems of pasture traits and genetic gain through plant breeding using phalaris (Phalaris aquatica L.) as an example. Grass and Forage Science 71, 66–78.
| A comparison of methods to assess the likely on‐farm value for meat production systems of pasture traits and genetic gain through plant breeding using phalaris (Phalaris aquatica L.) as an example.Crossref | GoogleScholarGoogle Scholar |
McWilliam JR (1963) Selection for seed retention in Phalaris tuberosa L. Australian Journal of Agricultural Research 14, 755–764.
| Selection for seed retention in Phalaris tuberosa L.Crossref | GoogleScholarGoogle Scholar |
McWilliam JR, Gibbon CN (1981) Selection for seed retention in Phalaris aquatica L. In ‘Proceedings XIV International Grassland Congress’. (Eds JA Smith, V Hays) pp. 269–272. (International Grassland Congress)
McWilliam JR, Latter BDH (1970) Quantitative genetic analysis in Phalaris and its breeding implications. Theoretical and Applied Genetics 40, 63–72.
| Quantitative genetic analysis in Phalaris and its breeding implications.Crossref | GoogleScholarGoogle Scholar | 24435673PubMed |
Neal-Smith CA (1955) Report on herbage plant exploration in the Mediterranean region. FAO Report No. 415. Food and Agriculture Organization of the United Nations, Rome.
Nichols PGH, Francis CM (1993) Sardinian subterranean clovers: their importance to Australia. In ‘Proceedings 10th Australian Plant Breeding Conference’. Gold Coast, Qld, Vol. 2. (Eds BC Imrie, JB Hacker) pp. 19–20. (Conference Organising Committee: Canberra)
Nichols PGH, Revell CK, Humphries AW, Howie JH, Hall EJ, Sandral GA, Ghamkhar K, Harris CA (2012) Temperate pasture legumes in Australia: their history, current use, and future projects. Crop & Pasture Science 63, 691–725.
| Temperate pasture legumes in Australia: their history, current use, and future projects.Crossref | GoogleScholarGoogle Scholar |
Nichols PGH, Foster KJ, Piano E, Pecetti P, Kaur K, Ghamkhar K, Collins WJ (2013) Genetic improvement of subterranean clover (Trifolium subterraneum L). 1. Germplasm, traits and future prospects. Crop & Pasture Science 64, 312–346.
| Genetic improvement of subterranean clover (Trifolium subterraneum L). 1. Germplasm, traits and future prospects.Crossref | GoogleScholarGoogle Scholar |
Oram RN (1970) Genetic and environmental control of the amount and composition of toxins in Phalaris tuberosa L. In ‘Proceedings XI International Grassland Congress’. pp. 785–788. (International Grassland Congress)
Oram RN (1984) Inheritance of conditional dormancy in over-summering buds of Phalaris aquatica L. Euphytica 33, 313–319.
| Inheritance of conditional dormancy in over-summering buds of Phalaris aquatica L.Crossref | GoogleScholarGoogle Scholar |
Oram RN, Culvenor RA (1994) Phalaris improvement in Australia. New Zealand Journal of Agricultural Research 37, 329–339.
| Phalaris improvement in Australia.Crossref | GoogleScholarGoogle Scholar |
Oram RN, Ferreira V, Culvenor RA, Hopkins AA, Stewart A (2009) The first century of Phalaris aquatica L. cultivation and genetic improvement: a review. Crop & Pasture Science 60, 1–15.
| The first century of Phalaris aquatica L. cultivation and genetic improvement: a review.Crossref | GoogleScholarGoogle Scholar |
Porqueddu C, Ates S, Louhaichi M, Kyriazopoulos AP, Moreno G, Del Pozo A, Ovalle C, Ewing MA, Nichols PGH (2016) Grasslands in ‘Old World’ and ‘New World’ Mediterranean-climate zones: past trends, current status and future research priorities. Grass and Forage Science 71, 1–35.
| Grasslands in ‘Old World’ and ‘New World’ Mediterranean-climate zones: past trends, current status and future research priorities.Crossref | GoogleScholarGoogle Scholar |
Reed KFM, Nie ZN, Clark B (2008) Field evaluation of phalaris, tall fescue and cocksfoot cultivars and accessions in western Victoria. Australian Journal of Agricultural Research 59, 971–981.
| Field evaluation of phalaris, tall fescue and cocksfoot cultivars and accessions in western Victoria.Crossref | GoogleScholarGoogle Scholar |
Requis J, Culvenor RA (2004) Progress in improving aluminium tolerance in the perennial grass, phalaris. Euphytica 139, 9–18.
| Progress in improving aluminium tolerance in the perennial grass, phalaris.Crossref | GoogleScholarGoogle Scholar |
Trumble HC (1933) The agricultural features of Phalaris tuberosa and allied forms. Journal of the Department of Agriculture, South Australia 37, 400–425.
Trumble HC, Cashmore AB (1934) The variety concept in relation to Phalaris tuberosa and allied forms. The Herbal Review 2, 1–4.
Volaire F, Norton M (2006) Summer dormancy in perennial temperate grasses. Annals of Botany 98, 927–933.