

Habitat for the endangered ephemeral Monotaxis macrophylla (Euphorbiaceae) in New South Wales: how do predictions compare with the evidence?
Stephen A. J. Bell A *
A *
A
Abstract
The endangered ephemeral herb Monotaxis macrophylla (Euphorbiaceae) has limited (~211) records throughout its known range in eastern Australia. In New South Wales (NSW), the 127 occurrences are spread over more than half of the state, and for regulatory purposes this has led to 262 Plant Community Types (PCTs) being predicted as suitable habitat for the species in the BioNet Threatened Biodiversity Data Collection (TBDC). Under the Biodiversity Assessment Method currently in use in NSW, Monotaxis macrophylla is to be specifically addressed for any development site where one or more of the 262 PCTs occur.
To assess how predicted habitats for Monotaxis macrophylla as shown in the TBDC compare with evidence of occurrence.
Examination of all available voucher collections, observations records, and the literature for eastern Australia was undertaken, and reviewed against the New South Wales PCT classification.
Monotaxis macrophylla was found to be affiliated with only 15 PCTs in NSW at best, but only 10 of these are currently included in the list of predicted PCTs for the species. The weight of evidence from existing presence data clearly shows that 10 heath and rocky scrub PCTs identified for Monotaxis macrophylla in NSW comprise 121 (95%) of all NSW records. Analysis of records in Queensland against Regional Ecosystems did not identify additional PCT-affiliated vegetation that might also support the species elsewhere in NSW.
The revised suite of 15 PCTs known or expected to support Monotaxis macrophylla occupies just 14,660 km2 or 1.8% of the State (potentially less if low-confidence PCTs are removed), compared with more than 27,000 km2 or 3.4% for the predicted 262 PCTs.
Currently, if any development is proposed within land that supports one or more of the 262 PCTs linked to Monotaxis macrophylla, appropriately timed targeted surveys are required to discount its presence, or it must be assumed to be present. This review has highlighted that predicted habitats should be evidence based by using known presence data, rather than assumed using broad understandings of possible habitat, and regular revision of predicted habitats within regulatory frameworks for all threatened species is recommended.
Keywords: endangered, ephemeral, evidence, habitat, Monotaxis macrophylla, PCT mapping, regulation, surrogate.
Introduction
Predicting where threatened plants occur is a key challenge in managing the landscapes that support them (Muscatello et al. 2021; Olivares-Rojas et al. 2024), and this need is becoming increasingly important to guide regulatory processes (Robinson et al. 2018; Scheele et al. 2018). For the vast majority of such plants, we know very little about their ecology, how they are adapted to their environment, and the drivers that determine their distribution (Murray et al. 2002; Wang et al. 2019; Wall et al. 2024). As a consequence, decisions made concerning the distribution of data-deficient species within the realm of conservation management and expected habitat are often little more than educated guesses (Scheele et al. 2018; Silcock and Fensham 2018). However, when such presumptive decisions are built into assessments required for regulatory purposes, they can incorrectly estimate suitable habitat, which, in some instances, may be prohibitive in their subsequent offsetting requirements (Briggs 2011; Abdo et al. 2021).
One common method to address this issue is through predictive statistical models such as species distribution models, generalised linear models, generalised additive models and classification tree analysis (Thuiller et al. 2003; Baker et al. 2021; Muscatello et al. 2021), or any of the many other available similar models (see Muñoz and Felicísimo 2004; Elith et al. 2006). Such models have a high reliance on presence–absence data to drive predictions, but this approach can be inappropriate for data-poor species, and often key ecophysiological predictors for plants (e.g. soil pH, nutrient profiles, biotic interactions) are not available across wide areas (Cramer and Verboom 2017; Mod et al. 2016), although this situation is improving (Wang et al. 2019; Lannuzel et al. 2021). Predictive mapping using satellite imagery has also garnered some support in the literature (e.g. Ponce-Fontenla et al. 2021), and there are examples where expert elicitation has been used to assess model accuracy (e.g. Mainali et al. 2020).
Less intensive alternate methods involve the use of species richness, landscape or habitat surrogates to represent the distribution of a species (Faith and Walker 1996; Pharo and Beattie 2001; Lombard et al. 2003; Crisfield et al. 2020; Tälle et al. 2023). The rapid advance in the production of vegetation community mapping across government jurisdictions over recent decades (e.g. DAWE 2020; Roff et al. 2022) has facilitated adoption of these products as surrogates for biodiversity (Ware et al. 2018; Olivares-Rojas et al. 2024). In this sense, Hunter et al. (2016) defined two uses of surrogates, either where one component of an ecosystem is used to represent another, or where a specific element within that ecosystem is used to assist biodiversity management. Use of landscape or habitat features might be considered an example of the first, an indicator surrogate.
Regulatory authorities in several Australian jurisdictions have incorporated the use of indicator surrogates for the assessment of biodiversity across broad areas (e.g. DPE 2023). Under New South Wales (NSW) legislation, the Biodiversity Conservation Act 2016 (BC Act) and Biodiversity Conservation Regulation 2017 (BC Regulation) provide the framework for addressing impacts on biodiversity from development and clearing. This framework requires a proponent to avoid, minimise or offset impacts on biodiversity from proposed threatening actions using the Biodiversity Offsets Scheme (DPE 2023). The Biodiversity Assessment Method (BAM), enabled by the BC Act, provides a standardised method for assessing the biodiversity values at proposed development and biodiversity stewardship sites, and is to be followed when documenting native flora, fauna and their habitats (DPIE 2020). Part of the BAM requires a proponent to identify and map the habitat on a site, and to nominate (with justification) the associated Plant Community Types (PCTs) present. Plant Community Types comprise the NSW State Vegetation Map (DCCEEW 2023), which for eastern parts of the state incorporates the recently revised Eastern New South Wales (ENSW) PCT classification (DPE 2022). Within the BAM, threatened plant species are treated as species credits, and expected suitable habitats for each are as listed in the BioNet Threatened Biodiversity Data Collection (TBDC) (https://www.environment.nsw.gov.au/topics/animals-and-plants/biodiversity/nsw-bionet/about-bionet-atlas/threatened-biodiversity-profiles). When assessing a site, a proponent is obliged to address through survey and/or documentation all the threatened entities listed for the relevant PCTs as shown in the TBDC.
Under the NSW BAM (DPIE 2020), threatened species require assessment against PCTs in which they have been predicted to potentially occur, essentially operating as indicator surrogates. Many of these PCTs exist as broad collections of similar vegetation associations which, because of data limitations, effectively operate at low resolution. Allocation of PCTs to each threatened species for the BAM was partly expert driven through a specific contracted alignment project, backed by a series of prior government-run workshops to gather information on ecology and habitat, but was otherwise compiled by government (often regional) staff following the development of PCTs. In effect, the PCTs listed as surrogates for predicted habitats of threatened plant species were completed in an ad hoc way and based on the often-limited data available at the time of their allocation. They are largely untested yet can carry enormous implications and costs when, under BAM, a proponent is required to prove absence from a proposed development site or accept the liability and purchase credits. Additionally, whereas a species must be assumed present within a development site, the same cannot be applied to a proposed offset site, and the only way to address this is through payment to the NSW Biodiversity Conservation Fund.
This paper uses the endangered ephemeral herb Monotaxis macrophylla as a case study to assess how predictions within the TBDC for suitable habitat, using PCTs as indicator surrogates, align with evidence of occurrence gleaned from databased and specimen records. Predicting occurrence of threatened species always includes an element of risk (Lombard et al. 2003; Wilson et al. 2005), and particularly so for ephemeral species that may be absent from the above-ground flora for extended periods of time. The key question being asked in this paper is how well do predicted habitats for Monotaxis macrophylla shown in the TBDC compare with those known to support it?
Study species
Monotaxis Benth. (Euphorbiaceae) is a small Australian genus of monoecious shrubs or herbs present in all mainland States except Victoria. Halford and Henderson (2002) outlined the taxonomic history of the genus and recognised eight species in their revision, which remains current. Two species (Monotaxis linifolia and Monotaxis macrophylla) are currently known from NSW (PlantNET 2024), with Monotaxis macrophylla listed as Endangered in this State under the BC Act. In Queensland (Qld), the only other Australian State where Monotaxis macrophylla occurs, it is considered of Least Concern under the Nature Conservation Act 1992 (https://apps.des.qld.gov.au/species-search/details/?id=11317). From their global assessment of the conservation status of vascular plant species, Bachman et al. (2024) predicted the extinction risk for this species as ‘not threatened’ at a ’confident’ level, meaning that it is likely to be a species of Least Concern if reassessed under IUCN Red List criteria.
The Type collection for Monotaxis macrophylla was made by Allan Cunningham (with no date), from the ‘summit of Mount Dangar near Moreton Bay’ (Bentham 1873, p. 79). However, there is some confusion over the legitimacy of this location, because there is no Mount Dangar known from the Moreton Bay region of Qld (https://www.qld.gov.au/environment/land/title/place-names). Instead, there is a location by that name ~40 km south-west of Bowen, ~1000 km further to the north, and Cunningham did collect various specimens from McCane’s Bay at Bowen during his third voyage on the Mermaid from Sydney in 1820. However, none of those collections were made by him from this Mount Dangar, nor anywhere away from the immediate coast in that region. There are several Cunningham collections of other species from the Moreton Bay region from between 1828 and 1829, and it is likely that this is where he first collected Monotaxis macrophylla given the number of contemporary collections that exist for the wider Brisbane area. Although Cunningham did ascend and collect various plant specimens from a Mount Dangar in the Hunter Valley of NSW in 1825 (Bell et al. 2022), there is no indication that he saw or collected Monotaxis macrophylla there.
Monotaxis macrophylla occurs sporadically within parts of NSW and Qld, and almost all detections have occurred following the passage of fire (Hunter 1998; Gibson 2002; Bell 2019; Saunders et al. 2024) or other ground-disturbance mechanisms (e.g. fire trail construction; MEL 2012602A, BRI AQ0668464). Hunter (1998) noted that, in Qld, fire did not appear to be a prerequisite; however, this observation may relate to limited ground-disturbance mechanisms occurring outside of fire in rocky habitat. There are also some anecdotal comments (e.g. Voucher collections NSW1098864 and NSW716868) that germination has followed on from extended dry periods without fire or other physical ground disturbance, suggesting breaking of dormancy through non-direct means. Little data are available on pollination and dispersal, although Saunders et al. (2024) documented two separate native bee species from the Halictidae family, and a bee-fly (Bombyliidae: Villiini) as potential pollinators.
Details on habitat supporting Monotaxis macrophylla within the literature are scant. In the western districts of NSW, this includes isolated plants on rocky hillsides in vegetation characterised by Acacia doratoxylon (Ayers et al. 1996; Cunningham et al. 2011) or sandstone ridges (Hunter 2011), and further east on granite heaths of the Northern Tablelands (Hunter and Bruhl 1997; Hunter 1998, 2005; Hunter and Copeland 2007; Saunders et al. 2024), and scrubby heath on sandstone on the Central Western Slopes (Bell and Holzinger 2015; Bell 2019). In Qld, Stanley and Ross (1995) considered stony or sandy soils east of the ranges as favoured habitat. In their review of the genus, Halford and Henderson (2002) summarised known habitat at the time as coastal heathland and open eucalypt forests on deep white coastal sands, and heath, woodland and open forest on rocky mountain slopes and sandstone plateaux, although it is unclear how comprehensive or representative all of these habitats are in NSW.
Methods
Data audit
A review was undertaken of all voucher specimen and observational records of Monotaxis macrophylla contained within the Atlas of Living Australia (ALA; accessed July 2024), incorporating specimen data from Australia’s Virtual Herbarium, the NSW BioNet observational database, and other personal data sources. Data fields associated with all records were examined and details extracted on (1) number and extent of populations, including date of observation, location, and collector, (2) reproductive condition and any other phenology or ecological data, and (3) habitat including notes on fire history. Care was taken to ensure that collections made from the same location but on different dates or by different observers were retained as independent entries. True duplicates (i.e. identical location, date, observer and/or notes fields) and records noting uncertain identification were removed. For data originating from the NSW BioNet database, affiliations with systematic vegetation plot sampling were identified to allow tracing within the ENSW PCT classification (DPE 2022). In those cases, original plot data were examined within the Flora Surveys module of BioNet to glean further information on habitat and associated species. The more expansive PCT classification for the remainder of NSW does not allow such tracing of survey plots.
Meta-populations and bioregions
The analysis dataset was viewed in GIS to delineate meta-populations of Monotaxis macrophylla. A meta-population was defined as a more-or-less connected group of records present within a defined region. Assessment was also made of bioregions in which meta-populations occurred, following the bioregionalisation of Environment Australia (2000).
Plant Community Type
Plant Community Types predicted to provide habitat for Monotaxis macrophylla in NSW were extracted from the NSW BioNet database. These were as contained in the TBDC under the ‘Vegetation type’ tab of the ‘Ecological data’ module. Spatial representation of the geographical spread of these PCTs was assessed in GIS through the NSW State Vegetation Type Map (SVTM) (ver. C2.0M2.0; DCCEEW 2023), which provides the best available comprehensive depiction of PCTs across the State.
Using Monotaxis macrophylla occurrence records, PCTs were identified to delineate known occupied habitat for the species in NSW. This was undertaken by reviewing written notes associated with point records, including associated species, and comparing these with PCT descriptions from DPE (2022), aerial imagery and the PCT mapping of DCCEEW (2023), acknowledging the variable geospatial accuracy provided with each record. Further interrogation of systematic plot data within the Sharing and Enabling Environmental Data (SEED) resource in BioNet was also possible for those records that originated from plot surveys. This enabled identification of records and their component survey plots that had been incorporated into the PCT classification of NSW vegetation (DPE 2022), thereby providing a traceable link to occupied habitat for Monotaxis macrophylla. In these cases, the relevant PCT is shown within the data fields of each plot in SEED and was taken as a point of truth.
All identified PCTs were attributed a confidence level (high, medium, low) on the basis of the nature and extent of supporting evidence for Monotaxis macrophylla. Highest confidence was placed in PCTs where floristic plots contributed directly to the ENSW classification (which comprised >50,000 plots; DPE 2022), or where detailed collection and observation notes defined key aspects of the PCT, or where habitat photographs were viewed, or that were based on the author’s own field knowledge. Medium confidence was attributed to PCTs after viewing aerial imagery with PCT mapping, mostly supported by collector’s habitat notes. Low confidence was placed in PCTs that had very few and often dated records, or had little or no supporting collection and observation notes, or occurred only in broadly defined habitats or PCTs, or where PCT identities were gleaned only from the SVTM. These subjective allocations were categorised into the following four levels to facilitate interpretation: ENSW plots, plots/habitat, PCT inference/habitat, PCT inference only.
A comparison of PCTs was undertaken where Monotaxis macrophylla was identified or strongly suspected of inhabiting specific PCTs (known) against those where the species was assumed to potentially occur as listed in the TBDC (predicted). Examination was also made of those PCTs not present in either dataset, but which, on the basis of similarity of habitat and structure, might also be found to support the species. Notes associated with Monotaxis macrophylla records in Qld were also reviewed to determine whether additional insights on habitat there might also apply to NSW. The Regional Ecosystems (RE) mapping for Qld assisted these assessments, in combination with observer or collector notes associated with each record.
Results
Data audit
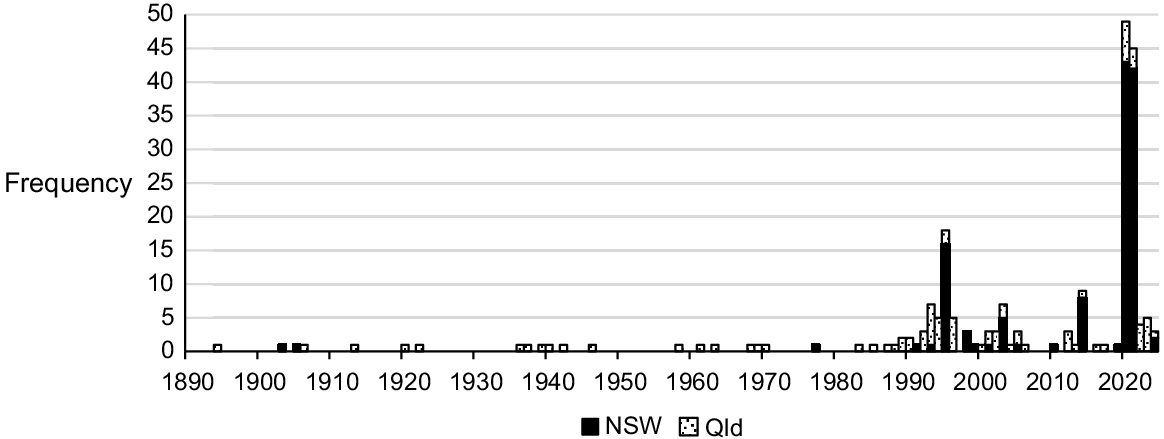
Data auditing found a total of 221 unique records for Monotaxis macrophylla within NSW (127) and Qld (84), and the majority have been made over the past 30 years (Fig. 1). On the basis of all records, 28 meta-populations of Monotaxis macrophylla can be delineated (Fig. 2). The earliest confirmed record from NSW was from Hermitage Plains near Cobar (NSW252064, NSW252069; August 1903), whereas the most recent record was from Boona Mount, also near Cobar (SDFKI0228891; April 2024). For Qld, the first record is a voucher specimen from Bundaberg in 1894 (BRI AQ0204092). The Cunningham collection, supposedly from Moreton Bay in the early-mid 1800s, would predate this record by >60 years if the exact location could be confirmed; however, until such time, this remains unclear. The most recent Qld record is from 2024 at Moogerah Peaks National Park near Boonah (ALA 226145104; June 2024).
History of voucher collections and observations (combined; n = 211) for Monotaxis macrophylla in New South Wales (NSW) and Queensland (Qld). (Source: ALA, accessed July 2024).
Monotaxis macrophylla records (●) and meta-populations within Australia (n = 211). (Source: ALA database, accessed July 2024).
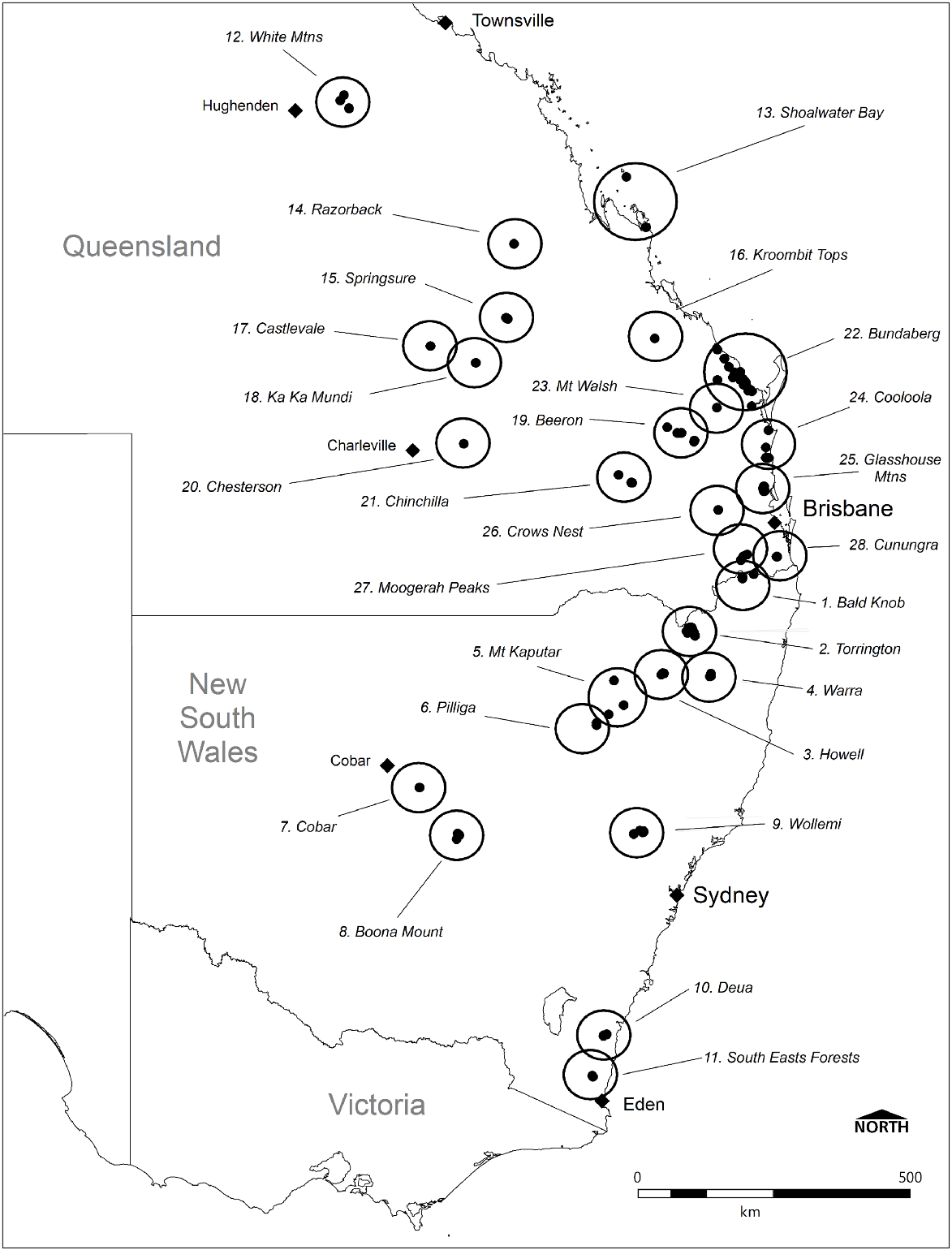
Records are distributed across 10 bioregions within both NSW and Qld. In NSW, Monotaxis macrophylla has been recorded from the South East Corner (SEC), Sydney Basin (SB), Cobar Peneplain (COP), Brigalow Belt South (BBS), Nandewar (NAN), New England Tablelands (NET), and South Eastern Queensland (SEQ) bioregions. In Qld, records exist for SEQ, BBS, Brigalow Belt North (BBN), Desert Uplands (DEU), and Central Mackay Coast (CMC) bioregions.
Many of the 221 unique records are likely to represent repeat visitations to the same populations, often several decades apart. For example, at Miles, west of Brisbane in Qld, Monotaxis macrophylla was first recorded in 1946 (BRI AQ0516180), then again in 1994 (BRI AQ0626680). It was also detected on the top of Mount French (now in Moorgerah Peaks National Park) in 1942 (BRI AQ0204096, BRI AQ0204101) and again 70 years later in 2012 (BRI AQ0908063, MEL 2460354A).
Overall, there are at least 22 populations of Monotaxis macrophylla within conservation reserves. In NSW, these include Deua, Mount Kaputar, Nymadgee, Warra, Wollemi, and South East Forest National Parks, and Torrington State Conservation Area, Willala and Pilliga Aboriginal Areas, and Bornhardtia Voluntary Conservation Area. In Qld, reserved populations occur in White Mountains, Minerva Hills, Mundi, Ka Ka Mundi, Kroombit Tops, Chesterton, Crow’s Nest, Moorgerah Peaks, Mount Walsh, Kinkuna, Woodgate, and Littabella National Parks.
Nearly all voucher specimens within the collated dataset are of fertile material, not unexpected given the short life span of this species and the continual flowering that occurs soon after germination (Hunter 1998). Consequently, Monotaxis macrophylla has been recorded flowering across all months of the year (Fig. 3), and the triggers for flowering appear related to disturbance history rather than any other environmental cue (but this remains untested). There is a slight peak in September, which may coincide with plants maturing after germination following wildfire in the previous Summer.
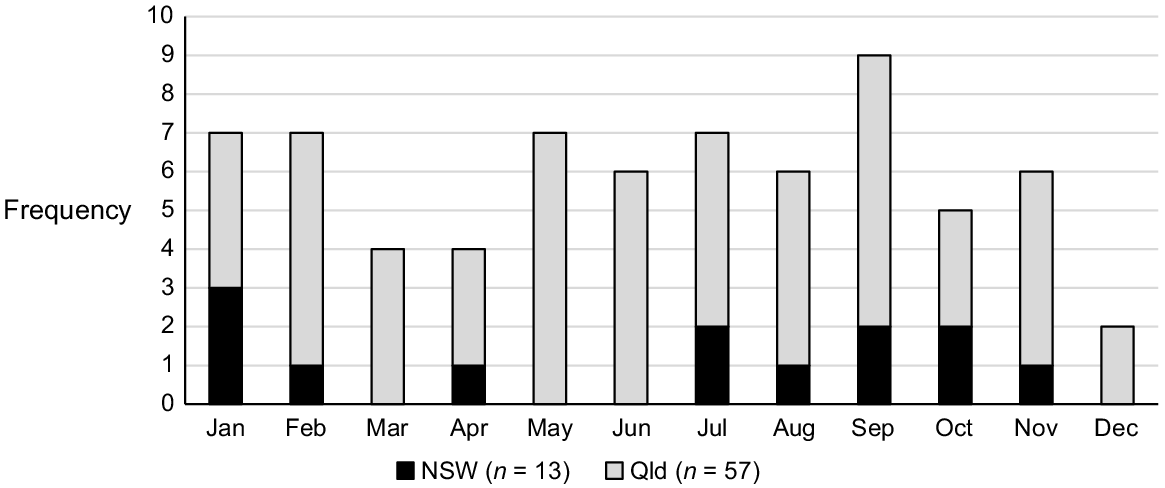
Notes associated with habitat and disturbance for Monotaxis macrophylla as made by the original collectors and observers are summarised in Supplementary Table S1 (NSW) and Table S2 (Queensland).
In general terms, NSW populations of Monotaxis macrophylla are closely linked to rocky and shrubby heath habitats, of volcanic or sedimentary origin. This is clearly the situation within the SEQ, NET, NAN, SB, and much of the SEC bioregions. Of the 11 meta-populations included in NSW, only those in the BBS and COP (~ west of Pilliga) did not include heath or scrub habitats in their descriptions. Similarly, Qld populations predominantly occur in rocky outcrops supporting heath vegetation, or open rocky woodland with a heath understorey. Woodland habitats on sandstones are also apparent in the DEU and BBS bioregions, and wallum heath and related forests on coastal sandy soils in the SEQ bioregion. Both heathlands and forests/woodlands on rhyolite are evident in the SEQ bioregion.
Vegetation types
For Monotaxis macrophylla, the BioNet TBDC showed 262 PCTs as potentially supporting the species in NSW (Table S3). Under the classification system of Keith (2004), these encompass 221 Dry Sclerophyll Forest, 14 Heathland, 14 Semi-arid Woodland, 11 Grassy Woodland, and 1 Rainforest formations (Table 1). The most populous classes are the Western Slopes Dry Sclerophyll Forests (94 PCTs), North-west Slopes Dry Sclerophyll Woodlands (44 PCTs) and the Northern Tableland Dry Sclerophyll Forests (34 PCTs), which combined comprise 66% of the 262 PCTs. Collectively, the 262 PCTs comprise at least 27,009 km2 or 3.4% of the total land area of NSW (Fig. 4), although 64 of the 261 PCTs currently have no mapping within the SVTM. Although approved as part of the NSW classification, these 64 PCTs remain unmapped at present owing to insufficient plot survey data to support them, and/or they cannot be differentiated sufficiently through modelling, unsupervised remote sensing or environmental layers (Roff et al. 2022). This raises the possibility for significantly more land area to comprise PCTs affiliated with Monotaxis macrophylla, but there is no way to determine the magnitude of this.
| Formation | Class | PCTs predicted | PCTs known | |
|---|---|---|---|---|
| Dry Sclerophyll Forests (shrub/grass subformation) | North-west Slopes Dry Sclerophyll Woodlands | 44 | – | |
| New England Dry Sclerophyll Forests | 12 | – | ||
| Pilliga Outwash Dry Sclerophyll Forests | 4 | – | ||
| Hunter–Macleay Dry Sclerophyll Forests | 1 | – | ||
| Northern Gorge Dry Sclerophyll Forests | 1 | – | ||
| Dry Sclerophyll Forests (shrubby subformation) | Western Slopes Dry Sclerophyll Forests | 94 | 2A | |
| Northern Tableland Dry Sclerophyll Forests | 34 | – | ||
| Yetman Dry Sclerophyll Forests | 17 | – | ||
| Southern Tableland Dry Sclerophyll Forests | 4 | – | ||
| North-west Slopes Dry Sclerophyll Woodlands | 3 | – | ||
| Sydney Hinterland Dry Sclerophyll Forests | 2 | – | ||
| New England Dry Sclerophyll Forests | 1 | – | ||
| North Coast Dry Sclerophyll Forests | 1 | – | ||
| Northern Escarpment Dry Sclerophyll Forests | 1 | – | ||
| South Coast Sands Dry Sclerophyll Forests | 1 | – | ||
| South East Dry Sclerophyll Forests | 1 | 1B | ||
| Grassy Woodlands | New England Grassy Woodlands | 6 | – | |
| Coastal Valley Grassy Woodlands | 3 | – | ||
| Western Slopes Grassy Woodlands | 2 | – | ||
| Heathlands | Northern Montane Heaths | 14 | 7 | |
| Sydney Montane Heaths | – | 1 | ||
| Southern Montane Heaths | – | 1 | ||
| Semiarid Woodlands (shrubby subformation) | Inland Rocky Hill Woodlands | 14 | 1 | |
| Western Peneplain Woodlands | – | 1 | ||
| Rainforests | Dry Rainforests | 1 | – | |
| Southern Warm Temperate Rainforests | – | 1C | ||
| Not applicable | [Rocky cliffs, major rock outcrops etc.] | 1 | – | |
| Total | 262 | 15 |
(Source: NSW BioNet Threatened Biodiversity Data Collection and ALA, accessed July 2024). See full list of PCTs in Table S3.
Predicted NSW Plant Community Types (PCTs) associated with Monotaxis macrophylla. Mapping depicts selected PCTs from the NSW BioNet SVTM, as delineated within the TBDC and shown with known records from the ALA database (accessed July 2024).
Of the 127 unique records of Monotaxis macrophylla existing in NSW, 20 could be traced directly to a PCT as used in the ENSW PCT classification. In addition, a further eight could be confidently aligned with PCTs through habitat descriptions and/or proximity to floristic plots included in the ENSW classification, and 18 records could be assigned on the basis of habitat descriptions and viewing of PCT mapping. Eighty records from Torrington SCA were assigned on the basis of descriptions in Saunders et al. (2024) and PCT mapping, whereas one record (Rocklea North BSA) was allocated by field workers associated with the finding (AMBS 2022). The 15 assigned PCTs are shown within Keith (2004) formations and classes in Table 1, summarised in Table 2, and their spatial distribution is depicted in Fig. 5.
| Meta-population | State | PCT/RE | |
|---|---|---|---|
| 1. Bald Knob | NSW | Woodenbong Plugs Rocky Scrub (PCT 3856) | |
| Qld | Heath and Rock Pavement with Scattered Shrubs or Open Woodland on Cainozoic Igneous Hills and Mountains (RE12.8.19) | ||
| 2. Torrington | NSW | New England Rockplate Shrubland (PCT 3854) | |
| Northern New England Rock Outcrop Shrubland (PCT 4128) | |||
| Western New England Rocky Granite Shrubland (PCT 4132) | |||
| 3. Howell | NSW | Western New England Rocky Granite Shrubland (PCT 4132) | |
| 4. Warra | NSW | Eastern New England Leucogranite Mallee Scrub (PCT 3827) | |
| 5. Mount Kaputar | NSW | Heathy Outcrop Shrublands on Volcanic Sediments of the Nandewar Bioregion and Brigalow Belt South Bioregion (PCT 520) | |
| Mount Kaputar Kunzea–Five Star Heath–Spur–wing Wattle Shrubland (PCT 521) | |||
| 6. Pilliga | NSW | A White Bloodwood–Red ironbark–Black Cypress Pine Shrubby Sandstone Woodland of the Pilliga Scrub and Surrounding Regions (PCT 405) | |
| 7. Cobar | NSW | A Poplar Box–Gum Coolabah–White Cypress Pine Shrubby Woodland (PCT 103) | |
| 8. Boona Mount | NSW | Dwyer’s Red Gum–White Cypress Pine–Currawang Low Shrub–Grass Woodland (PCT 184) | |
| 9. Wollemi | NSW | Western Hunter Rocky Scrub (PCT 3784) | |
| Wollemi Rockplate Scrub (PCT 3866) | |||
| 10. Deua | NSW | Far South-east Mountain Rock Scrub (PCT 3870) | |
| A South Coast Temperate Gully Rainforest (PCT 3045) | |||
| 11. South East Forests | NSW | Brogo Scarp Mallee Scrub (PCT 3642) | |
| 12. White Mountains | Qld | Acacia shirleyi Open Forest on Sandstone Ranges (RE 10.10.1) | |
| 13. Shoalwater Bay | Qld | A Corymbia spp. and/or Eucalyptus spp. Open Forest to Low Woodland (RE 8.2.8a) | |
| A Allocasuarina littoralis and/or Leptospermum neglectum and/or Leptospermum polygalifolium and/or Baeckea frutescens Dwarf Shrubland to Low Open Forest (RE 8.2.3d) | |||
| 14. Razorback | Qld | Shrubland to Low Open Forest on Cainozoic Igneous Rocks (RE 11.8.7) | |
| 15. Springsure | Qld | Shrubland to Low Open Forest on Cainozoic Igneous Rocks (RE 11.8.7) | |
| 16. Kroombit Tops | Qld | Eucalyptus decorticans, Lysicarpus angustifolius ± Eucalyptus spp., Corymbia spp., Acacia spp. (RE 11.10.4) | |
| 17. Castlevale | Qld | Acacia shirleyi or A. catenulata Open Forest on Coarse-grained Sedimentary Rocks (RE 11.10.3) | |
| 18. Ka Ka Mundi | Qld | A Eucalyptus cloeziana ± E. melanoleuca ± Corymbia bunites ± E. sphaerocarpa Woodland to Open Forest (RE 11.10.13a) | |
| 19. Beeron | Qld | Corymbia citriodora or Eucalyptus crebra Woodland on Cainozoic Lateritic Duricrust (RE 11.7.6) | |
| Corymbia spp., Eucalyptus baileyana, E. dura, E. exserta Woodland on Igneous Rocks (RE 11.12.20) | |||
| Open Forest Complex with Corymbia citriodora subsp. variegata on Subcoastal Remnant Tertiary Surfaces. Usually deep red soils (RE 12.5.1) | |||
| 20. Chesteron | Qld | Acacia shirleyi or A. catenulata Open Forest on Coarse-Grained Sedimentary Rocks (RE 11.10.3) | |
| 21. Chinchilla | Qld | Acacia spp. Woodland on Cainozoic Lateritic Duricrust (RE 11.7.2) | |
| A Acacia aprepta Shrubland (RE 11.7.5b) | |||
| 22. Bundaberg | Qld | Banksia aemula Low Open Woodland on Dunes and Sand Plains. Usually deeply leached soils (RE 12.2.9) | |
| Banksia aemula Low Woodland on Alluvial Plains usually near Coast (RE 12.3.14) | |||
| Eucalyptus latisinensis and/or Banksia aemula Low Open Woodland on Complex of Remnant Tertiary Surface and Tertiary Sedimentary Rocks (RE 12.5.10) | |||
| Corymbia intermedia ± Lophostemon confertus ± Banksia spp. ± Callitris columellaris Open Forest on Beach Ridges usually in Southern Half of Bioregion (RE 12.2.5) | |||
| A Gahnia sieberiana, Empodisma minus, Gleichenia spp. Closed Sedgeland in Coastal Swamps (RE 12.2.15) | |||
| A Eucalyptus tereticornis Woodland on Quaternary Alluvium (RE 12.3.3) | |||
| A Melaleuca quinquenervia ± Eucalyptus tereticornis, Lophostemon suaveolens, Corymbia intermedia Open Forest on Coastal Alluvial Plains (RE 12.3.6) | |||
| A Eucalyptus tereticornis ± Eucalyptus siderophloia, Corymbia intermedia Open Forest on Alluvial Plains usually near Coast (RE 12.3.11) | |||
| 23. Mount Walsh | Qld | Shrubland of Rocky Peaks on Mesozoic to Proterozoic Igneous Rocks (RE 12.12.10) | |
| 24. Cooloola | Qld | Heath and Rock Pavement with Scattered Shrubs or Open Woodland on Cainozoic Igneous Hills and Mountains (RE 12.8.19) | |
| A Tall Open Forest often with Eucalyptus resinifera, E. grandis, E. robusta and Corymbia intermedia on Sedimentary Rocks, usually Coastal (RE 12.9-10.1) | |||
| 25. Glasshouse Mountains | Qld | Heath and Rock Pavement with Scattered Shrubs or Open Woodland on Cainozoic Igneous Hills and Mountains (RE 12.8.19) | |
| A Eucalyptus racemosa subsp. racemosa Woodland on Remnant Tertiary Surfaces (RE 12.5.3) | |||
| 26. Crows Nest | Qld | Eucalyptus dura Woodland usually on Rocky Peaks on Mesozoic to Proterozoic Igneous Rocks (RE 12.12.9) | |
| 27. Moogerah Peaks | Qld | Heath and Rock Pavement with Scattered Shrubs or Open Woodland on Cainozoic Igneous Hills and Mountains (RE 12.8.19) | |
| A Shrubby Woodland with Eucalyptus racemosa subsp. racemosa or E. dura on Cainozoic Igneous Rocks (RE 12.8.20) | |||
| 28. Canungra | Qld | Eucalyptus racemosa subsp. racemosa ± Lophostemon confertus, Syncarpia glomulifera, Eucalyptus acmenoides Woodland to Open Forest usually on Rocky near coastal areas on Mesozoic to Proterozoic Igneous Rocks (RE 12.12.14) |
Known NSW Plant Community Types associated with Monotaxis macrophylla. Mapping depicts selected PCTs from the NSW BioNet SVTM, as delineated through the present study and shown with known records from the ALA database (accessed July 2024).
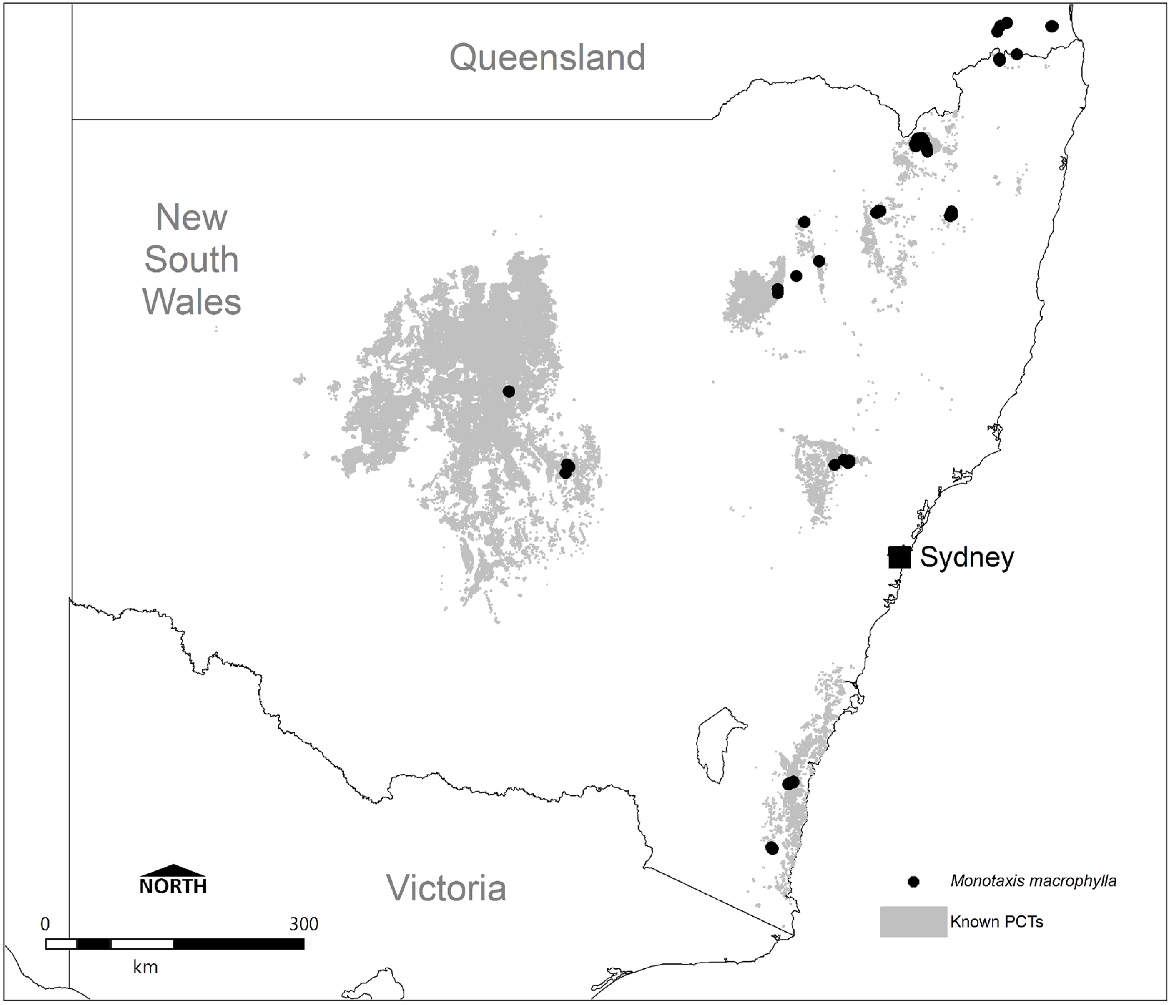
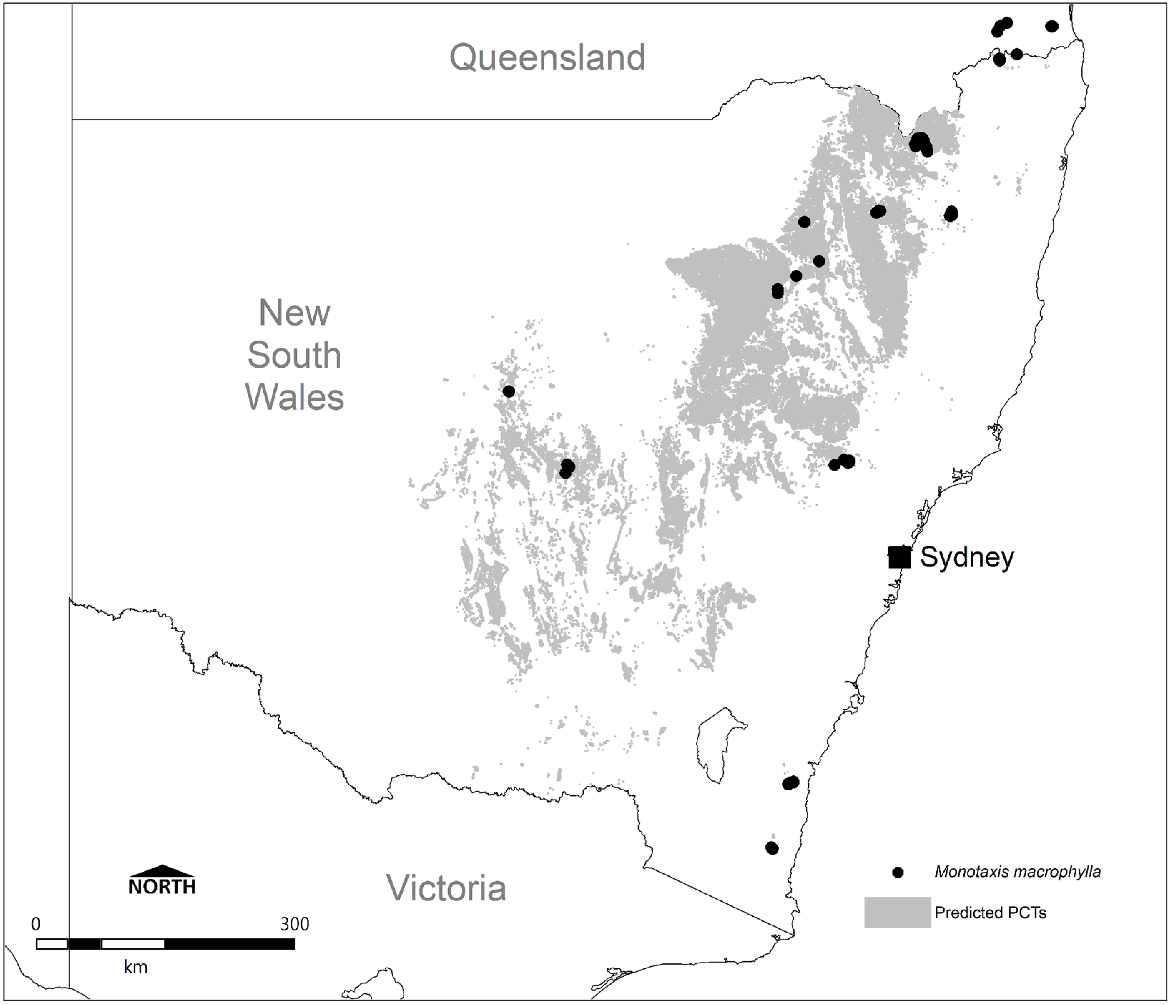
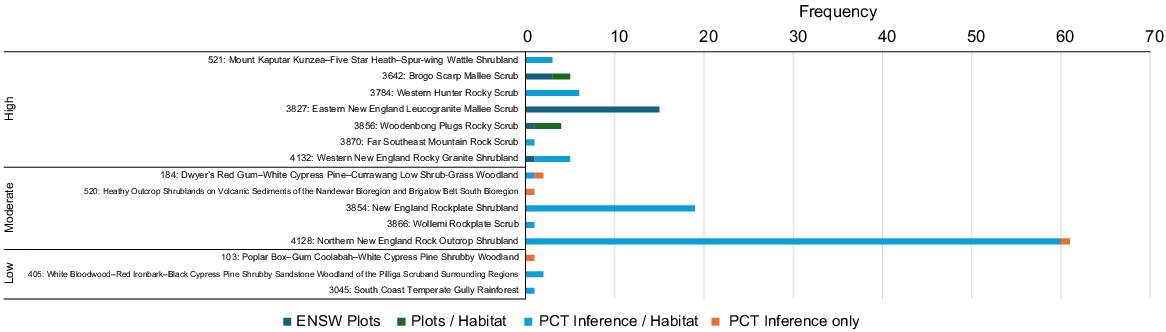
On the basis of occurrence data, seven PCTs are considered with high confidence to represent important habitat for the species in NSW, and a further five PCTs are of moderate confidence and may also be important, although this is less clear (Fig. 6). Driven by frequency of records alone, Northern New England Rock Outcrop Shrubland (PCT 4128), New England Rockplate Shrubland (PCT 3854), Eastern New England Leucogranite Mallee Scrub (PCT 3827) and Western New England Rocky Granite Shrubland (PCT 4132) are the most commonly represented PCTs, collectively comprising 100 of the 127 (79%) records in NSW. These habitats consequently encompass the major stronghold of records from the Northern Tablelands between Armidale and Inverell.
Frequency of NSW Monotaxis macrophylla records traceable to PCT. Categorised into high, medium or low confidence on the basis of direct inclusion in the ENSW PCT classification, or inferred through interrogation of PCT mapping and descriptions, habitat notes linked to records or field knowledge (n = 127).
In the far north of NSW, near the Qld border, the Monotaxis macrophylla population is represented by Woodenbong Plugs Rocky Scrub (PCT 3856), whereas on the far south coast it is Brogo Scarp Mallee Scrub (PCT 3642), Far Southeast Mountain Rock Scrub (PCT 3870), and potentially South Coast Temperate Gully Rainforest (PCT 3045). The 1991 record associated with the last PCT was made inland from Moruya, and notes show occurrence 2–3 months after construction of a ‘cleared fire break through dry rainforest’ of Backhousia myrtifolia, Phebalium squameum, Eriostemon trachyphyllus (syn. Philotheca trachyphylla), Acacia silvestris, and Parsonsia straminea. Review of mapping at this location suggests PCT 3045 to be appropriate, although Monotaxis macrophylla has clearly appeared following fire mitigation clearing, and it is questionable how representative undisturbed rainforest habitats are for this species. Given the mention of Acacia silvestris and Philotheca trachyphylla, it is possible that vegetation more similar to Brogo Wattle Scrub (PCT 3669), which is characterised by both of these species, once occurred here prior to rainforest development.
In central eastern NSW in and around the upper Hunter Valley, known populations occur within Western Hunter Rocky Scrub (PCT 3784) and Wollemi Rockplate Scrub (PCT 3866). For the Mount Kaputar area, Heathy Outcrop Shrublands on Volcanic Sediments of the Nandewar Bioregion and Brigalow Belt South Bioregion (PCT 520) and Mount Kaputar Kunzea–Five Star Heath–Spur-wing Wattle Shrubland (PCT 521) support the species, both heathlands occurring on elevated hills above the undulating plains. The eastern Pilliga is represented by only two records, from 1995 and 2010, attributable to White Bloodwood–Red Ironbark–Black Cypress Pine Shrubby Sandstone Woodland of the Pilliga Scrub and Surrounding Regions (PCT 405).
The central western districts of NSW are confounded by few records (1903, 1977, 2024), but important PCTs here include Dwyer’s Red Gum–White Cypress Pine–Currawang Low Shrub–Grass Woodland (PCT 184) near Boona Mount, and potentially Poplar Box–Gum Coolabah–White Cypress Pine Shrubby Woodland (PCT 103) near Cobar. Metcalfe et al. (2003) describe low woodlands of Eucalyptus dwyeri on outcropping granite on and around Boona Mount, corroborating collection notes for that location. Further clarification of habitat in these more western areas is required.
The weight of evidence from review of existing presence data clearly shows that the 10 heath and rocky scrub PCTs identified for Monotaxis macrophylla in NSW comprise 121 (95%) of all NSW records. Of the remaining data, five records (4%) occur west from the Pilliga, typically in rocky areas within western woodlands, and one (1%) occurs close to or in disturbed rainforest on the South Coast.
PCTs predicted to support Monotaxis macrophylla in NSW bear little resemblance to those known to harbour actual populations. Under NSW regulatory requirements, this species is predicted to potentially occur in 262 PCTs within six formations and 21 classes presented in the Keith (2004) classification, yet evidence from documented populations shows that just 15 PCTs from four formations and eight classes are represented. There is no evidence for the species to be present in any of the 62 PCTs classified within Dry Sclerophyll Forests (shrub/grass subformation) or 11 PCTs within the Grassy Woodlands formation. There are substantially fewer PCTs (3 vs 159) represented in the Dry Sclerophyll Forests (shrubby subformation), and two of these are structurally heathlands and would be better reclassified as such. The 14 Northern Montane Heaths from the Heathlands formation currently predicted are halved in number, and new additions in this formation previously not expected to carry the species include Sydney Montane Heath and Southern Montane Heath classes. One additional class is also evident in the Semiarid Woodlands (shrubby subformation), but the Dry Rainforest class formerly included from the Rainforest formation has been replaced by Southern Warm Temperate Rainforests (based on a single, perhaps opportunistic, occurrence).
Of the 15 PCTs shown to support populations of Monotaxis macrophylla in NSW on the basis of database records, 10 are already included in the 262 PCTs currently predicted for this species. Six of these are supported by the most recent ENSW PCT classification, where four PCTs include floristic plots that recorded Monotaxis macrophylla. However, despite evidence from existing records, the remaining five PCTs (33% of those identified here) are not currently considered suitable habitat for Monotaxis macrophylla within current regulatory guidelines. These comprise Western Hunter Rocky Scrub (PCT 3784) and Far Southeast Mountain Rock Scrub (PCT 3870), both with high confidence; Wollemi Rockplate Scrub (PCT 3866), with moderate confidence; and Poplar Box–Gum Coolabah–White Cypress Pine Shrubby Woodland (PCT 103) and South Coast Temperate Gully Rainforest (PCT 3045), with low confidence. Three of these new PCTs are heathlands, and together with the eight other heath- or scrub-identified PCTs highlight the importance of the Heathlands formation for Monotaxis macrophylla. Indeed, a clear difference between those PCTs predicted to support Monotaxis macrophylla in the TBDC and those known to support the species is the dominance of Heathlands as habitat. This formation was predicted to comprise just 5% of listed PCTs, but on the basis of known presence this has increased to 56% (or 69% if the two suspected misclassified Dry Sclerophyll Forests PCTs are included). As noted earlier, by weight of number alone, 95% of all Monotaxis macrophylla records occur within the Heathland formation, reinforcing this importance.
Collectively, the revision of NSW PCTs known to support Monotaxis macrophylla down from 262 to 15 has also reduced the total land area providing habitat. Currently, at least 27,009 km2 or 3.4% of the total land area of NSW has been mapped as supporting the 262 predicted PCTs, but this reduces to 14,660 km2 (1.8%) when only the 15 known PCTs are considered. Furthermore, disregarding the three low-confidence PCTs (103, 405, 3045), which are supported by just four records, further reduces this area to 1810 km2 or 0.2% of the State. Although there remains the possibility that additional affiliated PCTs may well be found to support the species, particularly other rocky heath, scrub or woodland habitats, on the basis of currently available evidence, the reduction in PCTs and total land area known to support Monotaxis macrophylla is substantial.
As for NSW, rocky heath and scrub was the most commonly reported habitat in Qld, although in western districts (DEU, BBS and BBN) dry woodland and/or forests on rocky sandstone landscapes were also evident in the data (Table 2). The latter are commonly associated with low woodlands typified by various Acacia species, but lancewood (Acacia shirleyi) is usually prominent, and eucalypts may be widely scattered. Of particular interest also was quite different habitat along the coast (SEQ) in deeper aeolian sand deposits (commonly wallum, Banksia aemula), a habitat type present but not yet recorded for NSW populations of Monotaxis macrophylla. Supplementary Table S4 provides summary descriptions for all identified REs relating to Qld populations, as described by Queensland Herbarium (2024).
Occupied habitats for Monotaxis macrophylla in Qld, on the basis of currently available data attributed to REs with some level of confidence, include the following: (1) rocky heath and scrub on a variety of geological substrates but particularly of volcanic and igneous origin, in near-coastal locations (34% of records); (2) wallum heath, scrub and heathy woodlands on whitish sand plains and dunes, along the coast (21%); (3) Acacia shrublands and low woodlands on sandstone, lateritic and other sedimentary substrates, in inland locations (17%); and (4) low and sparsely vegetated open woodlands on igneous substrates, in inland locations (6%). Review of dominant and characteristic species for each of these shows that none supports floristic associations such as the 262 PCTs currently predicted to support populations of Monotaxis macrophylla in NSW.
Discussion
Predictive species distribution models are an important component of conservation biology, and there are numerous analytical methods and algorithms where these have been successfully deployed (Muñoz and Felicísimo 2004; Guisan and Thuiller 2005; Elith et al. 2006; Wisz et al. 2008). However, their use tends to be less effective for species with few occurrence records (Stockwell and Peterson 2002; Wisz et al. 2008) or when specific ecophysiological plant requirements are neglected or unavailable (Mod et al. 2016; Cramer and Verboom 2017), and, hence, incorporation of landscape surrogates becomes more attractive. Such an approach has been undertaken for species conservation elsewhere (e.g. Lombard et al. 2003), and in the design of comprehensive reserve networks using landscape units or vegetation communities (e.g. Woinarski et al. 1996; Wilson et al. 2005), and currently in NSW, this is also the situation for predicting threatened species habitat within regulation.
Adopting habitat characteristics as a surrogate for likely presence of threatened plants is particularly appropriate for taxa where numerous records exist and there is good knowledge on the ecological processes that govern distribution (Lombard et al. 2003; Crisfield et al. 2020; Tälle et al. 2023; Olivares-Rojas et al. 2024). However, like Monotaxis macrophylla, such indicator surrogates (Hunter et al. 2016) perform poorly when distribution data are limited, or target species occupy specific niches. Landscape units of insufficient resolution are consequently limited in their value to predict distribution in some species (Guisan and Thuiller 2005), and this seems to be the case for Monotaxis when based on PCTs. Nevertheless, in the absence of better alternatives, predicting occurrence and distribution even with low-resolution landscape units is often the only option.
However, within the NSW BAM (DPIE 2020) this process of allocating predicted threatened species habitat via PCTs as indicator surrogates is unclear and appears inconsistent, and there is no publicly available documentation to show how this was accomplished. Acknowledging the enormity of this process to allocate several hundred threatened species across over 1800 PCTs, it is important that clear documentation and regular review be undertaken moving forward. Current PCT allocation appears more logical for species occupying known-small rather than unknown-large distributions, these relying more on recorded presence rather than predictions. For example, fewer PCTs tend to be associated with well-known or highly restricted species (e.g. three for Wollemia nobilis; five for Grevillea caleyi), but many more are listed for less well-known or widespread species (e.g. 162 for Tylophora linearis; 262 for Monotaxis macrophylla; 371 for Rhodamnia rubescens). Some species with many affiliated PCTs are justifiable, such as for the widespread and relatively common Rhodamnia rubescens now severely affected by myrtle rust and occurring in a wide range of habitats across eastern NSW. However, others are seemingly over-represented by the PCTs assigned to them, perhaps driven by ignorance and a conservative approach. Plant biology, field knowledge and available data will clearly influence how many PCTs should be allocated to a species, yet the process undertaken for the current assignment appears to have overlooked available resources (e.g. database records, herbarium collections) in the interests of expediency. One outcome of this is that proponents for a potential development where predicted PCT alignment results in offsetting obligations must accept or provide substantial evidence to the contrary.
Outside of NSW, review of habitat for Qld populations of Monotaxis macrophylla provides important additional insights, which may lead to new habitats and populations becoming uncovered and recognised in NSW. As in NSW, habitats and REs supporting the species are largely dominated by rocky heath and scrub on various geological substrates, or Acacia-dominated shrublands or sparsely vegetated woodlands on rocky inland geologies. However, a large proportion of records occur on coastal sand deposits in habitats characterised by wallum (Banksia aemula), a landscape that also occurs along much of the NSW coast, yet no populations of Monotaxis macrophylla are known there. This is somewhat surprising, given the higher populations of professional and amateur botanists along the coastal zones who frequent these landscapes, and perhaps explains why wallum habitats comprise the second-most important (by frequency of records) habitat for Monotaxis macrophylla in Qld. A small number of Qld records also exist in open forests and woodlands associated with, but not in, rocky or sandy landscapes, although none of these carries floristic associations such as the 247 NSW PCTs predicted to carry Monotaxis populations, but not supported by evidence.
It is important to consider the implications of this critical review of known habitat for Monotaxis macrophylla against predicted habitats currently included in the BAM regulatory process (DPIE 2020). Currently, if any development or other activity is proposed within land that supports one or more of the 262 PCTs linked to Monotaxis macrophylla, appropriately timed targeted surveys are required to discount its presence, or it must be assumed to be present. Additionally, the TBDC strongly recommends that an expert report (as defined under BAM) be prepared for any proposed development site where the relevant PCTs occur, or analysis of the soil seed bank be undertaken to discount species presence. Currently, there are no approved biodiversity experts for Monotaxis macrophylla within NSW, therefore precluding preparation of an expert report, and successful analysis of the soil seed bank to confirm the species presence is yet to be demonstrated as a viable detection method.
Surveys for the species in accordance with the TBDC are often not possible because of the absence of fire or ground disturbance within 6 months of the survey or are outside of the limited months in which the surveys need to occur (stipulated to be undertaken within the months of January, February, and August–December). These requirements severely constrain survey opportunities, and although the requirement for fire/ground disturbance is valid, evidence from collections and observational records presented here suggest that survey at any time of the year should be permissible. The additional survey and assessment requirements that are triggered for disturbance to one or more of the 262 PCTs linked to Monotaxis macrophylla, as well as the potential implications for assuming presence, highlights the need for predicted habitats to be evidence based using known presence data, rather than assumed using broad understandings of possible habitat.
Further clarification of the extent and habitat of Monotaxis macrophylla is required in some areas. Within NSW, the most westerly record occurs on the North Western Plains in the vicinity of Hermitage Plains near Cobar. This record comprises two collections by Baeuerlen from 1903 (NSW252064, NSW252069), accepted by Halford and Henderson (2002) in their revision of the genus, but there have been no other observations or collections during the subsequent 120 years. In effect, this adds nearly 11,000 km2 of potential habitat (PCT 103: Poplar Box–Gum Coolabah–White Cypress Pine Woodland) to the feasible distribution of Monotaxis macrophylla within NSW. This is substantial, and within the context of the other 14 PCTs delineated as known habitat in this study, comprises 74% of the total land area. New targeted surveys in areas following fire or other ground disturbances are required to substantiate this habitat.
Similarly, the single 1977 collection by Milthorpe from Boona Mount north of Condolobin (CANB 816114.1) was until 2024 the only record for Monotaxis macrophylla on the South Western Plains. The 1977 collection was made near a trig station on a sandstone hill with skeletal sandy soils supporting Eucalyptus dwyeri and Acacia doratoxylon, and the recent observational record (SDFKI0228891) occurs in similar habitat. Acceptance of this adds a further 9 km2 of PCT184 (Dwyer’s Red Gum – White Cypress Pine – Currawang Low Shrub-Grass Woodland) as potential habitat for this species. These historical collections, made over 120 and nearly 50 years ago respectively, raise the possibility that additional undetected populations of Monotaxis macrophylla may still occur on the Western Plains of NSW.
Equally important as PCTs known to support Monotaxis macrophylla are those PCTs where the species has not yet been reported. Taking the heathland formation as an example, where 11 of the 15 identified PCTs reside, there are an additional 47 heath PCTs within the Northern Montane Heaths, 18 in Sydney Montane Heaths, and 20 in Southern Montane Heaths. Additionally, as noted above, the species has also been observed on numerous occasions in Banksia aemula heath in south-eastern Qld, a broad vegetation type common along the NSW coast but currently with no known populations of Monotaxis macrophylla. All of these habitats should be prioritised for targeted survey of this species following fire events.
Sousa-Baena et al. (2014) highlighted that for many data deficient species, relevant data do actually exist but often remain unanalysed and not used in conservation decision-making. For Monotaxis macrophylla, the data analysed in this paper are held within existing publicly accessible databases, yet it is unclear whether they have been fully utilised in the preparation of regulatory guidelines. Admittedly, this may have occurred because of limited resources, and a desire to be conservative when identifying PCTs considered likely to support the species. However, it does present problems for the regulatory process of development and conservation, when reliance shifts from regulators to proponents to provide evidence of absence rather than the former providing evidence of presence.
Any model of species distribution, including those incorporating indicator surrogates, can be improved as additional data become available. As improvements are adopted into regulatory processes, a more holistic system of conservation management can then be achieved. Using publicly accessible presence data, the effectiveness of utilising indicator surrogates (NSW PCTs) to predict where Monotaxis macrophylla might occur was found wanting for this species. It may now be appropriate for government to build on this review and revise the list of predicted habitats, as encapsulated in PCTs, not only for Monotaxis macrophylla but for other cryptic or ephemeral plant species occupying wide distributions with limited records. Furthermore, given its importance in regulation, a process of regular systematic review of allocated PCTs (perhaps every 2 years) is warranted for all threatened species to ensure the most up-to-date data inform predicted PCTs within the BAM. At the very least, review should include an audit of records with assessment of PCT, utilising existing public databases and herbarium collections, and publication of any changes made. Expert panels comprising ecologists familiar with target species might also be convened to guide changes to PCT allocations, rather than this being permissible by accountable species officers within government as is the current situation.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Declaration of funding
This work was completed as part of several consultancy projects funded through various sources, supported in-kind by the author.
Acknowledgements
Thanks are extended to Phil Lamrock, Ryan Sims and Jamie Gleeson for discussions on the issues raised and review of an earlier draft of this paper. Two anonymous reviewers are also thanked for constructive feedback.
References
Abdo L, Griffin S, Kemp A, Coupland G (2021) Disparity in biodiversity offset regulation across Australia may reduce effectiveness. Australasian Journal of Environmental Management 28, 81-103.
| Crossref | Google Scholar |
Ayers D, Nash S, Baggett K (Eds) (1996) ‘Threatened Species of Western New South Wales.’ (NSW National Parks and Wildlife Service: Hurstville, NSW, Australia) Available at https://nswdpe.intersearch.com.au/nswdpejspui/handle/1/8497
Bachman SP, Brown MJM, Leão TCC, Lughadha EN, Walker BE (2024) Extinction risk predictions for the world’s flowering plants to support their conservation. New Phytologist 242, 797-808.
| Crossref | Google Scholar | PubMed |
Baker DJ, Maclean IMD, Goodall M, Gaston KJ (2021) Species distribution modelling is needed to support ecological impact assessments. Journal of Applied Ecology 58, 21-26.
| Crossref | Google Scholar |
Bell SAJ (2019) Additions and amendments to the rare or threatened vascular plants of Wollemi National Park, central eastern New South Wales. Cunninghamia 19, 43-56 Available at https://www.botanicgardens.org.au/sites/default/files/2023-06/Cunninghamia-10-3-331-Bell-331.pdf.
| Google Scholar |
Bell S, Holzinger B (2015) Wildfire reveals new populations of the endangered Commersonia rosea and Monotaxis macrophylla in northern Wollemi National Park, NSW. Australasian Plant Conservation 23, 2-4.
| Crossref | Google Scholar |
Bell SAJ, Lamrock P, Haines HA, Turney C (2022) Multiple lines of evidence infer centurial-scale habitat change and resilience in a threatened plant species at Mount Dangar, Hunter Valley, New South Wales. Australian Journal of Botany 70, 432-446.
| Crossref | Google Scholar |
Briggs S (2011) Avoiding madness in the method: best practice methodologies for regulatory biodiversity assessment. Pacific Conservation Biology 17, 94-96.
| Crossref | Google Scholar |
Cramer MD, Verboom GA (2017) Measures of biologically relevant environmental heterogeneity improve prediction of regional plant species richness. Journal of Biogeography 44, 579-591.
| Crossref | Google Scholar |
Crisfield VE, Dennett JM, Denny CK, Mao L, Nielsen SE (2020) Species richness is a surrogate for rare plant occurrence, but not conservation value, in boreal plant communities. Biodiversity and Conservation 29, 99-114.
| Crossref | Google Scholar |
DAWE (2020) National Vegetation Information System (NVIS) data products V5.0. Department of Agriculture, Water and the Environment. Available at https://www.environment.gov.au/land/native-vegetation/national-vegetation-information-system/data-products
DCCEEW (2023) NSW State Vegetation Type Map Release C2.0M2.0 (December 2023). Department of Climate Change, Energy, the Environment and Water. Available at https://datasets.seed.nsw.gov.au/dataset/nsw-state-vegetation-type-map
DPE (2022) A revised classification of plant communities of Eastern New South Wales. Department of Planning and Environment. Available at https://www.environment.nsw.gov.au/-/media/OEH/Corporate-Site/Documents/BioNet/a-revised-classification-of-plant-communities-of-eastern-new-south-wales-220178-2.pdf
DPE (2023) Biodiversity Assessment Method 2020 Operational Manual – Stage 2, April 2023. (NSW and Department of Planning and Environment). Available at https://www.environment.nsw.gov.au/-/media/OEH/Corporate-Site/Documents/Animals-and-plants/Biodiversity/biodiversity-assessment-method-operational-manual-stage-2-230164.pdf
DPIE (2020) Surveying threatened plants and their habitats: NSW survey guide for the Biodiversity Assessment Method, April 2020. (NSW Department of Planning, Industry and Environment) Available at https://www.environment.nsw.gov.au/-/media/OEH/Corporate-Site/Documents/Animals-and-plants/Biodiversity/surveying-threatened-plants-and-habitats-nsw-survey-guide-biodiversity-assessment-method-200146.pdf
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JMM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129-151.
| Crossref | Google Scholar |
Environment Australia (2000) Revision of the Interim Biogeographical Regionalisation of Australia (IBRA) and the development of Version 5.1 – Summary Report. Department of Environment and Heritage, Canberra, ACT, Australia. Available at https://www.dcceew.gov.au/sites/default/files/documents/revision-ibra-development-5-1-summary-report.pdf
Faith DP, Walker PA (1996) Environmental diversity: on the best-possible use of surrogate data for assessing the relative biodiversity of sets of areas. Biodiversity and Conservation 5, 399-415.
| Crossref | Google Scholar |
Gibson C (2002) Along the Prostanthera Trail. Lasianthos 3, 12-17.
| Google Scholar |
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecology Letters 8, 993-1009.
| Crossref | Google Scholar | PubMed |
Halford DA, Henderson RJF (2002) Studies in Euphorbiaceae A.L. Juss. sens lat. 4. A revision of Monotaxis Brongn. (Acalyphoideae Ascherson, Ampereae Mull.Arg.). Austrobaileya 6, 273-292 Available at https://archive.org/details/biostor-257094.
| Google Scholar |
Hunter JT (1998) Notes on the occurrence of Monotaxis macrophylla Benth. (Euphorbiaceae), with particular reference to New South Wales. The Queensland Naturalist 36, 21-24.
| Google Scholar |
Hunter JT (2005) Vegetation and floristics of Warra National Park and Wattleridge, Northern Tablelands, NSW. Cunninghamia 9, 255-274 Available at https://www.google.com/url?sa=t&source=web&rct=j&opi=89978449&url=https://www.researchgate.net/publication/228658066_Vegetation_and_floristics_of_Warra_National_Park_and_Wattleridge_Northern_Tablelands_NSW&ved=2ahUKEwiD7qjDvYWIAxXHp1YBHZnlAy0QFnoECBEQAQ&usg=AOvVaw2pfU2x-_7zn_p1FwEHolFR.
| Google Scholar |
Hunter JT (2011) Vegetation and Floristics of the Pilliga East State Conservation Area, Willala Aboriginal Area, Pilliga Nature Reserve and the Ukerbarley Addition. Available at https://www.researchgate.net/profile/John-Hunter-7/publication/279963204_Vegetation_and_flora_of_the_Pilliga_Nature_Reserve/links/55a0661008aef92d04ce371b/Vegetation-and-flora-of-the-Pilliga-Nature-Reserve.pdf
Hunter JT, Bruhl JJ (1997) Significant range extensions for 10 species of vascular plants in northern New South Wales. Austrobaileya 4, 691-694.
| Crossref | Google Scholar |
Hunter JT, Copeland LM (2007) Field Survey of Western Granite Threatened Flora Species, Identified as Priority by the Namoi Catchment Management Authority, January 2007. Report to the Namoi Catchment Management Authority. Available at https://www.google.com/url?sa=t&source=web&rct=j&opi=89978449&url=https://www.researchgate.net/publication/281366338_Field_Survey_of_Western_Granite_Threatened_Flora_Species_Identified_as_Priority_by_the_Namoi_Catchment_Management_Authority&ved=2ahUKEwj0kJDQuoWIAxWL7DQHHevcEPoQFnoECAYQAQ&usg=AOvVaw0YKQwu53s8eUbFKXG7v3jW
Hunter M, Westgate M, Barton P, Calhoun A, Pierson J, Tulloch A, Beger M, Branquinho C, Caro T, Gross J, Heino J, Lane P, Longo C, Martin K, McDowell WH, Mellin C, Salo H, Lindenmayer D (2016) Two roles for ecological surrogacy: indicator surrogates and management surrogates. Ecological Indicators 63, 121-125.
| Crossref | Google Scholar |
Lannuzel G, Balmot J, Dubos N, Thibault M, Fogliani B (2021) High-resolution topographic variables accurately predict the distribution of rare plant species for conservation area selection in a narrow-endemism hotspot in New Caledonia. Biodiversity and Conservation 30, 963-990.
| Crossref | Google Scholar |
Lombard AT, Cowling RM, Pressey RL, Rebelo AG (2003) Effectiveness of land classes as surrogates for species in conservation planning for the Cape Floristic Region. Biological Conservation 112, 45-62.
| Crossref | Google Scholar |
Mainali K, Hefley T, Ries L, Fagan WF (2020) Matching expert range maps with species distribution model predictions. Conservation Biology 34, 1292-1304.
| Crossref | Google Scholar | PubMed |
Metcalfe L, Sivertsen DP, Tindall D, Ryan KM (2003) Natural vegetation of the New South Wales wheat-belt (Cobar–Nyngan–Gilgandra, Nymagee–Narromine–Dubbo 1:250 000 vegetation sheets). Cunninghamia 8, 253-284 Available at https://www.google.com/url?sa=t&source=web&rct=j&opi=89978449&url=https://www.botanicgardens.org.au/sites/default/files/2023-06/Volume-8%25282%2529-2003-Cun8Met253-284.pdf&ved=2ahUKEwiZpPaovoWIAxVfslYBHcc_E8IQFnoECB8QAQ&usg=AOvVaw2DOo3mo8rkBQ5-Tp5oNTTp.
| Google Scholar |
Mod HK, Scherrer D, Luoto M, Guisan A (2016) What we use is not what we know: environmental predictors in plant distribution models. Journal of Vegetation Science 27, 1308-1322.
| Crossref | Google Scholar |
Muñoz J, Felicísimo AM (2004) Comparison of statistical methods commonly used in predictive modelling. Journal of Vegetation Science 15, 285-292.
| Crossref | Google Scholar |
Murray BR, Thrall PH, Lepschi BJ (2002) Relating species rarity to life history in plants of eastern Australia. Evolutionary Ecology Research 4, 937-950 Available at http://www.evolutionary-ecology.com/issues/v04n07/ccar1407.pdf.
| Google Scholar |
Muscatello A, Elith J, Kujala H (2021) How decisions about fitting species distribution models affect conservation outcomes. Conservation Biology 35, 1309-1320.
| Crossref | Google Scholar | PubMed |
Olivares-Rojas J, Cook CN, Ward MS, Walsh JC (2024) Species and ecological communities as management surrogates for threatened biodiversity. Biodiversity and Conservation 33, 987-1008.
| Crossref | Google Scholar |
Pharo EJ, Beattie AJ (2001) Management forest types as a surrogate for vascular plant, bryophyte and lichen diversity. Australian Journal of Botany 49, 23-30.
| Crossref | Google Scholar |
PlantNET (2024) The NSW Plant Information Network System. Royal Botanic Gardens and Domain Trust, Sydney. Available at https://plantnet.rbgsyd.nsw.gov.au [accessed 21 August 2024]
Ponce-Fontenla S, Serrano M, Carballal R, Regos A (2021) Sentinel 2 images enable reliable prediction of fine-scale habitat dynamics of narrow endemic plant species in serpentine soils. Applied Vegetation Science 24, e12614.
| Crossref | Google Scholar |
Queensland Herbarium (2024) Regional Ecosystem Description Database (REDD). Version 13.1 (May 2024). (Queensland Department of Environment, Science and Innovation: Brisbane, Qld, Australia) Available at https://www.qld.gov.au/__data/assets/file/0010/411112/REDD_v13.1_2024.zip
Robinson NM, Scheele BC, Legge S, Southwell DM, Carter O, Lintermans M, Radford JQ, Skroblin A, Dickman CR, Koleck J, Wayne AF, Kanowski J, Gillespie GR, Lindenmayer DB (2018) How to ensure threatened species monitoring leads to threatened species conservation. Ecological Management and Restoration 19, 222-229.
| Crossref | Google Scholar |
Roff A, Day M, Thonell J, Denholm B (2022) NSW State Vegetation Type Map: Technical Notes. NSW Department of Planning and Environment, Sydney, NSW, Australia. Available at https://www.google.com/url?sa=t&source=web&rct=j&opi=89978449&url=https://www.environment.nsw.gov.au/-/media/OEH/Corporate-Site/Documents/BioNet/nsw-state-vegetation-type-map-technical-notes-220275.pdf&ved=2ahUKEwichNGLu4WIAxUCqVYBHTKBL4AQFnoECAwQAQ&usg=AOvVaw3xn1gYsUBCT22j6wjEQUQX
Saunders ME, Andrew RL, Mitchell-Williams J, Pemberton P, Wandrag EM, Hunter JT (2024) Rapid on-ground assessment after the 2019–2020 megafires reveals new information on rare and threatened plants in northern New South Wales, Australia. Austral Ecology 49, e13514.
| Crossref | Google Scholar |
Scheele BC, Legge S, Armstrong DP, Copley P, Robinson N, Southwell D, Westgate MJ, Lindenmayer DB (2018) How to improve threatened species management: an Australian perspective. Journal of Environmental Management 223, 668-675.
| Crossref | Google Scholar | PubMed |
Silcock JL, Fensham RJ (2018) Using evidence of decline and extinction risk to identify priority regions, habitats and threats for plant conservation in Australia. Australian Journal of Botany 66, 541-555.
| Crossref | Google Scholar |
Sousa-Baena MS, Garcia LC, Peterson AT (2014) Knowledge behind conservation status decisions: data basis for ‘Data Deficient’ Brazilian plant species. Biological Conservation 173, 80-89.
| Crossref | Google Scholar |
Stockwell DRB, Peterson AT (2002) Effects of sample size on accuracy of species distribution models. Ecological Modelling 148, 1-13.
| Crossref | Google Scholar |
Tälle M, Ranius T, Öckinger E (2023) The usefulness of surrogates in biodiversity conservation: a synthesis. Biological Conservation 288, 110384.
| Crossref | Google Scholar |
Thuiller W, Araújo MB, Lavorel S (2003) Generalized models vs. classification tree analysis: predicting spatial distributions of plant species at different scales. Journal of Vegetation Science 14, 669-680.
| Crossref | Google Scholar |
Wall WA, Just MG, Huskins SD, Hohmann MG (2024) Enhancing rare plant population predictions through demographic modeling of seed predation, dispersal, and habitat suitability. Plant Ecology 225, 63-74.
| Crossref | Google Scholar |
Wang HH, Wonkka CL, Treglia ML, Grant WE, Smeins FE, Rogers WE (2019) Incorporating local-scale variables into distribution models enhances predictability for rare plant species with biological dependencies. Biodiversity and Conservation 28, 171-182.
| Crossref | Google Scholar |
Ware C, Williams KJ, Harding J, Hawkins B, Harwood T, Manion G, Perkins GC, Ferrier S (2018) Improving biodiversity surrogates for conservation assessment: a test of methods and the value of targeted biological surveys. Diversity and Distributions 24, 1333-1346.
| Crossref | Google Scholar |
Wilson KA, Westphal MI, Possingham HP, Elith J (2005) Sensitivity of conservation planning to different approaches to using predicted species distribution data. Biological Conservation 122, 99-112.
| Crossref | Google Scholar |
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A, NCEAS Predicting Species Distributions Working Group (2008) Effects of sample size on the performance of species distribution models. Diversity and Distributions 14, 763-773.
| Crossref | Google Scholar |
Woinarski JCZ, Price O, Faith DP (1996) Application of a taxon priority system for conservation planning by selecting areas which are most distinct from environments already reserved. Biological Conservation 76, 147-159.
| Crossref | Google Scholar |