

Limited evidence for the use of livestock for the conservation management of exotic plant cover
David J. Eldridge A F , Ian Oliver B C , James Val D and Samantha K. Travers E
A F , Ian Oliver B C , James Val D and Samantha K. Travers E
A New South Wales Department of Planning, Industry and Environment, c/- Centre for Ecosystem Science, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
B New South Wales Department of Planning, Industry and Environment, PO Box 1477, Gosford, NSW 2250, Australia.
C School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia.
D New South Wales Department of Planning, Industry and Environment, PO Box 363, Buronga, NSW 2739, Australia.
E Centre for Ecosystem Science, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
F Corresponding author. Email: d.eldridge@unsw.edu.au
Australian Journal of Botany 68(2) 137-145 https://doi.org/10.1071/BT19183
Submitted: 25 November 2019 Accepted: 12 April 2020 Published: 6 May 2020
Journal Compilation © CSIRO 2020 Open Access CC BY-NC-ND
Abstract
Livestock grazing has been used for conservation management in some situations to reduce exotic plant cover, but supporting empirical evidence is scant. This is particularly true for eastern Australian semiarid woodlands and forests. We surveyed 451 sites across three broad semiarid vegetation communities (cypress pine, black box, red gum) in eastern Australia to examine the effects of recent and longer-term (historic) grazing by livestock on exotic (and native) plant cover. Because our focus was the use of domestic livestock for conservation management, our sites were mostly located in conservation reserves, travelling stock routes and native production forests where domestic grazing is approved via permit. We also included recent grazing by kangaroos and rabbits, which occur widely with livestock across the study sites. Structural equation modelling revealed that recent sheep and cattle grazing was associated with increased exotic plant cover in two of the three communities (cypress pine, black box), and historic grazing was associated with lower exotic (and native) plant cover across all communities. Recent cattle grazing-induced reductions in exotic plant cover in the red gum community resulted from declines in the palatable exotic annual grass Ehrharta longiflora. Higher intensity grazing by rabbits, sheep and cattle in the cypress pine community increased exotic plant cover by suppressing the negative effect of native cover on exotic cover. Higher intensity rabbit grazing reinforced the negative effect of nitrogen on native plant cover, resulting in increases in exotic cover. Overall, we found strong evidence that domestic livestock grazing reduces native plant cover either directly or indirectly in all three communities, but limited evidence that it would be a useful conservation management strategy to reduce exotic plant cover in these communities.
Additional keywords: cattle, forb, grass, herbivory, rabbit, sheep, weeds.
Introduction
Livestock grazing of native plant communities has been used for millennia to support human populations and cultures worldwide (Pineiro et al. 2006); however, overgrazing can have deleterious effects on native plant communities and soils. It can reduce the richness of native plants (Eldridge Eldridge et al. 2018) and habitat for plants and animals (Socher et al. 2013). It can reduce soil condition and the capacity of soil to resist erosion (Vandandorj et al. 2017). Indirectly, overgrazing can reduce soil porosity, alter the decomposition and mineralisation of organic matter (Golluscio et al. 2009) thereby altering carbon, nitrogen and phosphorus cycles, which in turn allows the proliferation of exotic plant species (Eldridge et al. 2017). Similarly, the large body of literature from Australia indicates that grazing by European livestock leads to reductions in native plant cover and increases in exotic plant cover (Page and Beeton 2000; Yates et al. 2000; Spooner and Briggs 2008).
Notwithstanding the well-known negative effects of overgrazing, the past few decades have seen a renewed interest in the use of domestic livestock grazing for rehabilitating degraded native ecosystems by completely removing or reducing the cover of exotic plants (Olson and Lacey 1994; Popay and Field 1996). For example, high intensity sheep grazing has been used to control the thistle (Cirsium arvense) by increasing competition in the existing native plant community (Pywell et al. 2010), and continuous sheep grazing has been shown to control the aggressive Eurasian forb leafy spurge (Euphorbia esula; Olson and Wallander 1998). Goats are widely used to reduce the cover of woody plants such as blackberry (Rubus fruticosus; Mellado 2016) and grazing by cattle and horses has been used to prevent the encroachment of grasses in coastal dunes (Kooijman and van der Meulen 1996). Although there may be some situations where livestock grazing could be used for conservation management in Australia (Lunt et al. 2007), supporting empirical evidence is often scant, especially for semiarid woodlands and forests.
Although there are abundant literature on the effects of livestock grazing on plant community composition, ecosystem health and biodiversity from grazed Australian landscapes, few studies have compared the impacts of different herbivore types (but see Eldridge et al. 2018; Travers et al. 2019). This is critically important to the use of livestock for conservation management because free-ranging kangaroos (Macropus spp.), and residual populations of European rabbits (Oryctolagus cuniculus) co-occur with livestock over much of the rangelands of Australia and have been shown to have marked effects on plant community composition (Tiver and Andrew 1997; Prowse et al. 2019). Further, we know of no studies that have examined variable grazing history (recent cf. historic) across a range of environmental contexts in Australian semiarid woodlands and forests. In the present study we aim to test these effects on exotic and native plant cover using a large number of sites across an extensive area of rangelands in eastern Australian semiarid woodlands and forests.
We use a regional study of 451 sites in eastern Australia, spanning three broad vegetation communities, to test the effects of recent grazing, at different intensities by different herbivores, and historic grazing by livestock, on both native and exotic plant cover. As our focus is the use of domestic livestock for conservation management, our sites were largely located in conservation reserves, travelling stock routes and multiple use production forests where domestic grazing is approved via permit. Because we expected effects on cover to vary among different herbivores, we used sites that ranged from negligible grazing by any herbivore, to sites with variable intensities of livestock (sheep, cattle) grazing, with or without rabbits and kangaroos. Based on available evidence we expected that grazing would influence plant cover directly, by herbivory, and indirectly, by reducing soil health (Lunt et al. 2007). We expected increases in exotic plant cover with increasing grazing by sheep and cattle (Prober and Lunt 2009; Yates et al. 2000; Spooner and Briggs 2008), and by rabbits (Leigh et al. 1989), but fewer effects of kangaroo grazing (Travers et al. 2018). Thus we did not expect to find evidence to support the use of domestic stock for conservation management in these woodlands and forests. Climate (aridity) and soil characteristics (texture) would also be expected to affect native and exotic plant cover due to differences in precipitation and water holding capacity and therefore productivity (Lezama et al. 2013; Borer et al. 2014; Eldridge et al. 2018). Climate is also likely to influence the distribution of different herbivores, as more arid locations would tend to have a greater intensity of sheep grazing, but less kangaroo and cattle grazing.
We used structural equation modelling to examine the direct and indirect effects of grazing on native and exotic plant cover, after accounting for the effects of other variables such as climate, woody plant cover, soil chemistry and soil health (Fig. 1; Table 1). An extensive literature of grazing effects from Australian grazing systems (Eldridge et al. 2016), allowed us to develop an a priori model that described the expected relationships among our variables of interest. Using this model, we predicted that increasing intensity of livestock grazing would directly reduce the cover of native plants (Fig. 1; Pathway 1a) but increase the cover of exotic plants (Pathway 1b). We also expected that grazing would increase the cover of exotic plants, indirectly, by enhancing the negative effect of native plants on exotics (Pathway 2 via 1a; Fig. 1).
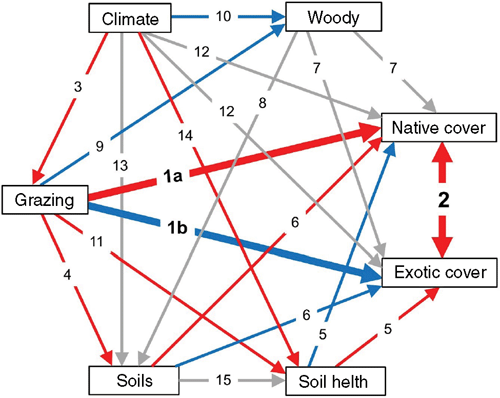
|

|
Methods
Study area
We conducted our study at 451 sites across 0.5 M km2 of dryland in eastern Australia. Sites were located in three markedly different semiarid woodland communities used that are used for livestock grazing, including conservation reserves (52%, native forests (23%), travelling stock reserves (20%) and presented a range of grazing histories and intensities. Climate across the study area is mainly Mediterranean (aridity 0.26–0.39), with slightly more rainfall during the six warmer months in the east-central and during the cooler months in the south and south-west. Average rainfall (385 to 460 mm year–1) and average temperatures (~18°C) varied little across the gradient.
The three communities were dominated by either black box (Eucalyptus largiflorens F.Muell.; hereafter black box), white cypress pine (Callitris glaucophylla F.Muell.; hereafter cypress pine) or river red gum (Eucalyptus camaldulensis Dehnh.; hereafter red gum). A description of the soil physical and chemical properties, grazing intensity and climatic information for the three communities as shown in Supplementary Material Appendix S1 (available at the journal’s website). Soil textures ranged from sandy clay-loams in the cypress pine community to silty loams in black box and red Gum communities (Eldridge et al. 2016).
Our study used data from 151 sites in cypress pine and 150 in each of black box and red gum (n = 451). The original selection of these sites from a potentially larger pool of sites across eastern Australia was based on several factors determined using Arc GIS and pre-measurement verification. These were (1) distance from permanent water, which has been used widely as a surrogate of grazing intensity (Fensham et al. 2009), (2) a range of grazing histories (e.g. ungrazed and lightly grazed conservation reserves, road verges subjected to intermittent grazing, and woodlands with long histories of more intense grazing), which enabled us to sample across a full spectrum of recent (last 2–3 years) and historic (up to 20 years) grazing intensities, and (3) >50 m from any vehicle track or road. Selecting a range of different distances from water enabled us to preselect sites that we expected would represent different levels of grazing intensity. Distance from water, however, was not used as our measure of grazing intensity. Rather, grazing intensity was based on dung counts and the intensity of livestock tracks (see below). Our models examined relationship among (1) climatic (aridity, rainfall), woody plant cover, grazing intensity (recent and historic), soil physical and chemical properties, and soil heath (see below), and the cover of exotic and native plants. The methods used to assess these attributes are described below.
Climatic variables
We had two climatic variables in our models; rainfall received in the nine months before sampling (lag rainfall), and aridity (FAO 2015). Lag rainfall was used because field work extended over a period of ~7 months from late autumn to early summer, hence the need to account for potential differences in rainfall that might affect our assessment of plant cover. The FAO (2015) Aridity Index (AI) was used because it is a useful method to account for the variability in climate among study sites that span a distance of >400 km from west to east (Delgado-Baquerizo et al. 2013). The AI expresses precipitation in relation to potential evapotranspiration using FAO’s Global Aridity Map (FAO 2015). For our purpose, we calculated Aridity as 1-AI to denote that increases in Aridity represent increasing dryness. Aridity was negatively related to annual mean precipitation in our database (Spearman’s ρ = –0.95, P < 0.001, n = 451).
Woody cover and exotic and native plant cover
At each site we established a 200 m long transect running perpendicular to the nearest livestock watering point at a predetermined distance based on the desktop Arc GIS assessment. Watering points were generally earthen dams or troughs. Along this transect we assessed the cover of woody plants (trees and shrubs >3 m tall) at 100 points every 2 m. We also placed five large (25 m2) quadrats every 50 m (hereafter ‘large quadrat’) along the transect, and nested a smaller (0.25 m2) quadrat (hereafter ‘small quadrat’) within the large quadrat. Within the large quadrat we identified all plant species, enabling us to record the total cover of exotic and native plant species.
Grazing intensity
At each site, we calculated grazing intensity based on two measures; recent and historic grazing. Recent grazing (during the past 2–3 years) was assessed by counting the number of pellets or pats of different herbivores within the large (cattle, sheep or goat, kangaroo) and small (kangaroo, rabbit, sheep or goat) quadrats. Dung and pellet counts are used widely to estimate herbivore abundance (Johnson and Jarman 1987). Counts were converted to mass using algorithms relating counts of dung to mass for different herbivores (Eldridge et al. 2017). Using counts and herbivore specific algorithms, allowed us to calculate the total oven-dried mass of dung per hectare for each herbivore type as our measure of recent grazing intensity. For sheep or goats and kangaroos, dung mass was based on average values using data from both the large and small quadrats. We were unable to discriminate between sheep and goat, or European rabbit and European hare (Lepus europaeus) dung. Historic livestock grazing was assessed by measured the width and depth of all livestock tracks crossing the 200 m transect to derive a total cross-sectional area of livestock tracks for each site (Pringle and Landsberg 2004).
Soil properties and soil health
A sample of the uppermost layer of the soil (5 cm) was collected from the centre of the small quadrat, air-dried, ground, and passed through a 2 mm sieve to remove any roots or organic debris. This was used to assess soil total C and N in the uppermost layers using high intensity combustion (LECO CNS-2000; LECO Corporation, St Joseph, MI, USA), available (Olsen) P (Colwell 1963), and particle size distribution (sand, silt and clay contents), using the hydrometer method (Bouyoucos 1962).
To assess soil health, we used a rigorous field-based protocol to assess the status and morphology of the soil surface within the small quadrats (Tongway 1995). Within these quadrats, we measured 10 attributes: surface roughness, crust resistance, crust brokenness, crust stability, surface integrity (cover of uneroded surface), cover of deposited material, biocrust cover, litter cover, litter origin, and the degree of litter incorporation (Appendix S2). The values of these attributes were used to calculate a combined measure of soil health that has been shown to be highly correlated with ecosystem functions associated with soil stability, nutrient cycling and infiltration (Maestre and Puche 2009; Eldridge and Delgado-Baquerizo 2018; see Appendix S2 for specific analytical methods).
Statistical analyses
We used structural equation modelling (SEM; Grace 2006) to clarify the direct and indirect effect of grazing, climate, and plant and soil variables on the cover of exotic and native plants. Structural equation modelling is a synthesis of path analysis, factor analysis, and maximum likelihood techniques (Appendix S3), and is now widely used in ecology to examine complex networks of relationships found in natural systems (Grace 2006). Structural equation modelling tests the plausibility of a causal model, based on a priori information, in explaining the relationships among a group of variables of interest. Its strength, over other contemporary methods such as generalised linear mixed models (GLMM), is its ability to partition direct and indirect effects of one variable upon another and to estimate the strengths of these multiple effects.
Our a priori model (Appendix S3) was compared with the variance-covariance matrix to assess an overall goodness-of-fit, using the χ2 statistic. The goodness of fit test estimates the long-term probability of the observed data given the a priori model structure. Thus, high probability values indicate that these models are highly plausible causal structures underlying the observed correlations. The stability of these models was evaluated by introducing other indicators to determine whether they represented additional contributing information (Reisner et al. 2013). Analyses were performed using the AMOS 22 (IBM, Chicago, IL, USA) software. The model with the strongest measures of fit (e.g. low χ2, high goodness of fit index [GFI] and high normal fit index [NFI]) were interpreted as showing the best fit to our data (Appendix S3). The strength and sign of relationships among the variables is represented by path coefficients, which are analogous to partial correlation coefficients (Grace 2006).
Finally, because our SEM analysis indicated that some exotic plant species declined with increasing livestock grazing, we used generalised linear mixed modelling (GLMM) to determine the identity of these species. We used herbivores (cattle, sheep, kangaroo, rabbit) and our measure of historic livestock grazing (tracks) as fixed effects and log (x + 1) transformed and standardised the data before modelling with the lme4 package (Bates et al. 2015; Appendix S4).
Results
Exotic plant cover was generally low (<20%) across all communities, and significantly lower in black box (7.1 ± 0.9%; mean ± s.e.) than either cypress pine (13.3 ± 1.2%) or red gum (17.2 ± 1.7%; Supplementary Material Table S1, available at the journal’s website). Exotic plants were dominated by grasses, and to a lesser extent, annual forbs, irrespective of whether we measured cover or abundance (Fig. 2). The standardised total effects (STE) derived from the structural equation modelling represent the overall effects of our variables of interest on exotic plant cover, taking into account both positive and negative, and direct and indirect, effects (Fig. 3a). The standardised total effects indicate three general trends. First, increased soil nitrogen was strongly associated with increased cover of exotic plants whereas increased soil carbon and woody cover were associated with reductions in exotic cover. Second, recent sheep and cattle grazing were generally associated with increased exotic cover in all communities (with the exception of cattle in red gum). Third, historic grazing was associated with lower exotic cover across all communities (Fig. 3a).
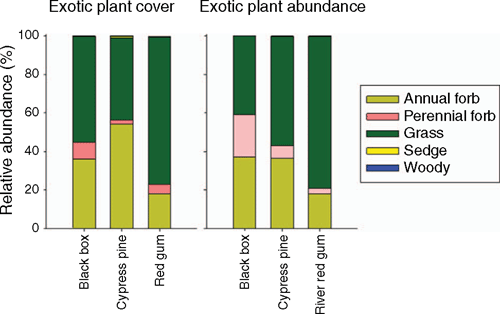
|

|
The SEMs revealed further details of the direct and indirect effects and the directions of these effects (Fig. 3). For example, in the red gum community, increasing intensity of recent cattle grazing and historic grazing was associated with direct overall reductions in exotic plant cover (Fig. 3d). However, in this community, indirect effects included increased exotic cover associated with reductions in native cover brought about by increased soil phosphorus due to recent sheep and cattle grazing (Fig. 3d). The strong correlation between exotic plant cover and abundance meant that our conclusions were similar when abundance was used in the models in place of cover.
In the cypress pine community there were no direct effects of domestic grazing on exotic cover but there were some significant indirect effects that increased exotic cover (Fig. 3c). For example, increased grazing by sheep and cattle (and rabbits) directly reduced native cover resulting in increased exotic plant cover. Similarly, increased grazing by sheep and cattle (and rabbits) reduced soil health, thereby reducing native plant cover, further increasing exotic cover. In addition to the effects of domestic livestock on exotic plant cover, path coefficients clearly revealed very strong indirect effects of rabbit grazing on both exotic and native cover in the cypress pine community (Fig. 3c).
In the black box community there were no direct effects of domestic grazing on exotic cover but here also, recent cattle and historic grazing reduced native cover (Fig. 3b). Our models also revealed some effects unrelated to grazing. For example, in the black box community, increased aridity suppressed the positive effect of carbon on native cover and suppressed the negative effect of increasing woody cover on exotic cover (Fig. 3b). In cypress pine, increasing aridity suppressed the positive effect of both soil nitrogen and carbon on soil health and its positive effect on native cover (Fig. 3c). For red gum, increasing aridity was associated with reduced native plant cover and reduced soil health; increased woody cover suppressed exotic cover, but enhanced native cover, and increasing soil nitrogen increased exotic cover (Fig. 3d).
Across all vegetation communities, generalised linear mixed models (GLMMS) revealed that historic grazing reduced the presence of both native and exotic species (Supplementary Material Figure S4.1, available at the journal’s website). They also revealed that the presence of the exotic grass Ehrharta longiflora (Sm.) was negatively correlated with increases in recent cattle grazing whereas the presence of the exotic annual grasses Lolium rigidum (Gaudin) and Bromus diandrus (Roth) were both positively correlated (Fig. S4.2). In contrast the GLMMs revealed that both Bromus diandrus (Roth) and Lolium rigidum (Gaudin) were highly negatively correlated with increasing historic grazing (Fig. S4.2).
Discussion
In our study we used a very large regional dataset to test the effects of livestock grazing on exotic (and native) plant cover within three common vegetation types across an extensive area of semiarid eastern Australia. Our results generally revealed that recent grazing increased exotic cover but historic grazing reduced it (Fig. 3a). We found some evidence of lower cover of exotic plants with higher levels of recent cattle grazing in the red gum community, but this was due to reductions in only a single, relatively palatable species of exotic grass Ehrharta longiflora (Fig. S4.2). However, recent and historic grazing also reduced native plant cover, either directly or indirectly, in all three vegetation communities.
Grazing effects on exotic plant cover vary with herbivore type and plant community
The effects of herbivores on exotic plant cover varied across the study area, but for two of the three communities, grazing either increased exotic cover directly, or reduced the cover of natives, sometimes by increasing soil P or reducing soil health, thereby indirectly increasing exotic cover. There were no effects in the black box community (Fig. 3b), but in cypress pine, grazing by cattle, sheep and rabbits was associated with increased exotic plant cover by reducing natives (Fig. 3c). In the red gum community, increasing cattle and sheep grazing was associated with increased exotic plant cover via a suppressive effect of increased phosphorus (P) on native plants (Fig. 3d). Native and exotic plant cover were negatively related in cypress pine (r = –0.46) and red gum (r = –0.16). Thus, although increased grazing may not have affected exotics directly, its effect was to create conditions that are conducive to the increase in exotic plants such as reducing the quality of the soil surface or reducing competition from native plants (Eldridge et al. 2017).
Grazing could also increase exotic cover by increasing concentrations of soil nutrients such as nitrogen (N) or phosphorus (P), which favour nitrophilous weedy species such as annual grasses (Aerts and Berendse 1988; Bobbink 1991; Driscoll 2019). We found empirical evidence that increased P from grazing by sheep and cattle (red gum) and increased P and N from rabbits (cypress pine), indirectly resulted in greater exotic plant cover (Fig. 3c, d). Increased soil P from grazing-induced disturbance and exposure of subsoils is a major source of ecosystem P (Delgado-Baquerizo et al. 2013) and can enhance exotic plant cover by suppressing native plants (Dorrough and Scroggie 2008). Although low levels of soil P are important for maintaining native plant richness on low productivity soils (Lambers et al. 2010), high levels, either from herbivore dung or exposed subsoils, can promote exotic, often annual plants at the expense of natives (Prober and Lunt 2009).
Increases in recent cattle grazing were associated with reduced cover of exotic plants in only one of the three communities, red gum (Fig. 3d), a highly productive community associated with large drainage tracts that are infrequently inundated by floodwaters. Soils in this community are relatively stable due to their high content of fine soil particles (silt + clay; Table S1) and the addition of fine material after flooding. Most of the historic grazing in this community can be attributed to cattle (Table S1). Those exotic plant species that did decline under historic and recent grazing in the red gum community were the highly palatable exotic annual grasses Ehrharta longiflora, Bromus diandrus and Lolium rigidum. This result highlights how important it is to consider the palatability of weed species and the specific contexts or scenarios in which grazing may reduce their cover. The Grazing Framework model of Lunt et al. (2007) suggests that grazing could be an appropriate tool for maintaining plant diversity by controlling dominant, grazing-sensitive species, but only in environments where there are no substantial negative impacts on landscape processes, for example, due to non-erodible soils (Lunt 2002). Ehrharta longiflora and its congener Ehrharta calycina (Sm.) are highly palatable exotic species, originally from Africa, that have become widely naturalised in southern Australia but are unable to withstand heavy grazing (Cunningham et al. 2011). Similarly, Lolium rigidum is a highly palatable annual grass that does not tolerate overgrazing under very hot summer conditions (Cunningham et al. 2011), and Bromus diandrus (Roth) is highly palatable in the early growth stages before it forms large awns. We acknowledge that these high biomass plants likely play a role in reducing erosion, nutrient cycling and hydrological processes, and may be important for carrying a fire in some fire-adapted communities. In the red gum community therefore, grazing may produce positive conservation outcomes (sensu Lunt et al. 2007) under some situations by reducing the cover of these palatable exotic plants of high biomass. Thus, grazing might prevent exotic plants from suppressing the richness of other smaller statute plants (Scenario D; Lunt et al. 2007).
Different effects of free-ranging herbivores (rabbits and kangaroos) on exotic cover
Increased rabbit grazing was associated, indirectly, with increases in exotic plant cover by reinforcing the suppressive effect of soil N on native plant cover, which tended to reduce exotic plant cover. Thus, rabbit grazing increased exotic cover indirectly by reducing native plant cover. The standardised total of effects indicated that increased soil N concentrations consistently increased exotic plant cover, consistent with the large body of research showing that increasing soil N increases the likelihood of exotic plant invasion (Groves et al. 2003; Chalmers et al. 2005). We also found a strong overall negative effect of soil carbon on exotic plant cover (Fig. 3a). Microbes can immobilise soil N (making it unavailable to plants) by consuming C stores in the soil (e.g. Chapman et al. 2006). This is particularly relevant for exotic plants, which have a high N demand. The effect of rabbit grazing was most strongly revealed in the cypress pine community where rabbits are largely restricted to sandy soils where they construct underground warrens. Rabbits have two main impacts on exotic plants; (1) increasing their cover by reducing competition from native plants, or (2) predisposing the soil surface to invasion by exotics by increasing surface disturbance. Rabbits are highly selective herbivores (Leigh et al. 1989), and reduce the cover of palatable, generally native, plants in the vicinity of their communal warrens (Leigh et al. 1989). Overgrazing by rabbits has been shown to favour high biomass Mediterranean weeds at the expense of smaller native forbs and grasses (Myers and Poole 1963). Rabbits also create multiple disturbances, both on and off their communal warrens that are suitable microsites for the establishment of weedy plant species (Eldridge and Simpson 2002).
There were no effects of kangaroo grazing on either native or exotic plant cover in any community, despite the fact that kangaroo grazing was recorded at all but two of the 451 sites, often in high densities (Eldridge et al. 2017). High densities of kangaroos have been shown to reduce plant richness (Driscoll 2019; Prowse et al. 2019) in the semiarid woodlands, but grazing by kangaroos in the study area was associated with sites of greater native plant richness in the cypress pine, but not the other communities (Eldridge et al. 2017; Travers et al. 2018). Differences in grazing effects between kangaroos, sheep and cattle likely relates to the fact that kangaroos are highly mobile (Caughley et al. 1987) and unrestrained by fences, unlike European livestock, which in the study area are generally managed by set stocking in large paddocks.
Woody plant and aridity effects on exotic cover
Our standardised total effects indicated that increasing cover of woody plants was associated with reductions in exotic plant cover (Fig. 3a). Woody plants are known to facilitate understorey species (Howard et al. 2012) due to a combination of habitat amelioration (Soliveres et al. 2011) and escape from predation through associational resistance (Smit et al. 2015). That native shrubs and trees facilitate native species has been well established and suggests that removal of shrubs is likely to suppress any positive effects on native understorey species. Our work has also shown that woody plants tend to increase richness of native plants (Eldridge at al. 2017; Travers et al. 2018). We also found that the negative effect of woody plants on exotic cover was suppressed by increasing aridity. In fact, in the red gum community, increased woody cover increased native cover and reduced exotic cover.
Overall our study provides strong evidence that within the context of conservation management of conservation reserves, native forests and travelling stock reserves, recent grazing by sheep, cattle and rabbits either increases exotic cover directly, or indirectly by reducing native plant cover. The only community where recent grazing had a positive impact on exotic cover was river red gum, which is characterised by high biomass exotic palatable grasses. Historic grazing revealed a legacy of general reductions of plant cover regardless of origin (native and exotic). Our study provides strong support for the conclusion that, at least in eastern Australian semiarid woodlands and forests which have a short evolutionary history of livestock grazing, the use of European livestock is unlikely to be a useful conservation management strategy for reducing exotic plant cover.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We thank Mark Peacock, Ross McDonnell and Sarah Carr (NSW National Parks and Wildlife Service), Greg Summerall (Office of Environment and Heritage (OEH), and the River Red Gum Scientific Advisory Committee for project management, and Forest Corporation NSW and landholders for access to field sites. Marta Ruiz-Colmenero, James Glasier and staff from OEH, Umwelt and Ecology Australia collected and entered field data.
References
Aerts R, Berendse F (1988) The effect of increased nutrient avail- ability on vegetation dynamics in wet heathlands. Vegetatio 76, 63–69.Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1–48.
| Fitting linear mixed-effects models using lme4.Crossref | GoogleScholarGoogle Scholar |
Bobbink R (1991) Effects of nutrient enrichment in Dutch chalk grassland. Journal of Applied Ecology 28, 28–41.
| Effects of nutrient enrichment in Dutch chalk grassland.Crossref | GoogleScholarGoogle Scholar |
Borer ET, Seabloom EW, Gruner DS, Harpole WS, Hillebrand H, Lind EM, Adler PB, Alberti J, Anderson TM, Bakker JD, Biederman L, Blumenthal D, Brown CS, Brudvig LA, Buckley YM, Cadotte M, Chu C, Cleland EE, Crawley MJ, Daleo P, Damschen EI, Davies KF, DeCrappeo NM, Du G, Firn J, Hautier Y, Heckman RW, Hector A, HilleRisLambers J, Iribarne O, Klein JA, Knops JMH, La Pierre KJ, Leakey ADB, Li W, MacDougall AS, McCulley RL, Melbourne BA, Mitchell CE, Moore JL, Mortensen B, O’Halloran LR, Orrock JL, Pascual J, Prober SM, Pyke DA, Risch AC, Schuetz M, Smith MD, Stevens CJ, Sullivan LL, Williams RJ, Wragg PD, Wright JP, Yang LH (2014) Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508, 517–520.
| Herbivores and nutrients control grassland plant diversity via light limitation.Crossref | GoogleScholarGoogle Scholar | 24670649PubMed |
Bouyoucos GJ (1962) Hydrometer method improved for making particle size analysis of soils. Agronomy Journal 54, 464–465.
| Hydrometer method improved for making particle size analysis of soils.Crossref | GoogleScholarGoogle Scholar |
Cardoso EJ, Vasconcellos RL, Bini D, Miyauchi MY, Santos CA, Alves PR, Paula AM, Nakatani AS, Pereira JD, Nogueira MA (2013) Soil health: looking for suitable indicators. What should be considered to assess the effects of use and management on soil health? Scientia Agricola 70, 274–289.
| Soil health: looking for suitable indicators. What should be considered to assess the effects of use and management on soil health?Crossref | GoogleScholarGoogle Scholar |
Caughley G, Shepherd N, Short J (1987). ‘Kangaroos: their ecology and management in the sheep rangelands of Australia’. (Cambridge University Press: New York, USA)
Chalmers A, McIntyre S, Whalley RDB, Reid N (2005) Grassland species response to soil disturbance and nutrient enrichment on the northern tablelands of New South Wales. Australian Journal of Botany 53, 485–499.
| Grassland species response to soil disturbance and nutrient enrichment on the northern tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Chapman SK, Langley JA, Hart SC, Koch GW (2006) Plants actively control nitrogen cycling: uncorking the microbial bottleneck. New Phytologist 169, 27–34.
| Plants actively control nitrogen cycling: uncorking the microbial bottleneck.Crossref | GoogleScholarGoogle Scholar | 16390416PubMed |
Cingolani AM, Noy-Meir I, Díaz S (2005) Grazing effects on rangeland diversity: a synthesis of contemporary models. Ecological Applications 15, 757–773.
| Grazing effects on rangeland diversity: a synthesis of contemporary models.Crossref | GoogleScholarGoogle Scholar |
Colwell JD (1963) The optimization of the phosphorus fertilizer requirements of wheat in southern New South Wales by soil analysis. Australian Journal of Experimental Agriculture and Animal Husbandry 3, 190–197.
| The optimization of the phosphorus fertilizer requirements of wheat in southern New South Wales by soil analysis.Crossref | GoogleScholarGoogle Scholar |
Cunningham GN, Mulham WE, Milthorpe PL, Leigh JH (2011). ‘Plants of Western New South Wales’. (CSIRO Publishing: Canberra).
Delgado-Baquerizo M, Maestre FT, Gallardo A, Bowker MA, Wallenstein MD, Quero JL, Ochoa V, Gozalo B, García-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira JA, Chaieb M, Conceição AA, Derak M, Eldridge DJ, Escudero A, Espinosa CI, Gaitán J, Gatica MG, Gómez-González S, Guzman E, Gutiérrez JR, Florentino A, Hepper E, Hernández RM, Huber-Sannwald E, Jankju M, Liu J, Mau RL, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez E, Ramírez-Collantes DA, Romão R, Tighe M, Torres D, Torres-Díaz C, Ungar ED, Val J, Wamiti W, Wang D, Zaady E (2013) Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 502, 672–676.
| Decoupling of soil nutrient cycles as a function of aridity in global drylands.Crossref | GoogleScholarGoogle Scholar | 24172979PubMed |
Ding J, Travers SK, Delgado‐Baquerizo M, Eldridge DJ (2019) Multiple trade‐offs regulate the effects of woody plant removal on biodiversity and ecosystem functions in global rangelands. Global Change Biology 26, 709–720.
| Multiple trade‐offs regulate the effects of woody plant removal on biodiversity and ecosystem functions in global rangelands.Crossref | GoogleScholarGoogle Scholar | 31518466PubMed |
Dorrough J, Scroggie MP (2008) Plant responses to agricultural intensification. Journal of Applied Ecology 45, 1274–1283.
| Plant responses to agricultural intensification.Crossref | GoogleScholarGoogle Scholar |
Driscoll DA (2019) The Kosciuszko science accord. Ecological Management & Restoration 20, 75–76.
| The Kosciuszko science accord.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ (1998) Trampling of microphytic crusts on calcareous soils, and its impact on erosion under rain-impacted flow. Catena 33, 221–239.
| Trampling of microphytic crusts on calcareous soils, and its impact on erosion under rain-impacted flow.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Delgado-Baquerizo M (2018) Grazing reduces the capacity of landscape function analysis to predict regional-scale nutrient availability or decomposition, but not total nutrient pools. Ecological Indicators 90, 494–501.
| Grazing reduces the capacity of landscape function analysis to predict regional-scale nutrient availability or decomposition, but not total nutrient pools.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Simpson R (2002) Rabbit Oryctolagus cuniculus L. impacts on vegetation and soils, and implications for management of wooded rangelands. Basic and Applied Ecology 3, 19–29.
| Rabbit Oryctolagus cuniculus L. impacts on vegetation and soils, and implications for management of wooded rangelands.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Soliveres S (2014) Are shrubs really a sign of ecosystem degradation? Disentangling the myths and truths of woody encroachment in Australia. Australian Journal of Botany 62, 594–608.
| Are shrubs really a sign of ecosystem degradation? Disentangling the myths and truths of woody encroachment in Australia.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Soliveres S, Bowker MA, Val J (2013) Grazing dampens the positive effects of shrub encroachment on ecosystem functions in a semi-arid woodland. Journal of Applied Ecology 50, 1028–1038.
| Grazing dampens the positive effects of shrub encroachment on ecosystem functions in a semi-arid woodland.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Poore AGB, Ruiz-Colmenero M, Letnic M, Soliveres S (2016) Ecosystem structure, function and composition in rangelands are negatively affected by livestock grazing. Ecological Applications 26, 1273–1283.
| Ecosystem structure, function and composition in rangelands are negatively affected by livestock grazing.Crossref | GoogleScholarGoogle Scholar | 27509764PubMed |
Eldridge DJ, Delgado-Baquerizo M, Travers SK, Val J, Oliver I (2017) Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient. Journal of Applied Ecology 54, 976–985.
| Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient.Crossref | GoogleScholarGoogle Scholar |
Eldridge DJ, Delgado-Baquerizo M, Travers SK, Val J, Oliver I (2018) Livestock grazing and forest structure regulate the assembly of ecological clusters within plant networks in eastern Australia. Journal of Vegetation Science 29, 788–797.
| Livestock grazing and forest structure regulate the assembly of ecological clusters within plant networks in eastern Australia.Crossref | GoogleScholarGoogle Scholar |
FAO (2015) Food and Agriculture Organisation global map of aridity – 10 arc minutes. Available at http://www.fao.org/geonetwork/srv/en/ [Verified 15 September 2015]
Fensham RJ, Fairfax RJ, Ward DP (2009) Drought-induced tree death in savanna. Global Change Biology 15, 380–387.
| Drought-induced tree death in savanna.Crossref | GoogleScholarGoogle Scholar |
Gilbert B, Lechowicz MJ (2005) Invasibility and abiotic gradients: the positive correlation between native and exotic plant diversity. Ecology 86, 1848–1855.
| Invasibility and abiotic gradients: the positive correlation between native and exotic plant diversity.Crossref | GoogleScholarGoogle Scholar |
Golluscio RA, Austin AT, García Martínez GC, Gonzalez-Polo M, Sala OE, Jackson RB (2009) Sheep grazing decreases organic carbon and nitrogen pools in the Patagonian Steppe: combination of direct and indirect effects. Ecosystems 12, 686–697.
| Sheep grazing decreases organic carbon and nitrogen pools in the Patagonian Steppe: combination of direct and indirect effects.Crossref | GoogleScholarGoogle Scholar |
Grace JB (2006) ‘Structural equation modeling and natural systems.’ (Cambridge University Press: Cambridge, UK)
Groves RH, Austin MP, Kaye PE (2003) Competition between Australian native and introduced grasses along a nutrient gradient. Austral Ecology 28, 491–498.
| Competition between Australian native and introduced grasses along a nutrient gradient.Crossref | GoogleScholarGoogle Scholar |
Howard KSC, Eldridge DJ, Soliveres S (2012) Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic and Applied Ecology 13, 159–168.
| Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland.Crossref | GoogleScholarGoogle Scholar |
Johnson CN, Jarman PJ (1987) Macropod studies at Wallaby Creek. VI: A validation of the use of dung- pellet counts for measuring absolute densities of populations of macropodids. Australian Journal of Wildlife Research 14, 139–146.
| Macropod studies at Wallaby Creek. VI: A validation of the use of dung- pellet counts for measuring absolute densities of populations of macropodids.Crossref | GoogleScholarGoogle Scholar |
Keith D (2004) ‘Ocean shores to desert dunes: the native vegetation of NSW and ACT’. (NSW Office of Environment and Heritage: Sydney)
Kooijman AM, van der Meulen F (1996) Grazing as a control against ‘grass-encroachment’ in dry dune grasslands in the Netherlands. Landscape and Urban Planning 34, 323–333.
| Grazing as a control against ‘grass-encroachment’ in dry dune grasslands in the Netherlands.Crossref | GoogleScholarGoogle Scholar |
Lambers H, Brundrett MC, Raven JA, Hopper SD (2010) Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant and Soil 334, 11–31.
| Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies.Crossref | GoogleScholarGoogle Scholar |
Leigh JH, Wood DH, Holgate MD, Slee A, Stanger MD (1989) Effect of rabbit and kangaroo grazing on two semi- arid grassland communities in central-western New South Wales. Australian Journal of Botany 37, 375–396.
| Effect of rabbit and kangaroo grazing on two semi- arid grassland communities in central-western New South Wales.Crossref | GoogleScholarGoogle Scholar |
Lezama F, Baeza S, Altesor A, Cesa A, Chaneton EJ, Paruelo JM (2013) Variation of grazing-induced vegetation changes across a large-scale productivity gradient. Journal of Vegetation Science 25, 8–21.
| Variation of grazing-induced vegetation changes across a large-scale productivity gradient.Crossref | GoogleScholarGoogle Scholar |
Li S, Cadotte MW, Meiners SJ, Hua Z, Shu H, Li J, Shu W (2015) The effects of phylogenetic relatedness on invasion success and impact: deconstruction Darwin’s naturalisation conundrum. Ecological Letters 18, 1285–1292.
| The effects of phylogenetic relatedness on invasion success and impact: deconstruction Darwin’s naturalisation conundrum.Crossref | GoogleScholarGoogle Scholar |
Lunt ID (2002) Grazed, burnt and, cleared: how ecologists have studied century-scale vegetation changes in Australia. Australian Journal of Botany 50, 391–407.
| Grazed, burnt and, cleared: how ecologists have studied century-scale vegetation changes in Australia.Crossref | GoogleScholarGoogle Scholar |
Lunt ID, Eldridge DJ, Morgan JW, Witt GB (2007) A frame-work to predict the effects of livestock grazing and grazing exclusion on conservation values in natural ecosystems in Australia. Australian Journal of Botany 55, 401–415.
| A frame-work to predict the effects of livestock grazing and grazing exclusion on conservation values in natural ecosystems in Australia.Crossref | GoogleScholarGoogle Scholar |
Maestre FT, Puche MD (2009) Indices based on surface indicators predict soil functioning in Mediterranean semi-arid steppes. Applied Soil Ecology 41, 342–350.
| Indices based on surface indicators predict soil functioning in Mediterranean semi-arid steppes.Crossref | GoogleScholarGoogle Scholar |
Mellado M (2016) Dietary selection by goats and the implications for range management in the Chihuahuan Desert: a review. The Rangeland Journal 38, 331–341.
| Dietary selection by goats and the implications for range management in the Chihuahuan Desert: a review.Crossref | GoogleScholarGoogle Scholar |
Myers K, Poole W (1963) A study of the biology of the wild rabbit, Oryctolagus cuniculus (L.), in confined populations IV. The effects of rabbit grazing on sown pastures. Australian Wildlife Research 2, 11–32.
Noble JC (1998) ‘The delicate and noxious scrub: CSIRO studies on native tree and shrub proliferation in the semi-arid woodlands of eastern Australia’. (CSIRO Publishing: Lyneham, Australia)
Olson BE, Lacey RJ (1994) Sheep: a method for controlling rangeland weeds. Sheep Research Journal 1994, 105–112.
Olson BE, Wallander RT (1998) Effect of sheep grazing on a leafy spurge-infested Idaho fescue community. Journal of Range Management 51, 247–252.
| Effect of sheep grazing on a leafy spurge-infested Idaho fescue community.Crossref | GoogleScholarGoogle Scholar |
Page MJ, Beeton RJS (2000) Is the removal of domestic stock sufficient to restore semi-arid conservation areas? Pacific Conservation Biology 6, 245–253.
| Is the removal of domestic stock sufficient to restore semi-arid conservation areas?Crossref | GoogleScholarGoogle Scholar |
Pineiro G, Paruelo JM, Oesterheld M (2006) Potential long-term impacts of livestock introduction on carbon and nitrogen cycling in grasslands of Southern South America. Global Change Biology 12, 1267–1284.
| Potential long-term impacts of livestock introduction on carbon and nitrogen cycling in grasslands of Southern South America.Crossref | GoogleScholarGoogle Scholar |
Popay I, Field R (1996) Grazing animals as weed control agents. Weed Technology 10, 217–231.
| Grazing animals as weed control agents.Crossref | GoogleScholarGoogle Scholar |
Pringle HJR, Landsberg J (2004) Predicting the distribution of livestock grazing pressure in rangelands. Austral Ecology 29, 31–39.
| Predicting the distribution of livestock grazing pressure in rangelands.Crossref | GoogleScholarGoogle Scholar |
Prober SM, Lunt ID (2009) Restoration of Themeda australis swards suppresses soil nitrate and enhances ecological resistance to invasion by exotic annuals. Biological Invasions 11, 171–181.
| Restoration of Themeda australis swards suppresses soil nitrate and enhances ecological resistance to invasion by exotic annuals.Crossref | GoogleScholarGoogle Scholar |
Prowse TAA, O’Connor PJ, Collard SJ, Rogers DJ (2019) Eating away at protected areas: Total grazing pressure is undermining public land conservation. Global Ecology and Conservation 20, e00754
| Eating away at protected areas: Total grazing pressure is undermining public land conservation.Crossref | GoogleScholarGoogle Scholar |
Pywell RF, Hayes MJ, Tallowin JB, Walker KJ, Meek WR, Carvell C, Warman LA, Bullock JM (2010) Minimizing environmental impacts of grassland weed management: can Cirsium arvense be controlled without herbicides? Grass and Forage Science 65, 159–174.
| Minimizing environmental impacts of grassland weed management: can Cirsium arvense be controlled without herbicides?Crossref | GoogleScholarGoogle Scholar |
Reisner MD, Grace JB, Pyke DA, Doescher PS (2013) Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. Journal of Applied Ecology 50, 1039–1049.
| Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems.Crossref | GoogleScholarGoogle Scholar |
Smit C, Ruifrok JL, van Klink R, Olff H (2015) Rewilding with large herbivores: the importance of grazing refuges for sapling establishment and wood-pasture formation. Biological Conservation 182, 134–142.
| Rewilding with large herbivores: the importance of grazing refuges for sapling establishment and wood-pasture formation.Crossref | GoogleScholarGoogle Scholar |
Socher SA, Prati D, Boch S, Müller J, Baumbach H, Gockel S, Fischer M (2013) Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions. Basic and Applied Ecology 14, 126–136.
| Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions.Crossref | GoogleScholarGoogle Scholar |
Soliveres S, Eldridge DJ, Maestre FT, Bowker MA, Tighe M, Escudero A (2011) Microhabitat amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: towards a unifying framework. Perspectives in Plant Ecology, Evolution and Systematics 13, 247–258.
| Microhabitat amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: towards a unifying framework.Crossref | GoogleScholarGoogle Scholar | 25914601PubMed |
Spooner PG, Briggs SV (2008) Woodlands on farms in southern New South Wales: a longer‐term assessment of vegetation changes after fencing. Ecological Management & Restoration 9, 33–41.
| Woodlands on farms in southern New South Wales: a longer‐term assessment of vegetation changes after fencing.Crossref | GoogleScholarGoogle Scholar |
Tiver F, Andrew MH (1997) Relative effects of herbivory by sheep, rabbits, goats and kangaroos on recruitment and regeneration of shrubs and trees in eastern South Australia. Journal of Applied Ecology 34, 903–914.
| Relative effects of herbivory by sheep, rabbits, goats and kangaroos on recruitment and regeneration of shrubs and trees in eastern South Australia.Crossref | GoogleScholarGoogle Scholar |
Tongway D (1995) Monitoring soil productive potential. Environmental Monitoring and Assessment 37, 303–318.
| Monitoring soil productive potential.Crossref | GoogleScholarGoogle Scholar | 24197857PubMed |
Travers SK, Eldridge DJ, Dorrough J, Val J, Oliver I (2018) Introduced and native herbivores have different effects on plant composition in low productivity ecosystems. Applied Vegetation Science 21, 45–54.
| Introduced and native herbivores have different effects on plant composition in low productivity ecosystems.Crossref | GoogleScholarGoogle Scholar |
Travers SK, Eldridge DJ, Val J, Oliver I (2019) Rabbits and livestock grazing alter the structure and composition of mid-storey plants in a wooded dryland. Agriculture, Ecosystems & Environment 277, 53–60.
| Rabbits and livestock grazing alter the structure and composition of mid-storey plants in a wooded dryland.Crossref | GoogleScholarGoogle Scholar |
van Auken OW (2009) Causes and consequences of woody plant encroachment into Western North American grasslands. Journal of Environmental Management 90, 2931–2942.
| Causes and consequences of woody plant encroachment into Western North American grasslands.Crossref | GoogleScholarGoogle Scholar | 19501450PubMed |
Vandandorj S, Eldridge DJ, Travers SK, Delgado-Baquerizo M (2017) Contrasting effects of aridity and grazing intensity on multiple ecosystem functions and services in Australian woodlands. Land Degradation & Development 28, 2098–2108.
| Contrasting effects of aridity and grazing intensity on multiple ecosystem functions and services in Australian woodlands.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Norton DA, Hobbs RJ (2000) Grazing effects on plant cover, soil and microclimate in fragmented woodlands in south-western Australia: implications for restoration. Austral Ecology 25, 36–47.
| Grazing effects on plant cover, soil and microclimate in fragmented woodlands in south-western Australia: implications for restoration.Crossref | GoogleScholarGoogle Scholar |