

Australian corticolous myxomycetes: models of distribution and development
Peter Wellman
17 Warragamba Avenue, Duffy, ACT 2611, Australia. Email: wellmanp@iinet.net.au
Australian Journal of Botany 67(8) 617-629 https://doi.org/10.1071/BT19155
Submitted: 22 September 2019 Accepted: 17 December 2019 Published: 10 February 2020
Journal compilation © CSIRO 2019 Open Access CC BY-NC-ND
Abstract
This paper presents an integrated model of the variation over a continental landmass of myxomycetes, a single-celled organism in the phylum Amoebozoa. Bark samples were collected on long traverses across Australia, and cultivated in Petri dishes by the moist chamber technique to obtain large assemblages of common species. The results of this survey and previous surveys are consistent with there being four major myxomycete assemblages: Tropical, Northern Arid, Southern Arid and Temperate. Where mapped, these species assemblage regions are consistent with the Australian phytogeographical regions. The myxomycetes differ between arid and non-arid areas; the arid areas have slightly higher productivity per wetting event, with members of the Physarales and Liceales relatively important and the Stemonitidales, Trichiales and Cribrariales less important. When the bark samples are placed in a moist culture there is a myxomycete growth cycle and then the population declines to resting phases. The population increase during a growth phase can be modelled by a linear plot of log(abundance) against species rank, where abundance is total harvested spore volume of a species. The population decline appears to be linear from two weeks after watering, declining to negligible activity 4 weeks after watering.
Additional keywords: biogeography, ecology, ecosystem dynamics.
Introduction
Myxomycetes (plasmodial slime moulds) are phagotropic eukaryotic single-celled organisms in the phylum Amoebozoa. Their life cycle has two very different feeding stages: a relatively small uninucleate phase; and a larger multinucleate plasmodium. They feed when there is moisture but otherwise survive in three types of dormant stages (microcysts, sclerotia and spores). Because of the numerous types of dormant phases myxomycetes are relatively successful in arid environments. Fruiting bodies (containing the spores) are permanent structures, and are the basis of the species identification. Myxomycetes are the only living organisms that have both a single celled phase living as a true microorganism and can be identified from a permanent structure. They are easily cultivated in the laboratory, so they are relatively easy to study.
Myxomycetes are normally identified from their fruiting bodies. The fruiting bodies can be obtained either from collecting fruiting bodies in the field after rain (which is difficult in central Australia), or from cultivating suitable substrates in the laboratory using the moist chamber culture technique (Stephenson and Stempen 1994; Novozhilov et al. 2017; Wrigley de Basanta and Estrada-Torres 2017). Previous overseas work had shown that dead bark off a live tree was the most productive material for moist chamber culture (e.g. Schnittler 2001), so this substrate was used in this study.
The result of all the previous worldwide work on myxomycetes was summarised by Stephenson and Rojas (2017). Most of the work to date in natural habitats has been in describing all species, finding which species are present in what substrates, and investigating which environmental variables affect the presence of a species, with each paper generally considering a single climatic area. Previous work on Australian myxomycetes with this objective consists of Ing and Spooner (1994), Mitchell (1995), McHugh et al. (2003), Black et al. (2004), Jordan et al. (2006), Rosing et al. (2007), Davison et al. (2008, 2017), Wrigley de Basanta et al. (2008), McHugh et al. (2009), Stephenson and Shadwick (2009), Knight and Brims (2010), Lloyd (2014), and Wellman (2017).
In this paper and two previous papers (see Wellman 2015, 2016) myxomycetes have been studied in a different way. Taken together, the sampling covers the whole continent, and the studies concentrate on the local population of common species rather than the rare species. To achieve these objectives the procedure and emphasis differed somewhat from normal; rather than collection of spot samples in a compact area, samples were collected systematically in very long traverses (to 2000 km long) sampling all the Australian regions. The growth procedure was not optimised to find the maximum number of species but to study competing, large multi-species populations. As a result of this different approach, topics can studied that were not previously explored, such as modelling of the myxomycete growth and decay after wetting, the variation of productivity and order dominance over a continent, and the evidence for myxomycete-geographical regions. The ecological conclusions of this paper are consistent with previous international work, but the paper shows how these conclusions can be integrated on a continental scale.
The moist chamber technique simulates a rainfall event. However, the results from moist chamber cultivation are unlikely to be replicated during actual normal rainfall events in the countryside because it is unusual in Australia for bark to be damp continuously for 4 weeks. The moist chamber experiments are important because the results determine the myxomycete productivity of the bark samples under standard conditions, and can be used to model population dynamics.
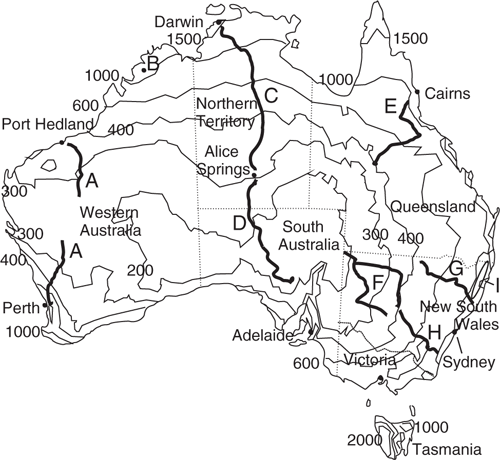
The intention of this paper, and the earlier traverses incorporated in this paper, is to sample the corticolous myxomycetes over the main Australian landmass. Because the main reason for changes in species is likely to be the change in the wetting events, the sampling was designed to determine the change in species assemblages with changes in rainfall. The mean annual rainfall (Fig. 1) has a low of 200 mm/year in the central south of Australia, and a gradual then abrupt increase towards the coastline. The coastal areas with high rainfall occur along the northern margin of Australia, the east coast and in the south-west corner. Both the previous and new sampling was carried out predominantly on long traverses on tar-sealed roads, in the direction of the rainfall gradient (Fig. 1). There is a long north–south traverse across Western Australia with a gap in the middle, a traverse across the centre of Australia, and several short traverses in eastern Australia. Together the traverses sample the main rainfall variation, importantly crossing the lowest annual rainfall area of Australia. A small study was made of the bark in trees in wet temperate rainforest in the north-east highlands of New South Wales.
|
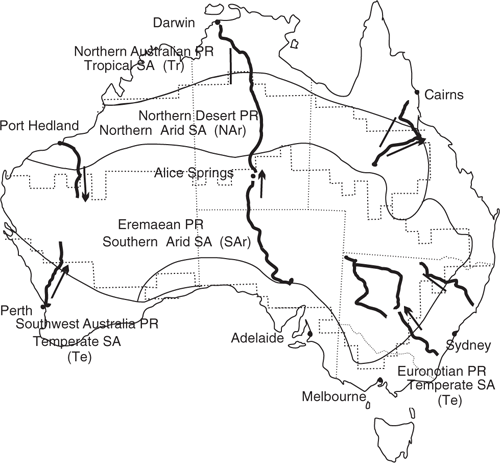
In a previous paper (Wellman 2016), I showed that in Australia, myxomycete species assemblages along long profiles appeared to be constant over long distances, and then there is a wide transition zone to another constant species assemblage. I recognised four species assemblages: a tropical, northern dry, southern dry, and temperate. As the word ‘dry’ is inaccurate, this paper uses the names Tropical (Tr), Northern Arid (NAr), Southern Arid (SAr), and Temperate (Te), for these species assemblages. To prove this model for all Australia we need to determine that: (1) species assembles are reasonably constant within profiles; (2) transition zones are where they would be expected geographically; and (3) the four species assemblages are reasonably constant east–west across Australia. Also of interest is whether the myxomycete species assemblage boundaries correlate with the phytogeographical regions defined by the maxima in plant species turnover. The collected myxomycete traverses are positioned to have two crossings of each of the three main phytogeographical region boundaries (Fig. 2).
|
Materials and methods
The method used in the previous long traverses (Wellman 2015, 2016), and the survey reported here, are similar and are described below. The bark sampling was generally conducted during the winter, as the winter temperature is pleasant and, for most of Australia, rain in winter is unlikely. Most samples were collected at least a month after the last rainfall. In Western Australia there was unseasonal rain a week before sampling both when the whole traverse was sampled in July and also when the southern end was re-sampled in November.
The moist culture technique was used in this study. Most people using the moist culture are mainly trying to obtain the range of species found at a site. Many of these studies used only one piece of material, or several related pieces, for each Petri dish. This study tried to obtain, for a large number of samples, the range of common species and their relative productivity. Because of this difference in survey objective, the field sampling and moist culture in this survey were conducted slightly differently from most previous studies. (1) At each ‘site’ dead-bark pieces were collected from 1–4 trees of the same species, from a range of 1–2.4 m above the ground, from random positions around the tree(s), and from the available range of the bark textures. (2) Five Petri dishes were cultivated from each site. Each Petri dish contained if possible a full range of the types of bark. If long strips of bark were collected, then each strip was cut into pieces and one piece put in each dish. (3) The bark samples in the Petri dishes were kept moist, and at weekly intervals the Petri dishes were inspected and the mature fruiting bodies harvested. Generally there were no further fruiting bodies after 4 weeks and the sample was dried out after ~6 weeks. Samples 860–1087 (of Wellman 2015, 2016) were subject to another cycle of soaking and cultivation, but few additional fruiting bodies were obtained. The fruiting bodies from the five Petri dish were not kept separate. The few fruiting bodies on the base of the bark pieces were only collected when the bark pieces had dried. The estimates from the number of fruiting bodies for each species is based on counting or estimating the number of harvested fruiting bodies, a number supplemented, when much of the bark was covered by fruiting bodies, by an estimate of the number of fruiting bodies never harvested. This procedure aimed at getting a large diverse species assemblage, and getting a large number of fruiting bodies so that their relative productivity can be better determined.
The pH of the bark was measured for 56 samples in Western Australia, 55 in Northern Territory and Queensland, and 11 near Canberra. The measurements were made to an accuracy of ~0.1 pH unit, by measuring deep fluid in a narrow plastic cup with an inexpensive pH instrument that was calibrated using standard solutions.
The myxomycete fruiting bodies were identified using a wide range of sources, but mainly Ing (1999), Poulain et al. (2011) and Discover Life (2018). In order that the bulk of the Australian myxomycete identifications were consistent, the more difficult to identify species of the previous publications (Wellman 2015, 2016) were re-identified. Identifications in some species groups were difficult. In Physarum/Badhamia this was because the capillitium structure was generally not clear and spore diameters of possible species were similar. Echinostelium minutum and Echinostelium arboreum could be identified, but reliable identification of the other species of Echinostelium could not be obtained because the material used for identification was dried (Whitney 1980). The separation and identification of most Paradiacheopsis species was not attempted because many samples were dominated by Comatricha elegans or Comatricha ellae, and the similarity of the smaller fruiting bodies of these species to Paradiacheopsis fruiting bodies made separation of Paradiacheopsis fruiting bodies, and their identification and counting, uncertain.
Results
Supplementary Material File S1 lists the sample localities, and File S2 the species collected and for each record the locality and number of fruiting bodies. The surveys resulted in spreadsheets listing, for each species-record, the sample number, species, and number of fruiting bodies (except for Echinostelium). The average size of the fruiting body for each species was estimated, mainly using Poulain et al. (2011). Using these sizes the spreadsheet was used to calculate the total volume of spores for each species record, and the total volume of spores for each sample. The errors of the spore volumes are substantial, but the paper is mainly using the logarithms of the volumes. Spore volumes have previously been used as mentioned in Novozhilov et al. (2017).
Species assemblages
The results of the new surveys along long profiles are presented in the form of tables of occurrences (see Tables S1–S4, available as Supplementary Material to this paper). To save space in the tables, a sample is not shown if it contains only one common species, and a species is not shown if it has only one occurrence. Where the survey abuts a previously published survey, then the species distribution of the adjacent part of the previous survey is summarised in one column.
Table S1 shows a 500 km long north-west-trending traverse in north-east New South Wales between Yarrowitch and Goodooga (Fig. 1, traverse G), collected July 2017, with samples numbers 1129–1172. The species are divided into three groups: 14 species that occur along the whole survey line, 10 SAr species, and six Te species. The transition zone is ~250 km wide with a midpoint ~29.6°S, 149.2°E, located near Narribri.
Table S2 shows a 450 km long north-west-trending traverse in south-east New South Wales between Canberra and Lake Cargelligo (Fig. 1, traverse H), with sample numbers 1101–1111 collected October 2016, and 1254–1291 mainly collected June–July 2017. The species are divided into three groups: twelve species that occur along the whole survey line, five SAr species, and eight Te species. The transition zone is ~200 km wide with a midpoint ~33.6°S, 146.1°E near West Wyalong, but it is poorly defined at its north-west margin.
Table S3 shows an 1100 km long south trending traverse in Northern Territory and South Australia between Alice Springs and Woomera (Fig. 1, traverse D), with sample numbers 1201–1251 collected June 2017. The species are divided into three groups. Most of the species (13) occur throughout the traverse and occur elsewhere in both the SAr and NAr assemblages. Three species should occur both in the NAr and SAr, but they only extend south to sample 1208. Three species characteristic of the SAr occur on the southern end of the traverse and only extend north to sample 1220, not as far north as the likely transition zone. In this traverse the SAr/NAr transition zone is very poorly defined in the north, but it is likely to be over samples 1201–1208, just south of Alice Springs, with a midpoint at 24.3°S, 133.8°E.
Table S4 shows a 1300 km long, south-trending traverse in Western Australia from near Port Hedland in the north to Perth in the south (Fig. 1, traverse A). Sample numbers 1312–1420 were collected July 2018, and 1450–1502 were collected November 2018. There is a 340 km gap in this traverse between 24.5 and 27.6°S. The data in this table are not shown in full because there are 89 samples along the profile. The table has been drastically shortened by summarising the occurrences for the two transition zones. The expected extent of the species that should terminate at a transition zone are shown by the underlining. The interpretation is the NAr/SAr transition zone is 16 samples wide, between samples 1362 and 1340, or further south, giving a transition zone of 250+ km wide with a midpoint ~23.3°S, 119.7°E, near Newman. The SAr/Te transition zone is interpreted to be 32 samples wide, between samples 1425 and 1454 or further north; that is 260 km wide and with a midpoint at 30.2°S, 116.7°E near Mount Magnet town-ship. The results from this traverse are puzzling, as the sample spore volumes are high, but key species of NAr and Te do not appear as frequently as expected. This may be due to the rain before the sampling.
Discussion
The analysis in this discussion section interprets the results of both the new and the previously published corticolous myxomycete work – the datasets shown in Fig. 1 and additional spot sample data. A total of 300 long traverse samples were used.
Table 1 shows the occurrence of myxomycete species in each of the four species assemblage areas using this new data, and the previously published data. Not all species can be shown because the table would be too large, so the contents of the table concentrate on the significant species found in the long profile studies, and leaves out many species found in low numbers in the wetter areas. Columns 2–11, 13 and 14 show the data from the long traverses of Wellman (2015, 2016) and of this paper. The myxomycete records within the transition zones are recorded as follows: if the species is present in both species associations then a record in the transition zone is put in the closest species association, if the record is found in only one of the species associations then all the records within the transition zone are allocated to that species association. The other columns sample the wetter parts of the Tropical and Temperate Species Assemblages. Column 1 (Tr2) is a composite sample of the wetter Tropical areas in the Kimberly, Darwin and Cairns areas: sites CB, CO, CT, EB, FI, MJ, WF of McHugh et al. (2003), sites 1–11, 13 of McHugh et al. (2009), and sites 10–12 of Davison et al. (2017). Column 12 is a composite sample of the temperate areas of SW of Western Australia combining the records of the traverse (Table S4), of Jordan et al. (2006) from Banksia bark for two sites, and spot samples of sites 19, 23–25 and 28 of McHugh et al. (2009). Column 15 (Te2) comprises spot samples from the wetter part of temperate south-eastern New South Wales (Wellman 2016) and the wetter part of temperate Victoria (Rosing et al. 2007). Column 16 (Te3) reports a few samples from cool-temperate closed rainforest in New South Wales, mainly Nothofagus bark (traverse I of Fig. 1). Columns 18–20 summarise the data for columns 1–16, giving the sum of the species records in each of the four species assemblages expressed as parts per thousand. In these four columns significant species occurrences are in bold. These are generally species with abundances >10‰.

|
For much of this table the interpretation is relatively simple. For instance, Licea kleistobolus is relatively common in all four species assemblages, and Cribraria confusa is present only in the tropical and temperate species assemblages. The difficulty is in determining the significance of relative abundance differences (such as Licea biforis in columns 17–20), and in estimating the abundance (occurrence rate) of outliers for this organism spread by spores.
Species characteristic of all four species assemblages (Tr, NAr, SAr, Te) are Arcyria cinerea, A. sp. H (Arcyria sp. of Davison et al. 2008), Calomyxa metallica, Comatricha elegans, C. ellae, Licea biforis, and L. kleistobolus. Species characteristic of the wetter species assemblages (Tr, Te) are Clasterderma debaryanum, Cribraria confusa, Cribraria minutissima, Cribraria violacea, and Echinostelium minutum. Species characteristic of the arid species assemblages (NAr, SAr) are Badhamiopsis ainoae, Didymium dubium, Mcbrideola oblonga, and Physarum decipiens. The Tr assemblage differs from the Te assemblage in having, or having in greater numbers, Hemitrichia minor, Perichaena chrysosperma, Stemonitis mussooriensis, Trichia erecta, and Trichia munda. The Te assemblage differs from the Tr assemblage in having, or having in greater numbers, Arcyria pomiformis, and Trichia contorta. The NAr assemblage differs from the SAr assemblage in having, or having in greater numbers Macbrideola decapilata and Perchaena corticalis. The SAr assemblage differs from the NAr assemblage in having, or having in greater numbers Dianema corticatum, Didymium dubium, Enerthenema papillatum, and Physarum crateriforme. There is a major change in the species assemblage between the non-arid (Tr, Te) and arid (NAr, SAr) species assemblages, and only a minor change between the non-arid assemblages (Tr, Te), and between the arid assemblages (NAr, SAr).
Table 1 shows the consistency of the occurrence of a species within a species association over a continental area. Table 1 and the profiles of Tables S1–S4 demonstrate that species can occur outside their normal range as outliers. The distinction between normal range and outliers is important in interpreting World maps of myxomycete record sites, such as in Discover Life (2018).
Tree bark acidity (pH) and characteristics
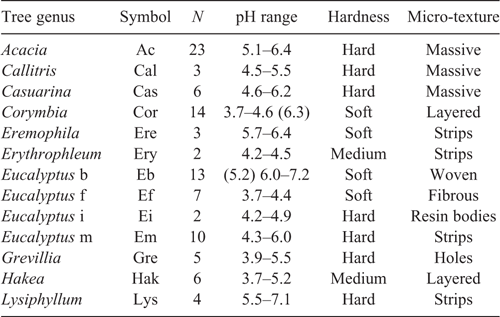
Table 2 shows for each myxomycete species the range of observed pH and the host trees, while Table 3 gives for the common tree genera the range of pH, and the type of bark. Each myxomycete species grows on a range of bark pH, and a range of bark texture. Examples of the preference differences between related myxomycete species are as follows. Arcyria cinerea grows over the enormous pH range of 3.7–7.4, but is mainly restricted to soft and fibrous barks, while Arcyria sp. H and A. pomiformis are both restricted to relatively acid bark but A. sp. H likes soft bark and A. pomiformis likes hard bark. Echinostelium arboreum differs from E. minutum in all measured factors - species association, preferred pH, and bark texture. The occurrence of myxomycete species on a tree bark is clearly determined by three factors: species association (climate), bark acidity and bark substrate properties such as hardness and texture. To sample all the potential species that would inhabit a species-assemblage area one has to sample trees with a range of bark textures and for each texture sample the whole range of bark pH. Over much of Australia the existing native trees have a full range of pH, but what is missing is generally a full range of bark textures. This is in part because the trees and tall shrubs use bark hardness and texture to limit tree damage during the wildfires, such fires being more frequent in the arid areas. Most of the tree genera collected had a relatively narrow range of bark pH and bark type. The exception is Eucalyptus, which has been divided into four bark types (Table 3). The myxomycete species observed range of substrate pH found in this survey can be compared with those found in overseas surveys with a different climate, different mean substrate range and different bark textures (Schnittler 2001; Novozhilov et al. 2006). The species in common between the three surveys have a similar tolerance of pH when allowance is made for the range of substrate pH available in a sampling area.

|
|
Table 4 shows that the bark acidity (pH) correlates with changes in the productivity and species composition for each sample. As the average productivity of the bark samples decreases from neutral (column with the range 6.0–7.3) to acidic (the range 3.1–3.9), the mean number of species records per sample decreases from 6.1 to 3.9, the mean volume of spores per sample decreases from 5.5 to 3.0 mm3, and there is an associated decrease in the ratio of Physarales records to total records from 28 to 2%. The trends shown in Table 4 are true when using only the Western Australian samples, and when only using only the rest of the samples.

|
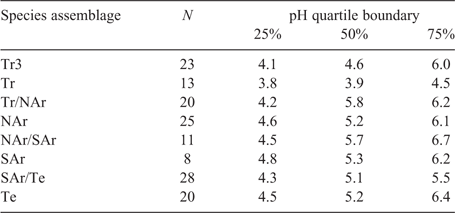
There appears to be a regional change in the average pH of tree bark within Australia (Table 5). Using the traverse data, the median pH is 5.1–5.8 over most of Australia (arid or temperate), while the median is lower for the tropical region, 3.9 for the savanna forest (Tr sample), and 4.6 for two sites in the wetter area (Tr3; sites 10 and 12 of Davison et al. 2017). Hence the influence of pH is at two scales, at a tree scale the myxomycete assemblage will vary with the pH of the individual tree bark, and at a regional scale the slight changes in the mean pH of tree bark may results in a slight regional change in myxomycete assemblage.
|
Changes in species assemblages across Australia
The information from the long traverses in Table 1 are consistent with there being four species assemblages. In the area of each species assemblage the assemblage is largely independent of mean annual rainfall changes inland from the coast, and independent of east–west changes across Australia such as timing of rainfall (winter or summer). The non-arid/arid boundary in the north is at ~650 mm mean annual rainfall, falling mainly during the summer. It is ~250 mm in the south-west with mainly winter rain, and ~500 mm in the south-east with equal amounts of summer and winter rain. Hence, on a continental scale, there is no point in plotting the various parameters against rainfall. To a first approximation the variation of myxomycetes within Australia can be summarised by plotting parameters against species assemblages. The traverse data is used to compile samples of Tr, NAr, SAr, Te and the transitions, the environments being mainly open forest, woodland with grass or scrub understory, and scrubland with isolated trees. Tr3 and Te2 document the wetter areas; Tr3 is composed of sites 10 and 12 of Davison et al. (2017) of 24 samples, and Te4 is composed of the data of Rosing et al. (2007), Wellman (2016), and column 16 of Table 1. However, Tr3 and Te4 are not ideal samples because they have low sample numbers, possibly biased sampling, and importantly are from relatively wet open forest, rather than very wet closed rainforest (which is thought to be the extreme environment).
Fig. 3 show the change, over Australia, in the character of the mean corticolous myxomycete harvest. The variation in productivity, shown by the total spore volume per sample (Fig. 3a) and by the number of species found at a site (Fig. 3b), is for a slight rise from Tr and Te to a high at the NAr/SAr transition zone. For both parameters the values are thought to be much lower in areas of wet tall open forest, and closed rainforest, but there is only poor data to support this in the Tr2 and Te2 values.
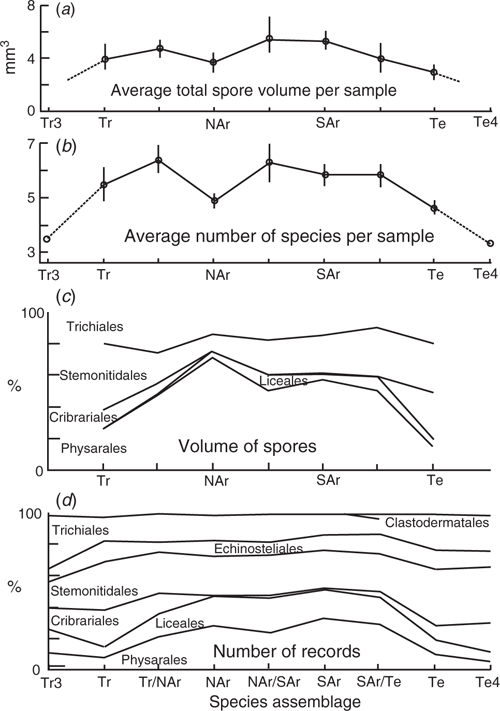
|
The relative importance of the myxomycete orders changes over Australia (expressed as a percentage of the assemblage) can be based either on the total volume of spores (Fig. 3c) or the number of records (Fig. 3d). With both methods the Echinosteliales and Clastodermatales have about the same importance in arid and non-arid areas, the Physarales and Liceales are more important in the arid areas, and the Trichiales, Cribraiales, and to a lesser extent the Stemonitidales, are more important in the non-arid areas. When measuring importance by the number of records, the orders have similar importance except for a constant low number of the Clastodermatales. When measuring importance by the volume of spores, the Physarales dominates the pattern of change. It is ~55% of the volume in the arid area, and ~20% in the non-arid areas. The Trichiales, Stemonitidales and Cribraiales together form 40% of the volume in the arid areas and 60–80% of the volume in the non-arid areas. The change from non-arid to arid characteristics is more gradual (in terms of region subdivisions and distance) in the north than in the south. In the north the change is between Tr through Tr/NAr to NAr, whereas in the south the change is between SAr/Te to Te.
Previous corticolous work, based on both the differences between local studies, and on larger regional studies (Novozhilov et al. 2017; Schnittler et al. 2017) have shown many of the correlations discussed above: decrease in abundance and richness in very high rainfall areas, increasing species diversity with increasing pH values towards neutral, and the higher proportion of Physarales in arid areas. This paper supplements these earlier papers as it shows how these variables change over a whole continent, and the difference between expressing these changes as number of species, and expressing them using volume of spores.
Histogram of total spore volume
Fig. 4 is a histogram of the total spore volume using 170 samples. It illustrates the various influences on sample total spore volume. To decrease the effects of regional variation in mean volume, the data used to construct the histogram were from a single area (Western Australia), and it excluded data from Tr and Te that Fig. 3a showed had lower mean volumes. The histogram plots volume on a logarithmic scale. The histogram has a reasonably symmetrical high, with a mean of 4 mm3, a mode at 6 mm3, and one-half the amplitude at 2 and 16 mm3. There is only a small tail on the high side, and a very long tail on the low side. In this histogram the higher volumes (>20 mm3) are likely to be samples where one species has carried considerable mass from a previous feeding event on the tree to the feeding event in the Petri dish. Lower volumes (<1 mm3) (12% of the samples) in part comprise trees with corky or insufficiently-weathered bark, possibly acid bark, and (notably) single trees that were not local, being exotic or from another part of Australia. The low volumes are thought to be due to bark with little food, or suitable myxomycete species not being available for one tree. In Fig. 4 the volumes vary by 2.5 orders of magnitude. A mean of the raw values would be dominated by the larger values. In this paper, each mean volume from many samples has been calculated using the logarithm of the values so the long tail of low values results in a mean that is much influenced by low values, hence the mean is significantly lower than the mode or median.
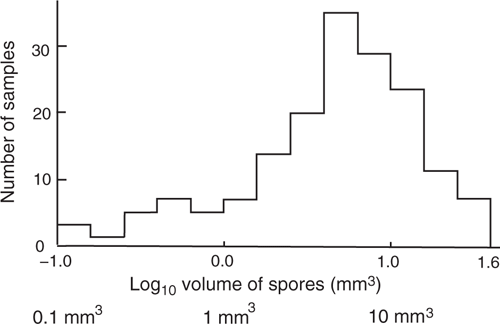
|
Modelling the pattern of fruiting during a growing event
The moist chamber technique simulates a rainfall event, so the results of the growing experiments can be used to model population dynamics. In the plant and animal kingdoms it is common to use abundance-rank plots to determine the abundance relationship of species in a large population (May 1976). Normally in these plots log(abundance) is shown on one axis, and the other axis is rank, plotted linearly, with the highest rank with a value of 1. The ecological importance of a species is dependent on the amount the species eats. With myxomycetes we can allow for the varying size of the fruiting body and the large range in the number of bodies by using total harvested fruiting body volume of each species as a measure of species abundance. A site never has a large number of species, so the abundance-rank plot of a single site is very irregular and not useful by itself. However we have a large number of sites with the same number of species, and rather than base an interpretation on a single site, we can base the interpretation on the mean path of the log(abundance)-rank plots of all sites with the same (or similar) number of species. Fig. 5 shows a log(abundance)-rank plot both for sites with 11 or 12 species, and sites with six species. The plots have been simplified in that a few sites with very different relationships have been omitted for simplicity, and species points are not shown on the plot if their total volume is <0.001 mm3, because these fruiting bodies are small and easy to overlook, so their abundance is likely to be greatly underestimated, and so incorrect. In this paper samples were plotted with a different number of species, hence the rank scale has been inverted, and species with the lowest rank have a value of 1.
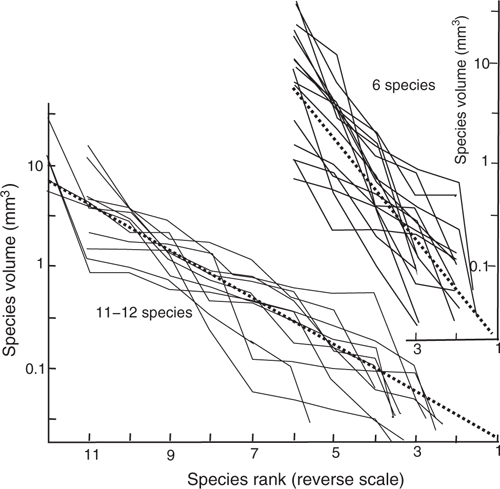
|
The log(abundance)-rank plots for samples with 11 or 12 species give tracks that are within a wide linear band for most of their extent. The average relationship for samples with 11–12 species is given by the thick linear dotted line. This line gives for the highest rank species a predicted mean total volume of ~7 mm3, and for the lowest rank species a predicted mean total volume of ~0.02 mm3. A similar pattern describes the log(abundance)-rank plots of samples with six species, with the mean line with similar highest and lowest rank values.
For flowering plants and animals a linear relation between log(abundance) and rank is taken to be consistent with species radiating into what is initially a vacant environment with food, the species growth for each species being exponential. In Fig. 5 the linearity of the mean relationship (dotted line) using a log(abundance) scale is consistent with each species having exponential growth. The volume of the highest rank species is generally only slightly less than the sample total volume, and the ~7 mm3 volume value is nearly the same as the mode value of Fig. 4. The predicted volume for the lowest rank species is ~0.02 mm3, which equates to one fruiting body with a diameter of ~0.35 mm. This value is reasonable as in this model for the lowest rank species the number of fruiting bodies should be one, and its predicted fruiting body diameter should approximate the mean of the fruiting body diameter of the sample. More importantly, the mean relationships for 11–12 species in the sample should be the same as that for six species in the sample, except that there should be 10–11 equally spaced volumes in one and six in the other. The model predicts that for samples with n species, then the n spore volumes will be equally spaced along the log scale, although the errors in the data fit will increase dramatically in samples with few species.
Individual fruiting body of different species in this study generally vary in radius from 0.03 mm for Echinostelium to ~1 mm, so their volumes range from 1.10–5 to ~0.5 mm2, and the number of fruiting body vary from 1 to 10 000, the higher numbers of fruiting body for species with the smallest fruiting body. The fruiting body volumes have a greater range than the number of fruiting body, so species with a larger fruiting body tend to have higher total volume, and species with smaller fruiting body tend to have low total volume. Two other effects may make a secondary contribution to the ranking: (1) if some species are much more efficient than others at growth and producing fruiting bodies they would be higher rank species than their fruiting body volume would warrant. Conversely if some species are relatively less efficient they would have a lower rank than their fruiting body volume would warrant; (2) predominant effects are random factors. Before growth starts in the Petri dish there is, for each species, near random numbers of the three dormant phases (spores, microcysts and sclerotium), each with different times to complete the life cycle and to develop mature fruiting bodies. So if there is no efficiency difference between species, the species position in the ranking would be random, but with a mean dependent on fruiting body volume. The observational data of this survey is that for an area with a near constant species assemblage (within a species assemblage area or transition zone), a tree genera, or Eucalyptus type, has a range of dominant myxomycete species and near dominant species, apparently at random. That is, the highest ranking species in each Petri dish are thought to be determined by random events not by a few species having more efficient growth.
Twelve samples have what appears to be a ‘swarm’ of relatively large number of large fruiting body of one species, this ‘swarm’ being of high total volume (>16 mm3), and larger than a normal high volume total harvest. A log(abundance)-rank plot was made for samples containing these events (Fig. 6). For all samples there is a bigger than expected volume change between the highest volume species and the next, shown by the difference in slope of the thick lines and thin lines in Fig. 6. Hence this sample data is not consistent with the linear log(abundance)-rank model. The data is consistent with there being an abnormal transfer of myxomycete mass from the previous myxomycete feeding event on the tree bark to the event in the Petri dish for the species forming a swarm. The genera forming these ‘swarms’ are Physarum (seven samples), Badhamia (three samples), Stemonitis (one sample) and Enerthenema (one sample). The number of fruiting bodies forming a ‘swarm’ ranged from 300 to 1000.
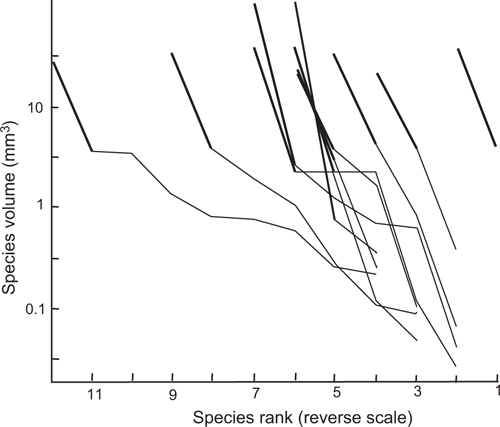
|
Modelling the decrease in myxomycete activity at the end of a growing event
In this survey the harvest was greatest at the first and second harvests, then decreased, and was negligible from harvest four onwards. Of interest is the shape of the rise and fall in fruiting body numbers, and its possible variation between species. Samples 1312 to 1391 were investigated. For each species of these samples the number of fruiting bodies was counted for each harvest, and if there were records for most weeks the sample-species could be used. Species were selected that had records that could be used derived from three or more samples. For each species the total number of fruiting bodies for the selected samples was added for each harvest. The results for each species were normalised to a value of 1.0 at harvest 2 (14 days). The graph of this data (Fig. 7) shows the mean paths of the various species are relatively consistent after harvest 2, with an approximately linear decrease from harvest 2 to harvest 4 and an average of very few fruiting bodies for each species at harvest 4 and later. The simplest explanation for this linear decrease is that the myxomycete numbers are limited by a linear decrease in food supply and that this food supply affects the species equally. Prior to week two the trajectories of the species are different, consistent with the exponential increase of each species starting at different times, and possibly increasing at a different rate. This overall model is roughly consistent with the published ‘development time’ information, such as by Schnittler (2001).
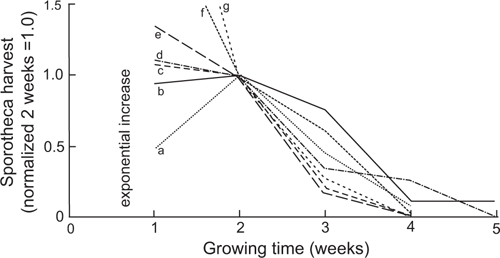
|
It is unusual for bark in the countryside to be dry for long periods then damp continuously for four weeks or longer. Hence moist chamber cultivation is unlikely to be replicate the pattern of myxomycete growth and decrease in activity during a rainfall event in the countryside. Figs 5 and 7 are important in that they are an indication of the controls on myxomycete growth and subsequent decrease in activity.
Biogeographical region boundaries
In order to put the transition zone widths for myxomycete species assemblages in perspective, it is necessary to compare them with transition zone widths for other Australian organisms. However no estimates of transition zone width and continuity have been published. Linder et al. (2012), in analysing geographical regionalisation, gave maps for Africa of a parameter ‘neighbourhood heterogeneity’. Bands of high heterogeneity correspond with the transition zones. The bands are generally 200–300 km wide, which is consistent with transition zone widths for Australian myxomycetes.
The magnitude of myxomycete change over a species assemblage boundary can be expressed by the proportion of species that terminate at the transition zone. Estimates for the Tr/NAr boundary are 80% in the Northern Territory and 80% in Queensland (Wellman 2016), for the NAr/SAr boundary it is 30% in the Northern Territory and ~43% in Western Australia, and at the SD/Te boundary it is 57% in north-east New South Wales, 53% in south-east New South Wales and ~64% in Western Australia. These proportions have low accuracy but they show there is generally considerable species change at species assemblage boundaries.
The restriction of certain myxomycete species to certain bark types, and the restriction of certain bark types to some phytogeographical regions makes it highly likely that at least in part myxomycete species associations are controlled by phytogeographical regions. However, the partial independence of the myxomycete species association is suggested by species that are characteristic of arid areas (Physarum decipiens, and Mabrideola oblonga).
Myxomycetes (a unicellular organism) appears to be geographically distributed in such a way that different species associations are found in distinct geographical regions. This leads to question as which plant and animal groups are also in similar (major) geographical regions, and how much the boundaries agree. The difference in flora and fauna of the arid and non-arid areas of Australia was recognised early (Tate 1889). It is a major feature of the phytogeographical and terrestrial zoogeographical regions (Kikkawa and Pearse 1969; Cogger and Heathwole 1981; Ebach et al. 2013, 2015; González-Orozco et al. 2014). There is approximate agreement between the various estimates of the arid/non-arid boundary position from plant, animal, myxomycete and arid ecology (Morton et al. 2011). A boundary between a northern and southern arid areas was recognised by Burbidge (1960). González-Orozco et al. (2014) showed that this boundary was the major floral boundary in Australia. Computer analysis showed that a similar boundary is a reasonably important boundary for birds (Kikkawa and Pearse 1969) and for reptiles (Cogger and Heathwole 1981). The present paper shows that there is a similar boundary with myxomycetes. Hence, there seems to be common first-order biogeographical regionalisation in Australia, and this includes myxomycetes (Fig. 2). In Europe and Africa computer analyses of flowering plant and animal distribution data have been used to determine if there is a common biogeographical region. Common regions was found in subSaharan Africa with mammal, bird, amphibian, reptile, and vascular plants (Linder et al. 2012), but common regions were not found in Europe (Rueda et al. 2010).
Similarity of the Australian species assemblages to those found in other continents
Three overseas corticolous surveys give species lists that contain a large proportion of the species found commonly in Australia. McHugh (2009) reported a small survey from two areas in Paraguay: an arid area near Filadelfia (mean annual rainfall 700 mm, 22°S), and from the higher rainfall area in eastern Paraguay (1200–1800 mm, ~26°S). A high proportion of the species in this survey are also found in the Australian arid and non-arid species associations. Novozhilov et al. (2003) reported on a survey in the Colorado Plateau of USA 30–38°S and a mean rainfall of 130–250 mm/year, but with a wide range in altitude and hence climate, and few tree species. These Colorado species are very similar to the Australian arid species (Wellman 2016). Pando and Lado (1990) and Wrigley de Basanta (1998) reported on surveys of Spain 36–44°S and a mean rainfall of 400–1000 mm/year. The reported species are similar to SAr and Te species assemblages in Australia (Wellman 2016). Wellman (2017) compared myxomyetes species assemblages field-collected on wood in temperate climates in Australia, Patagonia, Spain and Britain and found that there were no major differences in the species present or their relative abundance that could not be attributed to normal collecting variation. Hence the myxomycetes in Australia on bark and wood have a similar species assemblage to those elsewhere in areas with the same climate. The major differences between Australian corticolous myxomycetes and those overseas are that only in Australia is the common Comatricha species C. ellae and C. elegans, one common Australian species (Arcyria sp. H) has not been recognised elsewhere, and one rare species (Cribraria bicolor, Stephenson et al. 2018) has not been found outside Australia.
Other topics
Martiny et al. (2006) using data from molecular methods discussed controls on microorganism distribution. They concluded that the control is both the present environment and past events, but not random distribution over space. However, this is not supported by the present study. Control by present environment is supported by the existence of the species associations, and by the observation that almost all important species are found in similar environments overseas. The only significant species not found overseas is Arcyria sp. H. Hence, there is very little evidence for ‘past events’ controlling Australian corticolous myxomycete distribution. Random distribution is not dominant as the level of outliers in Tables S1–S4 is low. Hence this study is consistent with environmental control (the Baas-Becking model of ubiquity) being the overriding control of myxomycete distribution.
Myxomycetes may be a fairly obscure group of single cell organisms but models for their distribution and growth may be important in two contexts. First, single cell organisms constitute a major proportion of life on Earth. In a paper on the biomass distribution on Earth, Bar-On et al. (2018) estimated that of the total biomass of the Earth of 550 Gt of carbon, of which 450 Gt C was plants, 70 Gt C was bacteria, 7 Gt C was archaea, and 4 Gt C was protists. In the soil biomass myxomycetes form a major component of soil protozoans (Stephenson et al. 2011). Modelling of these other single cell organisms over the Earth is difficult except by molecular methods, while myxomycetes can be studied easily using normal botanical methods. Second, continental-scale distribution modelling of spore bearing plants/fungi (ferns, mosses, liverworts, lichen, fungi) is difficult in a mainly arid continent such as Australia. Distribution models have not been proposed for these groups. However the effect of spore distribution should be similar for myxomycetes and spore bearing plants.
There has been very little previous discussion of the relative abundance of myxomycete species within continental sized areas. It seems from this study that myxomycete distributions are an interaction between three distribution models; species distribution density having step-like changes, gradients in density, and random distribution of ‘outliers’. Step-like changes are supported by the species association model (Table 1). Ing (1982) made maps of the distribution over Britain, which are consistent with species having a limited tolerance to the Atlantic weather causing the distribution of species to terminate at different distances to the north-west. This gradient change is similar to the change across Tr and Te from the relatively arid short woodland to the wetter tall open forest and closed-canopy rainforest. These two observations support gradients in distribution density. The possibility of a low density of ‘outliers’ is consistent with the existence of scattered odd species observations within the long profiles of this paper (Tables S1–S3).
Using molecular techniques it has been found that marine amoeba-eating bacteria have a mean value of gross growth efficiency of 22% (Zubkov and Sleigh 1999) and 36% (Butler and Rogerson 1996). The total volume of myxomycete spores grown from five Petri dishes of each sample is ~4 mm3 over most of Australia (Fig. 3), so the total volume of bacteria (and other minor food groups) eaten can be estimated very approximately as 14 mm3 dry volume for the very artificial rainfall event of a moist culture experiment. The total mass of the bark sample in five Petri dishes is to 30 g, and the area of the weathered bark surface is generally ~70% of the areas of the dishes or 4500 mm2. It is thought that only the small thickness of the well weathered bark is hosting the myxomycetes and their food, so that area is probably the critical parameter to convert the harvest to a landscape value of corticolous myxomycete productivity.
Conclusions
The regional variation of myxomycetes across Australia can be determined by sampling long traverses across the continent along the rainfall gradient. For each substrate sample we can obtain relatively large populations of the stronger, common myxomycetes by collecting bark samples that are large and diverse in texture, and by cultivating more Petri dishes for each sample. With these changes in procedure a continent-wide ecological study of a unicellular organism (myxomycetes) is achievable over a relatively short period.
Along myxomycete long-traverses we find four distinct species associations (Tropical, Northern Arid, Southern Arid and Temperate). In the arid and semiarid areas the myxomycete species associations do not change much within the species association either north–south with amount of rainfall, or east–west with the proportion of rainfall falling in summer relative to winter. In the non-arid areas there is a strong change in species from the inland, dryer forests of short woodland to wetter forests (taller denser forest and closed rainforest), but this change is poorly defined with available data. The transition zones between these species associations have a width of 200–400 km, which is about one-half the width of many of the geographical regions, so between one-half and one-third of the Australian area is within a transition zone. Across a transition zone the species associations change by 30–80%. The four species assemblages can be distinguished by key common species, the species being previously known to occur in this climatic zone overseas. The regions defined by the four myxomycete species associations correspond with the major flowering-plant based phytogeographical regions. This is thought to be partly due by all bark types are not present in all the phytogeographical regions.
The average productivity of the myxomycete population, is relatively high and uniform over arid Australia, but it is lower in the open forest non-arid areas, and is likely to be much lower in wet tall open-forests and closed rainforests. The change in composition of the myxomycetes can be expressed using the proportion of the myxomycete orders. The Echinosteliales and Clastodermatales have about the same importance in arid and non-arid areas. The Physarales and Liceales are more important in the arid areas. The Trichiales, Cribraiales, and to a lesser extent Stemonitidales, are more important in the non-arid areas. When measuring importance by the volume of spores, the Physarales dominates the pattern of change, it is ~55% of the volume in the arid area, and ~20% in the non-arid areas. The myxomycete species clearly have preferred climatic area (species associations), bark preferences of pH, hardness and texture. The pH of the bark is important, effecting the number of records per sample, the total volume of spores produced, and the proportion of the Physarales.
For most biological groups the population dynamics during growth are determined by a single large population. With myxomycetes cultivation we have a large number of individual growth experiments, which together can be used to infer the average population dynamics after a watering event. Each growing phase can be modelled by a linear log(abundance)-rank plot. This model is consistent with the species growing exponentially in an initially vacant space. The population decline after this exponential growth is a linear decrease in harvested spore volume from the end of week two to negligible harvest at the end of week four. This model is consistent with a linear decrease in the amount of food available.
Conflict of interest
The author declares that there are no conflicts of interest.
Acknowledgements
I wish to thank the Australian National Herbarium for the use of laboratory facilities, and two anonymous reviewers for very helpful suggestions. I received help and encouragement from Christine Cargill, Judith Curnow and Heino Lepp of the Cryptogam Herbarium at the Australian National Herbarium, Steve Stephenson of University of Arkansas, and (during field work) from Mike Morriss of Canberra. The collections are in the Australian National Herbarium (CANB). The research did not receive any specific funding.
References
Bar-On YM, Philips R, Milo R (2018) The biomass distribution on Earth. Proceedings of the National Academy of Sciences of the United States of America 115, 6506–6511.| The biomass distribution on Earth.Crossref | GoogleScholarGoogle Scholar | 29784790PubMed |
Black DR, Stephenson SL, Pearce CA (2004) Myxomycetes associated with the aerial litter microhabitiat in tropical forests of northern Queensland, Australia. Systematics and Geography of Plants 74, 129–132.
BOM Bureau of Meteorology (2017) Average annual rainfall map for Australia. Product code IDCJCM004. Available at http://www.bom.gov.au/jsp/ncc/climate_averages/rainfall/index.jsp [Verified 22 January 2020].
Burbidge N (1960) The phytogeography of the Australian region. Australian Journal of Botany 8, 75–211.
| The phytogeography of the Australian region.Crossref | GoogleScholarGoogle Scholar |
Butler H, Rogerson A (1996) Growth potential, production efficiency and annual production of marine benthic naked amoebae (gymnamoebae) inhabiting sediments of the Clyde Sea area, Scotland. Aquatic Microbial Ecology 10, 123–129.
| Growth potential, production efficiency and annual production of marine benthic naked amoebae (gymnamoebae) inhabiting sediments of the Clyde Sea area, Scotland.Crossref | GoogleScholarGoogle Scholar |
Cogger HG, Heathwole H (1981) The Australian reptiles: origins, biogeography, distribution patterns and island evolution. In ‘Ecological biogeography of Australia’. (Ed. A Keast) pp. 1303–1373. (Dr W Junk: The Hague, The Netherlands)
Davison EM, Davison PJN, Brims MH (2008) Moist chamber and field collections of myxomycetes from the northern Simpson Desert, Australia. Australasian Mycologist 27, 129–135.
Davison EM, Davison PJN, Barrett MD, Barrett RL, McMullan-Fisher SJM (2017) Additions to the myxomycota of summer rainfall regions of tropical Australia. Nova Hedwigia 104, 47–64.
| Additions to the myxomycota of summer rainfall regions of tropical Australia.Crossref | GoogleScholarGoogle Scholar |
Discover Life (2018) Discoverlife. Available at https://www.discoverlife.org/ [Accessed January 2020].
Ebach MC, Gill AC, Kwan A, Ahyong ST, Murphy DJ, Cassis G (2013) Towards an Australian bioregionalisation atlas: a provisional area taxonomy of Australia’s biogeographical regions. Zootaxa 3619, 315–342.
| Towards an Australian bioregionalisation atlas: a provisional area taxonomy of Australia’s biogeographical regions.Crossref | GoogleScholarGoogle Scholar | 26131478PubMed |
Ebach MC, Murphy DJ, González-Orozco CE, Miller JT (2015) A revised area taxonomy of phytogeographical regions within the Australian bioregionalisation atlas. Phytotaxa 208, 261–277.
| A revised area taxonomy of phytogeographical regions within the Australian bioregionalisation atlas.Crossref | GoogleScholarGoogle Scholar |
González-Orozco CE, Ebach MC, Laffan S, Thornhill AH, Knerr NJ, Schmidt-Lebuhn AN, Cargill CC, Clements M, Nagalingum NS, Mishler BD, Miller JT (2014) Quantifying phytogeographical regions of Australia using geospatial turnover in species composition. PLoS One 9, e92558
| Quantifying phytogeographical regions of Australia using geospatial turnover in species composition.Crossref | GoogleScholarGoogle Scholar | 24658356PubMed |
Ing B (1982) ‘A provisional atlas of British myxomycetes.’ (Biological Records Centre, Institute of Terrestrial Ecology: Huntingdon, UK)
Ing B (1999) ‘The myxomycetes of Britain and Ireland.’ (The Richmond Publishing Co. Ltd: Slough, UK)
Ing B, Spooner BM (1994) Myxomycetes from the Kimberley region, Western Australia. Botanical Journal of the Linnean Society 116, 71–76.
| Myxomycetes from the Kimberley region, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Jordan CC, Brims MH, Speijers EJ, Davison EM (2006) Myxomycetes on the bark of Banksia attenuata and B. menziesii (Proteaceae). Australian Journal of Botany 54, 357–365.
| Myxomycetes on the bark of Banksia attenuata and B. menziesii (Proteaceae).Crossref | GoogleScholarGoogle Scholar |
Kikkawa J, Pearse K (1969) Geographical distribution of land birds in Australia – a numerical analysis. Australian Journal of Zoology 17, 821–840.
| Geographical distribution of land birds in Australia – a numerical analysis.Crossref | GoogleScholarGoogle Scholar |
Knight KJ, Brims MH (2010) Myxomycota census of Western Australia. Nuytsia 20, 283–307.
Linder HP, de Klerk HM, Born J, Burgess ND, Fjeldså J, Rahbek C (2012) The partitioning of Africa: statistically defined biogeographical regions in sub-Saharan Africa. Journal of Biogeography 39, 1189–1205.
| The partitioning of Africa: statistically defined biogeographical regions in sub-Saharan Africa.Crossref | GoogleScholarGoogle Scholar |
Lloyd S (2014) ‘Where the slime mould creeps, the fascinating world of myxomycetes.’ (Tympanocryptis Press: Birralee, Tas.)
Martiny JBH, Bohannan BJM, Brown JH, Colwell RK, Fuhrman JA, Green JL, Horner-Devine MC, Kane M, Krumins JA, Kuske CR, Morin PJ, Naeem S, Øvreås L, Reysenbach A-L, Smith VH, Staley JT (2006) Microbial biogeography: putting microorganisms on the map. Nature Reviews. Microbiology 4, 102–112.
| Microbial biogeography: putting microorganisms on the map.Crossref | GoogleScholarGoogle Scholar |
May RM (1976) Patterns in multispecies communities. In ‘Theoretical ecology: principals and applications’. (Ed. RM May), pp. 142–162. (Blackwell Scientific Publications: Oxford)
McHugh R (2009) Field and moist chamber collections of Paraguay myxomycetes. Karstenia 48, 49–56.
| Field and moist chamber collections of Paraguay myxomycetes.Crossref | GoogleScholarGoogle Scholar |
McHugh R, Stephenson SL, Mitchell DW, Brims MH (2003) New records of Australian myxomycota. New Zealand Journal of Botany 41, 487–500.
| New records of Australian myxomycota.Crossref | GoogleScholarGoogle Scholar |
McHugh R, Mitchell DW, Brims MH, Stephenson SL (2009) New additions to the myxomycota of Australia. Australasian Mycologist 28, 56–64.
Mitchell DW (1995) The myxomycota of Australia. Nova Hedwigia 60, 269–295.
Morton SR, Stafford Smith DM, Dickman CR, Dunkerley DL, Friedel MH, McAllister RRJ, Reid JRW, Roshier DA, Smith MA, Walsh FJ, Wardle GM, Watson IW, Westoby M (2011) A fresh framework for the ecology of arid Australia. Journal of Arid Environments 75, 313–329.
| A fresh framework for the ecology of arid Australia.Crossref | GoogleScholarGoogle Scholar |
Novozhilov YK, Mitchell DW, Schnittler M (2003) Myxomycete biodiversity of the Cororado Plateau. Mycological Progress 2, 243–258.
| Myxomycete biodiversity of the Cororado Plateau.Crossref | GoogleScholarGoogle Scholar |
Novozhilov YK, Zimlianskaia IV, Schnittler M, Stephenson SL (2006) Myxomycete diversity and ecology in the arid regions of the Lower Volga River Basin (Russia). Fungal Diversity 23, 193–241.
Novozhilov YK, Rollins AW, Schnittler M (2017) Ecology and distribution of myxomycetes. In ‘Myxomycetes: biology, systematics, biogeography and ecology’. (Eds SL Stephenson, C Rojas), pp. 253–297. (Academic Press: London)
Pando F, Lado C (1990) A survey of the corticolous myxomycetes in peninsular Spain and Balearic Islands. Nova Hedwigia 50, 127–137.
| A survey of the corticolous myxomycetes in peninsular Spain and Balearic Islands.Crossref | GoogleScholarGoogle Scholar |
Poulain M, Meyer M, Bozonnet J (2011) ‘Les myxomycÈtes.’ (Fédération mycologique et botanique Dauphiné-Savoie: Sevrier, 2 volumes)
Rosing WC, Mitchell DW, Stephenson SL (2007) Corticolous myxomycetes from Victoria. Australasian Mycologist 26, 9–15.
Rueda A, Rodríguez MÀ, Hawkins BA (2010) Towards a biogeographic regionalization of the European biota. Journal of Biogeography 37, 2067–2076.
| Towards a biogeographic regionalization of the European biota.Crossref | GoogleScholarGoogle Scholar |
Schnittler M (2001) Ecology of myxomycetes of a winter-cold desert in western Kazakhstan. Mycologia 93, 653–669.
| Ecology of myxomycetes of a winter-cold desert in western Kazakhstan.Crossref | GoogleScholarGoogle Scholar |
Schnittler M, Dagamac NHA, Novozhilov YK (2017) Biogeographical patterns in myxomycetes. In ‘Myxomycetes: biology, systematics, biogeography and ecology’. (Eds SL Stephenson, C Rojas) pp. 299–331 (Academic Press: London)
Stephenson SL, Rojas C (2017) ‘Myxomycetes: biology, systematics, biogeography and ecology.’ (Academic Press: London)
Stephenson SL, Shadwick JDL (2009) Nivicolous myxomycetes from alpine areas of south-eastern Australia. Australian Journal of Botany 57, 116–122.
| Nivicolous myxomycetes from alpine areas of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Stephenson SL, Stempen H (1994). ‘Myxomycetes, a handbook of slime moulds.’ (Timber Press: Portland, OR, USA)
Stephenson SL, Fiore-Donno AM, Schnittler M (2011) Myxomycetes in soil. Soil Biology & Biochemistry 43, 2237–2242.
| Myxomycetes in soil.Crossref | GoogleScholarGoogle Scholar |
Tate R (1889) On the influence of physiological changes in the distribution of life in Australia. Report of the first Meeting of the Australian Association for the Advancement of Science. pp. 312–326, (Australian Association for the Advancement of Science: Sydney, Australia).
Stephenson SL, Novozhilov YuK, Wellman P (2018) A new species of Cribraria (Myxomycetes) from Australia. Новости систематики низших растений (Novosti sistematiki nizshikh rastenii) 52, 379–385.
| A new species of Cribraria (Myxomycetes) from Australia.Crossref | GoogleScholarGoogle Scholar |
Wellman P (2015) Myxomycetes (slime moulds) of arid and semi-arid areas of northwest New South Wales, Australia. Cunninghamia 15, 153–162.
Wellman P (2016) Distinctive corticolous myxomycete assemblages of tropical, temperate and arid regions of Australia. Cunninghamia 16, 101–114.
Wellman P (2017) Temperate-climate myxomycetes in Australia and the Southern Hemisphere. Cunninghamia 17, 1–13.
Whitney KD (1980) The myxomycete genus Echinostelium. Mycologia 72, 950–987.
| The myxomycete genus Echinostelium.Crossref | GoogleScholarGoogle Scholar |
Wrigley de Basanta D (1998) Myxomycetes from the bark of the evergreen oak Quercus ilex. Anales del Jardin Botanico de Madrid 56, 3–14.
| Myxomycetes from the bark of the evergreen oak Quercus ilex.Crossref | GoogleScholarGoogle Scholar |
Wrigley de Basanta D, Estrada-Torres A (2017) Techniques for recording and isolating myxomycetes. In ‘Myxomycetes: biology, systematics, biogeography and ecology’. (Eds SL Stephenson, C Rojas) pp. 333–363. (Academic Press: London)
Wrigley de Basanta D, Stephenson SL, Lado C, Estrada-Torres A, Nieves-Rivera AM (2008) Lianas as a microhabitat for myxomycetes in tropical forests. Fungal Diversity 28, 109–125.
Zubkov MV, Sleigh MA (1999) Growth of amoebae and flagellates on bacteria deposited on filters. Microbial Ecology 37, 107–115.
| Growth of amoebae and flagellates on bacteria deposited on filters.Crossref | GoogleScholarGoogle Scholar | 9929399PubMed |