

Evaluating conservation strategies for the endangered daisy Schoenia filifolia subsp. subulifolia (Asteraceae): fitness consequences of genetic rescue and hybridisation with a widespread subspecies
Leonie Monks A B * , David Coates A , Shelley McArthur A and Rachel J. Standish B
A B * , David Coates A , Shelley McArthur A and Rachel J. Standish B
A Biodiversity Conservation Science, Department of Biodiversity, Conservation and Attractions, Locked Bag 104, Bentley Delivery Centre, Bentley, WA 6983, Australia.
B Harry Butler Institute, Murdoch University, 90 South Street, Murdoch, WA 6150, Australia.
Australian Journal of Botany 70(5) 344-357 https://doi.org/10.1071/BT22006
Submitted: 21 January 2022 Accepted: 4 July 2022 Published: 27 July 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: To establish translocated populations of threatened plants with the genetic resources to adapt to changing environmental conditions, the source of propagation material is an important consideration.
Aim: We investigated the fitness consequences of genetic rescue and admixture for the threatened annual daisy Schoenia filifolia subsp. subulifolia, and the common S. filifolia subsp. filifolia, to inform seed-sourcing strategies for translocations of the threatened subspecies.
Methods: We evaluated genetic diversity of two populations of S. filifolia subsp. subulifolia and four populations of S. filifolia subsp. filifolia by using microsatellite markers. We grew seedlings from each study population and cross-pollinated inflorescences within and among populations of the same subspecies, and between subspecies. We evaluated the fitness consequences of each cross by using seed set, seed weight and seed viability.
Key results: There was a lower genetic diversity in the small (<50 plants, Nar = 3.28, He = 0.42) compared to the large (>10 000 plants, Nar = 4.42, He = 0.51) population of S. filifolia subsp. subulifolia, although none of the measures was significantly different, and seed fitness was slightly, although not significantly, reduced in interpopulation crosses compared with the small population. Genetic diversity was similar between the threatened and widespread subspecies; however, the subspecies were genetically divergent (Fst = 0.242–0.294) and cross-pollination between subspecies produced negligible amounts of seeds (<3% seed set).
Conclusions: Although genetic rescue or admixture of S. filifolia subsp. subulifolia would not necessarily result in greatly increased levels of genetic diversity or seed fitness, we still consider it a potential option. Negligible seed set in crosses between subspecies indicates that deliberate hybridisation is not a possibility.
Implications: Studies of fitness consequences of admixture or genetic rescue are rare yet critical to assessing the benefits of different translocation strategies.
Keywords: admixture, Asteraceae, demographic rescue, environmental change, genetic diversity, genetic rescue, reintroduction, seed fitness, threatened plants, translocation.
Introduction
Many causes of plant population extinction operate at a large scale, making it challenging, and in some cases impossible, to effectively manage threatened plant species in situ. Increasingly, conservation initiatives such as translocation are needed to ensure the persistence of a population or species. Translocations are defined as the intentional transfer of plants or plant regenerative material to a new location and are used to augment numbers of individuals in small and declining populations, to reintroduce plants to extinct populations, and to introduce plants to new safe locations (Commander et al. 2018).
The aim of translocation is to establish self-sustaining populations with sufficient genetic diversity to enable adaptive evolutionary change (Guerrant 1996). The levels of genetic diversity in the founding plants will therefore affect the likelihood of adaptation and, so, the long-term persistence of the translocated population (Montalvo and Ellstrand 2001). However, sourcing propagation material to establish translocated populations with sufficient genetic diversity can often be challenging. Small, isolated populations of threatened plants may be affected by inbreeding, genetic drift and reduced geneflow, which can negatively influence the levels of genetic diversity and fitness of propagules (Broadhurst et al. 2008; Weeks et al. 2011; Frankham et al. 2017). Additionally, these small population sizes can limit the amount of plant material (i.e. seeds, root stock, cuttings) available for propagation (Guerrant 1996). Where new populations are to be established (introduction translocations), or populations re-established (reintroduction translocations), genetic diversity can potentially be maximised through the carefully considered admixture of several populations (e.g. van Rossum et al. 2020). For the augmentation of small, isolated, and inbred populations, genetic rescue, the introduction of new genetic material (Tallmon et al. 2004; Whiteley et al. 2015; Ralls et al. 2018), may benefit species persistence.
Admixture involves the relatively even mixing of plant material from isolated populations and establishment of these plants at a new location to facilitate hybridisation and increase genetic diversity (Shi et al. 2018). Selection pressure could be strong at the new location. In contrast, genetic rescue involves the addition of a small number of plants from other populations, into small populations that are inbred, have low or no geneflow from other populations, and is not attempted where selection pressure is strong (Tallmon et al. 2004). The addition of new genetic material into an inbred population for genetic rescue, and the hybridisation that occurs in admixture, can benefit threatened plants by increasing genetic diversity, population size, and offspring fitness (e.g. growth rate) through heterosis or adaptive evolution, thereby reducing extinction risk (Pickup et al. 2013; Whiteley et al. 2015; Shi et al. 2018). However, there are concerns that the use of genetic rescue for augmenting populations or admixture to establish new populations can have adverse consequences. Genetic rescue may lower the fitness of the population being augmented by swamping the gene pool of the locally adapted plants with hybrids between the local and introduced plants, which are less adapted to the local conditions (Hufford and Mazer 2003; Whiteley et al. 2015). Admixture or genetic rescue can result in a population with reduced fitness because of outbreeding depression when the new genetic material is introduced from a genetically differentiated, ecologically dissimilar or geographically distant population (Hufford and Mazer 2003; Edmands 2007; Shi et al. 2018). Additionally, there are doubts as to how long the fitness benefits of heterosis last, with maximum fitness benefits predicted for the F1 and F2 generations before a decline in benefits for subsequent generations (Bell et al. 2019). Other concerns are that environmental conditions and threatening processes could play a larger, more imminent role in population decline than does a lack of geneflow, and these may be overlooked in the rush to undertake a translocation (Bouzat et al. 2009; Whiteley et al. 2015; Bell et al. 2019). Instead, proponents argue that the simple addition of immigrants to a population (i.e. demographic rescue) may have the same effect in decreasing probability of population extinction (Hufbauer et al. 2015; Bell et al. 2019) regardless of their contribution to genetic diversity and the gene pool.
The magnitude of the change in the level of fitness following admixture or genetic rescue can be associated with geographic distance between populations (Waser and Price 1989). That is, fitness measures can be lower in crosses between plants from the same or nearby populations (inbreeding depression) and between widely geographically separated populations (outbreeding depression), than in crosses with plants from populations over an intermediate geographic distance (Waser and Price 1989; Hufford et al. 2012). Cross-pollination experiments among different populations have demonstrated this effect, with greater germination and survival of F1 seedlings of Stylidium hispidum from intermediate-distance crosses, than from within-population, and long-distance crosses (Hufford et al. 2012). Similarly, improved seed set and seedling survival of Jacquemontia reclinata were observed over increasing geographic crossing distance (Maschinski et al. 2013) and higher seed viability; faster seed germination and increased seedling recruitment were recorded for Zostera marina from an intermediate crossing distance than from the near and far crossing distances (Billingham et al. 2007). These findings suggest that there is an optimal intermediate outcrossing distance that avoids both inbreeding depression and outbreeding depression that can inform genetic rescue of small populations or admixture to establish translocated populations.
As well as the biological concerns about the mixing of gene pools, cultural and legislative concerns have contributed to minimising attempts at genetic rescue (Frankham 2015; Ralls et al. 2018) and, until recently, limited the use of admixture to establish or re-establish populations (Maschinski et al. 2012). To counteract some of these concerns, risk frameworks have been developed to guide decisions on when and how to use genetic rescue or admixture to minimise potential negative consequences (Byrne et al. 2011; Frankham et al. 2011; Weeks et al. 2011; Frankham 2015). These frameworks indicate that the risk of outbreeding depression is low if the taxonomy is adequately resolved, there are no fixed chromosomal differences between the populations, gene flow has occurred within the past 500 years, and the populations are adapted to similar environments (Frankham et al. 2011; Frankham 2015). Additionally, the fitness benefits of genetic rescue have been demonstrated to occur to the F3 generation and beyond in numerous plant species (Pickup et al. 2013; Frankham 2016), which has helped allay concerns.
Some threatened plant species may exist only as a single population, or genetically, geographically, or ecologically divergent populations may be the only source of propagation material (Hedrick and Garcia-Dorado 2016; Kronenberger et al. 2017). In these cases, there is a question as to whether positive benefits of admixture or genetic rescue from more genetically divergent populations, or even closely related taxa such as subspecies, offset the risks of outbreeding depression (Baskett and Gomulkiewicz 2011; Hedrick and Garcia-Dorado 2016; Kronenberger et al. 2017). Empirical testing of the benefits of admixture or genetic rescue between genetically divergent populations will assist in assessing the trade-off between risks and benefits, but such experiments should be undertaken ex situ to allow for contained assessment of the consequences of such actions (Harrisson et al. 2016).
Schoenia filifolia (Turcz.) Paul G.Wilson is an annual herb from the Asteraceae family that grows in patches across the south-west of Western Australia. The species consists of the following three subspecies: subsp. subulifolia, subsp. arenicola and subsp. filifolia. The first subspecies, S. filifolia subsp. subulifolia, has a conservation listing of endangered (Smith and Jones 2018). Translocations of this subspecies are an important component of its conservation. However, seed is available only from two extant populations, one of which is large (>10 000 individuals) and the other very small (<50) and declining. To assess the benefits and risks of admixture to augment the small population or translocation to establish a new population, we investigated interpopulation hybridisation in S. filifolia subsp. subulifolia and in the more common widespread S. filifolia subsp. filifolia, and crosses between the subspecies, to inform the development of a translocation strategy for S. filifolia subsp. subulifolia. We coupled this investigation with an assessment of patterns of genetic diversity across populations of subsp. subulifolia and subsp. filifolia. Specifically, we asked the following questions: (1) what are the levels of genetic diversity and genetic divergence between and among the study populations of both subspecies; (2) what are the consequence of cross-pollination among and within populations of each subspecies on the early fitness measures of seed set, seed weight and seed germinability; and (3) can cross-pollination between subspecies occur and what are the early fitness consequences? Answers to these questions will inform whether genetic rescue of the small population of subsp. subulifolia is likely to be beneficial, whether admixture to establish new populations or re-establish extinct populations of subsp. subulifolia will be an effective conservation strategy, and which source population or populations (intra- or interspecific) will maximise genetic diversity and therefore evolutionary potential, while minimising the negative consequences of outbreeding depression. We discuss the implication of our findings for conservation of other endangered annual herbs.
Materials and methods
The species
Schoenia filifolia, from the family Asteraceae, is an annual erect herb growing up to 0.5 m in height. Multiple flowers are held in a capitula surrounded by bright yellow involucre bracts (Wilson 1992). Flowers are hermaphroditic and flowering occurs between July and November. The species is distributed across the south-west of Western Australia (Fig. 1).

|
Three subspecies are described. The first, S. filifolia subsp. subulifolia is distinguished from the other two subspecies by having a larger capitula and a longer ray, and a hemispherical involucre. Flowers of subspecies subulifolia are self-incompatible and predominantly insect pollinated, although wind pollination may occur over short distances (Png 2012). The subspecies is currently known from three extant populations (although only two populations were known at the time this study was undertaken) in a small area around the inland town of Mingenew (https://florabase.dpaw.wa.gov.au/; Fig. 1; Table 1). Subspecies arenicola has not been seen since 1977 and, consequently, is not included in this study. Subspecies filifolia has a widespread distribution from Cue in the north, inland to Coolgardie and south to Kondinin (https://florabase.dpaw.wa.gov.au/; Fig. 1) and is not listed as a taxon of conservation concern (Table 1). Like pollination of subsp. subulifolia, that of subsp. filifolia is likely to be insect- or wind-mediated, although this has not been confirmed.

|
Sampling
Four populations of subsp. filifolia were chosen for sampling because they represented populations from across the range of the subspecies (Fig. 1). Two populations of subsp. subulifolia were chosen for sampling as they were the only two extant populations known when the study commenced (Fig. 1). The six populations were visited during flowering in September 2016. Up to 24 plants were sampled from each population (all above-ground parts) and stored as individuals in envelopes. Samples were freeze-dried and then stored on silica until required. A representative specimen was also collected from five of the sampled populations, identification was verified, and the specimens were lodged at the Western Australian Herbarium (PERTH 08938008, PERTH 08938016, PERTH 08938024, PERTH 08938032, PERTH 08937990). The Lockier population of subsp. subulifolia was previously verified (PERTH 07772041, https://florabase.dpaw.wa.gov.au/). In November 2016, seeds were collected from the four populations of subsp. filifolia from >25 plants across each population and then held at 15°C and 15% relative humidity until required. Seeds of two populations of subsp. subulifolia, already held at the Western Australian Seed Bank, Kensington, were used for this study. These seeds were collected in October 2007 (Lockier) and October 2015 (Mingenew) and stored at −18°C and 15% relative humidity.
Genetic structure and genetic diversity
DNA was extracted from freeze-dried leaf material (approximately 50 mg) following the Doyle (1991) CTAB extraction method, with the addition of polyvinylpyrrolidone (PVP-40) and sodium sulfite to the extraction buffer (Byrne et al. 2001) and a second chloroform extraction step. DNA from one individual was sent to the Monash University Malaysia genomics facility for dye-based sequencing on an Illumina MiSeq desktop sequencer (Illumina, San Diego, California, USA). Unique loci were identified, and primers designed for 138 potential loci by using the default parameters in QDD v3.1 (Meglécz et al. 2010).
Eight individuals were randomly selected to trial amplification and polymorphism of microsatellite regions. Forty-eight of the primer pairs were tested and of these three primers showed no amplification, and 32 primers showed no variation or did not consistently amplify across all populations. From 13 remaining primers, seven were chosen (SCF01, SCF04, SCF08, SCF16, SCF20, SCF23, SCF35, Supplementary Table S1), because they reliably amplified across all populations.
The G5 label set was used to label forward primers with a fluorescent tag (either VIC, PET, NED or FAM). Microsatellite regions were then amplified by polymerase chain reaction (PCR) in multiplexes of two or three sets, by using a Qiagen Multiplex kit (Qiagen, Hilden, Germany). Each multiplex reaction contained 1 μL of DNA, 3.75 μL of Qiagen mix, 0.75 μL primer mix and 2 μL water (from Qiagen kit). Primer mix contained VIC, PET, NED, or FAM. The PCR amplification program was as follows: 95°C for 15 min, followed by 35 cycles of 94°C for 30 s, 60°C for 90 s, 72°C for 60 s, and a final extension of 60°C for 30 min. Two or three randomly selected individuals were added twice to each plate to serve as plate-level negative controls. The PCR product was diluted with sterile deionised water at a rate of 1:20 and then 1 μL of diluted PCR product was added to 12 μL of Hi-Di-formamide/0.125 μL GeneScan LIZ 500 (−250) size Standard (Applied Biosystems, Waltham, MA, USA). Fragments were viewed with automated fluorescent scanning detection using an Applied Biosystems 3730 DNA Analyser. Genotypes were scored in GeneMapper V5 (Applied Biosystems). The allele bins were assigned manually, checked automatically and, where required, the alleles were manually adjusted.
GENEPOP ver. 4.0 (Raymond and Rousset 1995) was used to test for Hardy–Weinberg equilibrium (HWE) across the seven loci with exact tests by the Markov-chain method. Tests for linkage disequilibrium (LD) between loci were performed in GENEPOP ver. 4. MICRO-CHECKER (van Oosterhout et al. 2004), using a Bonferroni adjusted 95% confidence interval and 10 000 repetitions, was used to look for evidence for large allele drop-out, scoring error due to stutter and null alleles. We also estimated the frequency of null alleles within a population, considering the possibility of inbreeding using INEST (Chybicki and Burczyk 2009).
GenAlEx v6.5 (Peakall and Smouse 2006, 2012) was used to calculate the mean number of individuals genotyped, averaged over all loci (N), and the measures of microsatellite diversity, including expected (He) and observed (Ho) heterozygosity and the fixation index (Fis). Because of the differences in sample sizes among populations, HP-Rare (Kalinowski 2005) was used to calculate allelic richness (Nar). Analysis of genetic diversity measures (Nar He, Ho and Fis) compared between subspecies using a nested one-way ANOVA and among populations within each subspecies by using a one-way ANOVA in R v3.6.2. Differences between population pairs were assessed using a Tukey HSD test (using the ‘TukeyHSD’ function in R v3.6.2).
Pairwise genetic differentiation among populations was estimated using unbiased Fst, Dst and 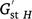 (Hedrick 2005; Meirmans and Hedrick 2011) in GenAlEx 6.5 based on the G-statistic option, with significance being tested using 999 permutations. Relationships among populations were analysed using principal coordinate analysis (PCoA) in GenAlEx based on a standardised covariance matrix using pairwise genetic-distance measures. Genetic structure was inferred using Bayesian model-based clustering in STRUCTURE v2.3.3, by using the admixture model and correlated allele frequencies (Pritchard et al. 2000). The analyses used Markov-chain Monte Carlo (MCMC) parameters with a burn-in period of 100 000 and 100 000 iterations, with 10 replicate runs for each value of K from 1 to 7. The most appropriate K value was found using STRUCTURE HARVESTER web v0.6.94 (Earl and vonHoldt 2012), as described by Evanno et al. (2005). The optimal alignment of 10 replicates at the optimal K was determined using CLUMPP 1.1.2 (Jakobsson and Rosenberg 2007), and a similarity coefficient, h′, was calculated to assess the similarity of individual replicates. A hierarchical approach was used to infer the number of clusters from the full dataset.
(Hedrick 2005; Meirmans and Hedrick 2011) in GenAlEx 6.5 based on the G-statistic option, with significance being tested using 999 permutations. Relationships among populations were analysed using principal coordinate analysis (PCoA) in GenAlEx based on a standardised covariance matrix using pairwise genetic-distance measures. Genetic structure was inferred using Bayesian model-based clustering in STRUCTURE v2.3.3, by using the admixture model and correlated allele frequencies (Pritchard et al. 2000). The analyses used Markov-chain Monte Carlo (MCMC) parameters with a burn-in period of 100 000 and 100 000 iterations, with 10 replicate runs for each value of K from 1 to 7. The most appropriate K value was found using STRUCTURE HARVESTER web v0.6.94 (Earl and vonHoldt 2012), as described by Evanno et al. (2005). The optimal alignment of 10 replicates at the optimal K was determined using CLUMPP 1.1.2 (Jakobsson and Rosenberg 2007), and a similarity coefficient, h′, was calculated to assess the similarity of individual replicates. A hierarchical approach was used to infer the number of clusters from the full dataset.
Glasshouse cross-pollination
In May 2017, seeds of subsp. subulifolia were removed from storage and equilibrated to room temperature over 24 h. To maximise germination, seeds of the six study populations were heat-shocked at 80°C for 14 days. Following heat-shock treatment, the seeds from each population were divided into three equal portions (replicates) and then plated onto individual petri dishes containing 7.5 g/L agar and incubated at 15°C. When the radicle had emerged at least 2 mm, the seed was scored as viable and was removed from the agar and transferred into soil mix in an individual pot (90 mm × 90 mm × 180 mm). Soil mix consisted of two parts composted pine bark, two parts course river sand and one part coco peat and was steam sterilised at 80°C for 2.5 h, then cooled prior to being used. Each potted seedling was labelled with source population information and an individual number and given 0.35 g of slow-release fertiliser. Seedlings were grown under nursery conditions until flowering (between 63 and 135 days).
When flowering commenced, plant height, widest crown width and crown width perpendicular to widest width, were measured for each plant. One inflorescence, where flowers had not yet opened, was selected on each plant, and covered with an organza mesh bag that excluded insect pollinators. Plants for crosses were chosen by selecting plants from the required cross type that were at the same stage of flowering. Crosses were reciprocal, with each plant donating and receiving pollen from another plant within the same cross type. Cross types and number of plants for each cross type are shown in Table S2. Flowering time overlapped between subsp. subulifolia and the Tardun, Morawa and Black Rd populations of subsp. filifolia. However, flowering time was much later in the Kondinin population of subsp. filifolia and did not overlap with flowering of subsp. subulifolia and, subsequently, we were unable to undertake any crosses between these populations. Flowering time of the Kondinin population, although overlapping with Tardun, Morawa and Black Rd populations, was delayed and resulted in fewer crosses between Kondinin and these populations.
Once flowering commenced, cross-pollination was undertaken every second day by gently rubbing the inflorescence of each plant against the selected cross. Flowerheads were re-bagged between cross-pollination events. Cross-pollinations occurred until 2 days after all flowers in the inflorescence had opened. Self-pollinations were undertaken on a subset of plants, using the same technique as for cross-pollination, but between inflorescences on the same plant. The plants were watered until seeds ripened. At seed release (inflorescences dried, and seeds loose within the bracts), the seeds of individual plants were collected and placed into a paper bag. Thereafter, all the above-ground shoot material was collected and placed into a paper bag.
Shoot material was oven-dried at 60°C for 48 h, then weighed. Inflorescences from cross-pollinations and self-pollinations were dissected under a microscope and seeds were sorted into whole seeds (firm, plump and dark brown seed) and non-viable seed (soft, shrivelled, and white/cream seed). Any flower parts and non-viable seed were weighed, and weight added to whole plant dry weight. Whole seeds were also weighed. The viability of whole seeds was assessed using a germination test, following the same technique as described above. After 22 weeks, the viability of seeds that remained ungerminated, but still whole and firm, was assessed using a stain. Seed were soaked in a 1% w/v solution of 2,3,5-triphenyl tetrazolium chloride incubated at 30°C, in the dark, for 48 h. Seed were cut to assess staining. Seed that had stained red were classed as viable, seed that did not stain (remained white), or where only a small proportion of the seed stained, were classed as non-viable.
For the plants grown for the study (P generation) percentage germination, plant height, mean crown width, crown volume (calculated as height × crown width × perpendicular crown width) plant dry weight and number of seeds per inflorescence were compared between subspecies by using a nested one-way ANOVA (using ‘aov’ function in R v3.6.2), where population was the nested factor. Percentage germination data were arcsin-transformed and crown volume was log-transformed prior to analysis. A one-way ANOVA was used to compare among populations within each subspecies for the same variables. For each analysis, when the P-value was significant (P < 0.05), mean differences between population pairs were assessed using a Tukey HSD test (using the ‘TukeyHSD’ function in R v3.6.2).
For the cross-pollinations (F1 generation), the percentage of whole seeds, weight of whole seeds, and percentage of viable seeds, were compared for the different cross-types. These variables were first compared for intrapopulation crosses within each subspecies; interpopulation crosses within subsp. filifolia, and between subspecies crosses, by using one-way ANOVA (using ‘aov’ function in R v3.6.2). Second, the variables were compared for each subspecies among the three cross-categories (all intrapopulation crosses, compared with all interpopulation crosses within each subspecies and compared with all between the subspecies crosses) by using nested one-way ANOVA with cross-type as the nested factor. In each case, percentage data were arcsin-transformed prior to analysis and when the P-value was significant (P < 0.05), mean differences between cross pairs were assessed using a Tukey HSD test (using the ‘TukeyHSD’ function in R v3.6.2).
For the self-pollinations, the percentage of whole seeds, weight of whole seeds and percentage of viable seeds were compared among the populations within each subspecies by using one-way ANOVA (using ‘aov’ function in R v3.6.2). These same measures were also compared between self-pollination crosses and intrapopulation crosses within subspecies, again by using one-way ANOVA (using ‘aov’ function in R v3.6.2). In each case, percentage data were arcsin-transformed prior to analysis.
Results
Genetic structure and genetic diversity
Sixty-one alleles were detected across seven loci. After adjusting for multiple comparisons (n = 33), significant departure from HWE (P < 0.05) in the form of heterozygote deficiency was observed at locus SCF01 in Tardun and Black Rd, locus SCF16 in Mingenew, Tardun and Morawa, locus SCF08 in Tardun and Morawa, and locus SCF20 in Morawa and Tardun. At no locus was there evidence of null alleles within any population and there was no evidence of large allele drop-out. When considering inbreeding using INEST, no locus had a frequency of null alleles of >0 across all populations. After adjusting for multiple comparisons (n = 21), no locus pairs were in linkage disequilibrium across any population.
There was significant differentiation among the S. filifolia populations, with high values for overall genetic fixation and allelic differentiation (Fst = 0.407 [P = 0.001]; Dst = 0.607; [P < 0.001], GstH = 0.759 [P < 0.001]). Between populations of the two subspecies, pairwise measures of Dst and GstH were high (Dst ≥ 0.691, GstH ≥ 0.762, Table 2). Within subspecies, pairwise measures of Dst and GstH were low for subsp. subulifolia (Dst = 0.023; GstH = 0.035, Table 2) and moderate for subsp. filifolia (Dst between 0.214 and 0.272; GstH between 0.274 and 0.340, Table 2). The PCoA clearly distinguished two discrete groups, with the two populations of subsp. subulifolia in one group, and the four populations of subsp. filifolia in the second group (Fig. 2). The first axis explained 44.8% of the variation in genetic composition and the second axis 15.3% of the variation (Fig. 2). STRUCTURE analysis confirmed the presence of two clearly differentiated groups, with the two populations of subsp. subulifolia occurring in one cluster and the four populations of subsp. filifolia occurring in the second cluster (Fig. 3). These results infer limited geneflow between the subspecies. These two clusters were supported by very high similarity among 10 replicates (h′ = 0.998).

|
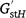 for two populations of Schoenia filifolia subsp. subulifolia and four populations of S. filifolia subsp. filifolia.
for two populations of Schoenia filifolia subsp. subulifolia and four populations of S. filifolia subsp. filifolia.

|

|
Comparisons of genetic diversity measures showed no differences between subspecies for Nar, Ho and He. However, Fis was significantly lower in subsp. subulifolia than in subsp. filifolia (P = 0.037; Table 3). Within subspecies, the two populations of subsp. subulifolia were not significantly different from each other for any genetic-diversity measure, although there was a trend towards lower genetic diversity in the small Lockier population. The four populations of subsp. filifolia were also not significantly different from each other for any genetic-diversity measure. There was no variation for the Kondinin population at any of the seven loci between any of the 24 samples, which may be due to apomixis, and, subsequently, this population was excluded from analysis for Ho, He and Fis.

|
Cross-pollination study
Parental generation
All fitness parameters, except height, were significantly different between subsp. subulifolia and subsp. filifolia (germination, P < 0.0001; crown P = 0.0018; log crown volume, P < 0.0001; dry weight, P = 0.0004; total number of seed per inflorescence, P < 0.0001; Table 4). Within subsp. subulifolia, plants from Mingenew were significantly larger than were plants from Lockier (height P = 0.0044, log crown volume P = 0.0139, dry weight P = 0.0015); however, germination was significantly lower (P = 0.00875; Table 4). All parameters within subsp. filifolia were significantly different among populations (initial viability P = 0.00598, height P < 0.0001, crown width P < 0.0001, log crown volume P < 0.0001, dry weight P < 0.0001), with plants from Tardun and Morawa generally taller and having more substantial crown volume than plants from Black Rd, and Black Rd plants larger than plants from Kondinin (Table 4). Germination was significantly lower at Kondinin than at Black Rd (P = 0.034) and Tardun (P = 0.007) and the number of seeds per inflorescence at Kondinin was significantly lower than that at Tardun (P < 0.0001), Morawa (P < 0.0001) and Black Rd (P < 0.0001).

|
Crosses
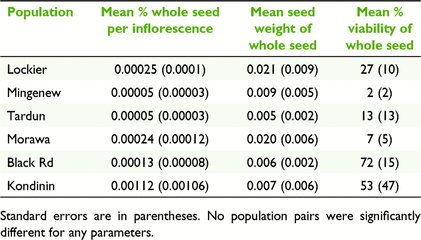
Seed set, seed weight and seed viability from self-pollination did not differ significantly among populations within the same subspecies. Inflorescences that were self-pollinated had less than 0.01% seed set in the six populations studied (Table 5) and this level of seed set was significantly less than that produced from intrapopulation crosses for subsp. subulifolia (P > 0.0001) and subsp. filifolia (P > 0.0001). Seed weight and seed viability of self-pollinated seed were also significantly less than those of intrapopulation crosses for subsp. subulifolia (P = 0.01; P > 0.0001) and subsp. filifolia (P = 0.0004; P = 0.04; Tables 5, 6).
|

|
Intrapopulation crosses of subsp. subulifolia resulted in greater seed set, seed weight and seed viability in the smaller Lockier population compared to the larger Mingenew population (P = 0.002; P = 0.02; P = 0.02; Table 6). Within subsp. filifolia, there were significant differences among populations for seed set (P < 0.0001) and seed weight (P = 0.002), but not seed viability, with Kondinin producing no viable seed from intrapopulation crosses, and Black Rd having significantly reduced seed set compared with Tardun (P > 0.0001) and Morawa (P = 0.002), and significantly reduced seed weight compared with Tardun (P = 0.002; Table 6).
Seed set from the subsp. subulifolia interpopulation cross was 13.6% (Table 6). The six subsp. filifolia interpopulation crosses resulted in seed set ranging from 0 to 29.1%, with a significant difference between the combination of populations crossed (P < 0.0001, Table 6). Many of the differences lay with crosses involving the Kondinin population, with no whole seed produced between this population when crossed with Morawa, and only very small amounts when crossed with Tardun (5.7%) and Black Rd (0.4%) (Table 6). However, there were no significant differences between the various subsp. filifolia interpopulation cross combinations for seed weight or seed viability.
Between subspecies, crosses resulted in a very low seed set, ranging from 0.6% to 3% of flowers per inflorescence producing whole seed. Of the seed set from the between-subspecies crosses, viability ranged between 39.2% and 97.8% (Table 6). There was no difference in seed set, seed weight or seed viability between the different subspecies interpopulation crosses (Table 6).
Comparisons between the different cross-types showed no difference between intrapopulation and interpopulation within-subspecies crosses for any of the seed fitness measures for both subspecies. However, both seed set and seed weight were significantly different for subsp. subulifolia when intrapopulation and interpopulation within-subspecies crosses were compared with between-subspecies crosses (P < 0.0001, P < 0.0001, P < 0.0001, P = 0.03; Table 6). Seed set, in particular, was substantially higher in intrapopulation (mean of 14.8%) and interpopulation within-subspecies (13.6%) compared with between-subspecies crosses (mean of 1.7%) (Table 6). Interestingly, there was no difference for seed viability between intrapopulation (80.6%) and interpopulation within-subspecies (74.1%) crosses compared with between-subspecies crosses (69.9%) in subsp. subulifolia (Table 6), so although there was only a very small number of seeds set for between-subspecies crosses, levels of viability in the small number of seeds was like that of the other cross-types. Differences were also seen in subsp. filifolia when intrapopulation and interpopulation within-subspecies crosses were compared with between-subspecies crosses, and these differences were not only for seed set (P < 0.0001, P < 0.0001) and seed weight (P = 0.0002, P = 0.0058), but also seed viability (P = 0.03, P = 0.006; Table 6). Seed set was substantially reduced for between-subspecies crosses (mean of 1.7%) compared with intrapopulation (mean of 18%) and interpopulation within-subspecies crosses (mean of 14.9%; Table 6). However, between-subspecies crosses (69.9%) resulted in higher seed viability than did intrapopulation (52.6%) and interpopulation within-subspecies (48.9%) crosses for subsp. filifolia (Table 6), which contrasts with findings for subsp. subulifolia.
Discussion
Inbreeding, genetic drift and reduced geneflow can adversely affect the levels of genetic diversity and fitness of propagules in small, isolated populations of threatened plants (Broadhurst et al. 2008; Frankham 2012; Leffler et al. 2012; Frankham et al. 2017). Our investigation can be used to assess options for managing genetic diversity in the endangered S. filifolia subsp. subulifolia. These options include augmentation of the small population with different genetic material from the large population, admixture of the two known populations to establish a new population, and hybridisation between this subspecies and its sister taxon S. filifolia subsp. filifolia, for augmentation and/or establishing a new population. In contrast to expectations, we found that the small population of the subsp. subulifolia had similar levels of genetic diversity as, and increased seed-fitness measures compared with, the large population of the subspecies, although there was a trend towards a lower genetic diversity in the smaller population. Furthermore, levels of genetic diversity of both populations of subsp. subulifolia were comparable to that of the more widespread sister taxon, subsp. filifolia, and both subspecies showed high levels of outcrossing, with limited seed production from selfing. We also found that interpopulation cross-pollination within subsp. subulifolia, an option that could be used to augment the small population or establish a new admixed population, resulted in increased seed fitness only relative to the large population, and that between-subspecies cross-pollination resulted in minimal seed set and substantially reduced seed weight. Our findings suggest, first, that genetic rescue of the very small population of subsp. subulifolia or admixture to establish a new translocation would not necessarily result in large increases in the levels of genetic diversity, but is still a viable management option, and, second, that deliberate hybridisation between the two subspecies to increase genetic diversity is not an option, given the negligible number of seeds produced. Instead, demographic rescue of the small population (Lockier) of subsp. subulifolia, sourcing individuals from the large population (Mingenew), is likely to be the best management option to reduce extinction risk of the small Lockier population, given its limited seed availability. Admixture to establish a new population of subsp. subulifolia is also a practical management option to reduce extinction risk across the subspecies.
Admixture of several source populations to establish a translocated population or augmentation of a small population with propagules from a large genetically diverse population can increase genetic diversity and enhance establishment success and persistence of the new or augmented population (Weeks et al. 2011; Whiteley et al. 2015; Shi et al. 2018). For S. filifolia subsp. subulifolia, our data suggested that the case for admixture is not clear-cut. We found that in subsp. subulifolia, genetic diversity was not significantly reduced in the small declining Lockier population compared with the substantially larger population, although there was a trend towards lower levels of genetic diversity in this small population. There was very low genetic differentiation between the two populations, and seed-fitness measures were significantly higher in the smaller population than in the large population. However, fitness of seeds resulting from crosses between these two populations was not significantly reduced relative to intrapopulation crosses for the small population, although there was a trend towards lower seed-fitness measures in the interpopulation crosses. Although this does suggest that there is no major fitness advantage in crossing these populations, there is the possibility of a slight increase in genetic diversity. This possibility, combined with the practicality of sourcing a greater number of seed to establish a translocation, makes genetic rescue of the small population and admixture to establish new populations, viable management options.
Our findings indicated that inbreeding depression and reduced genetic diversity are not significantly reduced in the small population of subsp. subulifolia compared with the larger populations of this and the other subspecies, and this could be due to the Lockier population once being much larger. Several dormancy mechanisms operate on the seed in this subspecies (Plummer et al. 2001), which means that the soil seed bank can persist for some time and could harbour genetic diversity that has not yet been lost from the population. Nevertheless, it is evident that there is a trend towards lower genetic diversity in the Lockier population and there could be significant negative demographic effects associated with its very small (<50 plants) population size and ongoing decline. Whereas reproductive output (i.e. seed set) and germination rate were higher than those of the larger population, other demographic processes such as recruitment failure or mass mortality, may be a greater driver of extinction risk of small populations than are genetic factors (Lande 1988). Indeed, the actual minimum viable population size of the subspecies is unknown but is likely to be more than 50 plants. Therefore, options for demographic rescue to significantly increase population size are still a high priority. This could be achieved by augmentation of the population with ex situ-stored seed or ex situ-grown seedlings from the small population in the first instance, or new material from the large population in the second instance. In addition, the establishment of a new population through admixture is a practical and viable strategy. Although this may not result in substantially higher levels of genetic diversity in the translocated population, maintaining larger numbers of plants and more populations will contribute to a reduction in extinction risk (Maschinski and Albrecht 2017).
Although translocation is not currently needed to conserve S. filifolia subsp. filifolia, information from this study could be of benefit when planning translocations of the closely related subsp. subulifolia. Where admixture to establish a new translocated population is considered as a way of increasing genetic diversity, knowledge of the patterns and levels of genetic differentiation can be used to maximise genetic diversity within translocations or augmented populations and minimise potential of outbreeding depression (Weeks et al. 2011; Maschinski et al. 2013). Our study suggests that admixture of the Tardun, Morawa and Black Rd populations is likely to result in a population with an increased genetic diversity relative to the three source populations, with minimal risk of outbreeding depression because of the low level of genetic differentiation, and with generally equivalent seed-fitness measures to those resulting from intrapopulation cross-pollination. However, our findings suggested that Kondinin be excluded from this admixed population. Although possessing several alleles that could enhance genetic diversity in an admixed population, there was a near-complete lack of seed set from crosses involving this population, much variation in habitat and morphology of plants from this population compared with the other three populations, and all adult individuals had the same multilocus genotype. The latter result may be due to apomixis, a common phenomenon in Asteraceae (Bicknell and Koltunow 2004). However, if it is a new apomictic taxon, further investigation is needed.
Another approach that can help guide seed-sourcing decisions for admixture and translocation planning is to investigate an optimal outcrossing distance to avoid both inbreeding and outbreeding depression (Waser and Price 1989; Hufford et al. 2012). We suggest that knowledge of optimal outcrossing distance would benefit translocation planning, both in the seed sourcing strategy, and locating translocation sites to optimise gene flow with wild populations. However, in practice, we found no clear pattern of increased seed fitness over an intermediate geographic distance compared with short or wide geographic distances for the subsp. filifolia crosses. Although the small number of populations used is a potentially limiting factor, other studies with relatively few populations have shown clear indications of an optimal distance over a range of geographic distances where outcrossing results in increased progeny fitness (Waser and Price 1989; Billingham et al. 2007; Hufford et al. 2012; Ayre et al. 2019). The same rule does not appear to apply to subsp. filifolia and this information can be used to guide translocation planning for the threatened subsp. subulifolia.
A key question was whether hybridisation between the subspecies could occur, because it offers potential to increase the genetic diversity in translocated populations of S. filifolia subsp. subulifolia. Inter-taxon hybridisation is rarely performed intentionally because of the perceived risk of outbreeding depression (Edmands 2007; Baskett and Gomulkiewicz 2011; Hedrick and Garcia-Dorado 2016; Kronenberger et al. 2017) often highlighted in risk frameworks (Byrne et al. 2011; Frankham et al. 2011; Weeks et al. 2011; Frankham 2015). However, in rare cases, inter-taxon hybridisation has been used to increase genetic diversity in threatened animal and plant species (Johnson et al. 2010; Valdiani et al. 2012; Zecherle et al. 2021). Further, hybridisation among different taxa occurs widely in Asteraceae (Whitney et al. 2010), and so there may be options to increase genetic diversity in the threatened subsp. subulifolia through hybridisation with the sister taxon subsp. filifolia. Additionally, with only two populations of subsp. subulifolia being known at the time of this study, seed was considered a significant limiting factor for translocation, and so hybridisation between the subspecies could have been an avenue to increase seed availability. Unfortunately, artificial cross-pollination between the two subspecies resulted in minimal levels of seed set and, although this seed was viable, such a low rate of seed production would not be a practical way of helping establish a new viable population.
Previous observations by Wilson (1992) suggested that taxon boundaries within S. filifolia require further investigation and we also note that the very low level of seed production from crosses between these two subspecies, compared with within the subspecies, seems indicative of reproductive barriers that might be expected between different species. This highlights the importance of adequate taxonomic resolution when considering mixing divergent populations for augmenting or establishing translocated populations of threatened plants. As the results of our study clearly indicated two distinct taxa, consideration should be given to further taxonomic revision of the two subspecies, S. filifolia subsp. subulifolia and S. filifolia subsp. filifolia, possibly elevating both to species level, taking into consideration the high level of genetic differentiation between the two subspecies, and the substantially reduced seed set when these subspecies are crossed. S. filifolia subsp. filifolia also warrants further taxonomic evaluation because there is variation in morphology and some evidence of reproductive isolation between Kondinin and the other three study populations.
The results of our study can inform translocations of other herbaceous or annual species from Asteraceae. Although other studies of herbaceous Asteraceae species have shown increased fitness following inter-population hybridisation, (DeMauro 1993; Pickup et al. 2013; Gavin-Smyth et al. 2021), our results for S. filifolia subsp. subulifolia showed that this is not the case and that genetic rescue of the small population (Lockier) or admixture to establish a new translocation is unlikely to result in substantially increased levels of genetic diversity or increased seed fitness. Our study highlighted that research into the levels of genetic diversity and genetic divergence among populations will be a valuable tool to support seed-sourcing decisions (Hufford and Mazer 2003), particularly before undertaking translocation actions such as admixture and genetic rescue. Where consideration is being given for sourcing new genetic material for translocations that maybe more distantly related (even between subspecies), these genetic studies will provide essential information, as will ex situ cross-pollination studies, to reduce the potential for outbreeding depression. In our case, genetic data coupled with cross-pollination studies suggested that demographic, rather than genetic, rescue is the best management option for S. filifolia subsp. subulifolia.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Funding for this study was received from the Australian government’s National Environmental Science Program Threatened Species Recovery Hub and Murdoch University.
Acknowledgements
We thank Alanna Chant for assistance with leaf and seed collection, Mike Hislop for specimen identification, Andrew Crawford for seed collection of S. filifolia subsp. subulifolia and assistance with seed germination and seedling propagation, the property owners for permission to sample from the S. filifolia subsp. subulifolia populations, Margaret Byrne for editing the manuscript and Matthew Williams for statistical advice.
References
Ayre BM, Roberts DG, Phillips RD, Hopper SD, Krauss SL (2019) Near-neighbour optimal outcrossing in the bird-pollinated Anigozanthos manglesii. Annals of Botany 124, 423–436.| Near-neighbour optimal outcrossing in the bird-pollinated Anigozanthos manglesii.Crossref | GoogleScholarGoogle Scholar |
Baskett ML, Gomulkiewicz R (2011) Introgressive hybridization as a mechanism for species rescue. Theoretical Ecology 4, 223–239.
| Introgressive hybridization as a mechanism for species rescue.Crossref | GoogleScholarGoogle Scholar |
Bell DA, Robinson ZL, Funk WC, Fitzpatrick SW, Allendorf FW, Tallmon DA, Whiteley AR (2019) The exciting potential and remaining uncertainties of genetic rescue. Trends in Ecology & Evolution 34, 1070–1079.
| The exciting potential and remaining uncertainties of genetic rescue.Crossref | GoogleScholarGoogle Scholar |
Bicknell RA, Koltunow AM (2004) Understanding apomixis: recent advances and remaining conundrums. The Plant Cell 16, S228–S245.
| Understanding apomixis: recent advances and remaining conundrums.Crossref | GoogleScholarGoogle Scholar |
Billingham MR, Simões T, Reusch TBH, Ester A, Serrão EA (2007) Genetic sub-structure and intermediate optimal outcrossing distance in the marine angiosperm Zostera marina. Marine Biology 152, 793–801.
| Genetic sub-structure and intermediate optimal outcrossing distance in the marine angiosperm Zostera marina.Crossref | GoogleScholarGoogle Scholar |
Bouzat JL, Johnson JA, Toepfer JE, Simpson SA, Esker TL, Westemeier RL (2009) Beyond the beneficial effects of translocations as an effective tool for the genetic restoration of isolated populations. Conservation Genetics 10, 191–201.
| Beyond the beneficial effects of translocations as an effective tool for the genetic restoration of isolated populations.Crossref | GoogleScholarGoogle Scholar |
Broadhurst LM, Lowe A, Coates DJ, Cunningham SA, McDonald M, Vesk PA, Yates C (2008) Seed supply for broadscale restoration: maximizing evolutionary potential. Evolutionary Applications 1, 587–597.
| Seed supply for broadscale restoration: maximizing evolutionary potential.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Macdonald B, Francki M (2001) Incorporation of sodium sulfite into extraction protocol minimizes degradation of Acacia DNA. BioTechniques 30, 742–748.
| Incorporation of sodium sulfite into extraction protocol minimizes degradation of Acacia DNA.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Stone L, Millar MA (2011) Assessing genetic risk in revegetation. Journal of Applied Ecology 48, 1365–1373.
| Assessing genetic risk in revegetation.Crossref | GoogleScholarGoogle Scholar |
Chybicki IJ, Burczyk J (2009) Simultaneous estimation of null alleles and inbreeding coefficients. Journal of Heredity 100, 106–113.
| Simultaneous estimation of null alleles and inbreeding coefficients.Crossref | GoogleScholarGoogle Scholar |
Commander LE, Coates DJ, Broadhurst L, Offord CA, Makinson RO, Matthes M (2018) ‘Guidelines for the translocation of threatened plants in Australia.’ 3rd edn. (Australian Network for Plant Conservation: Canberra, ACT, Australia)
DeMauro MM (1993) Relationship of breeding system to rarity in the Lakeside Daisy (Hymenoxys acaulis var. glabra). Conservation Biology 7, 542–550.
| Relationship of breeding system to rarity in the Lakeside Daisy (Hymenoxys acaulis var. glabra).Crossref | GoogleScholarGoogle Scholar |
Doyle J (1991) DNA protocols for plants. In ‘Molecular techniques in taxonomy. Vol. 57’. NATO ASI Series H: cell biology. (Eds GM Hewitt, AW Johnston, JPW Young) pp. 283–293. (Springer-Verlag: Berlin, Germany)
Earl DA, vonHoldt BM (2012) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conservation Genetics Resources 4, 359–361.
| STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method.Crossref | GoogleScholarGoogle Scholar |
Edmands S (2007) Between a rock and a hard place: evaluating the relative risks of inbreeding and outbreeding for conservation and management. Molecular Ecology 16, 463–475.
| Between a rock and a hard place: evaluating the relative risks of inbreeding and outbreeding for conservation and management.Crossref | GoogleScholarGoogle Scholar |
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software structure: a simulation study. Molecular Ecology 14, 2611–2620.
| Detecting the number of clusters of individuals using the software structure: a simulation study.Crossref | GoogleScholarGoogle Scholar |
Frankham R (2012) How closely does genetic diversity in finite populations conform to predictions of neutral theory? Large deficits in regions of low recombination. Heredity 108, 167–178.
| How closely does genetic diversity in finite populations conform to predictions of neutral theory? Large deficits in regions of low recombination.Crossref | GoogleScholarGoogle Scholar |
Frankham R (2015) Genetic rescue of small inbred populations: meta-analysis reveals large and consistent benefits of gene flow. Molecular Ecology 24, 2610–2618.
| Genetic rescue of small inbred populations: meta-analysis reveals large and consistent benefits of gene flow.Crossref | GoogleScholarGoogle Scholar |
Frankham R (2016) Genetic rescue benefits persist to at least the F3 generation, based on a meta-analysis. Biological Conservation 195, 33–36.
| Genetic rescue benefits persist to at least the F3 generation, based on a meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Frankham R, Ballou JD, Eldridge MDB, Lacy RC, Ralls K, Dudash MR, Fenster CB (2011) Predicting the probability of outbreeding depression. Conservation Biology 25, 465–475.
| Predicting the probability of outbreeding depression.Crossref | GoogleScholarGoogle Scholar |
Frankham R, Ballou JD, Ralls K, Eldridge M, Dudash MR, Fenster CB, Lacy RC, Sunnucks P (2017) ‘Genetic management of fragmented animal and plant populations.’ (Oxford University Press: Oxford, UK)
Gavin-Smyth N, Kramer AT, Urbina-Casanova R, Vitt P, Fant JB (2021) Genetic rescue reduces mate limitation in a threatened, clonal, and self-incompatible plant species. Restoration Ecology 29, e13458
| Genetic rescue reduces mate limitation in a threatened, clonal, and self-incompatible plant species.Crossref | GoogleScholarGoogle Scholar |
Guerrant EO (1996) Designing populations: demographic, genetic, and horticultural dimensions. In ‘Restoring diversity: strategies for reintroduction of endangered plants’. (Eds DA Falk, CI Millar, M Olwell) pp. 127–155. (Island Press: Covelo, CA, USA)
Harrisson KA, Pavlova A, Gonc A, da Silva A, Rose R, Bull JK, Lancaster ML, Murray N, Quin B, Menkhorst P, Magrath MJL, Sunnucks P (2016) Scope for genetic rescue of an endangered subspecies though re-establishing natural gene flow with another subspecies. Molecular Ecology 25, 1242–1258.
| Scope for genetic rescue of an endangered subspecies though re-establishing natural gene flow with another subspecies.Crossref | GoogleScholarGoogle Scholar |
Hedrick PW (2005) A standardized genetic differentiation measure. Evolution 59, 1633–1638.
| A standardized genetic differentiation measure.Crossref | GoogleScholarGoogle Scholar |
Hedrick PW, Garcia-Dorado A (2016) Understanding inbreeding depression, purging, and genetic rescue. Trends in Ecology & Evolution 31, 940–952.
| Understanding inbreeding depression, purging, and genetic rescue.Crossref | GoogleScholarGoogle Scholar |
Hufbauer RA, Szucs M, Kasyon E, Youngberg C, Koontz MJ, Richards C, Tuff T, Melbourne BA (2015) Three types of rescue can avert extinction in a changing environment. Proceedings of the National Academy of Sciences 112, 10557–10562.
| Three types of rescue can avert extinction in a changing environment.Crossref | GoogleScholarGoogle Scholar |
Hufford KM, Mazer SJ (2003) Plant ecotypes: genetic differentiation in the age of ecological restoration. Trends in Ecology & Evolution 18, 147–155.
| Plant ecotypes: genetic differentiation in the age of ecological restoration.Crossref | GoogleScholarGoogle Scholar |
Hufford KM, Krauss SL, Veneklaas EJ (2012) Inbreeding and outbreeding depression in Stylidium hispidum: implications for mixing seed sources for ecological restoration. Ecology and Evolution 2, 2262–2273.
| Inbreeding and outbreeding depression in Stylidium hispidum: implications for mixing seed sources for ecological restoration.Crossref | GoogleScholarGoogle Scholar |
Jakobsson M, Rosenberg NA (2007) CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 23, 1801–1806.
| CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure.Crossref | GoogleScholarGoogle Scholar |
Johnson WE, Onorato DP, Roelke ME, Land ED, Cunningham M, Belden RC, McBride R, Jansen D, Lotz M, Shindle D, Howard J, Wildt DE, Penfold LM, Hostetler JA, Oli MK, O’Brien SJ (2010) Genetic restoration of the Florida panther. Science 329, 1641–1645.
| Genetic restoration of the Florida panther.Crossref | GoogleScholarGoogle Scholar |
Kalinowski ST (2005) HP-RARE 1.0: a computer program for performing rarefaction on measures of allelic richness. Molecular Ecology Notes 5, 187–189.
| HP-RARE 1.0: a computer program for performing rarefaction on measures of allelic richness.Crossref | GoogleScholarGoogle Scholar |
Kronenberger JA, Funk WC, Smith JW, Fitzpatrick SW, Angeloni LM, Broder ED, Ruell EW (2017) Testing the demographic effects of divergent immigrants on small populations of Trinidadian guppies. Animal Conservation 20, 3–11.
| Testing the demographic effects of divergent immigrants on small populations of Trinidadian guppies.Crossref | GoogleScholarGoogle Scholar |
Lande R (1988) Genetics and demography in biological conservation. Science 241, 1455–1460.
| Genetics and demography in biological conservation.Crossref | GoogleScholarGoogle Scholar |
Leffler EM, Bullaughey K, Matute DR, Meyer WK, Ségurel L, Venkat A, Andolfatto P, Przeworski M (2012) Revisiting an old riddle: what determines genetic diversity levels within species? PLoS Biology 10, e1001388
| Revisiting an old riddle: what determines genetic diversity levels within species?Crossref | GoogleScholarGoogle Scholar |
Maschinski J, Albrecht MA (2017) Center for plant conservation’s best practice guidelines for the reintroduction of rare plants. Plant Diversity 39, 390–395.
| Center for plant conservation’s best practice guidelines for the reintroduction of rare plants.Crossref | GoogleScholarGoogle Scholar |
Maschinski J, Albrecht MA, Monks L, Haskins KE (2012) Centre for Plant Conservation best reintroduction practice guidelines. In ‘Plant reintroduction in a changing climate: promises and perils’. (Eds J Maschinski, KE Haskins) pp. 277–306. (Island Press: Washington, DC, USA)
Maschinski J, Wright SJ, Koptur S, Pinto-Torres EC (2013) When is local the best paradigm? Breeding history influences conservation reintroduction survival and population trajectories in times of extreme climate events. Biological Conservation 159, 277–284.
| When is local the best paradigm? Breeding history influences conservation reintroduction survival and population trajectories in times of extreme climate events.Crossref | GoogleScholarGoogle Scholar |
Meglécz E, Costedoat C, Dubut V, Gilles A, Malausa T, Pech N, Martin J-F (2010) QDD: a user-friendly program to select microsatellite markers and design primers from large sequencing projects. Bioinformatics 26, 403–404.
| QDD: a user-friendly program to select microsatellite markers and design primers from large sequencing projects.Crossref | GoogleScholarGoogle Scholar |
Meirmans PG, Hedrick PW (2011) Assessing population structure: FST and related measures. Molecular Ecology Resources 11, 5–18.
| Assessing population structure: FST and related measures.Crossref | GoogleScholarGoogle Scholar |
Montalvo AM, Ellstrand NC (2001) Nonlocal transplantation and outbreeding depression in the subshrub Lotus scoparius (Fabaceae). American Journal of Botany 88, 258–269.
| Nonlocal transplantation and outbreeding depression in the subshrub Lotus scoparius (Fabaceae).Crossref | GoogleScholarGoogle Scholar |
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6, 288–295.
| GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research.Crossref | GoogleScholarGoogle Scholar |
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research: an update. Bioinformatics 28, 2537–2539.
| GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research: an update.Crossref | GoogleScholarGoogle Scholar |
Pickup M, Field DL, Rowell DM, Young AG (2013) Source population characteristics affect heterosis following genetic rescue of fragmented plant populations. Proceedings of the Royal Society B: Biological Sciences 280, 20122058
| Source population characteristics affect heterosis following genetic rescue of fragmented plant populations.Crossref | GoogleScholarGoogle Scholar |
Plummer JA, Rogers AD, Turner DW, Bell DT (2001) Light, nitrogenous compounds, smoke and GA3 break dormancy and enhance germination in the Australian everlasting daisy Shoenia filifolia subsp. subulifolia. Seed Science and Technology 29, 321–330.
Png GK (2012) Pollination biology of Western Australia’s showy everlasting daisy (Schoenia filifolia subsp. subulifolia). Australasian Plant Conservation 21, 23–24.
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
| Inference of population structure using multilocus genotype data.Crossref | GoogleScholarGoogle Scholar |
Ralls K, Ballou JD, Dudash MR, Eldridge MDB, Fenster CB, Lacy RC, Sunnucks P, Frankham R (2018) Call for a paradigm shift in the genetic management of fragmented populations. Conservation Letters 11, e12412
| Call for a paradigm shift in the genetic management of fragmented populations.Crossref | GoogleScholarGoogle Scholar |
Raymond M, Rousset F (1995) GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. Journal of Heredity 86, 248–249.
| GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism.Crossref | GoogleScholarGoogle Scholar |
Shi J, Joshi J, Tielbörger K, Verhoeven KJF, Macel M (2018) Costs and benefits of admixture between foreign genotypes and local populations in the field. Ecology and Evolution 8, 3675–3684.
| Costs and benefits of admixture between foreign genotypes and local populations in the field.Crossref | GoogleScholarGoogle Scholar |
Smith MG, Jones A (2018) ‘Threatened and priority flora list, 5 December 2018.’ (Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia)
Tallmon DA, Luikart G, Waples RS (2004) The alluring simplicity and complex reality of genetic rescue. Trends in Ecology & Evolution 19, 489–496.
| The alluring simplicity and complex reality of genetic rescue.Crossref | GoogleScholarGoogle Scholar |
Valdiani A, Kadir MA, Saad MS, Talei D, Tan S-G (2012) Intra-specific hybridization: generator of genetic diversification and heterosis in Andrographis paniculata Nees. A bridge from extinction to survival. Gene 505, 23–36.
| Intra-specific hybridization: generator of genetic diversification and heterosis in Andrographis paniculata Nees. A bridge from extinction to survival.Crossref | GoogleScholarGoogle Scholar |
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P (2004) MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Resources 4, 535–538.
| MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data.Crossref | GoogleScholarGoogle Scholar |
Van Rossum F, Hardy OJ, Le Pajolec S, Raspé O (2020) Genetic monitoring of translocated plant populations in practice. Molecular Ecology 29, 4040–4058.
| Genetic monitoring of translocated plant populations in practice.Crossref | GoogleScholarGoogle Scholar |
Waser NM, Price MV (1989) Optimal outcrossing in Ipomopsis aggregata: seed set and offspring fitness. Evolution 43, 1097–1109.
| Optimal outcrossing in Ipomopsis aggregata: seed set and offspring fitness.Crossref | GoogleScholarGoogle Scholar |
Weeks AR, Sgro CM, Young AG, Frankham R, Mitchell NJ, Miller KA, Byrne M, Coates DJ, Eldridge MDB, Sunnucks P, Breed MF, James EA, Hoffmann AA (2011) Assessing the benefits and risks of translocations in changing environments: a genetic perspective. Evolutionary Applications 4, 709–725.
| Assessing the benefits and risks of translocations in changing environments: a genetic perspective.Crossref | GoogleScholarGoogle Scholar |
Whiteley AR, Fitzpatrick SW, Funk WC, Tallmon DA (2015) Genetic rescue to the rescue. Trends in Ecology & Evolution 30, 42–49.
| Genetic rescue to the rescue.Crossref | GoogleScholarGoogle Scholar |
Whitney KD, Ahern JR, Campbell LG, Albert LP, King MS (2010) Patterns of hybridization in plants. Perspectives in Plant Ecology, Evolution and Systematics 12, 175–182.
| Patterns of hybridization in plants.Crossref | GoogleScholarGoogle Scholar |
Wilson PG (1992) The Lawrencella complex (Asteraceae: Gnaphalieae: Angianthinae) of Australia. Nuytsia 8, 361–377.
Zecherle LJ, Nichols HJ, Bar-David S, Brown RP, Hipperson H, Horsburgh GJ, Templeton AR (2021) Subspecies hybridization as a potential conservation tool in species reintroductions. Evolutionary Applications 14, 1216–1224.
| Subspecies hybridization as a potential conservation tool in species reintroductions.Crossref | GoogleScholarGoogle Scholar |