

Carving up Australia’s arid zone: a review of the bioregionalisation of the Eremaean and Eyrean biogeographic regions
M. C. Ebach A C and D. J. Murphy B
A C and D. J. Murphy B
A Changing Earth Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
B Royal Botanic Gardens Victoria, Melbourne, Vic. 3004, Australia.
C Corresponding author. Email: mcebach@gmail.com
Australian Journal of Botany 68(3) 229-244 https://doi.org/10.1071/BT19077
Submitted: 24 April 2019 Accepted: 20 April 2020 Published: 1 June 2020
Journal Compilation © CSIRO 2020 Open Access CC BY-NC-ND
Abstract
The Eremaean region, Australia’s arid biome and biogeographic region, has been discussed by botanists (and as the Eyrean, its counterpart for zoogeographers) for over 150 years, yet little progress was made in defining it as an area of endemism until the 2000s. As Australia’s largest biome and biogeographic region, the Eremaean has been defined in a climate sense, but is a historically composite biogeographic area. Taxa that inhabit the Eremaean (Eyrean) tend to display sister relationships to those outside the biome in temperate and monsoonal biome areas, indicating that two or more temporally discordant distributional patterns exist in the Australian flora and fauna. The future of Eremean and Eyrean bioregionalisation will need to incorporate these temporal patterns when constructing new bioregionalisations and historical and climate-based biogeographic models.
Additional keywords: Bassian, biomes, comparative biogeography, Euronotian, Torresian.
This Zone is considered to cover the whole of arid Australia [Burbidge 1960, p. 79].
This includes the whole of the interior, southern and western part of the continent, the coastal ranges on the east and south-east… [Spencer 1896, p. 198].
Introduction
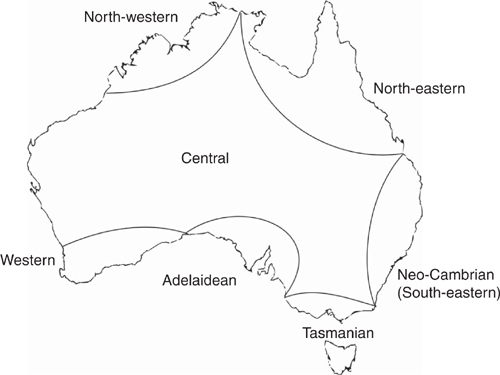
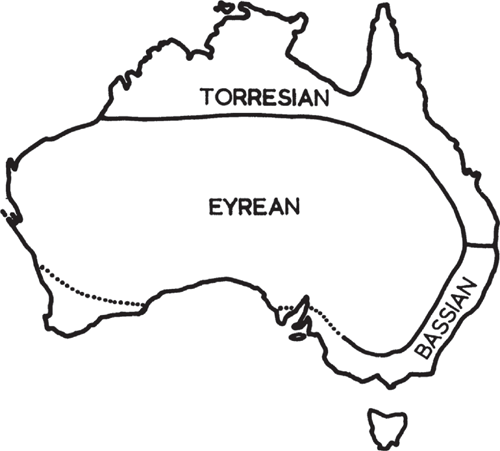
The arid Australian inland is a geographical feature that has been highlighted by several notable early naturalists. For instance, Hooker described the ‘vegetation of Australia’ as being ‘confined to a belt of more or less fertile land surrounding an arid desert, which occupies perhaps two-thirds of its total area’ (Hooker 1859, p. xxx). The first naturalist to group the vegetation of the Australian interior was Ferdinand von Mueller (1858; see Table 1), whereas Ralph Tate was the first to propose actual phytogeographic regions, namely the Euronotian, Autochthonian and the Eremaean (Tate 1889; Fig. 1). The first zoogeographic regions were proposed earlier, by Edmund Julian Tenison-Woods, who identified seven regions including ‘The Central, comprising all the inland waters and central regions’ (Tenison-Woods 1882, p. 49; see Ebach 2012) but considered the north-western and western divisions ‘arbitrary, as of these regions so little is known’ (Tenison-Woods 1882, 49 footnote; Table 2; Fig. 2). The regions of Tenison-Woods were largely ignored by zoologistsA who preferred a revised version of Tate’s regions that were proposed by Baldwin Spencer (1896), namely the Bassian, Torresian and Eyrean (Fig. 3). Although both botanists and zoologists used a similar biogeographic scheme or bioregionalisation, few could decide on the boundaries of the arid inland region. For example, Tate defined the Eremaean as the:
dominant in the dry region, which has its centre in the Lake Eyre Basin […]. It is bounded on the north and north-east by the Indo-Australian vegetation; on the east and south-east by the typical Euronotian Flora, and on the extreme south-west by the Autochthonian [Tate 1889, p. 315]B.
Spencer (1896), however, ignored the Autochthonian, rather his Eyrean subregion:
includes the whole of the interior, southern and western part of the continent, the coastal region on the east and south-east separating it from the Torresian subregion in the north-east and the Bassian sub-region in the south-east [Spencer 1896, p. 198; see also Main et al. 1958, fig. 4; Serventy and Whittell 1976, fig. 1; Fig. 4].

|

|

|

|
|
In fact, zoologists ignored the south-west subregion as an area of endemism for half a century. For instance, ornithologists Serventy and Whittell (1951, 1976) stated that the:
South-West is an area which at present cannot be classified as a simple zoo-geographic region, neither as a uniform portion of the Eyrean sub-region, as Baldwin Spencer originally thought […] It is a district occupied by an intermingling of two faunas, the Eyrean and the Bassian [Serventy and Whittell 1976, pp. 49–50].
Other zoologists had similar sentimentsC. Entomologist Ian Murray Mackerras, for example, defines the Eyrean province as comprising ‘the whole of the semiarid to arid interior within about the 500 mm (20-inch) isohyet’ (Mackerras 1960, p. 196), with an impoverished insect fauna. Mackerras also noted that:
Tate’s additional Autochthonian province in the south-west has been discarded, because of its fauna, although rather distinctive, proved to be predominantly Bassian in relationships [Mackerras 1960, p. 196].
Similarly, herpetologists Main et al. (1958, p. 228) defined the Eyrean as ‘a large central, western and southern area comprising the rest of the continent’, with the south-west containing both Bassian and Eyrean forms. However, not all zoologists dismissed Tate’s Autochthonian subregion. George Edward Nicholls, a zoologist, attempted to incorporate the Autochthonian (he renamed it the Hesperonotian) into a general biogeographic scheme. Interestingly, Nicholls also included the Mesohesperian, an overlap zone between ‘the EremianD and agreeing fairly closely with Prescott’s Sclerophyll Woodland and Scrub’ (Nicholls 1933, p. 94). Neither Nicholls’ general biogeographic scheme nor regions were adopted; however, his Mesohesperian does appear as Interzone Area 1 in Burbidge (1960, fig. 1; see below).
The division of Australia into regions and subregions has resulted in the broad acceptance of three regions by both zoologists and botanists. The largest of these, the Eremaean and Eyrean, have been defined differently by various authors, some have included the south-west, whereas others have treated it as a separate subregion. Regardless, both zoologists and botanists have further subdivided the Eremaean and Eyrean region into provinces. At this level the contrast between different bioregionalisations is profound and in this paper we review and explore the history and implications of these bioregionalisations of Australia’s arid zone.
Attempts at subdividing the Eremean and Eyrean
The idea that biogeographic areas have hard boundaries was questioned most of all in Australia by US herpetologist John A. Moore, who sought criteria to identifying and quantify zoogeographic regions in Australia (see below). Moreover, faunal and floral surveys had clearly shown areas of ‘intermingling’ or overlap of taxa. However, this had not deterred both zoogeographers and phytogeographers from subdividing the Eremaean and Eyrean into smaller provinces. The first attempt was by Tate (1890, p. 204), who divided up the Eremaean into five subdistricts (Table 1). Two of the districts overlapped with three districts in the Euronotian. Unfortunately, Tate’s description of each area is very brief and the areas are not mapped, making it difficult to pinpoint where these overlap zones occur. By 1896, Tate had included the Larapintine regionE,
from the native [Aranda; see Shergold et al. 1991, p. 6] name, Larapinta, of the upper and middle Finke River
which includes:
the southern watersheds of the Levi and George Gill Ranges, and the northern and western drainages that originate from the western extremity of the Mereenie Escarpment and its westerly extensions. This region is bounded on the south-east by remnants of the Cretaceous table-land, and further west by the depressed area centering in Lake Amadeus; on the north the Larapintine table-land abuts on and partially enters into the conformation of the McDonnell Range (sensu stricto, as deliminated by Stuart), which forms the southern boundary to the elevated plain called Burt Plain [Tate 1896, p. 118].
The Larapintine region, with its seemingly precise borders, however, is rarely used in the literature to refer to a biogeographic region, but rather to a flora or biota. Furthermore, later botanists do not use it in continental scale bioregionalisations. For example, Ludwig Diels, a German botanist had, like von Mueller before him, divided the Australian flora into floristic districts (including elements), which to all intents and purposes function as biogeographic areas, that is, they incorporate the distributions of more than one taxon (i.e. species). Diels (1906, fig. 72) had divided the Western Australian part of the Eremaean into the Coolgardie and Austin districts, which both contain Northern (tropical) and Autochthonous (endemic) elements. Gardner (1944) identified only five vegetation types in his Eremean province, whereas Beard (1969, fig. 5) proposed regions, based on the ‘natural regions’ of Clarke (1926), namely a combination of climatic, geographic and geological factors. Beard’s natural regions consisted of Northern, Eremaean and South-western Provinces as well as seven botanical districts, a concept they borrowed from Gardner and Bennetts (1956). Doing (1970) provided the most concise phytogeographical treatment of the Eremaean, dividing the Australian Kingdom into the Central Australian and Eucalyptus sub-kingdoms (Fig. 5). Doing (1970) considered the Centralian sub-kingdom to be ‘more or less identical with the Eremaean region of most Australian authors’ (Doing 1970, p. 87). If so, why not use the same terminology as previous authors? Also, the Centralian subkingdom only included the deserts regions, meaning much of the Eremaean was in the Eucalyptus subkingdom. Beard (1981) was critical of Doing’s bioregionalisation:
[h]is nomenclature, boundaries, and the descriptions of vegetation of the provinces are open to objection and suggest both inadequate consultation of available sources and inadequate knowledge of the country by the author [Beard 1981, p. 346].

|
For example, no subregions of the Eremaean proposed in Diels (1906), Beard (1985), Gardner (1944) or Doing (1970) overlap, including the desert regions. The development of the zoogeographic provinces in the Eyrean subregion is equally discordant.
The first bioregionalisation attempt for the Eyrean region was by entomologist and grazier Thomas Sloane (Fig. 6), who included the Riverina, South Australia, South West Australia, North West Australia and Central Australia as entomological districts within the Eyrean region, the latter corresponding to the botanical ‘Larapinta district of Tate’ (Sloane 1915, p. 147). Sloane, like many late 19th and early 20th century naturalists who proposed these areas, never revealed a method or procedures for delimiting regions and subregions. In contrast, for over 20 years, ornithologist Archibald George Campbell had used species data to produce a list that he had transferred to outline maps. By combining these maps, Campbell created a single map:
showing the main groupings of divisions of the birds as a whole into faunal regions and territories, according to their natural habitats [Campbell 1943, p. 242; Fig. 7].

|

However, fellow ornithologist Allan Keast was unimpressed. Keast rejected Sloane and Campbell’s zones as insignificant ‘from the reptile viewpoint’ (Keast 1959, p. 132). For Keast, the same discernible patterns should occur throughout Australia’s subregions if ‘the subregion concept has any real basis in Australian zoology’ (Keast 1959, p. 129). This is interpreted to mean that subregions should either be seen as areas of origin or as a general grouping of elements. Entomologist Kenneth H. L. Key thought instead zoogeographers should ‘recognise the fundamental importance of ecology, especially in intraregional biogeography’ (Key 1959, p. 207), which would make regions fluid rather than fixed. Eric Pianka also understood the importance of ecology and proposed 14 subregions within the Eyrean based on the behavioural ecology of seven species of lizard (Ctenotus). Although the study of Pianka (1969) also depicted fluid faunas, that were difficult to quantify across multiple genera, which may have different behavioural strategies. Moreover, ‘the responses of populations to these changing environments are different according to the historical differences of the faunal elements’ according to Kikkawa and Pearse (1969, p. 839), who attempted to used bird distributions at species and genus level to generate dendrograms in order to map multiple distributions on geographic maps as faunal provinces. Interestingly, Kikkawa and Pearse recovered Spencer’s Eyrean, Torresian (albeit narrower in the north-east) and Bassian regions, the latter being renamed the Kosciuskan. Here, possibly for the first time, a numerical method had reproduced the ‘static’ areas proposed 70 years earlier. Fluid or dynamic faunal distributions and the ‘fundamental importance of ecology, especially in intraregional biogeography’, seems to be a moot point. Critics of ‘static’ or ‘fixed’ biogeographic areas had changed their position from an ecological to an evolutionary one:
Modern zoogeography must have the same relationship to evolution as does modern taxonomy [Horton 1973, p. 195].
One of the first evolutionary approaches to Australian zoogeography was an endemicity analysis conducted by US ornithologist Joel Cracraft (Fig. 8). Using the distributions of vertebrates, Cracraft plotted 14 areas of endemism based on the bird distributions of Blakers et al. (1984), mammal distributions of Strahan (1983) and herpetological distributions of Cogger (1983). Cracraft (1991) does not explain in detail how this was done (presumably by hand); however, the areas do not resemble the regions of Spencer nor do they share common borders. In fact, much of the Eyrean is unaccounted for, presumably due to lack of data for taxa distributed in these ‘blank’ parts of Cracraft’s map and these are unassigned as areas. Cracraft (1991) ignored a century of zoogeographic work in Australia, which should have rung alarm bells in the wider zoogeographic community. This lack of data or quantitative analysis, such as Parsimony Analysis of Endemicity (Rosen 1988), strongly suggest that Cracraft’s areas may be arbitrary, as botanists who adopted these areas later discovered when attempting to apply the areas to biogeographic analyses, which was met with limited success (e.g. Crisp et al. 1995, 1999).

|
Area relationships: the fall of bioregionalisation and the increased use of biomes
Cracraft’s areas were more about establishing area relationships than testing the areas of endemism themselves (Cracraft 1991, p. 211). The aim of Cracraft (1991) was to investigate:
the extent to which there is hierarchical information about area-relationships contained in the distributions of Australian vertebrates [Cracraft 1991, p. 211].
If ‘modern zoogeography must have the same relationship to evolution as does modern taxonomy’ then the areas should be analogous to taxa and area cladograms and area phenograms the evolutionary lineages. Unlike organismal taxonomy, however, no concern was given to the area taxonomy, that is, the classification that forms the bioregionalisation. The areas that Cracraft used are akin to using paraphyletic or polyphyletic (i.e. aphyletic) taxa in order to create a phylogeny: the problem is that paraphyly and polyphyly ensue (i.e. aphyly). Yet this did not concern most cladistic biogeographers of the 1990s, who were more interested in area relationships and exploring the application of the comparative biogeographic methods, than in discovery of exact boundaries of the areas themselves. This latter endeavour, quantitative bioregionalisation, has become the focus of recent analysis of Australian phytogeography and zoogeography (see, for example, Ebach et al. 2015; Murphy and Crayn 2016); as area definition is the essential first step to historical biogeography. Cracraft’s (1991) areas were initially used by Crisp et al. (1995) and Crisp et al. (1999), who attempted a multi-taxon cladistic biogeographic analysis (Fig. 8a, b). Although their areas differed slightly, it was the first time – at least in Australian biogeography – that botanists had adopted zoogeographic areas. Another concept Crisp et al. (1999) had adopted from Cracraft (1991, fig. 7C) was that all the arid–desert areas were more closely related to each other than they were to any other area. Given that Cracraft’s phenogram was based on the presence of taxa in areas, rather than on cladistic similarities, it is now recognised that it was highly likely that this arid grouping is artificial. Only limited consideration was given to this other than Cracraft ‘included groups in his analysis that have radiated in the arid zone’ (Crisp et al. 1999, p. 352). Very little attention was placed on area definition, or if the data available contained and was heavily influenced by widespread taxa, and based on the results of their cladistics biogeographic analysis, Crisp et al. (1999) concluded:
general phylogenetic relationships among areas, have not been successful. This is probably due to the relative ease of dispersal within a continuous continent [Crisp et al. 1999, p. 355].
Murphy et al. (2019) have recently shown that widespread taxa obscure some patterns, likely due to the ‘layering’ of discordant relationships of different clades. Regardless, the relationships between the regions in continental distributions of taxa, depicted in Crisp et al. (2004) clearly show that the Eremaean shares a closer relationship to other subregions than it does with itself (Ebach and Parenti 2015). Most research in recent years discontinued analysis and discovery of finer-scale natural regions and subregions within Australia (see Murphy and Crayn 2016), and instead used the large regions first described by Tate (1889) as biomes, meaning that areas of endemism were abandoned in favour of areas based largely on climatic definitions (e.g. Crisp et al. 2004).
Quantifying Eremaean and Eyrean distributions: the rise of bioregionalisation
Before the statistical bioregionalisation of Kikkawa and Pearse (1969), an attempt was made by Moore (1961) to empirically distinguish Australian zoogeographic subregions using four criteria, namely:
1. A zoogeographical region must have boundaries, and it should be possible to determine these boundaries with a moderate degree of precision. A zoogeographical region must have boundaries, and it should be possible to determine these boundaries with a moderate degree of precision
2. A zoogeographical region should have a fauna markedly different from that of the adjacent zoogeographical regions
3. A significant proportion of the fauna characteristic of a zoogeographical region should have ranges approximately coextensive with the region
4. A zoogeographical region should be an area isolated, at the present time or during its past, for a sufficient period of time to have allowed the faunas to differentiate […] For zoogeographical sub-regions, such as the divisions of Australia, all four criteria must be met [Moore 1961, pp. 365–366].
Of the four criteria, Moore considered the third the most important as:
by ignoring it, animal geographers have frequently delimited zoogeographical regions that are little more than the ranges of one or several species […] Thus a zoogeographical region must be more than an area with endemic species (criterion 2): many of the endemic species should occur throughout the region and be characteristic of it.
Moore believed that only south-west WA could be distinguished using all four criteria, and that support for the rest of Spencer’s regions comes ‘more from the zoogeographers than the from the fauna’. What is ironic about Moore’s comment is that Spencer dismissed the south-west as a valid region, but regarded the south-west as an overlap zone or a region of ‘intermingling’ of the Eyrean and the Bassian faunas. Main et al. (1958, fig. 4), who, like Moore (1961), were also working on frog distributions, included the south-west as part of the Bassian. So did Iredale, who claimed that ‘Spencer’ ornithological data were:
of the scantiest and most imperfect, so that it is now shown that a peculiar avifauna, closely related to that of the Peronian Area [i.e. Bassian] and very distinct from that of the Centralian Area, lives in that remote district [Iredale 1929, p. 245].
Moreover, Spencer’s Eyrean, Torresian and Bassian regions were uncovered by the quantitative analysis by Kikkawa and Pearse (1969). If we adhere to Moore’s criteria we find that the Eyrean is the most nebulous region, an area that ‘lumped’ the non-Torresian and non-Bassian subregions with the south-west, which ‘coincides roughly with the 20-inch isohyet’ rather than with Spencer’s own boundaries (Moore 1961, pp. 364, 366, 367).
In phytogeographical regionalisation, however, the Eremaean had featured prominently in many quantitative analyses based on distributional data (e.g. Crisp et al. 2004; Bui et al. 2014). These biomes ‘defined using a combination of climate, vegetation structure and ecophysiology’ (Crisp et al. 2004, p. 1551) have dominated Australian phytogeography due to their recovery in large scale quantitative geospatial analyses (see Murphy and Crayn 2016). Moreover, these biomes are largely the same as the areas proposed by Tate (1889) and adopted by Spencer (1896). Previously Australia’s phytogeographical areas were defined in part by endemism (Cracraft 1991; Crisp et al. 1995, 1999), tracks (Ladiges 1998), or by the overlap of taxic distributions (Ladiges et al. 2011, 2012). These areas did not match the biomes recovered by qualitative analysis, more importantly, the area relationships did not support any biome other than the south-west (Ebach and Parenti 2015). Why did a quantitative analysis recover the original areas and biomes but not the biogeographic endemic areas and vice versa? An answer may lie in how quantitative analysis handles distributional data.
Before large-scale quantitative bioregionalisation was adopted by Australian botanists, the Interim Biogeographic Regionalisation for Australia (IBRA) regions were used as a national standard for Australia’s natural regions (Environment Australia 2000). The problem with IBRA was that it included distributional, ecological, soil, water catchments, elevational and climatic data, meaning that it was difficult to tell which data were driving the area definition of regions. Although the IBRA regions are useful for their intended purpose, namely to designate areas for conservation, they proved somewhat problematic for biogeography (see Ebach 2012) as they were not of a comparable scale across Australia and were generated by data that were not easily tested. By using only plant distributions, biogeographers are assured that it is the plant data that is solely responsible for the recovered regions, rather than climatic, soil, water catchments or other data. Regardless, the sole use of plant species distributional data also has its problems, for example, the distributions have no temporal information and rely on the taxonomic rank of species being comparable across plants. For instance, are Eucalyptus species distributions contemporaneous to Banksia species distributions? From the viewpoint of biomes they may be, as climate is likely a strong driver of plant distributions. However, climate is not the sole driver and other stochastic events are now known to impact phylodiversity (Laity et al. 2015). Moreover, changes in climatic cycles scale at a rate that does not fully reflect the taxon distributions and phylogenetic diversity across landscapes, meaning that the impact of refugia and other factors enabling the survival and distribution of lineages is not fully understood. For example, ever since Tate 1896, many have speculated that arid taxa may have closer relationships to coastal temperate or monsoonal taxaF (see Murphy and Crayn 2016), meaning that the Eremaean may be a mix of historical and climatic distributions. A quantitative analysis of Australia’s flora had revealed six regions and 22 subregions (including New Guinea) (González-Orozco et al. 2014; Ebach et al. 2015; Fig. 9). These areas were used in a large-scale comparative biogeographic analysis of Eucalyptus, Banksia and Acacia by Murphy et al. (2019), who discovered that the relationships of all 21 subregions (excluding New Guinea) support both biomes and historical phytogeographic areas, depending on how widespread taxa are treated in the analyses; once widespread taxa were removed from the analysis, historical phytogeographic areas were recovered. In contrast, clustering of the 21 subregions into clades that closely resembled Australian biomes as defined by Crisp et al. (2004), were found if the widespread taxa were left in the analysis. The conclusions of Murphy et al. (2019) were that widespread taxa have affected the way we view the Australian flora in an evolutionary sense, and may represent more recent distributions that are controlled by climate. The impact of Murphy et al. (2019) is not that widespread taxa are driving biomes, but that quantitative data is far more complicated than previously thought, meaning that the data themselves need to be examined and perhaps partitioned before any quantitative analysis is undertaken. In addition, this may explain the discrepancy between the criteria of Moore (1961) and the confirmation of Spencer’s areas by Kikkawa and Pearse (1969). Moore’s criteria are clearly aimed at historical endemism rather than current distributions influenced by recent climatic changes, whereas the opposite is true for the method by Kikkawa and Pearse (1969). If this is the case, then the long debate between what constitutes a true natural phytogeographic area (climate or endemism) may finally have moved on to ‘are our distributional data referring to historical or climatic areas?’. However, how this can be resolved in an ideal way for historical biogeographic analyses remains to be investigated.

|
Future directions in Eremaean bioregionalisation
Over the past 130 years, few biogeographers were able agree on the exact boundaries of the Eremaean and Eyrean region and even fewer could decide upon its subregions. Later in the 20th and 21st centuries there was debate over whether these regions were a result of recent climatic events (e.g. biomes) or of older historical events (e.g. biogeographic areas). Other areas such as Tate’s Autochthonian subregion or the south-west has, with greater sampling over time, appeared in the bioregionalisations of both botanists and zoologists, but yet the arid inland region still eludes biogeographers, probably because it isn’t a monophyletic, that is, a natural biogeographic area.
Eremaean as a biome
Biogeographers need to allow that the Australian flora contains two or more distinct patterns, which are perhaps temporally discordant. The areas where this pertains to the most are the Eremaean and Eyrean areas for two reasons: (1) the Eremaean and Eyrean are distinctively arid (sensu Burbidge 1960; Crisp et al. 2004) and; (2) there are few phylogenetic relationships that support the Eremaean as a natural biotic region (Ebach and Parenti 2015; Murphy et al. 2019). In this sense, the Eremaean and Eyrean are biomes, and not historical biogeographic areas, and should therefore not be used as a single large unit or area in historical biogeographic analyses. However, the approach of using the Eremaean as a biome (e.g. Crisp et al. 2004; Mishler et al. 2014) is empirically valid as it reflects the ecophysiological and climatic factors of an arid region. Historically, however, the Eremaean and Eyrean region is a composite, meaning that taxic ranges have recently expanded into it (e.g. Sloane 1915, p. 146; also Tate 1889 and Nicholls 1933G), and it therefore historical attributes, such as biotic endemism, will be complicated by range expansion and contractions. Because of this there are many fruitful studies that explore more recent evolutionary patterning, such as those approaches to population studies and discovery of potential biological refugia (Byrne et al. 2008). The biome does raise the question of more large-scale overlap (i.e. transition zones or interzones), as in the case of the ‘intermingling’ of Eremaean and Bassian biotas.
Overlap zones in the Eremaean: the Centralian
Early naturalists identified a new region within the Australian fauna, namely the Central or Centralian, first used by Tenison-Woods ‘as comprising all the inland waters and central regions’ (Tenison-Woods 1882, p. 49). In bird bioregionalisation, Gregory M. Mathews used the same term (Mathew 1917, p. 180) without referring to Tenison-Woods, whereas Campbell used:
‘Central’ – the country from the Macdonnell to the Warburton Ranges including the Musgrave and Everard Ranges [Campbell 1943, p. 243].
Sloane also refers to division in the Eyrean subregion based on Carabid distributions, including the ‘Flinders Range, MacDonnell Ranges, ranges of South-western Australia’ (Sloane 1915, p. 146). The Centralian region also refers to the freshwater biogeographic regions of Australia (Iredale 1937, p. 289), which were developed from his earlier avifauna (Iredale 1929). Iredale (1937) separates the faunula (freshwater biota) from the geographic area in the same way early naturalists separated elements from regions. Iredale considers the Centralian to be a geographic area that is inhabited by the ‘Eremian and Eyrean faunula’ (Iredale 1938, p. 287).
Burbidge (1960) identified three interzones, areas that overlap the Eremaean region (Interzones 1–3), and so some description of these is necessary in any review of the bioregionalisation of the Eremaean. Two of these interzones also overlap with the Temperate and one with the Tropical zones. The three interzones were described as having ‘been defined where special circumstances prevent the drawing of zonal boundaries’ (Burbidge 1960, p. 75; Fig. 10). Interzone 1:
has a dry climate with a rainfall mainly winter in incidence […]; Interzone 2 covers much of the central parts of New South Wales
and Interzone 3 is defined as:
North of the Barwon River there is the boundary of the ‘black soil’ system which covers much of Central Queensland and extends as far to the northwest as the Barkly Tableland of the Northern Territory [Burbidge 1960, p. 80].

|
Smith (1984) was the only zoogeographer to note a significant overlap of ‘six major faunal regions of land molluscs […] each with a transition of overlap zone with adjacent regions’ (Smith 1984, p. 178). The overlap of these regions with the arid Centralian region is reminiscent of Burbidge’s Interzones. Unlike Smith, however, Burbidge dismissed Interzones 1 and 2 as not being ‘true floristic units’ due their mosaic distributional character (Burbidge 1960, p. 91). Rather, Burbidge places importance on Interzone 1, an area in the south-west subregion that is mixed with ‘equally characteristic Eremaean elements’ (Burbidge 1960, p. 108). The composite nature of the south-west (or at least part of it) was an observation made independently by both zoogeographers and phytogeographers (e.g. Iredale 1939, Serventy and Whittell 1976; Main et al. 1958).
The possibility of two arid regions
Campbell (1943) was possibly the first to propose the idea of three distinct arid regions, the Desert, ‘a huge continuous area […] mainly in Western Australia […] entirely rivers […] stricken with chronic aridity’, the Central ‘region of mountains contours’, and the Interior ‘all directly traceable to [eastern] coastal stock’ (Campbell 1943, p. 245; Fig. 7). Léon Croizat, citing Lester C. King, also compared the division of the Australian landscape into ‘Great Western Plateau’ and ‘Central Eastern Lowlands’ (Croizat 1964, p. 154, fig. 39; King 1962) with Burbidge’s Eremaean region (Fig. 10). The division may be due to the:
immigration into it from the Torresian and Bassian sub-regions. The Carabidae of the Eyrean sub-region are not numerous, considering its great area; the eastern parts have more genera and species than the western parts, owing to the numerous Torresian and Bassian forms which have invaded the eastern borders of the sub-region [Sloane 1915, p. 146].
The same observation has been made by Murphy et al. (2019) using paralogy-free subtree analysis and the transparent method on the areagrams, namely, the area relationships based on the cladograms of three plant major Australian clades: the eucalypts, Acacia and Banksia. The result clearly shows that there is no relationship between the western and eastern arid subregion once widespread taxa have been removed. Moreover, the Eastern Desert subregion is more closely related to the temperate eastern regions than it is to any other desert region. Perhaps earlier theories of range expansion of coastal taxa into arid regions made by naturalists such as Sloane (1915), Tate (1889) and Nicholls (1933), and later by Burbidge (1960) and Schodde (1989), may explain the east–west disparity of the arid region? The range expansion of arid taxa into arid areas may explain the arid biome, however, phylogenetic evidence points to two distinct and phylogenetically unrelated arid zones: the higher and drier western plateau, and the lower and wetter eastern lowland. The western arid areas are more closely related to the western coastal areas than to any other arid area and the eastern arid region is more closely related to the eastern and south-eastern coastal areas (sensu Murphy et al. 2019).
Distinguishing overlap zones and fuzzy boundaries
Burbidge’s description of Interzones 2 and 3 are similar to the overlapping boundaries found between regions and subregions. For example, Wallacea has an intermingling of both south-east Asian and Australasian taxa due to the tectonic convergent margin between the Australian, Pacific and Asian plates (Hall 2012). The overlap is due in part to the long history of a convergent plate margin (~40 Ma) and the subsequent interchange of taxa during that time. These overlap zones represent a substantial transition of major taxa between regions (Ferro and Morrone 2014). Along smaller subregions, however, the breaks between areas are more subtle to identify, although extreme in terms of species turnover levels. For example, areas such as the south-eastern break between Victoria and the south-eastern subregions represent ~30% turnover in plant and mammal species despite limited apparent edaphic differentiation (Di Virgilio et al. 2013). However, along such boundaries the intermingling is geographically minor and constitutes a ‘fuzzy’ transition barrier rather than a large area of overlap. Burbidge too acknowledged this ‘fuzziness’:
the boundaries between these Zones are not always sharp, owing to differing tolerances of characteristic genera but it is hoped that the data presented will serve to justify their delineation as phytogeographic units [Burbidge 1960, p. 78].
We could speculate that if the ‘fuzziness’ increases over time, a fuzzy boundary may evolve in to an area of overlap, in which two subregions expand into each other.
If Burbidge’s Interzones 2 and 3 are fuzzy barriers that are not ‘adequate for a comparable treatment’ and, Interzone 1 is an actual area of overlap, then we may hypothesise that: (1) the South-west subregion has been isolated for a long time, either by an arid or semi-arid subregion to the east and north-east and; (2) Interzones 2 and 3 are younger ecological boundaries that shift with climate changes into the temperate zone. These two hypotheses may be corroborated using the comparative biogeographic approach used in Murphy et al. (2019) to see if smaller areas (e.g. provinces) in the Victoria, South-eastern and Atherton subregions share greater relationship with those in the Eastern Desert (Fig. 11).

|
Once biogeographers come to accept that the boundaries between regions and subregions are areas in which taxa intermingle, and perhaps over time evolve into overlap zones, then further progress in bioregionalisation may be made. This progress would manifest itself in defining and dating the evolution of smaller arid subregions of Australia, discovering whether geographical overlap also includes temporal overlap (sensu King and Ebach 2017) and whether these subregions share a relationship with regions elsewhere in the Austral realm, including New Zealand, the Sahul or the Andean region.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This research did not receive any specific funding.
References
Anon. (1882a) Review. In The Sydney Morning Herald, 17 May 1882, 9.Anon. (1882b) The Reviewer. In The Queenslander, 13 May 1882, 594.
Anon. (1896) The Horn expedition to Central Australia. Nature 55, 185–187.
| The Horn expedition to Central Australia.Crossref | GoogleScholarGoogle Scholar |
Barlow BA (1984) Proposal for delineation of botanical regions in Australia. Brunonia 7, 195–201.
| Proposal for delineation of botanical regions in Australia.Crossref | GoogleScholarGoogle Scholar |
Barlow BA (1985) A revised natural regions map for Australia. Brunonia 8, 387–392.
| A revised natural regions map for Australia.Crossref | GoogleScholarGoogle Scholar |
Beard JS (1969) The natural regions of the deserts of Western Australia. Journal of Ecology 57, 677–711.
| The natural regions of the deserts of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Beard JS (1981) The history of the phytogeographic region concept in Australia. In ‘Ecological Biogeography of Australia’. (Ed. A Keast) pp. 335–375. (Dr W. Junk Publishers: The Hague, Netherlands)
Beard JS (1985) Comment on Barlow’s ‘Proposal for delineation of botanical regions in Australia’. Brunonia 8, 381–385.
| Comment on Barlow’s ‘Proposal for delineation of botanical regions in Australia’.Crossref | GoogleScholarGoogle Scholar |
Blake SF, Attwood AC (1942) Geographical guide to the floras of the word. An annotated list with special reference to useful plants and common plant names Part 1. United States Department of Agriculture Miscellaneous Publication 401, 1–336.
Blakers M, Davies SJJF, Reilly PN (1984) ‘The Atlas of Australian Birds.’ (Melbourne University Press: Melbourne, Vic., Australia)
Bui EN, González-Orozco CE, Miller JT (2014) Acacia, climate, and geochemistry in Australia. Plant and Soil 381, 161–175.
| Acacia, climate, and geochemistry in Australia.Crossref | GoogleScholarGoogle Scholar |
Burbidge N (1960) The phytogeography of the Australian region. Australian Journal of Botany 8, 75–211.
| The phytogeography of the Australian region.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Yeates DK, Joseph L, Kearney M, Bowler J, Williams MA, Cooper S, Donnellan SC, Keogh JS, Leys R, Melville J, Murphy DJ, Porch N, Wyrwoll K-H (2008) Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota. Molecular Ecology 17, 4398–4417.
| Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota.Crossref | GoogleScholarGoogle Scholar | 18761619PubMed |
Campbell G (1943) Australian faunal regions. Emu 42, 242–245.
| Australian faunal regions.Crossref | GoogleScholarGoogle Scholar |
Chewings C (1886) ‘The Sources of the Finke River.’ (Thomas and Co.: Adelaide, SA, Australia)
Clarke EC (1926) Natural regions in Western Australia. Journal and Proceedings of the Royal Society of Western Australia 12, 117–132.
Cogger HG (1983) ‘Reptiles and Amphibians of Australia’, 3rd edn. (A.H. & A.W. Reed: Sydney, NSW, Australia)
Cogger HG, Heatwole H (1981) The Australian reptiles: origins, biogeography, distribution patterns and island evolution. In ‘Ecological Biogeography of Australia’. (Ed. A Keast) pp. 1331–1374. (Dr W Junk: The Hague, Netherlands)
Cracraft J (1991) Patterns of diversification within continental biotas: hierarchical congruence among the areas of endemism of Australian vertebrates. Australian Systematic Botany 4, 211–227.
| Patterns of diversification within continental biotas: hierarchical congruence among the areas of endemism of Australian vertebrates.Crossref | GoogleScholarGoogle Scholar |
Crisp MD, Linder HP, Weston PH (1995) Cladistic biogeography of plants in Australia and New Guinea: congruent pattern reveals two endemic tropical tracks. Systematic Biology 44, 457–473.
| Cladistic biogeography of plants in Australia and New Guinea: congruent pattern reveals two endemic tropical tracks.Crossref | GoogleScholarGoogle Scholar |
Crisp MD, West JG, Linder HP (1999) Biogeography of the terrestrial flora. In ‘Flora of Australia, Vol. 1’, 2nd edn. (Eds AE Orchard, HS Thompson) pp. 321–367. (CSIRO Publishing: Melbourne, Vic., Australia)
Crisp MD, Cook LG, Steane DA (2004) Radiation of the Australian flora: what can comparisons of molecular phylogenies across multiple taxa tell us about the evolution of diversity in present-day communities? Philosophical Transactions of the Royal Society of London – B. Biological Sciences 359, 1551–1571.
| Radiation of the Australian flora: what can comparisons of molecular phylogenies across multiple taxa tell us about the evolution of diversity in present-day communities?Crossref | GoogleScholarGoogle Scholar |
Croizat L (1964) ‘Space, Time, Form. The Biological Synthesis.’ (L. Croizat: Caracas, Venezuela)
Deane H (1896) President’s address. Proceedings of the Linnean Society of New South Wales 10, 619–667.
| President’s address.Crossref | GoogleScholarGoogle Scholar |
Di Virgilio G, Laffan SW, Ebach MC (2013) Quantifying high resolution transitional breaks in plant and mammal distributions at regional extent and their association with climate, topography and geology. PLoS One 8, e59227
| Quantifying high resolution transitional breaks in plant and mammal distributions at regional extent and their association with climate, topography and geology.Crossref | GoogleScholarGoogle Scholar | 23560044PubMed |
Diels L (1906) Die Pflanzenwelt von West-Australien südlich des Wendekreises: mit einer einleitung über die Pflanzenwelt Gesamt-Australiens in Grundzügen. In ‘Vegetation der Erde VII’. (Eds O Drude, A Engler) pp. 1–143. (Wilhelm Engelmann: Leipzig, Germany)
Doing H. (1970) Botanical geography and chorology in Australia. Mededelingen van de botanische tuinen en het Belmonte Arboretum der Landbouwhogeschool te Wageningen 13, 81–89.
E. G. B. (1896) The Horn Expedition to Central Australia. Journal of Botany, British and Foreign 34, 403–405.
Ebach MC (2012) A history of biogeographical regionalisation in Australia. Zootaxa 3392, 1–34.
| A history of biogeographical regionalisation in Australia.Crossref | GoogleScholarGoogle Scholar |
Ebach MC, Parenti LR (2015) The dichotomy of the modern bioregionalisation revival. Journal of Biogeography 42, 1801–1808.
| The dichotomy of the modern bioregionalisation revival.Crossref | GoogleScholarGoogle Scholar |
Ebach MC, González-Orozco CE, Miller JT, Murphy DJ (2015) A revised area taxonomy of phytogeographical regions within the Australian Bioregionalisation Atlas. Phytotaxa 208, 261–277.
| A revised area taxonomy of phytogeographical regions within the Australian Bioregionalisation Atlas.Crossref | GoogleScholarGoogle Scholar |
Environment Australia (2000) Revision of the Interim Biogeographic Regionalisation for Australia (IBRA) and development of Version 5.1: summary report. Environment Australia, Canberra, ACT, Australia.
Ferro I, Morrone JJ (2014) Biogeographical transition zones: a search for conceptual synthesis. Biological Journal of the Linnean Society. Linnean Society of London 113, 1–12.
| Biogeographical transition zones: a search for conceptual synthesis.Crossref | GoogleScholarGoogle Scholar |
Gardner CA (1944) The vegetation of Western Australia with special reference to the climate and soils. Journal of the Royal Society of Western Australia 28, 11–87.
Gardner CA, Bennetts HW (1956) ‘The Toxic Plants of Western Australia.’ (West Australian Newspapers Ltd: Perth, WA, Australia)
González-Orozco CE, Thornhill AH, Knerr N, Laffan SW, Miller JM (2014) Biogeographical regions and phytogeography of the eucalypts. Diversity & Distributions 20, 46–58.
| Biogeographical regions and phytogeography of the eucalypts.Crossref | GoogleScholarGoogle Scholar |
Hall R (2012) Late Jurassic–Cenozoic reconstructions of the Indonesian region and the Indian Ocean. Tectonophysics 570–571, 1–41.
| Late Jurassic–Cenozoic reconstructions of the Indonesian region and the Indian Ocean.Crossref | GoogleScholarGoogle Scholar |
Hedley C (1894) The faunal regions of Australia. Reports of the Australian Association for the Advancement of Science 5, 444–446.
Hedley C (1904) The effect of the Bassian isthmus upon the existing marine fauna: a study in ancient geography. Proceedings of the Linnean Society of New South Wales 28, 876–883.
Hooker JD (1859) ‘The Flora of Australia, its Origin, Affinities, and Distribution; Being an Introductory Essay to the Flora of Tasmania.’ (Lovell Reeve: London, UK)
Hopper SD (1979) Biogeographical aspects of speciation in the southwest Australian flora. Annual Review of Ecology and Systematics 10, 399–422.
| Biogeographical aspects of speciation in the southwest Australian flora.Crossref | GoogleScholarGoogle Scholar |
Horton D (1973) The concept of zoogeographic subregions. Systematic Zoology 22, 191–195.
| The concept of zoogeographic subregions.Crossref | GoogleScholarGoogle Scholar |
Iredale T (1929) The avifaunal districts of Australia. In ‘Report of the Nineteenth Meeting of the Australasian Association of the Advancement of Science’. (Ed. CE Lord) pp. 244–245. (Australasian Association for the Advancement of Science: Sydney, NSW, Australia)
Iredale T (1937) Basic list of the land Mollusca or Australia. Australian Zoologist 12, 287–333.
Iredale T (1938) The faunal divisions of Western Australia. Australian Journal of Science 1, 102–103.
Iredale T (1939) A review of the land Mollusca of Western Australia. Records of the Western Australian Museum and Art Gallery 2, 1–85.
Iredale T, Whitley GP (1938) The fluvifauna of Australia. South Australian Naturalist 18, 64–68.
Keast A (1959) Australian birds: their zoogeography and adaptations to an arid continent. In ‘Biogeography and Ecology in Australia’. (Eds A Keast, RL Crocker, CS Christian) pp. 89–114. (Dr W. Junk Publishers: The Hague, Netherlands)
Kerle JA, Fleming MR (1996) A history of vertebrate fauna observations in central Australia: their value for conservation. In ‘Exploring Central Australia: Society, the Environment and the 1894 Horn Expedition’. (Eds SR Morton, DJ Mulvaney) pp. 341–366. (Surrey Beatty & Sons: Sydney, NSW, Australia)
Key KHL (1959) The ecology and biogeography of Australian grasshoppers and locusts. In ‘Biogeography and Ecology in Australia’. (Eds A Keast, RL Crocker, CS Christian) Monographiae Biologicae, pp. 192–210. (Springer: Dordrecht, Netherlands)
Kikkawa J, Pearse K (1969) Geographical distribution of land birds in Australia-a numerical analysis. Australian Journal of Zoology 17, 821–840.
| Geographical distribution of land birds in Australia-a numerical analysis.Crossref | GoogleScholarGoogle Scholar |
King LC (1962) ‘Morphology of the Earth.’ (Hafner: New York, NY, USA)
King AR, Ebach MC (2017) A novel approach to time-slicing areas within biogeographic-area classifications: Wallacea as an example. Australian Systematic Botany 30, 495–512.
| A novel approach to time-slicing areas within biogeographic-area classifications: Wallacea as an example.Crossref | GoogleScholarGoogle Scholar |
Krause K (1912) Goodeniaceae und Brunoniaceae. Das Pflanzenreich: Regni vegetabilis conspectus 54, 1–213.
Ladiges PY (1998) Biogeography after Burbidge. Australian Systematic Botany 11, 231–242.
| Biogeography after Burbidge.Crossref | GoogleScholarGoogle Scholar |
Ladiges P, Parra OC, Gibbs A, Udovicic F, Nelson G, Bayly M (2011) Historical biogeographical patterns in continental Australia: congruence among areas of endemism of two major clades of eucalypts. Cladistics 27, 29–41.
| Historical biogeographical patterns in continental Australia: congruence among areas of endemism of two major clades of eucalypts.Crossref | GoogleScholarGoogle Scholar |
Ladiges PY, Bayly MJ, Nelson G (2012) Searching for ancestral areas and artifactual centers of origin in biogeography: with comment on east–west patterns across southern Australia. Systematic Biology 61, 703–708.
| Searching for ancestral areas and artifactual centers of origin in biogeography: with comment on east–west patterns across southern Australia.Crossref | GoogleScholarGoogle Scholar | 22234419PubMed |
Laity T, Laffan SW, González-Orozco CE, Faith DP, Rosauer DF, Byrne M, Miller JT, Crayn D, Costion C, Moritz CC, Newport K (2015) Phylodiversity to inform conservation policy: an Australian example. The Science of the Total Environment 534, 131–143.
| Phylodiversity to inform conservation policy: an Australian example.Crossref | GoogleScholarGoogle Scholar | 25976346PubMed |
Mackerras IM (1960) Composition and distribution of the fauna. In ‘The Insects of Australia’. pp. 187–203. (Melbourne University Press: Melbourne, Vic., Australia)
Main AR, Lee AK, Littlejohn MJ (1958) Evolution in three genera of Australian frogs. Evolution 12, 224–233.
| Evolution in three genera of Australian frogs.Crossref | GoogleScholarGoogle Scholar |
Mathew GM (1917) Some new Australian birds. Emu 216, 180–185.
Mishler BD, Knerr N, González-Orozco CE, Thornhill AH, Laffan SW, Miller JT (2014) Phylogenetic measures of biodiversity and neo- and paleo-endemism in Australian Acacia. Nature Communications 5, 4473
| Phylogenetic measures of biodiversity and neo- and paleo-endemism in Australian Acacia.Crossref | GoogleScholarGoogle Scholar | 25034856PubMed |
Moore JA (1961) The frogs of eastern New South Wales. Bulletin of the American Museum of Natural History 121, 151–385.
Murphy DJ, Crayn DM (2016) Australian comparative phytogeography: a review. In ‘Handbook of Australasian Biogeography’. (Ed. MC Ebach) pp. 129–153. (CRC Press: Boca Raton, FL, USA)
Murphy DJ, Ebach MC, Miller JT, Laffan SW, Cassis G, Ung V, Thornhill AH, Kerr N, Tursky ML (2019) Do phytogeographic patterns reveal biomes or biotic regions? Cladistics 35, 654–670.
| Do phytogeographic patterns reveal biomes or biotic regions?Crossref | GoogleScholarGoogle Scholar |
Nicholls GE (1933) The composition and biogeographical relations of the fauna of Western Australia. In ‘Report of the Twenty-First Meeting of the Australian and New Zealand Association for the Advancement of Science’, August 1932, Sydney, NSW, Australia. (Ed. AB Walkompp) pp. 93–138. (Australasian Association for the Advancement of Science: Sydney, NSW, Australia)
Pianka E (1969) Sympatry of desert lizards (Ctenotus) in Western Australia. Ecology 50, 1012–1030.
| Sympatry of desert lizards (Ctenotus) in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Pianka ER (1981). Diversity and adaptive radiations of Australian desert lizards. In ‘Ecological Biogeography of Australia’. (Ed. A Keast) pp. 1375–1392. (Dr W Junk: The Hague, Netherlands)
Rosen BR (1988) From fossils to earth history: applied historical biogeography. In ‘Analytical Biogeography: an Integrated Approach to the Study of Animal and Plant Distributions’. (Eds AA Myers, PA Giller) pp. 437–481. (Chapman and Hall: London, UK)
Schodde R (1989) Origins, radiations and sifting in the Australian biota – changing concepts from new data and old. Australian Systematic Botany Society Newsletter 60, 2–11.
Serventy DL, Whittell HM (1951) ‘A Handbook of the Birds of Western Australia.’ (University of Western Australia Press: Perth, WA, Australia)
Serventy DL, Whittell HM (1962) ‘Birds of Western Australia’, 3rd edn. (Paterson Brokensha: Perth, WA, Australia)
Serventy DL, Whittell HM (1976) ‘A Handbook of the Birds of Western Australia.’ (University of Western Australia Press: Perth, WA, Australia)
Shergold JH, Gorter JD, Nicoll RS, Haines PW (1991) Stratigraphy of the Pacoota sandstone (Cambrian–Ordovician), Amadeus Basin, NT. Bulletin of the Bureau of Mineral Resources Geology and Geophysics 237, 1–14.
Sloane TG (1915) On the faunal subregions of Australia. Proceedings of the Royal Society of Victoria 28, 139–148.
Smith BJ (1984) Regional endemism of the south-eastern Australian land mollusc fauna. In ‘World-wide Snails: Biogeographical Studies on Non-marine Mollusca’. (Eds A Solem, AC van Bruggen) pp. 178–188. (Backhuys: Leiden, Netherlands)
Spencer WB (1896) Report on the work of the Horn Scientific Expedition to Central Australia: Part I. Introduction, narrative, summary of results, supplement to zoological report, map. Melville, Mullen & Slade, Melbourne, Vic., Australia.
Strahan R (1983) ‘Complete Book of Australian Mammals.’ (Angus & Robertson: Sydney, NSW, Australia)
Tate R (1889) On the influence of physiological changes in the distribution of life in Australia. In ‘Report of the First Meeting of the Australasian Association for the Advancement of Science’, August–September 1888, Sydney. (Eds A Liversidge, R Etheridge) pp. 312–326. (Australasian Association for the Advancement of Science: Sydney, NSW, Australia)
Tate R (1890) ‘A Handbook of the Flora of Extratropical South Australia, Containing the Flowering Plants and Ferns.’ (The Education Department: Adelaide, SA, Australia)
Tate R (1896) Section II – botany. Part III. Geology and botany. In ‘Report on the Work of the Horn Scientific Expedition to Central Australia’. pp. 171–194. (Melville, Mullen & Slade: Melbourne, Vic., Australia)
Tenison-Woods JE (1882) ‘On the Natural History of New South Wales: an Essay.’ (Thomas Richards, Government Printers: Sydney, NSW, Australia)
Thackway R, Cresswell ID (1995) ‘An Interim Biogeographic Regionalisation for Australia: a Framework for Establishing the National System of Reserves, Ver. 4.0.’ (Australian Nature Conservation Agency: Canberra, ACT, Australia)
von Mueller F (1858) Botanical report on the North-Australian Expedition, under the command of A. C. Gregory, Esq. Journal of the Proceedings of the Linnean Society 2, 137–163.
Webby BD (1978) History of the Ordovician continental platform shelf margin of Australia. Journal of the Geological Society of Australia 25, 41–63.
| History of the Ordovician continental platform shelf margin of Australia.Crossref | GoogleScholarGoogle Scholar |
A Hedley (1904) dismissed the regions of Tenison–Woods as ‘neither natural nor well-defined’ as well as having ‘been overlooked by Tate, Spencer and other writers on zoogeography’ (Hedley 1904, p. 880; Ebach 2012). Reviews at the time of publication were not as negative: ‘In the vegetable kingdom representative forms are most strongly marked [sic] in the various geographical provinces, and the differences are often not only generic but tribual, yet never ordinal’ (Anon. 1882a, p. 9). And, ‘The essay is, although small, a work of considerable importance, its contents evidently being the result of great research, and it is well calculated to occupy a permanent place in the literature of science’ (Anon. 1882b, p. 594).
B Tate is here referring to Hooker who stated: ‘there is a greater specific difference between two quarters of Australia (south-eastern and south-western) than between Australia and the rest of the globe (Hooker 1859, p. xxviii). The idea that there was a distinct southwestern region is prominent in the botanical rather than in the zoological literature.
C Ornithologists have named several birds after Baldwin Spencer’s and Hedley’s terms, such as the Eyrean grass-wren (Amytornis goyderii; formerly Goyder grass-wren), the Bassian Thrush (Zoothera lunuata; formerly White’s thrush), as well as Torresian Crow (Corvus orru; formerly Australian crow), and the Papuan Frogmouth (Podargus papuensis; formerly plumed frogmouth).
D As pointed out in Burbidge (1960, pp. 79–80; as well as Ebach 2012, footnote 15), the Eremaean has had several different spellings since Tate (1889) had coined it Eremian. Diels (1906, p. 372) renamed it Eremaea, which was ignored by Nicholls (1933) and adopted by Gardner (1944) in its Anglicised form (i.e. Eremean). Burbidge (1960) changed the spelling again to its present-day form Eremaean. Burbidge’s spelling is used herein.
E Tate’s description of the Larapintine region is difficult to interpret. A review of Spencer’s Horn Expedition (see also Anon. 1896, p. 187; E. G. B. 1896) describes the Larapintine region as ‘the southwestern part of Northern Territory [sic], from the MacDonnell Range south to Engoordina (Mount Encurdina, 25°10′S, 134°16′E) and west to Lake Amadeus’ (Blake and Attwood 1942, p. 68). We are unsure how Blake and Attwood sourced this additional information, given that ‘Mount Encurdina’ is not listed in the Gazetteer of Australia (http://www.ga.gov.au/placename, accessed 24 April 2020). Henry Dean described the Larapintine region as ‘the central portion of the Eremian Region’ (Deane 1896, p. 659) and Kurt Krause as a ‘table land’ (Krause 1912, pp. 61, 66). Tate based his Larapintine region on Chewings (1886) description of the geology and flora found in the Macdonnell Ranges, mostly the basaltic and granitic areas. Tate’s Larapintine area is still used by geologist to describe the Ordovician Larapintine Sea (~470 Ma), which covers a much larger area, extending northwest and southeast (sensu Webby 1978, fig. 6, 7), and the Larapinta Group stratigraphic unit (https://asud.ga.gov.au/search-stratigraphic-units/results/10184, accessed 14 March 2019).
F ‘That in Post-Pliocene time there originated in Central Australia as Eremian flora developed from Autochthonian and Euronotian elements, and largely modified by Oriental immigrants and the species evolved from them’ (Tate 1896, p. 123).
G Iredale (1939) noted the opposite, namely that the ‘Eremian or Eyrean Faunula […] ranges across the whole of the centre of Australia and runs into the south-west, meeting members of the Autochthonian Faunula (Iredale 1939, p. 2).