

Consequences of prenatal and preweaning growth for feedlot growth, intake and efficiency of Piedmontese- and Wagyu-sired cattle
L. M. Cafe A B C , D. W. Hennessy A B D , H. Hearnshaw A B E , S. G. Morris A F and P. L. Greenwood A C GA Cooperative Research Centre for Beef Genetic Technologies, University of New England, Armidale, NSW 2351, Australia.
B New South Wales Department of Primary Industries, Agricultural Research and Advisory Station, Grafton, NSW 2460, Australia.
C New South Wales Department of Primary Industries, Beef Industry Centre, Armidale, NSW 2351, Australia.
D Present address: 187 Fitzroy Street, Grafton, NSW 2460, Australia.
E Present address: PO Box 433, Grafton, NSW 2460, Australia.
F New South Wales Department of Primary Industries, Wollongbar Agricultural Institute, Wollongbar, NSW 2477, Australia.
G Corresponding author. Email: paul.greenwood@dpi.nsw.gov.au
Animal Production Science 49(6) 461-467 https://doi.org/10.1071/EA08089
Submitted: 7 March 2008 Accepted: 4 August 2008 Published: 13 May 2009
Abstract
Consequences of low (mean 28.0 kg, n = 77) and high (mean 38.4 kg, n = 77) birthweight followed by slow (mean 548 g/day, n = 75) or rapid (mean 859 g/day, n = 79) growth to weaning for feedlot growth, intake and efficiency from 26 to 30 months of age were determined in Wagyu × Hereford (n = 81) and Piedmontese × Hereford (n = 73) cattle. Cattle were selected for study based on birthweight and preweaning growth rate, from multi-modal distributions achieved by imposition of low or high maternal nutrition during pregnancy and lactation, with the objective of achieving as close as possible to a 30% difference in birthweight and a 2-fold difference in preweaning growth rate between progeny groups. High birthweight cattle entered the intake test 57 kg heavier, grew 100 g/day more rapidly, and ate 1.0 kg dry matter /day more than the low birthweight cattle. The high birthweight cattle tended to have a higher feed conversion ratio than low birthweight cattle, but net feed intake did not differ due to birthweight group. Cattle grown rapidly to weaning entered the intake test 29 kg heavier, grew at an equivalent rate, and ate 0.7 kg dry matter/day more than the cattle grown slowly to weaning. No differences in feed conversion ratio or net feed intake were observed between the preweaning groups. When assessed at the same liveweight, differences in dry matter intake and/or feed conversion ratio due to birthweight or preweaning growth were no longer apparent. Interactions between prenatal and preweaning growth, or between sire genotype and early-life growth, were not evident for feedlot growth, intake or efficiency. It is concluded that severely restricted growth during prenatal life or from birth to weaning results in cattle that are smaller and consume less feed at the same age as their well grown counterparts; however, long-term effects of growth during early life on efficiency of utilisation of feed are not evident.
Additional keywords: calf, fetal programming, newborn.
Introduction
Cattle within Australian beef production systems typically spend the earlier phases of their life at pasture and may be finished on a grain-based diet in a feedlot. Pasture-based production systems can vary enormously in the growth rates they elicit in young cattle, due to variations across both space (soil types, climate) and time (season, drought). This often results in widely varying growth paths that may have long-term effects on their growth and the efficiency with which they utilise feed during the feedlot finishing period, with potential economic consequences for the feedlot sector.
Many studies have evaluated the consequences of growth during the preweaning period for subsequent growth and efficiency of cattle (reviewed by Berge 1991). Hennessy and Arthur (2004) reported that Australian Bos taurus cattle with restricted preweaning growth were smaller at feedlot entry, grew similarly in the feedlot, but consumed less feed and had a lower (better) feed conversion ratio (FCR) than cattle that were grown quickly from birth to weaning. There have been few studies on the extent to which variations in prenatal growth rate or birthweights influence efficiency of feed utilisation in later life, or on whether prenatal growth interacts with growth rate from birth to weaning to influence efficiency later in life.
More extreme intrauterine growth retardation of cattle can result in slower growth throughout postnatal life (Greenwood and Cafe 2007). Some evidence of fetal programming effects on feed efficiency was presented by Martin et al. (2007). These authors found that heifer progeny from dams that received enhanced maternal nutrition grew faster to 3 years of age, and tended to have reduced efficiency in the feedlot. However, a related study of Stalker et al. (2006) did not find effects on feedlot growth or efficiency in the steer component of the study. Studies of twin and singleton cattle reported by de Rose and Wilton (1991) showed that any reduction in feed consumption in the feedlot in twins was primarily due to the smaller size of the twin-born cattle.
The objective of this study was to test the hypotheses that severely restricted growth rates during prenatal and/or preweaning life have long-term effects on feed intake and the efficiency of feed utilisation for growth of cattle during feedlot finishing. It was also hypothesised that sire genotype may interact with early-life growth to influence growth, intake and efficiency in the feedlot. To test these hypotheses a steer and a heifer cohort comprising Wagyu × Hereford and Piedmontese × Hereford cattle of low and high birthweights grown either slowly or rapidly from birth to weaning were investigated. Descriptive statistics of the growth paths of the cattle used in this study are provided as background information.
Materials and methods
Animals, experimental design and growth
The cattle investigated in the present study were a subset of progeny that formed part of a larger study on the consequences of birthweight and growth to weaning on subsequent growth, carcass and meat quality characteristics of cattle to 30 months of age. The subset of cattle and their management were described in detail by Greenwood et al. (2006), and further details of the prenatal and preweaning treatments on the entire herd were provided by Cafe et al. (2006).
Briefly, the experimental design was a 2 (low and high birthweight groups) × 2 (slow and rapid preweaning growth groups) × 2 (Piedmontese and Wagyu sire genotypes) × 2 (2001-born heifer and 2002-born steer cohorts) factorial, with 9–11 cattle in each of the 16 cells within the design. The cohorts investigated were female (n = 76) or castrate (n = 78) progeny of nine Piedmontese and eight Wagyu bulls mated to the same herd of Hereford cows for two breeding cycles.
Progeny were selected based on birthweight and preweaning growth rate from multi-modal distributions achieved by imposition of low or high maternal nutrition during pregnancy and lactation (see Cafe et al. 2006; Greenwood et al. 2006). The objective was to achieve a 30% difference in birthweight and a 2-fold difference in preweaning growth rate between groups, these differences being representative of the extremes in prenatal and preweaning growth within the New South Wales North Coast region.
The early-life treatments concluded at weaning at ~7 months of age. The cattle were subsequently backgrounded within their cohort on improved temperate perennial pastures until feedlot entry at ~26 months of age.
Descriptive statistics of the growth characteristics of the two cohorts from birth to feedlot exit are presented in Table 1. The cattle in the low birthweight group were on average 10.4 kg (27%) lighter at birth, 27 kg (14%) lighter at weaning, and 50 kg (10%) lighter at feedlot entry than those in the high birthweight group. Those in the low birthweight group showed no compensatory growth at any stage relative to the high birthweight cattle. There was no difference in birthweight between the cattle in the slow and rapid preweaning growth groups. The slow preweaning group was on average 66 kg (44%) lighter at weaning and 28 kg (5%) lighter at feedlot entry. The cattle in the slow preweaning group did show some compensatory growth compared with their rapidly grown counterparts during backgrounding.

|
General feedlot management
Following backgrounding, the cattle were transported to the Cooperative Research Centre for Beef Genetic Technologies’ ‘Tullimba’ research feedlot near Kingstown (30°20′S, 151°10′E, altitude 560 m) in New South Wales, for grain finishing. The 2001-born heifer cohort arrived at the feedlot in October 2003, and the 2002-born steer cohort arrived in October 2004. On arrival at the feedlot the animals were vaccinated with 5-in-1 vaccine, treated for external and internal parasites, and tagged with an electronic identification device.
The grain-based diet was formulated to be as similar as possible in composition for both cohorts. The diet for heifer cohort contained 12.1 MJ metabolisable energy (ME), 133 g crude protein and 101 g fibre (ADF) per kg dry matter (DM); and that for steer cohort comprised 12.0 MJ ME, 157 g crude protein and 125 g ADF per kg DM; as determined at the laboratory of CASCO Agritech, Toowoomba, Queensland. The heifer cohort remained in the feedlot for a total of 123 days and the steer cohort for 120 days. Each cohort exited the feedlot to be slaughtered as a single group at ~30 months of age. The heifer cohort exited in March 2004, and the steer cohort exited in February 2005.
Feed intake measurement
After an initial 4-week adaptation period of feeding from an open bunk, the cattle were allocated to eight intake pens such that each pen contained at least one but not more than two representatives from each treatment group, and pens held animals of a similar liveweight mean and variance. Each intake pen contained an automated feed intake recorder that measured individual animal intake (Bindon 2001). The intake test period commenced following a 2–3-week period of adaptation to the pens. During the test period, the cattle were removed from their pens weekly and weighed. Both cohorts remained in the intake pens until feedlot exit. An animal from each cohort did not eat from the automatic feeder, and these were removed from the study.
Climatic conditions
The intake testing was conducted through summer for both cohorts. Both summers were warm, but the climatic conditions experienced by the heifer cohort were more severe than those experienced by the steer cohort. The heifer cohort was noticeably heat stressed for periods during their intake measurement whereas the steer cohort was not (M. Wolcott and J. Siddell, pers. comm.). To quantify the differences in the prevailing climate experienced by the two cohorts, the temperature humidity index (THI) at the time of maximum temperature for each day during feed testing was calculated using the equation of Mader et al. (2006):
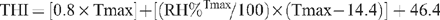
where Tmax is maximum temperature for the day, and RH%Tmax is the relative humidity at the time of Tmax.
Rainfall data but not temperature and humidity data were collected on site, hence extrapolated temperature and humidity data for Tullimba (Silo Data Drill, www.nrw.qld.gov.au/silo/datadrill) were used to assess the prevailing climate during the two feedlot periods. The extrapolated data compared well with the rainfall data collected on site, and with recorded comments from feedlot staff on the timing of the hot conditions.
A THI of 75–78 is rated as dangerous, and 79–83 as an emergency situation for B. taurus cattle in confined feedlot conditions (Mader et al. 2006). During the 80 days in the intake pens, 33 days were within the 75–78 range at the hottest time of the day, and 20 days were within the 79–83 range for the heifer cohort. For the steer cohort, 20 days were within the 75–78 range at the hottest time of the day, and 15 days were within the 79–83 range. A large rainfall event (143 mm in 24 h) near the midpoint of the intake test coincided with an ~30% decline in daily feed intake by the heifer cohort. Intakes increased from this point, but did not return to the pre-event levels. The steer cohort was not subjected to such an extreme rainfall event, and their daily intake was very consistent throughout the intake test period.
The consequences of these climatic extremes on the growth, feed intake and feed efficiency of the cattle, and the differences in the severity of effect between the cohorts will be discussed where relevant.
Feed efficiency and net feed intake calculations
The growth rate for each animal over the intake test period was modelled by linear regression of its weekly weights on day of test. The regression coefficient provided the average daily gain over the test period (ADGt), and the regression estimates were used to predict the liveweight at the start and end of the test period. Mean test liveweight was calculated as the average of the start and end of test liveweights. Mean metabolic liveweight (MMW) was calculated as mean liveweight0.73. The growth rate for each animal over the entire feedlot period (ADGf) was calculated as the difference between the feedlot exit and entry weights on the number of days in the feedlot.
Within each test, net feed intake (NFI) was calculated using a multiple linear regression of daily feed intake (DMI, kg DM/day) on MMW and ADGt. The coefficient of determination (R2) for the regression model was 0.88. The NFI for each animal was then calculated as the difference between its measured daily feed intake and the expected value from the regression (Arthur et al. 2001). Across-test NFI was also determined to allow comparison between the cohorts. Because the fit for the intake regression was better for the within-test analyses than for the across-test analyses, and using the across-test NFI did not show a cohort effect, the within-test NFI was used and the data for the combined analyses are not presented.
The FCR for each animal was calculated as its DMI on ADGt. In addition to the two cattle removed from the study due to failure to consume feed, data from four heifers and one steer were excluded from the analyses due to erratic liveweight measurements or erroneous intake measurements.
Statistical analyses
Analyses of the data were conducted using linear mixed models within the Genstat software package (VSN International, Hemel Hempstead, UK). The model included fixed effects of cohort, sire genotype, birthweight group and preweaning growth group; and random effects of sire nested within sire genotype, and intake pen nested within cohort. Where appropriate, test start weight was included as a covariate in the model to allow assessment of traits at an equivalent liveweight at the start of the intake test.
Results
Effects of birthweight
Cattle in the high birthweight group were 10% heavier at feedlot entry, grew faster in the feedlot, and had a higher DM intake than those from the low birthweight group. The high birthweight cattle tended (P = 0.094) to have a higher FCR (9.7 v. 9.2 kg DMI/kg gain), but there was no difference in NFI between the two birthweight groups.
There was an interaction between cohort and birthweight group in ADGt, as the high birthweight group grew more quickly than the low birthweight group within the steer cohort (1.70 v.1.52 kg, s.e.d. 0.076), while both groups grew at the same rate in the heifer cohort (1.08 v. 1.06 kg, s.e.d. 0.076). There was also a tendency for a significant interaction between cohort and birthweight for FCR (P = 0.076), with the high birthweight heifers having a higher FCR than the low birthweight heifers (10.89 v. 9.95 kg DM/kg gain, s.e.d. = 0.692), whereas there was no difference within the steer cohort.
When compared at an equivalent intake test start weight (591 kg), there were no differences in adjusted least-squares means for ADGt, DMI or FCR between the high and low birthweight groups (Table 2). The cohort by birthweight interaction was significant for adjusted AGDt and FCR, and there was a tendency towards a significant effect of birthweight on DMI (P = 0.076).

|
Effects of preweaning growth
Cattle from the rapid preweaning growth group were 5% heavier at feedlot entry and had higher feed intake than those from the slow preweaning growth group. There was no difference in feedlot growth rates nor were there any differences in FCR or NFI between the preweaning growth groups (Table 2).
When compared at an equivalent intake test start liveweight (591 kg), there were no differences in the adjusted least-squares means between preweaning growth groups for any of the traits.
Effects of sire genotype
No differences in liveweight, growth, intake or efficiency measurements were evident due to sire genotype, and there were no interactions between sire genotype and birthweight or preweaning growth for any of the measured traits.
When compared at an equivalent intake test start weight (591 kg), there were no differences between sire genotypes for any of the traits (Table 2).
Effects of cohort
The steer cohort was 13% heavier at feedlot entry, grew 50% more quickly during the intake test period and 35% more quickly during the entire feedlot period compared with the heifer cohort (Tables 1 and 2). Both cohorts had slower ADGt than for the entire ADGf.
The heifer cohort had lower DMI than the steer cohort and a higher FCR (Table 2). There was no difference in NFI between the two cohorts, which was expected given that within-test NFI calculations were used. When an across-test NFI was calculated, there was also no cohort difference.
Regression of DMI on MMW, within cohort, showed the steers to have a greater slope than the heifers (0.152 v. 0.113, respectively, P = 0.013). The regression accounted for 85.3% of the variance.
When compared at an equivalent start of feed test weight (591 kg), the differences between the two cohorts remained significant for all traits (Table 2).
Correlations between test traits
As expected, there was no correlation between NFI and its component traits MMW and ADGt in either cohort (Table 3). The MMW was not correlated with ADG in the heifer cohort, but was for the steer cohort (r = 0.08 v. r = 0.53, respectively). There was also a significant correlation between MMW and FCR in the heifer cohort but not in the steer cohort (r = 0.28 v. r = 0.08).
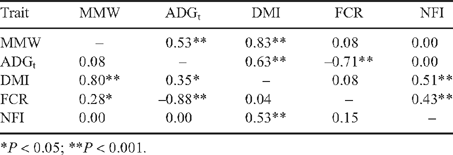
|
Correlations were also conducted on the residuals of the intake variables obtained from the linear mixed model analysis to check that the relationships were independent of the treatment groups. The residual correlations did not differ from the raw correlations and are not presented.
Discussion
This study shows that severely restricted growth during prenatal life or from birth to weaning resulted in cattle that were smaller and consumed less feed at the same age as their well grown counterparts. Cattle from the high birthweight group tended to have a poorer FCR than cattle from the low birthweight group, but NFI did not differ between birthweight groups. Cattle grown rapidly to weaning did not differ in FCR or NFI compared with those grown slowly to weaning. When assessed at the same liveweight, there were no differences evident in DMI or FCR due to birthweight or preweaning growth. Furthermore, there were no effects evident due to sire genotype, or interactions between prenatal and preweaning growth, or between sire genotype and prenatal or preweaning growth for any traits.
Cattle from the high birthweight group were heavier than those from the low birthweight group at feedlot entry. They also grew faster in the feedlot, had a higher DMI and a slightly higher FCR, but NFI did not differ. When they were compared at an equivalent test start weight (591 kg) there was no significant difference between birthweight groups for any trait (Table 2). This suggests that the differences between the birthweight groups in feedlot growth and efficiency were driven by the differing liveweights at the start of the intake test, or by underlying mechanisms correlated with the differences in starting liveweight. The high birthweight group grew faster than the low birthweight group through all production stages to 30 months of age. This suggests that they had greater growth potential, although the differences in growth do not appear to be due to differences in feed conversion efficiency. Hence, it is feasible that there is an influence of restricted nutrition and growth during gestation on hypothalamic appetite centres (Widdowson 1977) and/or tissue growth capacity (Greenwood et al. 1998, 1999, 2000; Greenwood and Bell 2003), which alone or in combination contributed to slower growth of the low birthweight cattle.
There was a tendency for some interactions between birthweight and cohort in growth and efficiency traits during the intake test. These interactions were due to the high and low birthweight heifers growing at the same rate during the intake test, whereas the high birthweight steers grew more rapidly than the low birthweight steers. During the entire feedlot period the high birthweight heifers did grow more quickly than the low birthweight heifers, but there was no difference in their growth during the intake test period. It is possible that the increased susceptibility of larger cattle to heat wave conditions (Mader et al. 2006) was the cause, with the growth rates of the heavier heifers being more affected during the hottest conditions within their intake test period. The steers did not suffer such severe heat wave conditions, and the high birthweight steers grew faster than the low birthweight steers at all times.
Cattle from the slow preweaning growth group were smaller than those from the rapid preweaning growth group at feedlot entry. The growth rates and feed efficiency differences between these groups trended in the same direction as the low and high birthweight groups, but only the difference in DMI was significant (Table 2). The difference in DMI was no longer evident when the preweaning groups were compared at an equivalent intake test starting liveweight (591 kg). This suggests that the only effect of the preweaning growth treatments on feedlot growth and efficiency were due to the long-term effect they had on the liveweight of the cattle at the same age. These results are consistent with those reported by Hennessy and Arthur (2004), where a restriction of preweaning growth of similar magnitude to the present study, imposed from 100 days of age, had little effect on efficiency of feedlot growth when cattle entered the feedlot at 14 months of age.
There were no effects on liveweight, growth or efficiency due to sire genotype in the cattle studied, nor were there any interactions with sire genotype. This was unexpected given the likely differences in the composition of gain during the feedlotting period, which contributed to markedly different carcass and yield characteristics (Greenwood et al. 2006, 2009; Greenwood and Cafe 2007). However, differences in lean body mass (see Greenwood et al. 2006) and hence maintenance energy requirements between the sire genotypes may have also influenced this outcome.
Differences in the climate between the two periods during which the cohorts were in the feedlot appear to have contributed to the variation between the cohorts in growth, intake and efficiency traits. Overall, the correlations (and residual correlations) between intake test growth and efficiency traits (Table 3) were similar to those obtained by Arthur et al. (2001) but varied somewhat between the two cohorts. Certainly the heifers suffered more severe climatic extremes than the steers during the intake test, although the steer cohort grew faster than the heifer cohort during all growth phases. This indicates that there were other differences in growth potential between the two cohorts and by feedlot exit at the same age the steers were 119 kg heavier than the heifers. During the intake test period the heifers had a poorer FCR than the steers. In this regard, the correlation between MMW and ADGt, was significant for the steers (r = 0.53) but not so for the heifers (r = 0.08). Corresponding with this, there was a significant correlation between MMW and FCR for the heifers (r = 0.28) but not for the steers (r = 0.08). The heavier cattle in the heifer cohort had higher DMI than the lighter cattle, but no difference in ADGt, resulting in a higher FCR in the heavier cattle. In comparison, the heavier cattle in the steer cohort had a higher DMI than the lighter cattle, but also had higher ADGt and no difference in FCR. Hence, it appears that the heifers’ intake of feed was limited to the extent that the ratio of gain to maintenance declined with increasing MMW. In support of this notion the slope for the association between MMW and DMI was lower for the heifers than the steers (0.113 v. 0.152, respectively, P = 0.013). This suggests that the differences in correlations between cohorts might be explained by the heifers suffering more from heat stress during the feed test period compared with the steers. This may have been associated with reductions in intake and nutrients available for growth beyond those used for maintenance.
Growth during the entire feedlot period was greater than during the intake test period for both cohorts (Tables 1 and 2) due to higher initial feedlot growth rates in both cohorts. The feed test was conducted during the latter part of the feedlot periods, on average from day 50 to day 120 in the feedlot, and it is not unusual for feedlot growth rates to decline towards the end of the feedlotting period in cattle fed for markets that require heavy carcasses (Robinson 2005). In addition, both cohorts underwent the intake test period during the hot summer months. Robinson (2005) suggested that the most accurate measurement of intake test gain can be calculated by including liveweights taken outside the intake test period in the liveweight regression, as this allows access to a longer growing period. Due to the variable weather around the period of the feed test it was not considered appropriate to attempt to include liveweight measurements taken outside the feed test period in the ADGt regression in the present study.
Conclusions
It is concluded that severely restricted growth during prenatal life or from birth to weaning results in cattle that are smaller and consume less feed at the same age as their well grown counterparts, but has no long-term effect on the efficiency of utilisation of feed. There is no evidence of any interactions between prenatal and preweaning growth or between early-life growth and sire genotype for the Wagyu- and Piedmontese-sired cattle in this study.
Acknowledgements
The financial and in-kind support of the Cooperative Research Centre for Cattle and Beef Quality, NSW Department of Primary Industries, CSIRO Livestock Industries and the University of New England to enable the conduct of this research is gratefully acknowledged. We also wish to acknowledge the considerable efforts of the following research, veterinary, technical and/or farm staff: Lewis Molloy, Keith Newby, William Lee, Max Johnson, Eric Donoghue and Albert Martin, NSW Department of Primary Industries Agricultural Research and Advisory Station at Grafton; Phil Dawes, Peter Kamphorst, Peter Newman and Ross Dicker, NSW Department of Primary Industries Agricultural Research and Advisory Station at Glen Innes; Stuart McClelland, Joe Brunner, Bill Johns, Steve Sinclair, Reg Woodgate, NSW Department of Primary Industries, Beef Industry Centre, Armidale; Reid Geddes, Matt Wolcott, Jason Siddell and Andrew Slack-Smith, Beef Quality CRC and ‘Tullimba’ feedlot. We are also grateful to James and Lyndon Mulligan of ‘Spelga Piedmontese’, the Australian Wagyu Association, and Peter Lee of ‘Waterview Wagyu’ who generously supplied the semen and the bulls used in this study. All experimental procedures were conducted under the approval of the NSW Department of Primary Industries’ North Coast Animal Care and Ethics Committee (Approval No. G2000/05).
Arthur PF,
Archer JA,
Johnston DJ,
Herd RM,
Richardson EC, Parnell PF
(2001) Genetic and phenotypic variance and covariance components for feed intake, feed efficiency, and other postweaning traits in Angus cattle. Journal of Animal Science 79, 2805–2811.
|
CAS |
PubMed |

Berge P
(1991) Long-term effects of feeding during calfhood on subsequent performance of beef cattle (a review). Livestock Production Science 28, 179–201.
| Crossref | GoogleScholarGoogle Scholar |
Bindon BM
(2001) Genesis of the Cooperative Research Centre for the Cattle and Beef Industry: integration of resources for beef quality research (1998–2000). Australian Journal of Experimental Agriculture 41, 843–853.
| Crossref | GoogleScholarGoogle Scholar |
Cafe LM,
Hennessy DW,
Hearnshaw H,
Morris SG, Greenwood PL
(2006) Influences of nutrition during pregnancy and lactation on birthweights and growth to weaning of calves sired by Piedmontese or Wagyu bulls. Australian Journal of Experimental Agriculture 46, 245–255.
| Crossref | GoogleScholarGoogle Scholar |
de Rose EP, Wilton JW
(1991) Productivity and profitability of twin births in beef cattle. Journal of Animal Science 69, 3085–3093.
|
CAS |
PubMed |
Greenwood PL, Bell AW
(2003) Consequences of intrauterine growth retardation for postnatal growth, metabolism and pathophysiology. Reproduction (Cambridge, England) Supplement 61, 195–206.
|
CAS |
PubMed |
Greenwood PL, Cafe LM
(2007) Prenatal and pre-weaning growth and nutrition of cattle: long-term consequences for beef production. Animal 1, 1283–1296.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL,
Hunt AS,
Hermanson JW, Bell AW
(1998) Effects of birthweight and postnatal nutrition on neonatal sheep: I. Body growth and composition, and some aspects of energetic efficiency. Journal of Animal Science 76, 2354–2367.
|
CAS |
PubMed |
Greenwood PL,
Slepetis R,
Hermanson JW, Bell AW
(1999) Intrauterine growth retardation is associated with reduced cell cycle activity, but not myofibre number, in ovine fetal muscle. Reproduction, Fertility and Development 11, 281–291.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Greenwood PL,
Hunt AS,
Hermanson JW, Bell AW
(2000) Effects of birthweight and postnatal nutrition on neonatal sheep: II. Skeletal muscle growth and development. Journal of Animal Science 78, 50–61.
|
CAS |
PubMed |
Greenwood PL,
Cafe LM,
Hearnshaw H,
Hennessy DW,
Thompson JM, Morris SG
(2006) Long-term consequences of birthweight and growth to weaning for carcass, yield and beef quality characteristics of Piedmontese- and Wagyu-sired cattle. Australian Journal of Experimental Agriculture 46, 257–269.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL,
Cafe LM,
Hearnshaw H,
Hennessy DW, Morris SG
(2009) Consequences of prenatal and preweaning growth for yield of beef primal cuts from 30-month-old Piedmontese- and Wagyu-sired cattle. Animal Production Science 49, 468–478.
| Crossref | GoogleScholarGoogle Scholar |
Hennessy DW, Arthur PF
(2004) The effect of preweaning growth restriction on the feed intake and efficiency of cattle on a grain-based diet before slaughter. Australian Journal of Experimental Agriculture 44, 483–488.
| Crossref | GoogleScholarGoogle Scholar |
Mader TL,
Davis MS, Brown-Brandl T
(2006) Environmental factors influencing heat stress in feedlot cattle. Journal of Animal Science 84, 712–771.
|
CAS |
PubMed |
Martin JL,
Vonnahme KA,
Adams DC,
Lardy GP, Funston RN
(2007) Effects of dam nutrition on growth and reproductive performance of heifer calves. Journal of Animal Science 85, 841–847.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Robinson DL
(2005) Assessing the accuracy of modelling weight gain of cattle using feed efficiency data. Livestock Production Science 95, 187–200.
| Crossref | GoogleScholarGoogle Scholar |
Stalker LA,
Adams DC,
Klopfenstein TJ,
Feuz DM, Funston RN
(2006) Effects of pre- and postpartum nutrition on reproduction in spring calving cows and calf feedlot performance. Journal of Animal Science 84, 2582–2589.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Widdowson EM
(1977) Undernutrition and retarded growth before and after birth. Nutrition and Metabolism 21, 76–87.
| Crossref | GoogleScholarGoogle Scholar |
CAS |