

Review of the potential impacts of freight rail corridors on livestock welfare and production
Tellisa R. Kearton A , Amelia Almeida A , Frances C. Cowley A and L. Amy Tait A *
A , Amelia Almeida A , Frances C. Cowley A and L. Amy Tait A *
A
Abstract
The proximity of rail corridors to livestock production enterprises poses potential risks to welfare and production. The association between these factors and production have been extensively investigated. This review aims to assess the potential impact on the basis of existing data in a livestock production context. Due to expansion of freight rail networks through agricultural land, there is a need to investigate potential impacts of rail (including train and track) noise, vibration and visual disturbance on the physiology and behaviour of the livestock and subsequent production traits. Additionally, the factors influencing the impact on animals were characterised broadly as noise, vibration, and visual and spatial disturbance. This information was used to develop conceptual frameworks around the contribution of rail impact on allostatic load, animal welfare and production. Placing rail noise in the context of other, known, noise impacts showed that proximity to the rail line will determine the impact of noise on the behaviour and physiology of the animal. Thresholds for noise levels should be determined on the basis of known noise thresholds, taking into account the impact of noise on allostatic load. Further research is recommended to investigate the behavioural, physiological and production impacts on livestock from proximity to rail corridors. Current literature suggests that the allostatic load will vary depending on the proximity of the animal to the source of stimulus, the type, size or level of stimuli, habituation and the individual animal variation in response to the stimuli.
Keywords: allostatic load, animal welfare, cattle, livestock, noise, rail, sheep, stress.
Introduction
The impacts of rail corridors traversing the landscape have been well studied in the context of wildlife and environmental considerations; however, research on the impact on livestock welfare and production is less prolific. This review considers whether noise, vibration and visual disturbance may have an impact on the welfare and behaviour of livestock. These may influence grazing behaviour, parturition and mothering behaviours, as well as any flow-on effects on predator–prey interactions. This review aims to analyse the literature in the context of potential impacts on sheep and cattle welfare and production from a freight rail network. This work may assist in the development of mitigation and/or management strategies where rail corridors may affect livestock.
Given the scarcity of research specific to domestic livestock and how rail corridors affect their behaviour, physiology and production systems, an interpolative approach was used, extrapolating the results from research investigating how the environmental changes expected from rail traffic affect livestock. Potential impacts, which are more speculative, can be inferred from research on wild sheep and other wild ungulate (e.g. deer, elk, antelope) populations, or from research on the effects of other stressors on domestic livestock. Work conducted by Trigg et al. (2023) also investigated the impact of freight rail noise and vibration on domestic livestock, suggesting that a wider scope of research was needed.
The review considers potential stressors during the construction and ongoing use of the rail corridor. The construction phase, which incorporates significant human activity, comprises a variety of novel sounds and sights, and substantial unpredictability for nearby livestock, and may introduce additional stressors. The novelty of this activity, which may last weeks to months, means that any stress arising can be considered acute (i.e. short-lived, although this descriptor does not imply a particular level of severity). Following construction, a period of adjustment to rail traffic may occur, because the sights and sounds of new rail traffic will appear ‘novel’ to the livestock in the vicinity of the rail corridor. Finally, in the long term, rail traffic will become a ‘normalised’ part of the landscape, after which livestock may learn to predict the ‘actions’ of trains, and, as they do so, will adjust their assessment of the threat that rail traffic provides in a process of habituation. The outcomes of this may include a reduction in stress associated with rail traffic as livestock down-grade their perception of the threat posed (habituation), changes in behaviour as mechanisms to cope with now-expected rail stressors, and/or chronic stress arising from situations where livestock are unable to be habituated or implement coping mechanisms. This review assesses current knowledge of livestock behavioural and physiological responses to the stressors associated with rail corridors, to provide advice on the expected outcomes in individual behaviour and physiology, and their implications for livestock production systems.
The first objective of this review was to identify the main areas of potential impact for animal welfare and production resulting from proximity to rail corridors. Second objective was to propose a conceptual model for ranking and assessing these impacts in the context of other stressors that animals may encounter under a normal production system. Finally, our findings will assist planners and producers in the mitigation of potential impacts and management of exposure of their animals to stressors. Identifying the gaps in the research in this area will help direct future research, developing a more comprehensive understanding of the impact of rail corridors on livestock.
Literature search methodology
Literature was reviewed using a qualitative integrative study methodology, identifying physiological and behavioural impacts on reproductive and productive performance. Research data were reviewed and analysed from across a range of sources and designs so as to form a more comprehensive and integrative understanding of the subject. The factors influencing potential impacts on animals were characterised broadly as noise, vibration, and visual and spatial disturbance.
Two hundred and twenty-six resources were identified as having relevance according to the inclusion criteria, comprising 12 book sections, four conference papers (Dumont and Boissy 2000), three government documents, 10 reports, six theses, three magazine and newspaper articles, two personal communications, and one unpublished work, the remainder (185, or 82%) were peer-reviewed studies. In total, 103 resources have subsequently been cited in this paper.
Literature search protocol
Initially, generic search engines were used to source news articles, opinions and other resources to help inform and direct the literature search terms to include public and producer concerns regarding the impact of rail infrastructure development. Then, the following protocol was utilised to identify relevant literature: first, a search was conducted investigating the impact of rail corridors on domestic livestock, and then the criteria were broadened from there on the basis of particular focus areas. Electronic databases (Google Scholar) and the University of New England library catalogue integrated with the Primo library discovery service were used, drawing from several scientific databases such as ProQuest, EBSCO, Scopus, and Web of Science. Additionally, news articles, opinions and other resources, including public and producer concerns regarding the impact of rail infrastructure development, were sourced. Search terms included, but were not limited to, livestock, rail corridors, noise impacts, anthropogenic disturbance, animal production, train, and livestock underpass.
Literature screening
Literature was assessed on a number of inclusion criteria. When assessing literature pertaining to wildlife, these were restricted primarily to studies on impact of rail and road traffic, as well as underpasses and wildlife corridors when looking at impacts on predation behaviours. Given the lack of specific research in rail impacts on domestic livestock, it was necessary to broaden search limits to allow the identification of literature describing a range of impacts and interpret how they might give insight in this context. Literature examining the impacts of noise on animals specifically focused on livestock in normal production environments such as paddocks and yards.
Research looking at the impacts of rail on wildlife biodiversity and the environmental impacts of rail corridors were excluded, except where this may affect the behaviour and welfare of livestock.
Searches were conducted using the common names and scientific names of the livestock species of interest (primarily sheep and cattle) and the terms ‘rail’, ‘anthropogenic disturbance’, ‘noise’, ‘impact’, ‘train’, ‘corridor’, ‘noise stress’, ‘sound’ ‘underpass’, ‘effects of noise’, ‘auditory’, ‘visual’, by using various combinations to broaden the search base. As well as searching literature published in English, we also encountered resources during our search that required some translation, which was performed using the ‘Google translate’ function.
Data included in Table 1 were assessed and included conditional on meeting the criteria of relevance to potential impact of rail noise and included details on species, noise level, and behavioural or production impact measures and results. Literature was selected for inclusion on the basis of relevance to each focus area, either in terms of impact on the animal (physiology, stress etc.) or the stimulus itself (noise, vibration etc.), by using the following inclusion-topic criteria:
Noise impacts on animals, particularly domestic livestock.
Rail, road and other anthropogenic impacts on wildlife behaviour and habituation, particularly grazing and predation behaviour.
Stress, particularly noise and anthropogenic factors, and animal production, behaviour, physiology and reproduction.
Auditory and visual recognition of ewes/lambs and cows/calves
Auditory communication in domestic animals
Underpass use, including predator behaviour
| Noise level | Duration (if known) | Type of noise | Species | Response | Source | |
|---|---|---|---|---|---|---|
| Unknown | 3 s every 10 s during test | White noise | Sheep | Reduced performance in maze test compared with silent | Doyle et al. (2014) | |
| 34–35 dB | Ambient farm noise A | |||||
| 45–55 dB, 2.7 kH | 2 s repeated three times, with 2 s intervals | Beep from collar | Sheep | Increased vigilance behaviour compared with control | Kearton et al. (2019) | |
| 53.6 dB(A) | Continuous during fattening period | Wind turbine 50 m away | Pigs | Meat quality impacts compared with pigs reared 500 and 1000 m away from the turbine | Karwowska et al. (2015) | |
| 56.3 dB (A) | 12 weeks | Wind turbine 50 m away | Geese | Meat quality reduced when compared with geese reared 460 m away from the turbine | Karwowska et al. (2014) | |
| 58–68 dB, 6.1 kH | 2 s repeated three times, with 2 s intervals | Recording of barking dog | Sheep | Increase in vigilance, and movement compared with contro. | Kearton et al. (2019) | |
| 88 | NA | Machine gun fire at a distance of 328 m | Sheep | No visible behavioural changes | Hauser and Wechsler (2013) | |
| 85–86 dB(c) | 1 min exposure once per day for 5 days | Recorded handling noise(human shouting and metal clanging) | Cattle | Increased heart rate and movement | Waynert et al. (1999) | |
| 90 dB | NA | White noise | Sheep | Decreased thyroid activity | Ames (1978) | |
| 97 dB | NA | Tractor engine sound | Dairy cows | Increased glucose concentration and leukocyte counts in the blood; reduced concentration of haemoglobin | Broucek et al. (1983) | |
| 100 dB | White noise | Sheep | Increased heart rate, respiration | |||
| NA | General noise (4 kHz) | Sheep | Increase in number of lambs per ewe | Ames (1978) | ||
| 105 dB | NA | General noise | Dairy cows | Reduces feed consumption, milk yield, and rate of milk release | Kovalcik and Sottnik (1971) | |
| 109 dB | NA | Machine gun fire at a distance of 80 m | Sheep | Moderate avoidance responses with a rapid return to normal behaviour | Hauser and Wechsler (2013) | |
| 110 dB | 3 min once per day for 4 days of testing,. | Helicopter flyover at 50 m | Goats | No significant change in behaviour or physiological response following habituation (goats) | van der Staay et al. (2011) | |
| NA | General noise | Dairy cows | Increase in glycaemia, nonesterified fatty acids, creatin; decrease in haemoglobin and thyroxin concentration | Broucek et al. (1983) | ||
| 102–120 dB | NA | General noise | Pigs | Influence on hormonal system | Borg (1981) | |
| 128 dB | NA | Machine gun fire at a distance of 16 m | Sheep | Persistent avoidance responses | Hauser and Wechsler (2013) | |
| 120–135 dB | NA | Recorded aircraft noise | Pigs | Increased heart rate | Bond (1963) |
The purpose of this review was to investigate the potential impacts of a freight rail corridor on neighbouring livestock production enterprises, with a (Sibbald et al. 2009) focus on impacts on the animals themselves and how noise, vibration and visual disturbance may affect the way in which they live their lives and respond to threats. This review does not aim to make animal welfare assessments or estimate production impacts.
Authors chose to work within focus areas that aligned with their fields of expertise to be as comprehensive as possible within each context of animal production. These fields encompassed animal welfare, animal health, stress physiology, animal behaviour, reproductive physiology and behaviour, grazing and spatial behaviour, predation, production, animal handling and farm practices. For this reason, screening was completed for each section separately, by one author per section. A framework was proposed utilising the data in the literature to attempt to place rail disturbance within the context of known stressors. Due to the lack of research on direct impacts of rail proximity to livestock, the framework was based on potential and likely impacts of the isolated and cumulative effects of stressors associated with rail corridors.
Personal communications were sought from fellow researchers who have worked with livestock within proximity to rail lines for their anecdotal observations to validate conclusions from the review of literature, particularly when findings from species other than domesticated ruminants were extended to these species.
Impacts of rail stimuli on livestock
Auditory (noise) and vibration disruption
Livestock in general, being prey animals, are sensitive to sounds and tend to respond to sounds by orientating their ears towards the sound source. Studies on livestock sensitivity to sound have predominantly focused on sound intensity (loudness) more than sound frequency (pitch). Pigs, sheep and dogs have similar auditory ranges (Heffner 1998), and may have similar aversion to high noise environments. Sheep and cattle have the ability to hear sounds in the ultrasound range (above 20 kHz), a range which is inaudible to humans (Adamczyk et al. 2015). Sounds in this range, such as rail wheel squeaks, may affect how the sheep respond to machinery (Fourie 2018). Previously, Brouček (2014) investigated the impact of noise across a range of sources, although not focusing on noise associated with rail corridors.
Noise can mask, or inhibit, the perception of sounds (Barber et al. 2010) and has the potential to affect behaviour and communication between animals. This masking is not just related to hearing acuity, but also the cognitive perception of auditory stimuli (Gutschalk et al. 2008). While local background ambient farm noise levels may be difficult to quantify, previous measurements by Thorne (2010) estimated ambient levels at a study farm to be in the range of 34 dBA and 35 dBA. Other characteristics of the sound (continuous or intermittent) contribute to its stressfulness. Sheep have shown ability to adapt to continuous noise, whereas loud, unpredictable, intermittent sounds are often recorded as the most aversive to sheep (Weeks 2008). Sheep show evidence of an ability to habituate, or adjust, to 100 dB intermittent noises (including electrical and diesel engines, jet and propeller planes) within 9 days (Ames and Arehart 1972), while cattle habituated to a recording of abattoir sounds more rapidly than the visual and physical activity associated with it (Grandin 1997). Further to this, Haas and Scrivener (2015) described the impact of high-speed rail on equines in terms of rapid onset rate, i.e. how quickly a loud noise is produced and is measured in decibels per second, resulting in a potential startle effect on animals. The recorded effects of noise on several species of domestic animals is described in Table 1 and demonstrate the knowledge gaps associated in the area of noise and livestock, particularly around habituation. Noises at almost three times that of ambient background farm noise generally result in responses such as increased movement or vigilance behaviours, and increased heart rate, but animals also show capacity for habituation, often within several days (Waynert et al. 1999; van Der Staay et al. 2011). While the impact of high-level (120 dB) noise is known to cause hearing loss in sheep and other animals (Griffiths et al. 1994), more subtle effects of lower-level noise such as masking of communication (Rosa and Koper 2018) are less well known.
Noise as a stressor has been shown to reduce the quality of farm animals’ life, with sounds of approximately 100 dB emerging from the literature as a possible threshold for stress (De la Fuente et al. 2007; Voslarova et al. 2011; Kim et al. 2014). Evidence for thresholds of noise having an impact on domestic ruminants (sheep, cattle and goats) is limited. Below 100 dB, sheep and cattle tend to respond behaviourally; movements such as startle reflexes, or moving away from the sound, show a level of aversiveness, and behavioural responses are a first-line defence against greater impacts on animal welfare at a physical or physiological level. However, even at volumes of ~80–100 dB, sheep and cattle show an ability to habituate to noise, reducing their behavioural responses, often within 5–10 days (Rylander et al. 1974; Waynert et al. 1999).
Exposure of domestic pigs to repeated noise stress cause changes in neuroendocrine regulations, which are characterised by temporal alterations in the responsiveness of the hypothalamic–pituitary–adrenal (HPA) system (Kanitz et al. 2005). That research concluded that repeated exposure of pigs to noise levels of 90 dB affects HPA function, resulting in a state of chronic stress that may have negative implications on animal productivity and welfare. Pigs exposed to 90 dB prolonged or intermittent noise had increased cortisol, and noradrenaline to adrenaline ratio (Otten et al. 2004). Pigs are very sensitive to noise and should not be exposed to constant or sudden noise. Therefore, noise levels above 85 dB must be avoided in buildings where pigs are kept (Fottrell 2009). Chloupek et al. (2009) determined a significant negative influence of noise exposure (80 and 100 dB) on the stress and fearfulness of broiler chickens, while Campo et al. (2005) exposed laying hens to 90 dB truck, train and aircraft noise for 60 min, finding increased stress and fear responses.
Distance from the source of the noise can also ameliorate many of the negative behavioural effects. For example, sheep exposed to the sound of a machine gun showed reduced behavioural signs of disturbance the further from the source they were, and no behavioural signs were observed at a distance of 328 m from the source of the noise (Hauser and Wechsler 2013). Because behavioural responses to stimuli are the primary reaction of animals, livestock will first seek to move away from any rail stimuli they find aversive, and once the stimulus has decreased (as the train passes), may return. Thus, impact of noise and vibration is likely to be greatest where livestock are detained in small areas, such as in handling yards, or in intensive grazing cells, and lowest where paddock dimensions allow stock to move away from the rail line. The impact of distance from the rail line can also be utilised in management strategies, such that stock that may be most vulnerable to the effects of rail stimuli, such as young lambs and new stock who have not been previously exposed to the rail line or ewes in late gestation, are located in paddocks at a greater distance from the tracks.
Noise created by the train will occur congruently with the vibration as the train approaches, and then passes by the livestock. It is therefore difficult to separate the two potential stressors from each other. Previous studies have reported the effect of vertical vibration in poultry, rodents and livestock during road transport (Randall et al. 1997; Abeyesinghe et al. 2001; Garcia et al. 2009; Doggett 2018). However, no known studies have assessed rail vibration on livestock productivity or welfare. It is likely that the intensity and duration of vibration experienced by animals being transported is far greater than that experienced by animals in a paddock near a railway, because the extent of animal behaviour and physiological effects rely on the frequency and magnitude of the vibration (Rabey et al. 2015). The vertical vibration levels experienced by animals being transported are constant for the duration of the trip and have been reported to vary from 0.5 Hz to 25 Hz, whereas railway-derived vibrations are generated primarily at the wheel-rail interface, ranging from 2 to 80 Hz depending on track structure and train velocity (Fahy and Thompson 2015), and then travel into the soil (i.e. ground–borne vibrations). One important aspect to be considered is that the ground-borne vibrations propagating from the track can be felt up to 50 m away (Sheng et al. 1999), which is likely to be below the threshold of effect in livestock.
Ground-borne vibrations also result in ground-borne noise; in that case, rail traffic at night-time brings vibration, noise and light to the landscape, possibly causing stress by disrupting sleep in nearby sheep. The effect of freight train noise and vibration on sleep in humans has been investigated (Croy et al. 2013), with sleep fragmentation (Smith et al. 2016) and increased heart rate during sleep having been observed. While the impact of rail noise and vibration on animal sleep is not known, chronic disturbed sleep can have long-term impacts on neurodegeneration (Owen and Veasey 2020). Sheep have shown sleep characteristics similar to those in humans (Schneider et al. 2020) and similar effects of sleep deprivation (Perentos et al. 2016). Interactions between sleep and noise have been studied in laboratory animals, with Rabat (2008) reporting environmental noise resulting in more sleep disturbance in rats than from white noise. Stress responses are activated by noise through the autonomic nervous system, with levels even below accepted thresholds showing impacts (Turner et al. 2005) and intermittency of noise having a greater impact (Rabat et al. 2004). Ground covering or vegetation, including groundcover, may help reduce or diffuse the direct travel of rail vibration through the paddocks or pastures in which the sheep are housed (Fahy and Thompson 2015). Offering livestock the option to move/rest away from the railway by avoiding the use of small paddocks in the proximity of the railway can give the animals the opportunity to choose whether or not to experience the negative stimulus (Kearton et al. 2020).
Visual disruption
There is some evidence that moving trains, carriages or vehicles per se are an aversive visual stimulus for domestic livestock (H. E. Salvin, pers. comm.). There is no evidence that particular colours or materials may be more aversive than others to livestock and nor is there any evidence that the increased speed of movement of the perceived threat, object or vehicle is associated with increased stress. At close proximity, in handling yards, moving vehicles can be a distraction to livestock, and cause them to baulk. In combination with the auditory and vibration effects when stock are housed close to the rail line, there could also be a tendency for sheep to exhibit startle or escape behaviours and flee when the train is in sight.
Designing yards and races with blocked-out lines of sight of moving vehicles is a long-standing recommendation. Location of yards close to rail corridors may cause problems with ease of handling of sheep, unless they are habituated to passing rail traffic. Tree lines or vegetation may be utilised to break up the visual impact of the trains as they pass by, creating a ‘safe’ barrier between the perceived threat and the animal. Over time, habituation to rail traffic will reduce the perceived threat of moving trains, and the accompanying stress.
Physical rail infrastructure
The physical infrastructure of the rail corridor consists of the railway line (fenced off) and level crossing or underpasses for stock passage across the railway line. The subdivision of paddocks by the rail corridor will reduce paddock size in a fashion similar to a fence line. However, where a simple fence line provides only a physical and visual barrier to free livestock movement across the landscape, a railway line has additional impact in the aversive nature of the rail stimulus (outlined above), but the impacts are not specific to the landscape division, and so are not addressed in greater detail here. Stock underpasses are a unique characteristic of a railway line, and potentially offer positive and negative impacts to livestock production systems.
Underpass and overpass use by wildlife has been extensively investigated, with a number of known and potential impacts. Impacts of use by domestic animals depends on factors such as drainage, maintenance, and whether the underpass will be designed for animals to travel freely across areas divided by the railway or whether it is only for controlled movement (Casburn and Cumming 2009).
While there is potential for predators to exploit the use of an underpass by prey animals, there is little evidence for this occurring (Little et al. 2002; Martinig et al. 2020). Underpasses could provide shade and shelter with the correct design enabling free access, and enables safe moving of livestock from one side of the track to the other (James 2020).
To mitigate the risk of predators congregating around underpasses, it is recommended that monitoring and control measures incorporate stock underpasses as a potential predation-risk area. If the animals have free access to the underpass, it is important to ensure that there is adequate drainage so that the animals are not traversing persistent stagnant or muddy conditions, which may lead to foot rot and other bacterial infections (Abbott and Egerton 2003). Design of the underpass should also consider options that facilitate the flow of movement through the area, avoiding dark openings, uneven footing, and sharp corners (James 2020). Inclusion of a light source in the underpass itself, such as, for example solar-powered lighting above animal passageway, will encourage animals to move effectively from entrance to exit. Vegetation around the entrance can reduce noise and visual disturbances around the underpass, facilitating stock movement (Casburn and Cumming 2009).
Behavioural responses to rail corridor stimuli
The combined stimuli resulting from the construction and on-going traffic of a rail corridor may elicit a range of behavioural and physiological effects on livestock, which can change over time. The characteristics of each specific stimulus need to be assessed in terms of their predictability by the livestock. If a certain level of predictability can be established or ‘learned’ by the animal, then a process of habituation can occur, whereby the stress that a particular stimulus elicits in the animal is reduced, either by a change in how stressful the animal perceives the stimulus to be, or by coping mechanisms implemented by the livestock in response to the stimulus. Stress responses and coping mechanisms are manifested in behavioural responses to the various rail stimuli. If the animal does not reduce its perception of the stressfulness of the stimulus or is not able to implement effective behavioural coping mechanisms, then physiological changes may occur. Temperament is an important consideration when investigating the impact of a novel stressor, and reactions can vary between and within species and breeds within species (Lanier et al. 2000; Atkinson et al. 2022).
Predictability of rail stimuli
Predictability of a negative stimulus is known to influence the subsequent stress and behavioural responses in animals (Weiss 1972), with lambs showing a reduced startle response to a sudden event that was signalled with a light cue compared with those that had received an un-signalled event (Greiveldinger et al. 2007). Furthermore, sheep that are able to control their negative experiences have shown reduced stress responses in virtual-fencing case studies (Kearton et al. 2020).
Although the timing of trains may not always be predictable or consistent, a passing train could be considered a predictable stimulus due to (1) considerable auditory and visual signalling as the train approaches, and (2) consistency of ‘behaviour’ from the train.
The ability of an animal to determine the speed of an object via temporal changes has been observed to have a relationship to body size and metabolism, with smaller animals with high metabolisms showing an ability to perceive changes over finer timescales than do larger animals (Healy et al. 2013). This ability may also be confounded in the case of vehicles and trains, which can move much more quickly than does a natural predator, meaning that timing of the sound and visual approach may be unable to be effectively processed by the sheep’s sensory and neurological capacities. Therefore, sheep and cattle may be less able than are smaller animals to accurately predict when an approaching train will arrive, possibly explaining observations of persistent startle and avoidance behaviours even when these species are accustomed to passing trains. However, due to the lack of literature available, it is difficult to determine whether this sensory capability affects the ability of sheep or cattle to effectively acclimate to passing trains.
During the construction phase of the project, a variety of human and machine activity will result in low predictability of stimuli, but sheep and cattle will quickly learn the constrained spatial impact of the activity. Once the phase of rail traffic adaptation begins, the consistency of train ‘behaviour’ will increase the speed at which sheep learn the limitations of train movement and the potential impact. This is essential for the process of habituation to occur, to minimise the duration of this phase, and for stock to move into the final phase of normalised rail traffic.
Acclimation to the new rail traffic and environment may take some time to occur, with some work finding that sheep adjusting to a new environment had not fully acclimatised to their new surroundings within a 9-week study period (Fordham et al. 1991). The low predictability of activity during construction phase may result in sheep avoiding the area close to the activity during this phase. High predictability during the adaptation to rail traffic phase will shorten this phase, but to reduce the incidence of a startle response in sheep, and promote neutral interactions with rail traffic, grazing sheep within sight and sound of rail traffic, but not containing them close to the track, is recommended in the initial weeks of Phases 1 and 2. Predictability does not only rely on a temporal pattern of rail traffic, but also on low-impact signalling of a more stressful stimulus, such as, for example, an increasing volume of noise from an approaching train before it passes close to stock. There is insufficient information in the literature to make a firm recommendation on a distance at which a startle response is less likely to occur, but allowing stock freedom to respond behaviourally, for example, with flight, will aid in the process of habituation.
Habituation
Habituation occurs when animals are exposed to the same stimulus repeatedly, and eventually stop responding to that stimulus. Suddenness, unpredictability and unfamiliarity combined serve to enhance physiological (e.g. heart rate) and behavioural (e.g. startle or flight) responses to a stimulus (e.g. a passing train), while familiarity can serve to modulate the behavioural response to a sudden event (Désiré et al. 2006). Sheep have also shown reduced responses over subsequent exposures to sonic booms, suggesting that sheep can become accustomed to loud noises, although some sheep still showed avoidance behaviours even after repeated exposure (Rylander et al. 1974). Avoidance behaviour was also observed in dairy cattle exposed to noise, with 81% of heifers choosing to avoid noise at 85 dB when offered the choice (Arnold et al. 2008). A strong habituation effect was observed in work by Rylander et al. (1974), investigating behavioural responses to sonic boom and subsonic flight. Cattle showed slight startle responses to the boom effect, returning to neutral within 10 s. Sheep showed reduced reactions to flyovers over time but were not exposed to sonic boom, so could not be compared to cattle. Ducks showed interrupted behaviour on exposure to the boom effect; however, avian responses varied greatly (Rylander et al. 1974). These differences are highlighted in the findings of Trigg et al. (2023), who found that avian species were affected at lower noise and vibration levels than were mammalian species.
Such physiological and behavioural responses can be energetically expensive, and so through the learning process, habituation allows animals to refine their assessment of the threat a stimulus, such as a passing train, poses, and only respond as necessary. The speed of habituation can be highly variable, depending on the temperament of the animals, the frequency of exposure to the stimulus, the predictability of the stimulus, and the learned consequences of the experiences the animals have with the stimulus.
Additionally, risk-disturbance theory may play a role here. This theory suggests that some areas may be avoided due to human disturbance, but can be regulated by the attractiveness of the resources (Lowrey and Longshore 2017). Therefore, while grazing animals may initially avoid grazing close to human activity, if the area is comparatively attractive such as a higher quality of grazing, they may show a higher tolerance to this activity.
Generational habituation (Guinn 2013) is also a potential benefit that may be utilised in some contexts, and has been observed in wild animals habituating to the proximity of humans (Schell et al. 2018). Generational learning also occurs in domestic livestock, the practice known as ‘hefting’ in some regions in which flocks remain in a home range (Davies et al. 2008), or the practice of selling properties with stock in place where generational knowledge may be preserved. Whether generational learning can occur in livestock in the context of noise and visual disturbances is less well understood.
Throughout this review, there is acknowledgement that habituation may occur and therefore mitigation strategies that may reduce impact during the initial exposure to rail stimuli will potentially become obsolete over time.
Behavioural responses of individuals
A behavioural response to a stimulus is the animal’s first line of defence against stress. A well-understood example is a flight-behaviour response to a perceived threat. Other changes in behaviour that could be expected from rail stimuli (at least initially, and potentially long term) include increased vigilance (visually checking for and assessing potential threats), startle and flight behaviours, changed use of the landscape close to the railway line, among others. These are discussed in detail later in this review. A change in animal behaviour alone, from rail stimuli, is not an indication of increased stress per se, but may be an animal seeking to avoid a potential negative impact from that stimuli. However, changes in behaviour can have negative impacts, for example, if increased vigilance reduces grazing time. Alerting distance can also vary across species and environments to novel stimuli; for example, polar bears detected the approach of a snowmobile over 5 km away, but avoidance responses varied significantly (Andersen and Aars 2008). Behavioural responses to rail stimuli will change over time as animals become habituated to the stimuli associated with it, learning about how trains ‘behave’ and refine their assessment of the threats they pose.
Behavioural responses to the introduction of a rail line in their environment will vary depending on the stage of development, human activity in the area, and time for habituation to occur. It is well known that many wildlife species change their behaviour in response to human-altered landscapes, but it is less evident how this may affect livestock, given their management already involves a certain degree of human disturbance.
Livestock that are habituated to handling show reduced stress responses (Tamioso et al. 2017; De Palo et al. 2018). Habituation results in a reduction of the magnitude of hypothalamic–pituitary–adrenal (HPA) axis activation; however, the degree of this response can be affected by other factors, such as predictability, in which the more predictable the stimulus the more rapidly an animal will habituate. An animal may become habituated to other stimuli in addition to the original stimulus, known as generalisation (Grissom and Bhatnagar 2009). This generalisation may have positive implications for production, such as reduced reactivity to novel stressors, conversely there may be a reduction in predator awareness and vigilance behaviours.
The behaviour of ruminants other than domestic sheep or cattle, such as wild elk and pronghorn, in response to road traffic noise has shown reduced responsiveness (vigilance, flight, travelling and defensive behaviours) with an increasing vehicle traffic (Brown et al. 2012). It was suggested by Brown et al. (2012) that these animals could not afford to maintain responsiveness to the most frequent stimuli, and while this might be beneficial in terms of energy investment, detection of predator activity and other cues may be affected.
Behavioural reactivity to stimuli can be affected by genetic temperament, breed, sex and age in sheep (Dodd et al. 2012), and there is considerable within-flock individual variation. As the stock habituate to the rail stimuli, their average flight distance from trains will reduce, but there may be some proportion of the flock that will always maintain a large flight distance, regardless of their experience. As herd animals, cattle to some extent, and sheep to a large extent, are allomimetic, meaning that the behaviour of one animal can cause a similar response in the rest of the group. A single nervous, highly reactive animal that startles in response to train stimuli can therefore cause disturbance in the rest of the mob. These highly reactive animals are potentially at greater risk for long-term effects on production (e.g. growth or reproduction,). Cattle can respond to stress in conspecifics and become more fearful (Boissy et al. 1998).
There is no quantified evidence to support a recommendation on a particular distance necessary to reduce behavioural impacts of rail traffic on stock. Allowing stock freedom to express behavioural responses during the adaptation phase will be important as a coping mechanism to prevent physiological impacts of stressful stimuli, during the process of habituation. While reduced behavioural responses to rail traffic over time are a good indication of habituation, it is important to consider that lack of reactivity does not mean absence of impact. An example might be a reduction in flight responses as sheep perception of the threat posed by rail traffic is down-graded, but if trains disturb animal sleep cycles, this could have a cognitive, psychological or physiological impact, without eliciting a behavioural response. Monitoring of impacts on production is recommended.
Physiological responses (acute, chronic, adaptation)
Until habituation occurs, if behavioural responses to rail traffic are insufficient to reduce the impact of the rail stimuli on sheep, then physiological responses may result, with acute, chronic and adaptive impacts. These physiological responses are not necessarily negative; for example, an acute flush of adrenaline, increasing blood pressure and heart rate, can stimulate a flight behavioural response, which reduces the risk of a more serious physical impact. However, if physiological responses to stimuli occur frequently and act for long periods, then animal health, welfare and productivity can be affected.
While the conditions of freight rail specifically have not yet been evaluated in terms of their impact on sheep and cattle physiology, it is possible to use known impacts on other species to make predictions about likely impacts of rail tracks on domestic stock.
Increased heart rate during sleep (Croy et al. 2013) responses to freight train noise and vibration have been observed in humans; however, no work has been conducted to investigate effects of rail disturbance on heart rate in sheep. Despite this lack of specific literature, much is known about the impact of various stressors on the health and physiology of sheep. An acute stressor, such as a barking dog, will elicit a plasma cortisol response; however, this response is multimodal and can vary in lactating and non-lactating sheep (Cook 1997). Similarly, repeated stress associated with handling and isolation has been shown to have longer-term impacts on immunological function in young lambs (Coppinger et al. 1991). When considering environmental stressors, the impact of disrupted rest and isolation resulting in chronic stress shows physiological impacts of HPA-axis dysregulation and judgement bias (Verbeek et al. 2019) and learning deficits (Destrez et al. 2013) in sheep.
Physiological impacts of stressors occur when animals are not able to either habituate to the stimulus (so that it is no longer stressful) or adjust their behaviour so that they can cope with a stressful stimulus. Although some acute physiological impacts are fleeting and are stimulants or associates to behavioural responses, such as, for example, increased heart rates associated with flight responses, chronic impacts of rail traffic stimuli on sheep physiology will indicate that the stock are not being offered freedom to respond to the stressful stimuli with a behavioural response. An example could be the freedom to move further away from the railway line to a location where the auditory stimulus and perceived threat of the visual stimulus is lessened. Any physiological impacts will usually become most readily evident in reduced growth or reproduction. More subtle signs of physiological responses, such as increased heart rate or impaired immune function may not be readily detected in grazed stock. Monitoring of sheep for productivity impacts, and ability of the sheep to implement behavioural adaptations so that physiological impacts do not occur, will be essential. Promoting habituation and allowing behavioural responses by initially grazing sheep where they can see and hear trains, but are not fenced close to the railway line, is recommended.
Impacts on production and reproduction
The main potential impacts of a rail corridor on grazing relate to (1) the time spent grazing, and whether the perception of rail traffic as a threat may increase vigilance behaviours at the expense of grazing time when they are located close to the rail corridor, and (2) paddock utilisation, and whether stock will spend less time and, therefore, utilisation of those parts of the paddock close to the rail corridor.
Foraging and vigilance are mutually exclusive activities, and if vigilance is increased, then grazing time may decrease (Lian et al. 2007). When stock perceive a risk or threat of predation, this affects their behavioural patterns and time spent grazing, because the time allocated to maintaining vigilance is increased (Hopewell et al. 2005). The individual behavioural response to a stimulus affects how quickly that individual returns to grazing. The magnitude of a startle response produced by a blast of air to the face of sheep was correlated with the time taken to return to eating following the startle (Salvin et al. 2020). Time spent in vigilance behaviours appears to be directly related to the frequency of traffic passage (Lian et al. 2011). Helicopter traffic in the Grand Canyon national park may be considered analogous to rail traffic in its frequency, volume and duration of auditory impact. Within a 3–5 min feeding bout, helicopter disruption was found to reduce foraging time by 50% for sheep within 50–200 m of a flying helicopter, and 10–30% for sheep within 100–450 m of a helicopter (Stockwell 1991). Sheep more than 450 m distant from flying helicopters were not disturbed during their feeding bouts (Stockwell 1991). Although this indicates that within an individual, short-term feeding bout, loud traffic noise can cause a disturbance in grazing behaviour, the proportion of total active time that sheep are likely to spend on vigilance behaviours, even in high-risk settings is low, being 1–2% of active time, compared to 60–80% of active time spent grazing, on average (Dumont and Boissy 2000; Hopewell et al. 2005). Bouts of induced vigilance (responding to an auditory or visual stimulus) are usually short-lived (seconds), and so the cost to grazing time of greater levels of induced vigilance is expected to be low (Mcdougall and Ruckstuhl 2018).
There is little research on the effect of human disturbance and traffic on paddock utilisation in domesticated ruminants. However, suggestions of the impact on the use of land close to rail corridors for grazing can be inferred from observations of wild ungulates. Proximity to a highway (between 0 and >500 m distance) had only a minor effect on the amount of time that Tibetan antelopes spent foraging, although bouts of foraging were shorter when the antelopes were closer to the highway, most likely due to interruptions to express vigilance behaviours (Lian et al. 2011). Wild bighorn sheep avoid heavy road traffic (Papouchis et al. 2001) and prefer to graze away from roads in their habitat. However, those sheep whose habitats were located in areas with heavy-use roads showed clear signs of habituation and, in fact, used road corridors more frequently than they did areas of their habitat away from road corridors (Papouchis et al. 2001).
When considering the effects of road corridors on wild ungulate grazing and vigilance behaviours, it is important to consider that road corridors may also bring a change in the environment in wilderness areas, in that road corridors are likely to be cleared of trees, whereas areas distant from a traffic corridor, but still utilised by wild ungulates, may be wooded. Thus, wild ungulates may have heightened vigilance as a result of the perception of increased risk in open spaces. In the commercial sheep and cattle production systems of central New South Wales and Victoria, domesticated stock will be habituated to grazing open grasslands, pastures and forage crops, and wooded areas are light in tree density. The implication is that evidence of road proximity affecting grazing behaviours in wild sheep breeds and other wild ungulates may be unrelated to traffic noise, and not relevant to domestic sheep and cattle production systems.
The short-lived duration of vigilance behaviours suggests that once habituation has occurred, it is unlikely that rail traffic will cause an increase in vigilance behaviours to the extent that they reduce grazing time. During the period of habituation after introduction of the rail corridor, while stock are expressing increased vigilance while they are learning and assessing the threat that passing trains pose, it is possible that grazing time may be reduced, as animals allocate more time to vigilance. However, the extent of this increased vigilance, combined with the frequency of expected rail traffic passage to stimulate vigilance behaviour, is unlikely to impede grazing time to the extent that productivity is affected.
Within a flock or herd, variation in fearfulness/boldness is expected. These animals may perceive, or respond differently to, rail traffic, and rail traffic may increase the anxiety of innately fearful individuals more than that of their bold flock-mates. Shy sheep have previously been shown to have a smaller range, and more restricted utilisation of dispersed grazing areas, even in a fairly homogenous grazing environment (Sibbald et al. 2009). Selection for temperament may be one way to improve the habituation of highly anxious flocks to a rail corridor in the long term, but would also be likely to bring other productivity benefits in growth, pasture utilisation, weaning rates and meat quality.
While the impact of rail noise has not yet been studied, an impact of varying levels of road noise has been found on behaviour and productivity in lambs (Quaranta et al. 2002). Most research on the effects of noise on sheep productivity has tested constant noise, which will be quite different in characteristic to the intermittent noise of rail freight. When lambs were exposed to the noise of a motorway (75–95 dB, frequency 100–6300 Hz), for long periods (8 h/day for 42 days), growth rate and feed efficiency reduced by 15%, compared with animals in a control group (Quaranta et al. 2002). In that particular research, the sheep did not show signs of adapting to the noise, rather, effects were cumulative, so that reductions in liveweight gain were greater in the last 2 weeks of the experiment than in the first 2 weeks of the experiment. Dry-matter intake was not affected, which indicates that the reduced performance was affected by increased stress and maintenance energy requirements of the sheep exposed to the noise. However, this contrasts with no effect of similar noise (54–95 dB, with a constant, high frequency of 2000 Hz) on growth performance (Sevi et al. 2001, in Quaranta et al. 2002), suggesting the specific characteristics of the noise, in particular the frequency band and range, may be more important than the volume, when predicting impact on productivity.
Promoting habituation and allowing performance of behavioural responses during the adaptation phase will be important to mitigating the effects of rail stimuli on growth. In research on the effects of noise on lamb growth, subject animals have typically been penned, and unable to change their behaviour to avoid or lessen the impact, for example, by moving away. If grazing animals are allowed space to move away from rail traffic, then the impact of rail noise on growth will be negated, and perhaps removed entirely, in all but the most anxious of animals. However, location of intensive facilities, such as weaning yards or feedlots close to railway lines and rail traffic will restrict the ability of stock held in those facilities to implement behavioural adaptations to lessen the stressfulness of the rail stimuli, and physiological and productivity impacts will be more likely. It is strongly recommended that feedlots are not located close to railway lines or rail traffic. Genetic selection for consistent calm temperament will reduce the perception of the scale of the threat posed by rail stimuli and promote habituation, leading to reduced impacts on growth.
Fertility is known to respond to chronic and acute stressors, with impacts on ovulation rate, abortions, and dystocia. Because the hypothalamus and pituitary gland are involved in reproduction and stress response, one may think that reproductive functions can be influenced by stress. In males and females, reproduction is mediated by the gonadotrophin-releasing hormone (GnRH) produced from the hypothalamus that stimulates the anterior pituitary to release luteinising hormone (LH), this then acts on the ovaries and testes to produce oestrogen or testosterone respectively (Dwyer and Phillips 2008). In this sense, oestrus can be delayed or blocked due to its effect on the preovulatory surge of LH (Ehnert and Moberg 1991). Similarly, stress may also reduce the expression of oestrus behaviour as well as libido in rams (Dobson et al. 2012).
Following a successful reproduction, the greatest cause of lamb losses is the starvation–mismothering complex (Refshauge et al. 2016), which is a well-characterised problem in sheep production systems. Rail traffic poses a risk for mismothering if ewes exhibit startle responses to passing trains, causing separation from their lambs, or if sound or olfactory stimuli from trains interferes with ewe–lamb bonding.
Sheep reactivity (temperament) is known to consistently affect weaning rate in sheep, with calm ewes weaning 10–17% more lambs than nervous ewes (van Lier et al. 2017). Nervous ewes have a lower ovulation rate, resulting in fewer multiple pregnancies than for calm ewes (van Lier et al. 2017). Although temperament is heritable, a stressful environment that results in higher than expected levels of anxiety in breeding ewes could result in a similar physiological response that could affect reproduction rate.
Sound stress, specifically, has shown equivocal relationships with ovulation rate, with continuous sound at high volumes (100 dB) possibly showing an increase in ovulation rate (Ames 1974). This result has not been replicated elsewhere. It is not possible to conclude whether rail noise will have a deleterious effect on sheep reproduction or lambing rates.
Olfactory recognition is an important aspect of social and maternal behaviour in sheep. The role of olfaction in bonding between ewes and lambs is well established, with ewes developing selective olfactory memory for their lambs shortly following birth (Mora-Medina et al. 2016), and interference from other ewes can inhibit this bond. Whether external olfactory stimuli can also interfere with this process is unknown and bears further investigation; however, it is known that following bonding, if olfaction is inhibited, an ewe may reject her lamb despite the presence of visual and auditory stimuli (Numan and Insel 2006). Due to the important role of olfactory stimuli in maternal behaviour, particularly suckling (Alexander and Stevens 1985), potential impacts of olfactory stimuli cannot be ruled out.
Vocal recognition of the mother plays an important role in the maintenance of the ewe–lamb contact. Within days after birth, most lambs can find their mother when hidden behind a canvas (Shillito and Alexander 1975; Nowak 1991). The acoustic features of a lambs’ bleat are an important part of the social bond between ewes and their lambs, when this cue is deficient, there is a negative impact on the maternal response (Morton et al. 2018). Ewes are able to discriminate between the bleats of their own lambs and those of an alien lamb 24 h following birth (Sèbe et al. 2007), and lambs able to discern the acoustic signature of their mother at 48 h old (Sèbe et al. 2007). Because of the lack of literature in this area, no conclusion can be made about whether noise disturbance may play a role in rates of mismothering in a population. Cattle also have highly developed vocal recognition systems and mothers and calves can recognise each other with high frequency calls from 10 days of age (Padilla de la Torre and McElligott 2017). While little is known about the importance of other types of vocalisation in cattle, it is hypothesised that it may be important for other interactions such as mating and dominance status.
The impact of rail traffic on reproduction is mediated by perception of the threat that trains pose. Habituation to rail traffic will reduce the perceived threat and the accompanying stress. Genetic selection in the flock or herd for consistent calm temperament will reduce the perception of the scale of the threat and promote habituation.
As potential rail-traffic interference with maternal bonding cannot be ruled out, it is recommended that young lambs and lambing ewes be situated at a distance from the rail line, which would reduce interference from rail stimuli to ambient levels. There is no existing literature that would suggest the distance at which an impact may be evident, along with the type of stimulus that may be implicated.
Predation is a significant cause of lamb losses in Australian sheep production systems, accounting for ~7% of neonatal lamb deaths (Refshauge et al. 2016). The impact of a rail line on predation behaviour has not been extensively studied in livestock; however, there is a large body of research looking at predator–prey interactions in wildlife affected by roadways and rail lines. The presence of human activity during the construction phase may reduce predation risk as domestic animals may perceive the humans the lesser risk (Muhly et al. 2010). Although no research has examined the effect of underpasses on lamb losses to predation, stock underpasses on rail lines may provide new sources of cover for lambs and ewes, but similarly, may provide cover to predators, such as foxes. In areas with predation risk by wolves in the USA, cattle responded to the presence of these predators by grazing closer to areas of human activity (Muhly et al. 2010).
Discussion
Conceptual model development
While short-term stressors and challenges are normal aspects of life, the ability of an animal to cope with these is affected by the type of stress event, the duration of the stress event, and the combined impact of stressors, known as the allostatic load (Edes et al. 2018). This is an important aspect when considering the impact of rail construction, noise, vibration and other factors.
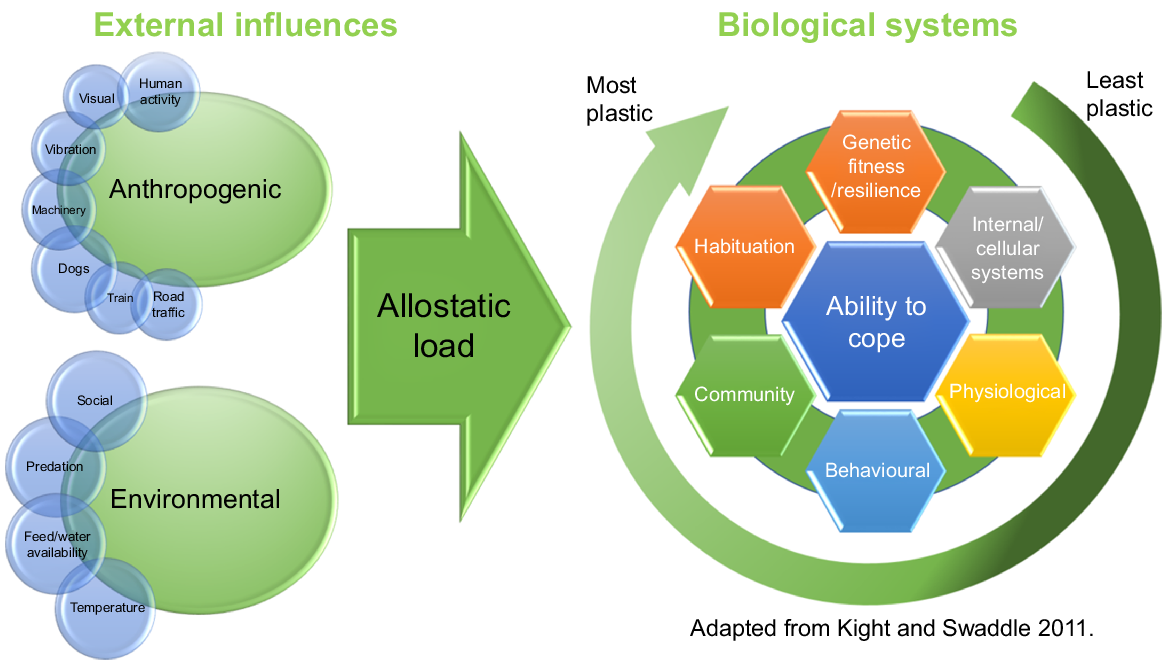
While each of these impacts may be minimal, and largely mitigated through habituation, when combined with other stressors encountered through normal farm activities, such as mustering, weaning, shearing and other husbandry practices, it may contribute to the allostatic load experienced by the animals (Fig. 1).
Diagram adapted from Kight and Swaddle (2011), demonstrating the impacts of anthropogenic and environmental influences on allostatic load and the animal’s ability to cope.
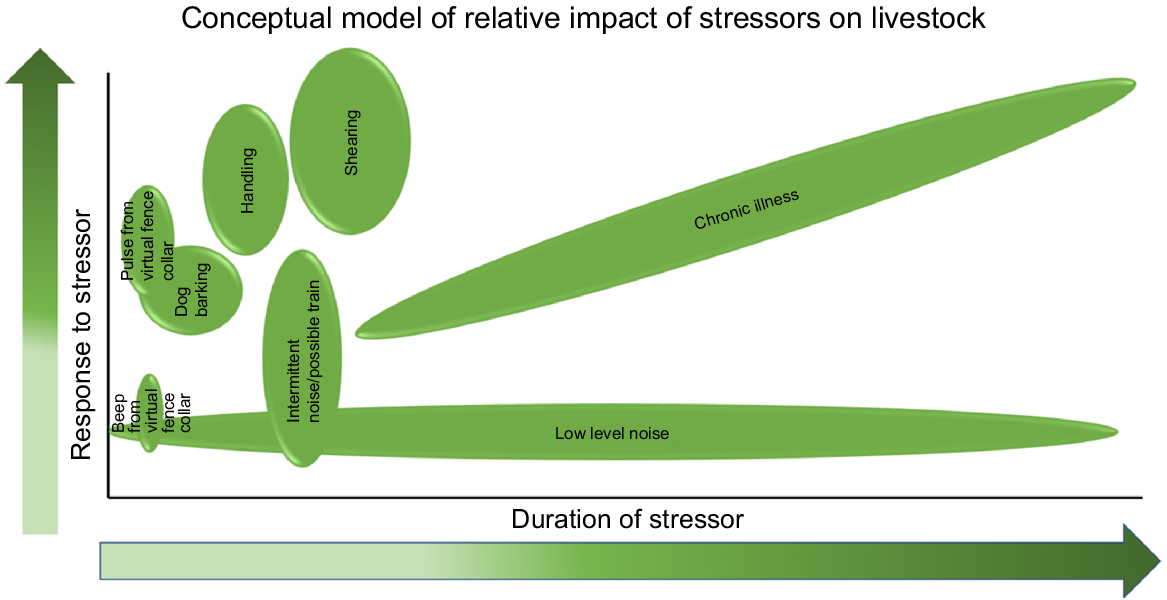
Once the stressors have been identified, we can then try to predict the level of response and duration of the stressors on the animal (Fig. 2). For example, shearing elicits a greater stress response than intermittent rail noise and both of these stressors may occur only for a fraction of the time compared with a consistent low-level noise (Fig. 2).
A conceptual model of the relative impact of certain stressors on livestock in relation to the level and duration of stressor.
A conceptual framework for understanding how noise stimuli affects wildlife has been described in Francis and Barber (2013); however, at present similar framework is not applicable to domestic livestock. This framework describes the impact of the types of anthropogenic noise, from sudden or erratic, through to more frequent sounds with spectral overlap, within the context of the biological impacts described in the literature. The relative lack of comprehensive literature on the impact of anthropogenic noise on domestic animals limits an adaptation of this framework for this purpose at this time. We have proposed the above conceptual models outlining factors likely to affect the ability of an animal to cope with environmental stressors, including noise, in Figs 1 and 2. These figures aim to provide context for the likely/possible impact, given the limited data available. To quantify the impacts of these stressors will require empirical data, which are not currently available.
Conclusions
This review has assessed the available literature across multiple species to provide an understanding of potential impacts on livestock situated close to the proposed rail corridor during multiple phases of construction and beyond. Although the literature testing specific effects of noise, vibration and visual stimuli similar to those presented by rail traffic on domestic sheep and cattle is scant, supplementary information on wild ungulates, and application of fundamental knowledge of sheep and cattle behavioural and physiological responses to stressors has been used to estimate an impact of a new rail freight corridor on grazed sheep and cattle production systems.
The main areas of focus that were found to be relevant in the available literature included, and in no particular order of importance, noise and vibration, visual, behavioural and physiological changes, potential impacts on livestock production, anthropogenic disturbance on livestock and predator behaviour.
Most research on noise or road traffic stimuli considers either acute/short-term or more-or-less continuous stimuli, neither of which fits the pattern of stimuli provided by freight trains passing at regular intervals throughout the day. There is evidence from a range of domestic livestock species to show that a threshold of approximately 100 dB could be considered the point at which noise stimulus moves from a moderately aversive stimulus to a stressor that animals are compelled to avoid. With most rail noise presenting impacts of <100 dB at 15 m suggests that the effect on sheep and cattle is likely to be limited.
The main response of sheep and cattle to rail freight stimuli is likely to be movement away from the passing train. The flight distance to which an animal will relocate, and initial distance from a train which will prompt relocation, are likely to be highly individualistic and dependent on temperament, but in herd animals such as sheep and cattle, may be affected by the behaviour of their conspecifics. The literature on livestock responses to noise indicates that at distances where rail noise is <90 dB, behavioural responses for many animals may be minor. For most animals, the behavioural response of moving away from a train as it passes will prevent higher-order impacts on physiology and productivity from occurring. In production environments where animals are prevented from moving away to a distance at which they feel the threat is suitably diminished, such as feedlots, stockyards, or intensive cell grazing systems, welfare may be affected so that there are physiological responses, or productivity impacts. For some animals, flight responses could have deleterious effects; lambing and recently lambed ewes are at particular risk, and it is recommended that lambing paddocks are not close to the rail tracks. While impacts of noise masking on animal communication have not been investigated in the literature, they should not be excluded as a potential impact. The limitations of currently available literature in this area have affected the accuracy and scope of the suggested impact model and impacts of rail stimuli on livestock; it is anticipated that future research will assist in further refinement of the model.
Habituation will play a major role in reducing stress and behavioural responses over time, particularly when human activity around construction reduces and the rail activity becomes more predictable. Increasing sound from approaching trains will enable animals to predict that a train is soon to pass, which will reduce the stress associated. Learning will refine animal assessment of the threat posed by passing rail freight, which will modulate behavioural responses over time. Part of this habituation involves learning that the threat has passed, and that it is safe to return to an area close to the rail line. It is not possible to predict how long a herd of sheep or cattle will take to habituate to rail freight traffic. There is some evidence that movement responses to noise stimuli reduce after five exposures, but it is likely that some animals will always exhibit a temporary flight response when they are close (within 20–50 m) to passing rail traffic.
Recommendations for future research
The impact of rail corridors on livestock welfare and production presents a number of opportunities for further research because much of the existing literature is focused on wildlife. Where domestic livestock noise and vibration disturbance research exists, the stressors are not easily comparable to the stimuli that freight rail corridors generate. Further research in this area would enable the conceptual models to be refined and applied more appropriately to livestock production contexts, including to species other than the ruminants reported here. Other rail-related issues that could be potential influencers on animal production include the possible redefinition of paddock and pasture size, disruption to drainage, transfer of weeds and other biosecurity issues. These issues were not a part of the current review but should not be discounted and further investigation is required to understand the gravity of railway corridors on animal production systems and the surrounding environment.
Real-time measures, behavioural assessments, physiological measures and production impacts are recommended for long-term impact assessments of rail corridor proximity to livestock. Spatial and land-use investigations in the paddocks adjoining rail corridors would provide additional information on the role of animal choice in mitigating the impact of noise and visual disturbance on livestock behaviour. The role that habituation and generational learning plays in mitigation of disturbance over time would also warrant investigation, assisting producers and managers in their decision-making.
Assessments of mitigation strategies such as buffer zones, reduced stocking densities and vegetation would provide valuable insight into the impact of rail noise and vibration on livestock behaviour, physiology and productivity.
Additionally, investigations into the contribution of rail noise and vibration to allostatic load would provide necessary context to livestock producers, managers and researchers when managing stressors. Development of an assessment tool would be of benefit when attempting to understand the cumulative impact of stressors affecting animals during daily life and when undergoing normal farm practices and procedures.
Data availability
Data sharing is not applicable as no new data were generated or analysed during this study.
Conflicts of interest
This review was generated from a report commissioned by the Australian Rail Track Corporation (ARTC) to investigate the potential impacts of rail corridors on livestock production.
Declaration of funding
This study was funded by the Australian Rail Track Corporation (ARTC), Adelaide, South Australia, Australia.
Acknowledgements
The authors thank and acknowledge the contributions of the University of New England and the Australian Rail Track Corporation (ARTC) for their support in funding this project.
References
Abbott KA, Egerton JR (2003) Effect of climatic region on the clinical expression of footrot of lesser clinical severity (intermediate footrot) in sheep. Australian Veterinary Journal 81, 756-762.
| Crossref | Google Scholar | PubMed |
Abeyesinghe SM, Nicol CJ, Wathes CM, Randall JM (2001) Development of a raceway method to assess aversion of domestic fowl to concurrent stressors. Behavioural Processes 56, 175-194.
| Crossref | Google Scholar | PubMed |
Adamczyk K, Górecka-Bruzda A, Nowicki J, Gumulka M, Molik E, Schwarz T, Earley B, Klocek C (2015) Perception of environment in farm animals – a review. Annals of Animal Science 15, 565-589.
| Crossref | Google Scholar |
Ames DR, Arehart LA (1972) Physiological response of lambs to auditory stimuli. Journal of Animal Science 34, 994-998.
| Crossref | Google Scholar | PubMed |
Andersen M, Aars J (2008) Short-term behavioural response of polar bears (Ursus maritimus) to snowmobile disturbance. Polar Biology 31, 501-507.
| Crossref | Google Scholar |
Arnold NA, Ng KT, Jongman EC, Hemsworth PH (2008) Avoidance of tape-recorded milking facility noise by dairy heifers in a Y maze choice task. Applied Animal Behaviour Science 109, 201-210.
| Crossref | Google Scholar |
Atkinson L, Doyle RE, Woodward A, Jongman EC (2022) Behavioural reactivity testing in sheep indicates the presence of multiple temperament traits. Behavioural Processes 201, 104711.
| Crossref | Google Scholar |
Barber JR, Crooks KR, Fristrup KM (2010) The costs of chronic noise exposure for terrestrial organisms. Trends in Ecology & Evolution 25, 180-189.
| Crossref | Google Scholar | PubMed |
Boissy A, Terlouw C, Le Neindre P (1998) Presence of cues from stressed conspecifics increases reactivity to aversive events in cattle: evidence for the existence of alarm substances in urine. Physiology & Behavior 63, 489-495.
| Crossref | Google Scholar | PubMed |
Borg E (1981) Physiological and pathogenic effects of sound. Acta Oto-Laryngologica 92, 1-64.
| Crossref | Google Scholar |
Brouček J (2014) Effect of noise on performance, stress, and behaviour of animals. Slovak Journal of Animal Science 47, 111-123.
| Google Scholar |
Broucek J, Kovalcikova M, Kovalcik K (1983) The effect of noise on the biochemical characteristics of blood in dairy cows. Zivocisna Vyroba 28, 261-267.
| Google Scholar |
Brown CL, Hardy AR, Barber JR, Fristrup KM, Crooks KR, Angeloni LM (2012) The effect of human activities and their associated noise on ungulate behavior. PLoS ONE 7, e40505.
| Crossref | Google Scholar | PubMed |
Campo JL, Gil MG, Dávila SG (2005) Effects of specific noise and music stimuli on stress and fear levels of laying hens of several breeds. Applied Animal Behaviour Science 91, 75-84.
| Crossref | Google Scholar |
Chloupek P, Voslarova E, Chloupek J, Bedanova I, Pistekova V, Vecerek V (2009) Stress in broiler chickens due to acute noise exposure. Acta Veterinaria Brno 78, 93-98.
| Crossref | Google Scholar |
Cook CJ (1997) Oxytocin and prolactin suppress cortisol responses to acute stress in both lactating and non-lactating sheep. Journal of Dairy Research 64, 327-339.
| Crossref | Google Scholar | PubMed |
Coppinger TR, Minton JE, Reddy PG, Blecha F (1991) Repeated restraint and isolation stress in lambs increases pituitary-adrenal secretions and reduces cell-mediated immunity. Journal of Animal Science 69, 2808-2814.
| Crossref | Google Scholar | PubMed |
Croy I, Smith MG, Waye KP (2013) Effects of train noise and vibration on human heart rate during sleep: an experimental study. BMJ Open 3, e002655.
| Crossref | Google Scholar |
De la Fuente J, Díaz MT, Ibáñez M, De Chavarri EG (2007) Physiological response of rabbits to heat, cold, noise and mixing in the context of transport. Animal Welfare 16, 41-47.
| Crossref | Google Scholar |
De Palo P, Maggiolino A, Albenzio M, Caroprese M, Centoducati P, Tateo A (2018) Evaluation of different habituation protocols for training dairy jennies to the milking parlor: effect on milk yield, behavior, heart rate and salivary cortisol. Applied Animal Behaviour Science 204, 72-80.
| Crossref | Google Scholar |
Destrez A, Deiss V, Lévy F, Calandreau L, Lee C, Chaillou-Sagon E, Boissy A (2013) Chronic stress induces pessimistic-like judgment and learning deficits in sheep. Applied Animal Behaviour Science 148, 28-36.
| Crossref | Google Scholar |
Désiré L, Veissier I, Després G, Delval E, Toporenko G, Boissy A (2006) Appraisal process in sheep (Ovis aries): interactive effect of suddenness and unfamiliarity on cardiac and behavioral responses. Journal of Comparative Psychology 120, 280-287.
| Crossref | Google Scholar | PubMed |
Dobson H, Fergani C, Routly JE, Smith RF (2012) Effects of stress on reproduction in ewes. Animal Reproduction Science 130, 135-140.
| Crossref | Google Scholar | PubMed |
Dodd CL, Pitchford WS, Hocking Edwards JE, Hazel SJ (2012) Measures of behavioural reactivity and their relationships with production traits in sheep: a review. Applied Animal Behaviour Science 140, 1-15.
| Crossref | Google Scholar |
Doggett F (2018) Minimizing animal stress due to facility noise and vibration. Laboratory Equipment, South San Francisco (5 December 2018). Available at https://login.ezproxy.une.edu.au/login?url=https://www.proquest.com/trade-journals/minimizing-animal-stress-due-facility-noise/docview/2153506842/se-2
Doyle RE, Freire R, Cowling A, Knott SA, Lee C (2014) Performance of sheep in a spatial maze is impeded by negative stimuli. Applied Animal Behaviour Science 151, 36-42.
| Crossref | Google Scholar |
Dumont B, Boissy A (2000) Grazing behaviour of sheep in a situation of conflict between feeding and social motivations. Behavioural Processes 49(3), 131-138.
| Crossref | Google Scholar |
Edes AN, Wolfe BA, Crews DE (2018) Evaluating allostatic load: a new approach to measuring long-term stress in wildlife. Journal of Zoo and Wildlife Medicine 49, 272-282.
| Crossref | Google Scholar | PubMed |
Ehnert K, Moberg GP (1991) Disruption of estrous behavior in ewes by dexamethasone or management-related stress. Journal of Animal Science 69, 2988-2994.
| Crossref | Google Scholar | PubMed |
Fordham DP, Al-Gahtani S, Durotoye LA, Rodway RG (1991) Changes in plasma cortisol and β-endorphin concentrations and behaviour in sheep subjected to a change of environment. Animal Science 52, 287-296.
| Crossref | Google Scholar |
Francis CD, Barber JR (2013) A framework for understanding noise impacts on wildlife: an urgent conservation priority. Frontiers in Ecology and the Environment 11, 305-313.
| Crossref | Google Scholar |
Garcia DB, Silva IJO, Barbosa Filho JAD, Vieira FMC, Dias CTS (2009) Evaluation of the effect of vibration in simulated condition of transport of broiler chickens. In ‘Livestock Environment VIII’, 31 August – 4 September 2008, Iguassu Falls, Brazil. (American Society of Agricultural and Biological Engineers)
Grandin T (1997) Assessment of stress during handling and transport. Journal of Animal Science 75, 249-257.
| Crossref | Google Scholar |
Greiveldinger L, Veissier I, Boissy A (2007) Emotional experience in sheep: predictability of a sudden event lowers subsequent emotional responses. Physiology & Behavior 92, 675-683.
| Crossref | Google Scholar |
Griffiths SK, Pierson LL, Gerhardt KJ, Abrams RM, Peters AJM (1994) Noise induced hearing loss in fetal sheep. Hearing Research 74, 221-230.
| Crossref | Google Scholar | PubMed |
Grissom N, Bhatnagar S (2009) Habituation to repeated stress: get used to it. Neurobiology of Learning and Memory 92, 215-224.
| Crossref | Google Scholar | PubMed |
Guinn JE (2013) Generational habituation and current bald eagle populations. Human–Wildlife Interactions 7, 69-76.
| Google Scholar |
Gutschalk A, Micheyl C, Oxenham AJ (2008) Neural correlates of auditory perceptual awareness under informational masking. PLoS Biology 6, e138.
| Crossref | Google Scholar | PubMed |
Hauser R, Wechsler B (2013) Assessment of noise exposure in sheep. Schweiz Arch Tierheilkd 155, 129-134.
| Crossref | Google Scholar | PubMed |
Healy K, McNally L, Ruxton GD, Cooper N, Jackson AL (2013) Metabolic rate and body size are linked with perception of temporal information. Animal Behaviour 86, 685-696.
| Crossref | Google Scholar | PubMed |
Heffner HE (1998) Auditory awareness. Applied Animal Behaviour Science 57, 259-268.
| Crossref | Google Scholar |
Hopewell L, Rossiter R, Blower E, Leaver L, Goto K (2005) Grazing and vigilance by Soay sheep on Lundy island: influence of group size, terrain and the distribution of vegetation. Behavioural Processes 70, 186-193.
| Crossref | Google Scholar | PubMed |
James D (2020) The benefits and costs of installing a cow underpass. Farmers Weekly 173, 26-27.
| Google Scholar |
Kanitz E, Otten W, Tuchscherer M (2005) Central and peripheral effects of repeated noise stress on hypothalamic–pituitary–adrenocortical axis in pigs. Livestock Production Science 94, 213-224.
| Crossref | Google Scholar |
Karwowska M, Mikołajczak J, Borowski S, Dolatowski ZJ, Marć-Pieńkowska J, Budziński W (2014) Effect of noise generated by the wind turbine on the quality of goose muscles and abdominal fat. Annals of Animal Science 14, 441-451.
| Crossref | Google Scholar |
Karwowska M, Mikolajczak J, Dolatowski ZJ, Borowski S (2015) The effect of varying distances from the wind turbine on meat quality of growing-finishing pigs. Annals of Animal Science 15, 1043-1054.
| Crossref | Google Scholar |
Kearton T, Marini D, Cowley F, Belson S, Lee C (2019) The effect of virtual fencing stimuli on stress responses and behavior in sheep. Animals 9, 30.
| Crossref | Google Scholar | PubMed |
Kearton T, Marini D, Cowley F, Belson S, Keshavarzi H, Mayes B, Lee C (2020) The influence of predictability and controllability on stress responses to the aversive component of a virtual fence. Frontiers in Veterinary Science 7, 580523.
| Crossref | Google Scholar |
Kight CR, Swaddle JP (2011) How and why environmental noise impacts animals: an integrative, mechanistic review. Ecology Letters 14, 1052-1061.
| Crossref | Google Scholar | PubMed |
Kim SJ, Chai SK, Lee KW, Park J-B, Min K-B, Kil HG, Lee C, Lee KJ (2014) Exposure-response relationship between aircraft noise and sleep quality: a community-based cross-sectional study. Osong Public Health and Research Perspectives 5, 108-114.
| Crossref | Google Scholar | PubMed |
Kovalcik K, Sottnik J (1971) Effect of noise on the milking efficiency of cows. [Vplyvhluku na mliekovu uzitkovost krav.]. Zivocisna Vyroba 16, 795-804.
| Google Scholar |
Lanier JL, Grandin T, Green RD, Avery D, McGee K (2000) The relationship between reaction to sudden, intermittent movements and sounds and temperament. Journal of Animal Science 78, 1467-1474.
| Crossref | Google Scholar | PubMed |
Lian X, Zhang T, Cao Y, Su J, Thirgood S (2007) Group size effects on foraging and vigilance in migratory Tibetan antelope. Behavioural Processes 76, 192-197.
| Crossref | Google Scholar | PubMed |
Lian X, Zhang T, Cao Y, Su J, Thirgood S (2011) Road proximity and traffic flow perceived as potential predation risks: evidence from the Tibetan antelope in the Kekexili National Nature Reserve, China. Wildlife Research 38, 141.
| Crossref | Google Scholar |
Little SJ, Harcourt RG, Clevenger AP (2002) Do wildlife passages act as prey-traps? Biological Conservation 107, 135-145.
| Crossref | Google Scholar |
Lowrey C, Longshore KM (2017) Tolerance to disturbance regulated by attractiveness of resources: a case study of desert bighorn sheep within the river mountains, Nevada. Western North American Naturalist 77, 82-98.
| Crossref | Google Scholar |
Martinig AR, Riaz M, St. Clair CC (2020) Temporal clustering of prey in wildlife passages provides no evidence of a prey-trap. Scientific Reports 10, 11489.
| Crossref | Google Scholar |
Mcdougall PL, Ruckstuhl KE (2018) Vigilance behaviour is more contagious when chewing stops: examining the characteristics of contagious vigilance in bighorn sheep. Behavioral Ecology and Sociobiology 72, 143.
| Crossref | Google Scholar |
Mora-Medina P, Orihuela-Trujillo A, Arch-Tirado E, Roldan-Santiago P, Terrazas A, Mota-Rojas D (2016) Sensory factors involved in mother-young bonding in sheep: a review. Veterinární Medicína 61, 595-611.
| Crossref | Google Scholar |
Morton CL, Hinch G, Small A, McDonald PG (2018) Flawed mothering or infant signaling? The effects of deficient acoustic cues on ovine maternal response. Developmental Psychobiology 60, 975-988.
| Crossref | Google Scholar | PubMed |
Muhly TB, Alexander M, Boyce MS, Creasey R, Hebblewhite M, Paton D, Pitt JA, Musiani M (2010) Differential risk effects of wolves on wild versus domestic prey have consequences for conservation. Oikos 119, 1243-1254.
| Crossref | Google Scholar |
Nowak R (1991) Senses involved in discrimination of merino ewes at close contact and from a distance by their newborn lambs. Animal Behaviour 42(3), 357-366.
| Crossref | Google Scholar |
Otten W, Kanitz E, Puppe B, Tuchscherer M, Brüssow KP, Nürnberg G, Stabenow B (2004) Acute and long term effects of chronic intermittent noise stress on hypothalamic–pituitary–adrenocortical and sympatho-adrenomedullary axis in pigs. Animal Science 78, 271-283.
| Crossref | Google Scholar |
Owen JE, Veasey SC (2020) Impact of sleep disturbances on neurodegeneration: insight from studies in animal models. Neurobiology of Disease 139, 104820.
| Crossref | Google Scholar | PubMed |
Padilla de la Torre M, McElligott AG (2017) Vocal communication and the importance of mother-offspring relations in cattle. Animal Behavior and Cognition 4, 522-525.
| Crossref | Google Scholar |
Papouchis CM, Singer FJ, Sloan WB (2001) Responses of desert bighorn sheep to increased human recreation. The Journal of Wildlife Management 65, 573-582.
| Crossref | Google Scholar |
Perentos N, Martins AQ, Cumming RJM, Mitchell NL, Palmer DN, Sawiak SJ, Morton AJ (2016) An EEG investigation of sleep homeostasis in healthy and CLN5 Batten disease affected sheep. Journal of Neuroscience 36, 8238-8249.
| Crossref | Google Scholar | PubMed |
Quaranta A, Sevi A, Nardomarino A, Colella GE, Casamassima D (2002) Effects of graded noise levels on behavior, physiology and production performance of intensively managed lambs. Italian Journal of Animal Science 1, 217-227.
| Crossref | Google Scholar |
Rabat A (2008) Extra-auditory effects of noise in laboratory animals: the relationship between noise and sleep. Journal of the American Association for Laboratory Animal Science 46, 35-41.
| Google Scholar |
Rabat A, Bouyer JJ, Aran JM, Courtiere A, Mayo W, Le Moal M (2004) Deleterious effects of an environmental noise on sleep and contribution of its physical components in a rat model. Brain Research 1009, 88-97.
| Crossref | Google Scholar | PubMed |
Rabey KN, Li Y, Norton JN, Reynolds RP, Schmitt D (2015) Vibrating frequency thresholds in mice and rats: implications for the effects of vibrations on animal health. Annals of Biomedical Engineering 43, 1957-1964.
| Crossref | Google Scholar | PubMed |
Randall JM, Duggan JA, Alami MA, White RP (1997) Frequency weightings for the aversion of broiler chickens to horizontal and vertical vibration. Journal of Agricultural Engineering Research 68, 387-397.
| Crossref | Google Scholar |
Refshauge G, Brien FD, Hinch GN, van de Ven R (2016) Neonatal lamb mortality: factors associated with the death of Australian lambs. Animal Production Science 56, 726-735.
| Crossref | Google Scholar |
Rosa P, Koper N (2018) Integrating multiple disciplines to understand effects of anthropogenic noise on animal communication. Ecosphere 9, e02127.
| Crossref | Google Scholar |
Rylander R, Sörensen S, Andrae BO, Chatelier G, Espmark Y, Larsson T, Thackray RI (1974) Sonic boom exposure effects – a field study on humans and animals. Journal of Sound and Vibration 33, 471-486.
| Crossref | Google Scholar |
Salvin H, Cafe L, Lees A, Morris S, Lee C (2020) A novel protocol to measure startle magnitude in sheep. Applied Animal Behaviour Science 228, 104996.
| Crossref | Google Scholar |
Schell CJ, Young JK, Lonsdorf EV, Santymire RM, Mateo JM (2018) Parental habituation to human disturbance over time reduces fear of humans in coyote offspring. Ecology and Evolution 8, 12965-12980.
| Crossref | Google Scholar | PubMed |
Schneider WT, Vas S, Nicol AU, Morton AJ (2020) Characterizing sleep spindles in sheep. eNeuro 7, ENEURO.0410-19.
| Crossref | Google Scholar |
Sèbe F, Nowak R, Poindron P, Aubin T (2007) Establishment of vocal communication and discrimination between ewes and their lamb in the first two days after parturition. Developmental Psychobiology 49, 375-386.
| Crossref | Google Scholar | PubMed |
Sheng X, Jones CJC, Petyt M (1999) Ground vibration generated by a harmonic load acting on a railway track. Journal of Sound and Vibration 225, 3-28.
| Crossref | Google Scholar |
Shillito E, Alexander G (1975) Mutual recognition amongst ewes and lambs of four breeds of sheep (Ovis aries). Applied Animal Ethology 1(2), 151-165.
| Crossref | Google Scholar |
Sibbald AM, Erhard HW, McLeod JE, Hooper RJ (2009) Individual personality and the spatial distribution of groups of grazing animals: an example with sheep. Behavioural Processes 82(3), 319-226.
| Crossref | Google Scholar |
Smith MG, Croy I, Hammar O, Persson Waye K (2016) Vibration from freight trains fragments sleep: a polysomnographic study. Scientific Reports 6, 24717.
| Crossref | Google Scholar | PubMed |
Stockwell CA (1991) Behavioural reactions of desert bighorn sheep to avian scavengers. Journal of Zoology 225, 563-566.
| Crossref | Google Scholar |
Tamioso PR, Boissy A, Boivin X, Chandeze H, Andanson S, Delval E, Hazard D, Taconeli CA, Molento CFM (2017) Does emotional reactivity influence behavioral and cardiac responses of ewes submitted to brushing? Behavioural Processes
| Google Scholar |
Trigg J, Naweed A, Kinnear S (2023) A scoping review of freight rail noise and vibration impacts on domestic animal health and welfare. Animal Welfare 31, 69-77.
| Crossref | Google Scholar |
Turner JG, Parrish JL, Hughes LF, Toth LA, Caspary DM (2005) Hearing in laboratory animals: strain differences and nonauditory effects of noise. Comparative Medicine 55, 12-23.
| Google Scholar | PubMed |
van Der Staay FJ, Joosse M, van Dijk H, Schuurman T, van Der Meulen J (2011) Physiological and behavioral reactions elicited by simulated and real-life visual and acoustic helicopter stimuli in dairy goats. BMC Veterinary Research 7, 16.
| Crossref | Google Scholar | PubMed |
van Lier E, Hart KW, Viñoles C, Paganoni B, Blache D (2017) Calm Merino ewes have a higher ovulation rate and more multiple pregnancies than nervous ewes. Animal 11, 1196-1202.
| Crossref | Google Scholar | PubMed |
Verbeek E, Colditz I, Blache D, Lee C (2019) Chronic stress influences attentional and judgement bias and the activity of the HPA axis in sheep. PLoS ONE 14, e0211363.
| Crossref | Google Scholar | PubMed |
Voslarova E, Chloupek P, Chloupek J, Bedanova I, Pistekova V, Vecerek V (2011) The effects of chronic intermittent noise exposure on broiler chicken performance. Animal Science Journal (Japan) 82, 601-606.
| Crossref | Google Scholar |
Waynert DF, Stookey JM, Schwartzkopf-Genswein KS, Watts JM, Waltz CS (1999) The response of beef cattle to noise during handling. Applied Animal Behaviour Science 62, 27-42.
| Crossref | Google Scholar |
Weeks CA (2008) A review of welfare in cattle, sheep and pig lairages, with emphasis on stocking rates, ventilation and noise. Animal Welfare 17, 275-284.
| Crossref | Google Scholar |
Weiss JM (1972) Psychological factors in stress and disease. Scientific American 226, 104-113.
| Crossref | Google Scholar | PubMed |