

Temporal variations in leaf traits, chemical composition and in vitro true digestibility of four temperate fodder tree species
Simone Ravetto Enri A , Massimiliano Probo B , Manuela Renna C D , Eleonora Caro A , Carola Lussiana A , Luca M. Battaglini A , Giampiero Lombardi A and Michele Lonati AA Dipartimento di Scienze Agrarie, Forestali e Alimentari, University of Torino, Largo Braccini 2 - 10095, Grugliasco (TO), Italy.
B Agroscope, Grazing Systems, Nyon 1, 1260, Switzerland.
C Dipartimento di Scienze Veterinarie, University of Torino, Largo Braccini 2 - 10095, Grugliasco (TO), Italy.
D Corresponding author. Email: manuela.renna@unito.it
Animal Production Science 60(5) 643-658 https://doi.org/10.1071/AN18771
Submitted: 17 December 2018 Accepted: 14 August 2019 Published: 31 January 2020
Journal compilation © CSIRO 2020 Open Access CC BY-NC-ND
Abstract
Context: Many tree and shrub species are underestimated fodder resources due to insufficient knowledge about their potential feeding value, especially for goats.
Aims: The present work aimed at assessing productive and nutritional attributes of the foliage of the following four temperate tree species widespread in Europe: Acer pseudoplatanus, Fraxinus excelsior, Salix caprea and Sorbus aucuparia.
Methods: Leaf length and biomass, proximate composition, fatty acid profile, phenolic composition and in vitro true dry matter digestibility were determined along the vegetative season.
Key results: The leaf length of the four species was significantly related to leaf biomass and can be considered as a proficient proxy for estimating leaf biomass. The differences found among the species were remarkable, although weakly related to temporal changes, especially when considering fatty acid and phenolic compositions. Fraxinus excelsior sprouts were the most productive, with a mean biomass of 13.2 g dry matter (DM) per sprout at the end of the growing season. Its foliage showed also the lowest phenolic concentrations (average total extractable phenols of 11.25 g/kg DM), resulting in the highest digestibility values (average in vitro true dry matter digestibility of 56.5 g/kg DM). Digestibility of S. aucuparia was similar, but its lower polyunsaturated fatty acid concentration (average value of 62.13 g/kg DM) could reduce the interest for this species as a feeding resource for goat dairy products with healthy properties. The lower digestibility found for A. pseudoplatanus and S. caprea (average values of 43.3 and 46.2 g/kg DM, respectively) may be related to their higher phenolic concentrations (average total extractable phenols of 45.9 and 47.3 g/kg DM, respectively).
Conclusions: The four species could represent an appealing feedstuff for goat nutrition, due to the valuable and complementary nutritional characteristics of their foliage.
Implications: The use of the studied species as fodder resource may be particularly relevant during drought periods and in the late summer when herbage quality decreases, especially in terms of crude protein and fatty acid profile.
Additional keywords: fatty acids, feed quality, goats, in vitro digestibility, tannins.
Introduction
Tree and shrub foliage is an important component of small ruminant diet in many parts of the world. It plays an essential role for browsing animals, especially where livestock systems are based on rangeland and grazable forestland exploitation (Leng 1997). Due to their deep root system, trees can maintain the high nutrient quality of their foliage also in dry periods, when herbage quality decreases (Papachristou and Papanastasis 1994). For this reason, the interest for fodder tree species is rising also in European temperate regions where drought periods are increasing in length and frequency over large areas due to global warming (Vandermeulen et al. 2018a). In this context, the so called ‘silvopastoral systems’, integrating fodder trees and grasslands, are spreading as a possible solution to face climate changes and to support a sustainable and resilient agriculture, especially in marginal areas (Novak 2014; Seidavi et al. 2018).
The knowledge of the importance of tree foliage for ruminant nutrition has led to several studies aimed at evaluating leaf forage productivity, chemical composition and digestibility (e.g. Boufennara et al. 2012). Proximate and fatty acid (FA) composition, phenolic concentration and digestibility have been largely assessed for various fodder tree species in several environments such as savanna (Tefera et al. 2008), tropics (Arigbede et al. 2012), subtropics (Prakash et al. 2009), American temperate region (Ricklefs and Matthew 1982), Mediterranean region (Kökten et al. 2012) and non-European alpine areas (Khanal and Subba 2001).
According to these and other studies, tree and shrub foliage is increasingly recognised as a high-quality feedstuff for small ruminants, either offered as sole feed or as dietary supplement (Leng 1997). Tree foliage has a superior mineral composition compared with that of some grasses (e.g. dry grasses), making foliage a significant source of minerals for the animals and rumen microbiota (Leng 1997). Some foliage has high protein contents and is, therefore, used as protein source to supplement other feeds, particularly when ruminants are offered roughages of a low nutritive value (Simbaya 2002; Kongmanila 2012). Tree and shrub foliage contains endogenous secondary compounds able to bind with proteins and protect them from microbial attack in the rumen afterwards. For this reason, foliage can represent a source of by-pass protein for the ruminant that, escaping rumen degradation, can be digested in the intestine, enhancing the animal protein status (Leng 1997). Plant secondary compounds are also able to alter the balance of microorganisms in the rumen (Vasta and Bessa 2012). Therefore, besides variations in FA, differences in concentrations of plant secondary compounds may also confer specific intrinsic sensory and chemical attributes to ruminant-derived dairy and meat products (Iussig et al. 2015a). Some plant secondary compounds contained in a large number of fodder tree species, such as phenolic compounds, are also characterised by potential anti-nutritional effects (Waghorn 2008). Among phenolic compounds, tannins can cause either adverse or beneficial effects on nutrient utilisation, health and animal production, in relation to their molecular characteristics and concentrations (Min et al. 2003). Particularly, tannins (depending on the type, chemical characteristics and amount ingested) can (1) increase intestinal escape protein availability, (2) reduce methane production, (3) defend against bloat and (4) wane gastrointestinal parasites (Mueller-Harvey 2006; Vandermeulen et al. 2018a). Francisco et al. (2015) reported also that inclusion of low concentrations (up to 16.3 g/kg DM) of condensed tannins (CT) in lamb diet could wax antioxidant properties in meat, improving its stability after storage. Similarly, milk from goats fed with low concentrations of hydrolysable tannins (HT) showed an increasing trend of conjugated linoleic acid isomers and oleic acid, as a result of an enhancement of Δ9-desaturase activity (Abo-Donia et al. 2017).
Temporal variations have also been analysed, since tree foliage productivity, chemical composition and digestibility can significantly change due to phenological advancement and climatic variations (Papachristou and Papanastasis 1994; Ndondo et al. 2004; Kökten et al. 2012).
In the last decades, extensive goat rearing has spread in European alpine areas, particularly in marginal areas (Battaglini et al. 2014; Álvarez‐Martínez et al. 2016). Here, due to their increasing presence and resistance to browsing and drought, extensive shrublands and small trees can provide abundant leaf fodder, which represents the basic component of goat diet. Particularly, a general increase of tree leaf consumption has been observed in late summer, when the availability and nutritive value of herbage decrease (Castro and Fernández Núñez 2016). Moreover, goats are well adapted to exploit low-quality forages due to their ruminal bacterial population, suitable for degradation of highly lignified material and resistance to tannin toxicity, resulting in an increased tree foliage digestibility (Manousidis et al. 2016).
Various research has been conducted on European temperate tree species (see, among others, Luske and van Eekeren 2015; Emile et al. 2016; Vandermeulen et al. 2018b). These plants have been a widespread feeding resource since Neolithic (Gómez García and Fillat 1984; Kühn et al. 2013; Hejcman et al. 2014), especially for goat nutrition (Leng 1997; Castro and Fernández Núñez 2016). However, the leaf biomass production of fodder tree species in these environments is poorly documented. Moreover, only a limited number of authors experienced the assessment of foliage production using allometric equations based on leaf traits (Forrester et al. 2017). In addition, there is a lack of knowledge on the FA profile of such tree foliage, even if this feature is of increasing interest for the possible effects on the nutraceutical properties of the derived dairy and meat products (Glasser et al. 2013).
The present study aimed to fill the gap of knowledge about the above-mentioned topics. For this purpose, we selected, in the western Italian Alps, the following four temperate tree species that are widespread on European uplands (San-Miguel-Ayanz et al. 2016): Acer pseudoplatanus L. (sycamore maple), Fraxinus excelsior L. (ash), Salix caprea L. (goat willow) and Sorbus aucuparia L. (rowan). These trees are among the browses most preferred and used by goats, either directly by browsing or by being fed after cutting (Guinier 1950; Thiebault 2005; Iussig et al. 2015a, 2015b). The objective of the work was to characterise their leaves across the whole vegetative season in terms of leaf traits (i.e. leaf length and biomass), proximate composition (i.e. dry matter (DM), ash, crude protein (CP), ether extract (EE), and fibre fractions), FA profile, phenolic compounds (i.e. total extractable phenols (TEP), non-tannin phenols (NTP), total tannins (TT; CT and HT) and in vitro true DM digestibility (IVTD).
Materials and methods
Study area
The study was conducted at Oasi Zegna, located in the Piedmont Region, north-western Italy (45°40′N, 8°09′E), within the boundaries of Valle Sessera Site of Community Interest (SCI IT11300002). The study area is characterised by a suboceanic climate (Köppen’s classification: Cfb), with annual mean temperature of 7.3°C and precipitation of 1700 mm (mean value for the years 2002–2015 of the pluviometric station of Bielmonte). Dominant soils have originated from siliceous parent rock. In the study area, tree stands are mixed broadleaved populations, dominated by Fagus sylvatica L. (European beech) and Betula pendula Roth (silver birch), currently managed by occasional selective cutting.
Experimental design
For each investigated species (i.e. A. pseudoplatanus, F. excelsior, S. caprea and S. aucuparia), a group of four trees was selected in December 2014, during the vegetative dormancy, in a 500 × 150 m2 area with homogeneous exposure (north–north-west) and elevation (1270–1320 m asl). Within each species group, the trees were located at a maximum distance of 125 m from each other and had a similar height, stem diameter, age, and no disease evidence. The selected trees were protected from animal browsing by using deer fences during the whole trial.
Leaf traits (i.e. leaf length and biomass) were recorded 12 times, while leaf sampling for laboratory analyses (i.e. chemical composition and IVTD) occurred four times during the trial, as detailed in Table 1.
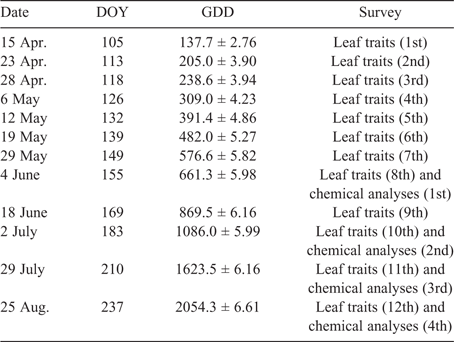
|
Leaf length and biomass
Four sprouts at bud stage per tree were selected at a maximum height of 1.80 m (i.e. as far as goats can browse buds and leaves, assuming a bipedal stance; Iussig et al. 2015b). Each sprout was located in a different cardinal direction to avoid differences due to light exposure. The elongation of emerging leaves was monitored on each sprout, from the budburst of the earliest sprout until all trees reached the maximum vegetative phenological stage, according to the extended BBCH scale (Hack et al. 1992). At each survey date, following the survey schedule provided in Table 1, the phenological stage (vegetative and, when present, reproductive) of each individual was recorded. In addition, one sprout, comparable to the monitored one in terms of leaf number and development, was harvested and transported to the laboratory. The emitted leaves were then separated from the sprout and weighed. Leaf traits (i.e. leaf length and biomass) were computed by cumulating the values of all unfolded leaves per sprout in each survey date, accounting for a total of 16 replicates per species (four trees × four sprouts).
Growing degree-days
Four HOBO data loggers (Onset Corporation, Pocasset, MA, USA), placed near the centroid of each species group, recorded air temperature every 30 min for the whole trial. Heat units from 1 January 2015 to the end of the trial, expressed as cumulative growing degree-days (GDD), were calculated for each species group from daily air temperature, by cumulating all mean daily temperatures of >5°C (Prislan et al. 2013). For each sprout, the GDD corresponding to the 25%, 50%, 75% and 100% of the final leaf length and biomass were assessed by data interpolation.
Leaf sampling
Since all the selected sprouts ended leaf emission (i.e. 4 June 2015), one leaf sample per tree (i.e. a total of four replicates per species) was collected for four dates from June to August, as detailed in Table 1. Such time interval approximately corresponds to the goat browsing season in these grazable forestlands. A quantity of ~400 g of fresh leaves (including petioles and rachises; Hejcmanová et al. 2014) was harvested all around the canopy, bottom-up to 1.80-m height, so as to simulate goat browsing (Iussig et al. 2015b). All leaves damaged by pathogens (insects, fungi) were avoided. The samples were placed in sealed polyethylene bags, immediately stored at 4°C in a portable refrigerator and transported to the laboratory. In the laboratory, each sample was divided into two homogeneous aliquots of ~200 g each. The samples were freeze-dried (Freeze Drying Equipments, Criofarma, Torino, Italy), grounded with a cutting mill to pass through a 1-mm-screen sieve (Pulverisette 15, Fritsch GmbH, Idar-Oberstein, Germany) and used for further laboratory analyses.
Proximate composition
AOAC (2000) procedures were used to determine DM (Method no. 930.15), ash (Method no. 942.05), CP (Method no. 984.13), acid detergent fibre (ADF) and acid detergent lignin (ADL; Method no. 973.18). The EE was determined following Method no. 920.39 of AOAC (2003). Neutral detergent fibre (NDF) was analysed according to Van Soest et al. (1991); α-amylase (Sigma Aldrich, Saint Louis, MO, USA), but no sodium sulfite was added and results were corrected for residual ash content. The proximate composition was expressed as g/kg DM.
Fatty acid composition
The FA composition was assessed using a combined direct transesterification and solid-phase extraction method, as described by Alves et al. (2008). The FA methyl esters were separated, identified and quantified as detailed by Renna et al. (2014). The FA composition was expressed as g/kg DM.
Phenolic composition
The TEP and phenol fractions (NTP and CT) were determined using standard protocols, as detailed in Iussig et al. (2015a). The absorbance was recorded at 725 nm (TEP and NTP, expressed as gallic acid equivalents) and 550 nm (CT, expressed as leucocyanidin equivalents) using a UV–VIS spectrophotometer (Shimadzu UVmini-1240, Shimadzu Corporation, Kyoto, Japan). The TT were computed as the difference between TEP and NTP. The HT were estimated as the difference between TT and CT. The phenolic composition was expressed as g/kg DM.
In vitro true dry matter digestibility
Four slaughtered adult male Alpine goats were used as rumen fluid donors. The male goats belonged to an extensive dairy goat farm located in Rueglio (Torino, Italy). They were fed on alpine grazable forestlands rich in fodder tree and shrub species, including those under investigation in the present trial. Slaughtering procedures followed Council Regulation (EC) No. 1099/2009 of the European Union. They occurred four times (once per each male goat), every three weeks between October and December 2015. A representative sample of the rumen contents was collected from each animal into a thermos container, which was kept at 39°C using a warm water-containing cooler, and immediately transported to the laboratory, where the sample was filtered through four layers of cheesecloth under continuous flush of CO2 at 39°C. The IVTD was determined according to the ANKOM Daisy procedure (ANKOM 2017). Following the slaughtering time schedule, four runs were performed. For each run, all 64 leaf samples were digested using the rumen fluid of one goat, accounting for a total of eight replicates per plant species and sampling date (two trees × four rumen fluids).
Leaf samples (0.25 g) were weighed into filter bags (ANKOM® #F57; pore size 25 μm, ANKOM Technology Corporation, Fairport, NY, USA), which were then heat-sealed. Rumen fluid was diluted into the buffer medium in a proportion of 1 : 4 (v/v). Thereafter, a volume of 2 L of buffered rumen fluid was transferred into four 5-L jars at 39°C under anaerobic conditions. Each jar, containing leaf samples and one blank, was placed in a revolving incubator (ANKOM DaisyII digestion system, ANKOM Technology Corporation) at 39°C for 48 h, under continuous rotation. After incubation, the samples were rinsed with cold water and subjected to an extraction with NDF solution at 100°C for 1 h, so as to remove microbial debris and any remaining endogenous products.
Statistical analyses
To evaluate species precocity in leaf development at different phenological stages, one-way ANOVA was used to analyse for differences among the GDD corresponding to budburst and to 25%, 50%, 75% and 100% of the final leaf trait values. Moreover, to test for differences of final leaf length and biomass values among species at the end of the growing season, a one-way ANOVA was performed. For both analyses, the sprout was considered as the statistical unit and tree species as a fixed factor.
The relationship between leaf length and biomass was then explored with linear regressions, using a separate dataset for each species. The regression equations were based on the best-fitting model and the sprout was considered as the statistical unit.
General linear models accounting for repeated measures were performed on proximate composition, FA profile (namely, total FA (TFA), groups of FA, and main represented individual FA), phenolic composition, and IVTD of each tree species. Tree was considered as the statistical unit, species as a fixed factor, and sampling date as a repeated measure.
To test for differences among species during the vegetative season, a one-way ANOVA was performed on the same variables (i.e. proximate composition, FA profile, phenolic composition and IVTD) for each of the four sampling dates, using tree as the statistical unit and tree species as the fixed factor. Additionally, the temporal variations in terms of proximate composition, FA, phenolic composition and IVTD within the same species were tested performing a one-way ANOVA, with tree as the statistical unit and sampling date as a fixed factor.
Assumptions of normality and homogeneity of variance were checked with Shapiro–Wilk and Levene’s tests respectively. Variables that were not normally distributed were log-transformed before further statistical analysis; results are presented as non-transformed data. When normal distribution or homogeneity of variances were not met, even after log-transformation, the non-parametric Kruskal–Wallis test or Welch one-way ANOVA were used, respectively. When significant differences were found, Tukey’s, Steel’s and Tamhane’s post hoc tests were performed for ANOVA, Kruskal–Wallis and Welch one-way ANOVA, respectively.
All statistical analyses were performed using SPSS (Version 24.0, IBM SPSS Statistics, Armonk, NY, USA). Significance was set at P < 0.05.
Results
Leaf traits
A total of 479 leaves was monitored during the trial. Mean number of emitted leaves per sprout was eight for A. pseudoplatanus and S. caprea, nine for F. excelsior and five for S. aucuparia.
The vegetative season started at 135 GDD (105th day of the year), with S. caprea unfolding of the first leaves (Table S1, available as Supplementary material to this paper). Budburst occurred first (at a lower GDD) for S. caprea and S. aucuparia, followed by F. excelsior and then by A. pseudoplatanus (Table 2). Salix caprea and S. aucuparia developed earlier in terms of leaf length and biomass for most of the considered stages, followed by F. excelsior and, then, by A. pseudoplatanus.

|
Differences among tree species in terms of final leaf traits were similar between leaf length and leaf biomass, with F. excelsior showing the greatest values, followed by A. pseudoplatanus, S. aucuparia and S. caprea (Fig. 1).
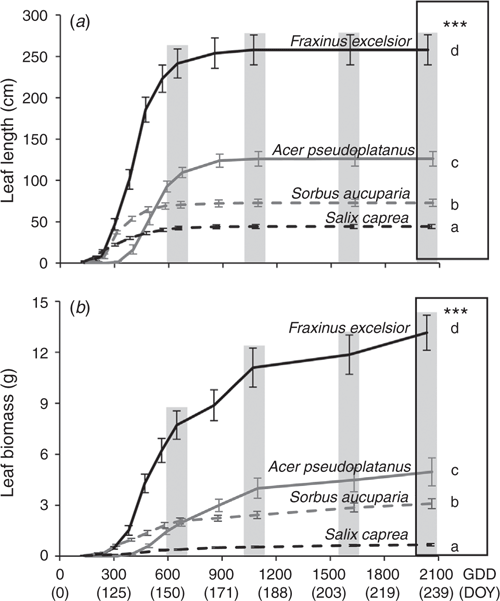
|
For each of the four species, a relationship between leaf length and leaf biomass was demonstrated. A remarkable amount of variance was explained by the regressions leaf length versus leaf biomass (regression parameters and equations are shown in Fig. 2).
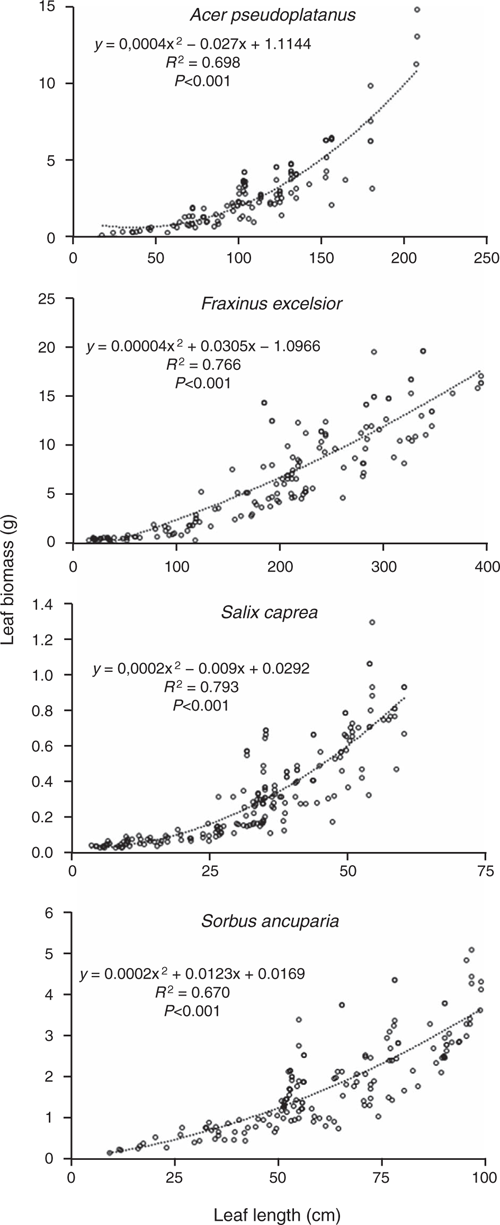
|
Proximate composition
The proximate composition was significantly different among the considered fodder tree species (Fig. 3). The average DM contents of S. caprea and S. aucuparia leaves were higher than those of A. pseudoplatanus and F. excelsior leaves. The ash content in S. aucuparia leaves was always lower than in the leaves of the other species. In F. excelsior, ash increased across time, displaying the highest values at the end of the season (Table 3). The highest EE values were observed in A. pseudoplatanus, followed by S. caprea, F. excelsior and S. aucuparia. Salix caprea was found to be the most fibrous species, with the highest concentrations of NDF, ADF and ADL. Fraxinus excelsior showed the lowest concentrations of ADF and ADL, while intermediate amounts of these fractions were found in A. pseudoplatanus and S. aucuparia leaves. Concerning NDF, F. excelsior showed values similar to those observed in S. caprea, whereas the lowest concentrations were found in A. pseudoplatanus and S. aucuparia.

|

|
The statistical analyses performed on the temporal variations in the proximate composition highlighted a general variability related to phenology advancement, with an overall decrease in CP and an increase in fibre content (Table 3).
Fatty acid profile
Regarding the FA profile of the leaves, the following 15 FA were detected in all samples: C12:0, C14:0, C15:0, C16:0, C16:1 c9, C18:0, C18:1 c9 (n9), C18:1 c11, C18:2 c9c12 (n6), C18:3 c6c9c12 (n6), C18:3 c9c12c15 (n3), C20:0, C20:1 c11, C22:0 and C20:4 c5c8c11c14 (n6). Among them, six FA, namely, palmitic acid (C16:0), stearic acid (C18:0), oleic acid (C18:1 c9), linoleic acid (C18:2 n6), α-linolenic acid (C18:3 n3) and γ-linolenic acid (C18:3 n6), comprised 92–97% of TFA and were then considered for further statistical analyses. The remaining FA were cumulated in the ‘Other FA’ group. In all tree species, C18:3 n3, C18:2 n6, and C16:0 were the main detected FA (Fig. 4). In S. aucuparia, the lowest total monounsaturated FA (MUFA, together with S. caprea), polyunsaturated FA (PUFA) and TFA concentrations were detected (Fig. 4). Concerning individual FA, S. aucuparia showed the highest C18:0 and the lowest C18:3 n6 and C18:3 n3 concentrations. Acer pseudoplatanus leaves had the highest C16:0 concentration, while F. excelsior showed the highest MUFA and C18:1 c9 concentrations. No differences were found for the ‘Other FA’ group among the species. The majority of individual FA and FA groups showed differences among the considered species during the whole trial. However, for total saturated fatty acids (SFA), C16:0, C18:3 n6, C18:3 n3, and ‘Other FA’, the differences among species tended to wane as DOY advanced.

|
Among sampling dates, FA profiles (Table 4) showed few significant variations. The concentration of C18:2 n6 was significantly (P < 0.001) higher at the first sampling date for all the species. Concerning F. excelsior and S. caprea, the sampling date influenced the concentration of total PUFA, which was significantly (P < 0.05 for both) higher at the first sampling date than at the third (F. excelsior) or the second and third sampling dates (S. caprea).

|
Phenolic compounds
On the basis of their mean phenolic compound values, the fodder tree leaves were significantly different (box plots in Fig. 5). Acer pseudoplatanus and S. caprea showed higher concentrations of TEP, TT and HT than did F. excelsior and S. aucuparia. The concentration of CT was lower in A. pseudoplatanus than in S. caprea and S. aucuparia. The CT were not detected in F. excelsior. No significant differences in NTP concentration were detected among the species. Variations in the phenolic composition among the sampling dates were not significant or negligible (Table 5).

|

|
In vitro true dry matter digestibility
The IVTD of leaves differed significantly among the fodder tree species (Fig. 6). On the basis of leaf mean digestibility values, F. excelsior and S. aucuparia were significantly more digestible than were A. pseudoplatanus and S. caprea.
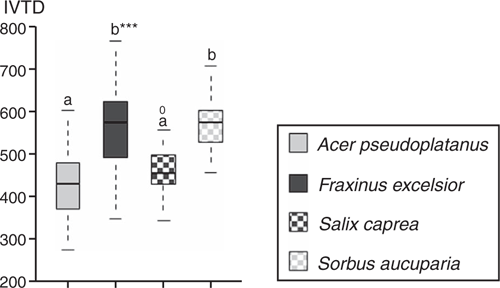
|
Sampling date affected A. pseudoplatanus (highest value: third sampling date; lowest value: final sampling date) and S. aucuparia (highest values: first and final sampling dates; lowest value: third sampling date) digestibility. Fraxinus excelsior and S. caprea leaves were characterised by a more stable digestibility throughout the vegetative season (Table 6).

|
Discussion
The results obtained in terms of the relationships among leaf traits confirmed the leaf length as an easy recordable in-field plant trait useful to estimate leaf biomass of a tree or shrub species via allometric equations (Itô and Sumida 2017; Škɹma et al. 2018). The use of allometric equations can, thus, represent a simple and effective tool to estimate foliage production, as it is a non-destructive and time-affordable method (Elzein et al. 2011; Konôpka et al. 2015; Pajtík and Konôpka 2015).
Concerning leaf development across the vegetative season, budburst dates for F. excelsior and S. aucuparia were consistent with those obtained in Swiss environments by Basler and Körner (2012). The same authors reported a later budburst for A. pseudoplatanus. As outlined in Table 2, precocity of leaf development appeared inversely proportional to final values of both leaf traits; leaves started to develop earlier (i.e. at a lower GDD) in the species with shorter and lighter leaves at the end of the vegetative season (i.e. S. caprea and S. aucuparia).
These considerations concerning variations in leaf length and biomass across the vegetative season can be used as proficient tools for fodder tree management, since productivity is one of the basic criteria for using a particular tree species as feeding resource (Papanastasis et al. 1998). Indeed, in European alpine regions, F. excelsior trees growing close to farm households were often used to forage ruminants (Gómez García and Fillat 1984). Trees were managed with pollard practice and yearling sprouts were given to animals either as fresh (in summer) or conserved (in winter) forage (Thiebault 2005).
Concerning the proximate composition, the results obtained for DM and EE were consistent with those already reported by other authors (Ricklefs and Matthew 1982; Kökten et al. 2012; Emile et al. 2016). The CP contents of the four species were higher than the minimum level of 70–80 g/kg DM required for optimum rumen function and feed intake in goats (Van Soest 1994), with A. pseudoplatanus showing the highest values. The range of CP content in tree foliage was comparable to the one of herbage growing in similar alpine environments across the vegetative season (Revello-Chion et al. 2011; Luske and van Eekeren 2015; Ravetto Enri et al. 2017). Conversely, in dry to arid environments, other authors reported higher CP contents in the foliage of deciduous fodder tree species than in local grass or hay (Yayneshet et al. 2009; Gardiner et al. 2013). Lu et al. (2005) showed that ADF concentrations of 180–200 g/kg DM or NDF concentrations of 410 g/kg DM meet the nutritional requirements for dairy goats. Other research highlighted that increasing the ADF or NDF contents over 180–200 g/kg DM and 600 g/kg DM respectively, may limit feed intake due to rumen fill (Santini et al. 1992; Mertens 1994; Riaz et al. 2014). Therefore, all the studied species showed adequate average NDF amounts according to these authors. However, if the leaves of the four species were given as sole feed, A. pseudoplatanus, S. aucuparia, and (with a stronger effect) S. caprea, may reduce the feed intake, due their high average ADF contents (Santini et al. 1992; Lu et al. 2005). In this respect, F. excelsior may exert the mildest effect, being slightly over the ADF limit. Other studies provided results sometimes different for the proximate composition, even if leaf samples were collected from the same tree species, in similar environments, and at comparable dates. Regarding F. excelsior, Emile et al. (2016) found lower DM and NDF and, together with Luske and van Eekeren (2015), comparable CP contents, while Masson et al. (1980) reported lower DM and higher ash and CP contents. Hejcman et al. (2016), instead, reported similar ADF but lower NDF, ADL and ash concentrations for S. aucuparia leaves in Iceland.
The temporal fluctuations in the proximate composition can be commonly observed in many fodder-tree species with similar trends, also in other than European environments (Papachristou and Papanastasis 1994; Yayneshet et al. 2009; Kökten et al. 2012), and in fresh grass from semi-natural pastures (Bovolenta et al. 2008).
The concentrations of the main FA were consistent with the findings for other fodder tree species and with grassland fodder values, also from environments other than the alpine ones (Ndondo et al. 2004; Uysal et al. 2015; Ravetto Enri et al. 2017). However, the concentration of C18:3 n3 in S. aucuparia leaves was lower than that observed in herbage collected in the same environment (Iussig et al. 2015a). Rosenqvist and Laakso (1991) reported a FA profile and main FA percentages comparable to those of the present study for S. caprea in northern Finland. Concerning A. pseudoplatanus, F. excelsior and S. aucuparia, no data on FA profile of leaves are currently available in literature. Other studies conducted in dryer environments have found amounts of some individual FA and FA groups similar to those of the fodder-tree species studied, while higher SFA and lower PUFA contents have been reported (Ndondo et al. 2004; Uysal et al. 2015). According to the obtained results on FA profile, A. pseudoplatanus, F. excelsior and S. caprea leaves could be used as feedstuff for goat nutrition, also in late summer, when quality (and particularly C18:3 n3 concentration) of grassland species decreases (Revello-Chion et al. 2011; Uysal et al. 2015; Ravetto Enri et al. 2017). Indeed, the lipid metabolism in the rumen and in the mammary gland can be affected by such decrease in the quality of grassland species, usually resulting in higher concentrations of hypercholesterolaemic SFA and lower concentrations of beneficial FA (i.e. vaccenic acid, rumenic acid and n3 FA) in the derived dairy and meat products (Ferlay et al. 2017).
The TEP, NTP, TT, CT and HT concentrations reported for other fodder tree species from drier regions were similar or even higher than those for the species here studied (Ammar et al. 2008; Kökten et al. 2012; Gemeda and Hassen 2015). Overall, CT concentrations were lower than the minimum threshold of 20 g/kg identified by Min et al. (2003) for ruminant nutrition. According to these and other authors, such CT values should have a limited negative effect on ruminal digestion, especially for goats. In fact, goats prefer tannin-rich foliage, minimising CT consumption through modification of the feed-intake pattern (Mkhise et al. 2018), and produce tannin-binding proteins in saliva, overcoming the negative impacts on digestibility in the rumen (Lamy et al. 2011).
Interesting and innovative results were shown by IVTD of the four species, which was similar to that of some Mediterranean tree and shrub species, either deciduous or evergreen (Ammar et al. 2005, 2008). Digestibility in rumen environment is a complex process, partially still unknown, in which several factors interact with each other (Aguerre et al. 2016; Seidavi et al. 2018). Chemical composition (i.e. mainly fibre fractions (NDF, ADF, ADL), FA profile (particularly PUFA), and phenolic compounds (above all, CT and HT) exert a crucial multi-layered influence on microbial activity (Waghorn 2008; Boufennara et al. 2012; Ferlay et al. 2017). The NDF constituents, being mainly represented by hemicellulose and cellulose, are the dominant feed fraction for grazing ruminants, supplying energy for maintenance and production. However, fibre has a slower ruminal passage rate than do other dietary elements because their chemical conformation is difficult to cleave or is even indigestible, resulting in a filling effect over time (Mertens 1994). The inverse relationship between the amount of fibre and digestibility rate reported for other fodder tree species in the Mediterranean region (Ammar et al. 2004; Boufennara et al. 2012; Gemeda and Hassen 2015) can be observed also in the present trial. This means high NDF, ADF and ADL and low IVTD in S. caprea, low NDF and high IVTD in S. aucuparia, and low ADF and ADL and high IVTD in F. excelsior. Interestingly, these relationships were not displayed in other cases, such as in A. pseudoplatanus (low NDF and low IVTD), F. excelsior (high NDF and high IVTD) and S. aucuparia (high ADF and ADL and high IVTD), as observed also in previous studies (Van Soest 1994; Riaz et al. 2014; Gemeda and Hassen 2015). This could be attributable to the additional influence of the other chemicals on the microbiome. The interaction of fibre with different PUFA concentrations could have also played an important role in digestibility, since PUFA can inhibit and/or alter the microbial activity and the biohydrogenation pathways in the rumen (Ferlay et al. 2017). For this reason, the high PUFA concentrations (especially concentrations of C18:3 n6, C18:3 n3 and C18:2 n6) in A. pseudoplatanus and S. caprea leaves, together with their high ADL contents, could have, consequently, contributed to the reduction of the degradation capacity of goat microbiome and IVTD. Indeed, ADL represents a physical barrier between digestible cell content and bacteria (Waghorn and McNabb 2003). In contrast, IVTD was promoted in F. excelsior because of the low amount of ADL (despite the high PUFA concentration) and in S. aucuparia because of the low PUFA concentration (despite the high ADL concentration). Concerning phenolic compounds, CT may have affected digestion mechanisms (i.e. by binding to dietary protein, fibre and enzymes and by reducing microbiome degradation activity; Makkar 2003; Mueller-Harvey 2006; Waghorn 2008). The chemical structure of CT can differ among tree species and can exert different binding activities, regardless of their concentration, resulting in different IVTD (Hove et al. 2001; McSweeney et al. 2001). In the present case, the lower IVTD associated with lower CT concentrations observed in A. pseudoplatanus leaves may be due to a higher binding activity of the CT molecules. The higher IVTD associated with the higher CT concentration in S. aucuparia leaves is probably due to a lower CT activity. The highest HT concentrations found in A. pseudoplatanus and S. caprea leaves (approximately 6-fold those of F. excelsior and S. aucuparia) may have negatively influenced the digestibility of these leaves, because of the potential release of compounds toxic for bacteria in the rumen (Makkar 2003; Waghorn 2008). Therefore, the high CT (with a probable high binding activity) and HT concentrations in S. caprea leaves could have combined and lowered the digestibility. Conversely, F. excelsior leaves showed a phenolic profile more suitable for ruminal digestion, since they were totally free of CT and had low HT concentrations.
The four species showed a consistent stability of IVTD throughout the vegetative season. This is in contrast with the results previously obtained for Quercus coccifera L. (an evergreen Mediterranean tree species), which showed a progressive decline of leaf digestibility while dry season advanced. However, it should be pointed out that evergreen species generally show lower quality at late phenological stages (Roukos 2016).
Additional research on productivity and chemical and digestibility features of other tree portions (such as sprouts) or other temperate tree species selected by goats in extensive European farming systems (e.g. Alnus viridis (Chaix) DC., B. pendula, Corylus avellana L., F. sylvatica, Quercus sp., Tilia cordata Mill., Ulmus spp.) would be advisable (Hejcman et al. 2014; Castro and Fernández Núñez 2016; Vandermeulen et al. 2018a). Moreover, the preferences by goats for the investigated tree species should be taken into consideration. Future trials should address topics such as the palatability of tree and shrub leaves and sprouts, the related goat diet selection and the effects of tree and shrub leaves and sprouts, used as sole feed or as supplement, on voluntary DM intake by goats, on digestion mechanisms (especially on protein and organic-matter digestibility and on total tract digestion), even through in vivo studies, and on goat milk yield, gross composition and FA profile.
Conclusions
The present study has provided original data with a comprehensive approach on leaf traits, and chemical features of four tree species widespread in Europe.
In situ, chemical, and in vitro measurements can be considered useful tools in initial screening studies to rank forages according to their nutritive quality. The present results have highlighted that the investigated species, due to their foliage production, proximate composition, FA profile, phenolic composition and digestibility, can represent good-quality feedstuff for goat nutrition. This is especially true in late summer when herbage quality decreases, particularly in terms of CP and the FA profile.
These species can supply goat feedstuff either for direct browsing or as fresh or dried fodder (Bestman et al. 2014), also depending on foliage production. Trees could be managed through copping and maintained at a maximum height of 1.80 m to facilitate direct browsing by goats. Alternatively, they could be pollarded to prevent damages from wild ungulates and their branches could be periodically cut for fresh- or dry-fodder delivery. Otherwise, new specific plantations for goat browsing could be established. In this case, the species of the stand and the relative abundance should be selected, taking into account goat feeding preferences as well as leaf nutritive values of the tree species.
The differences among the four selected species were remarkable, even if weakly related to phenological stage advancement, especially when considering the FA and phenolic composition. The high digestibility of F. excelsior leaves throughout the vegetative stage could be due to the positive influence of the low concentrations of ADL and phenolic compounds, irrespective of the low CP concentrations. This species could be regarded as potentially highly-nutritive feedstuff, also improving the quality of goat dairy products for human nutrition, as confirmed by its traditional use in European mountain areas (Gómez García and Fillat 1984; Thiebault 2005). Also S. aucuparia leaves showed similar digestibility values; however, the low CP and PUFA concentrations (especially C18:3 n3 concentration) can partially reduce the interest for this species as a feeding resource for goat dairy and meat products with healthy properties. Conversely, a lower digestibility was found in S. caprea samples, which was also the species with the less productive sprouts, especially at the beginning of the vegetative season, as a consequence of its high phenolic and ADL concentrations. Nevertheless, the FA profile of this species highlighted high C18:2 n6, C18:3 n6 and C18:3 n3 concentrations, generally being recognised as main lipid precursors in ruminant metabolism for the synthesis of FA considered beneficial to human health (e.g. vaccenic acid, rumenic acid and n3 FA). A modest digestibility was recorded for A. pseudoplatanus leaves, despite their high CP and medium–low ADL concentrations, which were probably insufficient to contrast the high concentrations of phenolics (TEP, TT and HT) and the high binding activity (CT), even if they displayed a good level of the same ‘healthy precursors’ as reported for S. caprea foliage.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
Financial support for this work was obtained by Fondazione Zegna, Trivero (BI). The authors thank Elisa Perotti for her essential help in fieldwork and Riccardo Fortina, Claudia Iseni, Vanda Malfatto and Sonia Tassone for their careful technical assistance.
References
Abo-Donia FM, Yang LY, Hristov AN, Wang M, Tang SX, Zhou CS, Han XF, Kang JH, Tan ZL, He ZX (2017) Effects of tannins on the fatty acid profiles of rumen fluids and milk from lactating goats fed a total mixed ration containing rapeseed oil. Livestock Science 204, 16–24.| Effects of tannins on the fatty acid profiles of rumen fluids and milk from lactating goats fed a total mixed ration containing rapeseed oil.Crossref | GoogleScholarGoogle Scholar |
Aguerre MJ, Capozzolo MC, Lencioni P, Cabral C, Wattiaux MA (2016) Effect of quebracho–chestnut tannin extracts at 2 dietary crude protein levels on performance, rumen fermentation, and nitrogen partitioning in dairy cows. Journal of Dairy Science 99, 4476–4486.
| Effect of quebracho–chestnut tannin extracts at 2 dietary crude protein levels on performance, rumen fermentation, and nitrogen partitioning in dairy cows.Crossref | GoogleScholarGoogle Scholar | 27060814PubMed |
Álvarez‐Martínez J, Gómez‐Villar A, Lasanta T (2016) The use of goats grazing to restore pastures invaded by shrubs and avoid desertification: a preliminary case study in the Spanish Cantabrian Mountains. Land Degradation & Development 27, 3–13.
| The use of goats grazing to restore pastures invaded by shrubs and avoid desertification: a preliminary case study in the Spanish Cantabrian Mountains.Crossref | GoogleScholarGoogle Scholar |
Alves SP, Cabrita ARJ, Fonseca AJM, Bessa RJB (2008) Improved method for fatty acid analysis in herbage based on direct transesterification followed by solid-phase extraction. Journal of Chromatography. A 1209, 212–219.
| Improved method for fatty acid analysis in herbage based on direct transesterification followed by solid-phase extraction.Crossref | GoogleScholarGoogle Scholar | 18814883PubMed |
Ammar H, López S, González JS, Ranilla MJ (2004) Seasonal variations in the chemical composition and in vitro digestibility of some Spanish leguminous shrub species. Animal Feed Science and Technology 115, 327–340.
| Seasonal variations in the chemical composition and in vitro digestibility of some Spanish leguminous shrub species.Crossref | GoogleScholarGoogle Scholar |
Ammar H, López S, González JS (2005) Assessment of the digestibility of some Mediterranean shrubs by in vitro techniques. Animal Feed Science and Technology 119, 323–331.
| Assessment of the digestibility of some Mediterranean shrubs by in vitro techniques.Crossref | GoogleScholarGoogle Scholar |
Ammar H, López S, Andrés S, Ranilla MJ, Bodas R, González JS (2008) In vitro digestibility and fermentation kinetics of some browse plants using sheep or goat ruminal fluid as the source of inoculum. Animal Feed Science and Technology 147, 90–104.
| In vitro digestibility and fermentation kinetics of some browse plants using sheep or goat ruminal fluid as the source of inoculum.Crossref | GoogleScholarGoogle Scholar |
ANKOM (2017) ‘ANKOM Technology method 3. In vitro true digestibility using the DAISY II incubator.’ Available at https://www.ankom.com/sites/default/files/document-files/Method_3_Invitro_D200_D200I.pdf [Verified 13 December 2018]
AOAC (Association of Official Analytical Chemists) (2000) ‘Official methods of analysis of AOAC International.’ (Association of Official Analytical Chemists: Gaithersburg, MD, USA)
AOAC (Association of Official Analytical Chemists) (2003) ‘Official methods of analysis of AOAC International.’ (Association of Official Analytical Chemists: Gaithersburg, MD, USA)
Arigbede OM, Tan ZL, Anele UY, Sun ZH, Tang SX, Han XF, Zhou CS, Zeng B (2012) Effects of age and species on agronomic performance, chemical composition and in vitro gas production of some tropical multi-purpose tree species. The Journal of Agricultural Science 150, 725–737.
| Effects of age and species on agronomic performance, chemical composition and in vitro gas production of some tropical multi-purpose tree species.Crossref | GoogleScholarGoogle Scholar |
Basler D, Körner C (2012) Photoperiod sensitivity of bud burst in 14 temperate forest tree species. Agricultural and Forest Meteorology 165, 73–81.
| Photoperiod sensitivity of bud burst in 14 temperate forest tree species.Crossref | GoogleScholarGoogle Scholar |
Battaglini L, Bovolenta S, Gusmeroli F, Salvador S, Sturaro E (2014) Environmental sustainability of Alpine livestock farms. Italian Journal of Animal Science 13, 3155
| Environmental sustainability of Alpine livestock farms.Crossref | GoogleScholarGoogle Scholar |
Bestman M, van Eekeren N, Luske B, Vonk M, Anssems E, Boosten M, van Bree M (2014) ‘Introducing trees in dairy and poultry farms: experiences dairy and poultry farmers’ networks in The Netherlands.’ Available at http://orgprints.org/23894/ [Verified 13 December 2018]
Boufennara S, Lopez S, Bousseboua H, Bodas R, Bouazza L (2012) Chemical composition and digestibility of some browse plant species collected from Algerian arid rangelands. Spanish Journal of Agricultural Research 10, 88–98.
| Chemical composition and digestibility of some browse plant species collected from Algerian arid rangelands.Crossref | GoogleScholarGoogle Scholar |
Bovolenta S, Spanghero M, Dovier S, Orlandi D, Clementel F (2008) Chemical composition and net energy content of alpine pasture species during the grazing season. Animal Feed Science and Technology 146, 178–191.
| Chemical composition and net energy content of alpine pasture species during the grazing season.Crossref | GoogleScholarGoogle Scholar |
Castro M, Fernández Núñez E (2016) Seasonal grazing of goats and sheep on Mediterranean mountain rangelands of northeast Portugal. Livestock Research for Rural Development 28,
Elzein TM, Blarquez O, Gauthier O, Carcaillet C (2011) Allometric equations for biomass assessment of subalpine dwarf shrubs. Alpine Botany 121, 129
| Allometric equations for biomass assessment of subalpine dwarf shrubs.Crossref | GoogleScholarGoogle Scholar |
Emile JC, Delagarde R, Barre P, Novak S (2016) Nutritive value and degradability of leaves from temperate woody resources for feeding ruminants in summer. In ‘The 3rd European agroforestry conference’, 23–25 May 2016, Montpellier, France.
Ferlay A, Bernard L, Meynadier A, Malpuech-Brugère C (2017) Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: a review. Biochimie 141, 107–120.
| Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: a review.Crossref | GoogleScholarGoogle Scholar | 28804001PubMed |
Forrester DI, Tachauer IHH, Annighoefer P, Barbeito I, Pretzsch H, Ruiz-Peinado R, Stark H, Vacchiano G, Zlatanov T, Chakraborty T, Saha S, Sileshi GW (2017) Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. Forest Ecology and Management 396, 160–175.
| Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate.Crossref | GoogleScholarGoogle Scholar |
Francisco A, Dentinho MT, Alves SP, Portugal PV, Fernandes F, Sengo S, Jerónimo E, Oliveira MA, Costa P, Sequeira A, Bessa RJB, Santos-Silva J (2015) Growth performance, carcass and meat quality of lambs supplemented with increasing levels of a tanniferous bush (Cistus ladanifer L.) and vegetable oils. Meat Science 100, 275–282.
| Growth performance, carcass and meat quality of lambs supplemented with increasing levels of a tanniferous bush (Cistus ladanifer L.) and vegetable oils.Crossref | GoogleScholarGoogle Scholar | 25460137PubMed |
Gardiner C, Kempe N, Hannah I, McDonald J (2013) PROGARDESTM: a legume for tropical/subtropical semi-arid clay soils. Tropical Grasslands–Forrajes Tropicales 1, 78–80.
| PROGARDESTM: a legume for tropical/subtropical semi-arid clay soils.Crossref | GoogleScholarGoogle Scholar |
Gemeda BS, Hassen A (2015) Effect of tannin and species variation on in vitro digestibility, gas, and methane production of tropical browse plants. Asian-Australasian Journal of Animal Sciences 28, 188–199.
| Effect of tannin and species variation on in vitro digestibility, gas, and methane production of tropical browse plants.Crossref | GoogleScholarGoogle Scholar | 25557814PubMed |
Glasser F, Doreau M, Maxin G, Baumont R (2013) Fat and fatty acid content and composition of forages: a meta-analysis. Animal Feed Science and Technology 185, 19–34.
| Fat and fatty acid content and composition of forages: a meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Gómez García D, Fillat F (1984) Utilisation du Frêne comme arbre fourrager dans les Pyrénées de Huesca. In ‘Écologie des milieux Montagnards et de haute altitude. Documents d’écologie Pyrénéenne, III–IV’. (Ed. Laboratoire d’Écologie Montagnarde) pp. 481–489. (Lazare: Bordeaux, France) [In French.]
Guinier P (1950) Botanique et économie rurale en montagne. Bulletin de la Société Botanique de France 97, 224–234.
| Botanique et économie rurale en montagne.Crossref | GoogleScholarGoogle Scholar |
Hack H, Bleiholder H, Buhr L, Meier U, Schnock-Fricke U, Weber E, Witzenberger A (1992) Einheitliche Codierung der phänologischen Entwicklungsstadien mono-und dikotyler Pflanzen – erweiterte BBCH-Skala, Allgemein. Nachrichtenblatt des deutschen Pflanzenschutzdienstes 44, 265–270.
Hejcman M, Hejcmanová P, Stejskalová M, Pavlů V (2014) Nutritive value of winter-collected annual twigs of main European woody species, mistletoe and ivy and its possible consequences for winter foddering of livestock in prehistory. The Holocene 24, 659–667.
| Nutritive value of winter-collected annual twigs of main European woody species, mistletoe and ivy and its possible consequences for winter foddering of livestock in prehistory.Crossref | GoogleScholarGoogle Scholar |
Hejcman M, Hejcmanová P, Pavlů V, Thorhallsdottir AG (2016) Forage quality of leaf fodder from the main woody species in Iceland and its potential use for livestock in the past and present. Grass and Forage Science 71, 649–658.
| Forage quality of leaf fodder from the main woody species in Iceland and its potential use for livestock in the past and present.Crossref | GoogleScholarGoogle Scholar |
Hejcmanová P, Stejskalová M, Hejcman M (2014) Forage quality of leaf-fodder from the main broad-leaved woody species and its possible consequences for the Holocene development of forest vegetation in central Europe. Vegetation History and Archaeobotany 23, 607–613.
| Forage quality of leaf-fodder from the main broad-leaved woody species and its possible consequences for the Holocene development of forest vegetation in central Europe.Crossref | GoogleScholarGoogle Scholar |
Hove L, Topps JH, Sibanda S, Ndlovu LR (2001) Nutrient intake and utilisation by goats fed dried leaves of the shrub legumes Acacia angustissima, Calliandra calothyrsus and Leucaena leucocephala as supplements to native pasture hay. Animal Feed Science and Technology 91, 95–106.
| Nutrient intake and utilisation by goats fed dried leaves of the shrub legumes Acacia angustissima, Calliandra calothyrsus and Leucaena leucocephala as supplements to native pasture hay.Crossref | GoogleScholarGoogle Scholar |
Itô H, Sumida A (2017) Allometric and growth data of an evergreen oak, Quercus glauca, in a secondary broadleaved forest. Ecological Research 32, 105
| Allometric and growth data of an evergreen oak, Quercus glauca, in a secondary broadleaved forest.Crossref | GoogleScholarGoogle Scholar |
Iussig G, Renna M, Gorlier A, Lonati M, Lussiana C, Battaglini LM, Lombardi G (2015a) Browsing ratio, species intake, and milk fatty acid composition of goats foraging on alpine open grassland and grazable forestland. Small Ruminant Research 132, 12–24.
| Browsing ratio, species intake, and milk fatty acid composition of goats foraging on alpine open grassland and grazable forestland.Crossref | GoogleScholarGoogle Scholar |
Iussig G, Lonati M, Probo M, Hodge S, Lombardi G (2015b) Plant species selection by goats foraging on montane semi-natural grasslands and grazable forestlands in the Italian Alps. Italian Journal of Animal Science 14, 484–494.
| Plant species selection by goats foraging on montane semi-natural grasslands and grazable forestlands in the Italian Alps.Crossref | GoogleScholarGoogle Scholar |
Khanal RC, Subba DB (2001) Nutritional evaluation of leaves from some major fodder trees cultivated in the hills of Nepal. Animal Feed Science and Technology 92, 17–32.
| Nutritional evaluation of leaves from some major fodder trees cultivated in the hills of Nepal.Crossref | GoogleScholarGoogle Scholar |
Kökten K, Kaplarn M, Hatipo?lu R, Saruhan V, Çinar S (2012) Nutritive value of Mediterranean shrubs. Journal of Animal and Plant Sciences 22, 188–194.
Kongmanila D (2012) Erythrina foliage as an alternative feed for growing goats in Lao PDR. Doctoral Thesis, Sveriges Lantbruksuniversitet, Acta Universitatis Agriculturae Sueciae, Uppsala, Sweden.
Konôpka B, Pajtík J, Šebeň V (2015) Biomass functions and expansion factors for young trees of European ash and Sycamore maple in the inner western Carpathians Biomassefunktionen und Expansionsfaktoren für junge Europäische Eschen und Bergahorne in den Inneren Westlichen Karpaten. Austrian Journal of Forest Science 132, 1–26.
Kühn M, Maier U, Herbig C, Ismail-Meyer K, Bailly ML, Wick L (2013) Methods for the examination of cattle, sheep and goat dung in prehistoric wetland settlements with examples of the sites Alleshausen– Täschenwiesen and Alleshausen–Grundwiesen (around cal 2900 BC) at Lake Federsee, south-west Germany. Environmental Archaeology 18, 43–57.
| Methods for the examination of cattle, sheep and goat dung in prehistoric wetland settlements with examples of the sites Alleshausen– Täschenwiesen and Alleshausen–Grundwiesen (around cal 2900 BC) at Lake Federsee, south-west Germany.Crossref | GoogleScholarGoogle Scholar |
Lamy E, Rawel H, Schweigert FJ, Capela E, Silva F, Ferreira A, Costa AR, Antunes C, Almeida AM, Coelho AV, Sales-Baptista E (2011) The effect of tannins on Mediterranean ruminant ingestive behavior: the role of the oral cavity. Molecules (Basel, Switzerland) 16, 2766–2784.
| The effect of tannins on Mediterranean ruminant ingestive behavior: the role of the oral cavity.Crossref | GoogleScholarGoogle Scholar |
Leng RA (1997) ‘Tree foliage in ruminant nutrition.’ Issue 139. FAO animal production and health paper. (Food and Agriculture Organization of the United Nations: Rome, IT)
Lu CD, Kawas JR, Mahgoub OG (2005) Fibre digestion and utilization in goats. Small Ruminant Research 60, 45–52.
| Fibre digestion and utilization in goats.Crossref | GoogleScholarGoogle Scholar |
Luske B, van Eekeren N (2015) Potential of fodder trees in high-output dairy systems. In ‘Proceedings of the 18th symposium on European Grassland Federation’, 14–17 June 2015, Wageningen, The Netherlands.
Makkar HPS (2003) Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Ruminant Research 49, 241–256.
| Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds.Crossref | GoogleScholarGoogle Scholar |
Manousidis T, Kyriazopoulos AP, Parissi ZM, Abraham EM, Korakis G, Abas Z (2016) Grazing behavior, forage selection and diet composition of goats in a Mediterranean woody rangeland. Small Ruminant Research 145, 142–153.
| Grazing behavior, forage selection and diet composition of goats in a Mediterranean woody rangeland.Crossref | GoogleScholarGoogle Scholar |
Masson C, Decaen C, Delamarche F, Faurie F (1980) Composition chimique et valeur alimentaire des jeunes pousses de peuplier (Populus) et de frêne (Fraxinus). Annales de Zootechnie 29, 195–200.
| Composition chimique et valeur alimentaire des jeunes pousses de peuplier (Populus) et de frêne (Fraxinus).Crossref | GoogleScholarGoogle Scholar |
McSweeney CS, Palmer B, McNeill DM, Krause DO (2001) Microbial interactions with tannins: nutritional consequences for ruminants. Animal Feed Science and Technology 91, 83–93.
| Microbial interactions with tannins: nutritional consequences for ruminants.Crossref | GoogleScholarGoogle Scholar |
Mertens DR (1994) Regulation of forage intake. In ‘Forage quality, evaluation and utilization’. (Eds GC Fahey Jr, M Collins, DR Mertens, LE Moser) pp. 450–493. (American Society of Agronomy: Madison, WI, USA)
Min BR, Barry TN, Attwood GT, McNabb WC (2003) The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: a review. Animal Feed Science and Technology 106, 3–19.
| The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: a review.Crossref | GoogleScholarGoogle Scholar |
Mkhize NR, Heitkönig IMA, Scogings PF, Hattas D, Dziba LE, Prins HHT, de Boer WF (2018) Seasonal regulation of condensed tannin consumption by free-ranging goats in a semi-arid savanna. PLoS One 13, e0189626
| Seasonal regulation of condensed tannin consumption by free-ranging goats in a semi-arid savanna.Crossref | GoogleScholarGoogle Scholar | 29293513PubMed |
Mueller-Harvey I (2006) Unravelling the conundrum of tannins in animal nutrition and health. Journal of the Science of Food and Agriculture 86, 2010–2037.
| Unravelling the conundrum of tannins in animal nutrition and health.Crossref | GoogleScholarGoogle Scholar |
Ndondo IB, Wilhelmi BS, Mabinya LV, Brand JM (2004) Alpha-tocopherol and fatty acids of major browse plant species of black rhinoceros in the Great Fish River Reserve: research article. South African Journal of Wildlife Research 34, 87–93.
Novak S (2014) Oasys: une expérimentation-système pour tester un système bovin laitier innovant, diversifié et durable. Available at http://www.nouvelle-aquitaine-poitiers.inra.fr%2FOutils-et-Ressources%2FDispositifs-experimentaux%2FOasys [Verified 13 December 2018]
Pajtík J, Konôpka B (2015) Quantifying edible biomass on young Salix caprea and Sorbus aucuparia trees for Cervus elaphus: estimates by regression models Quantifizierung der Biomasse von jungen Salweiden (Salix caprea) und Ebereschen (Sorbus aucuparia) die für Cervus elaphus genießbar ist: eine Bestimmung durch Regressionsmodelle. Austrian Journal of Forest Science 132, 61–80.
Papachristou TG, Papanastasis VP (1994) Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo-pastoral systems for goats. Agroforestry Systems 27, 269–282.
| Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo-pastoral systems for goats.Crossref | GoogleScholarGoogle Scholar |
Papanastasis VP, Platis PD, Dini-Papanastasi O (1998) Effects of age and frequency of cutting on productivity of Mediterranean deciduous fodder tree and shrub plantations. Forest Ecology and Management 110, 283–292.
| Effects of age and frequency of cutting on productivity of Mediterranean deciduous fodder tree and shrub plantations.Crossref | GoogleScholarGoogle Scholar |
Prakash B, Dhali A, Rathore SS, Das KC, Walling I, Vupru K, Mech A, Baruah KK, Rajkhowa C (2009) Chemical composition and nutritional evaluation of various foliages consumed by mithun (Bos frontalis). Animal Feed Science and Technology 150, 223–229.
| Chemical composition and nutritional evaluation of various foliages consumed by mithun (Bos frontalis).Crossref | GoogleScholarGoogle Scholar |
Prislan P, Gričar J, de Luis M, Smith KT, Čufar K (2013) Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agricultural and Forest Meteorology 180, 142–151.
| Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites.Crossref | GoogleScholarGoogle Scholar |
Ravetto Enri S, Renna M, Probo M, Lussiana C, Battaglini LM, Lonati M, Lombardi G (2017) Relationships between botanical and chemical composition of forages: a multivariate approach to grasslands in the western Italian Alps. Journal of the Science of Food and Agriculture 97, 1252–1259.
| Relationships between botanical and chemical composition of forages: a multivariate approach to grasslands in the western Italian Alps.Crossref | GoogleScholarGoogle Scholar | 27324464PubMed |
Renna M, Gasmi-Boubaker A, Lussiana C, Battaglini LM, Belfayez K, Fortina R (2014) Fatty acid composition of the seed oils of selected Vicia L. taxa from Tunisia. Italian Journal of Animal Science 13, 2.
| Fatty acid composition of the seed oils of selected Vicia L. taxa from Tunisia.Crossref | GoogleScholarGoogle Scholar |
Revello-Chion A, Tabacco E, Peiretti PG, Borreani G (2011) Variation in the fatty acid composition of alpine grassland during spring and summer. Agronomy Journal 103, 1072–1080.
| Variation in the fatty acid composition of alpine grassland during spring and summer.Crossref | GoogleScholarGoogle Scholar |
Riaz MQ, Südekum K-H, Clauss M, Jayanegara A (2014) Voluntary feed intake and digestibility of four domestic ruminant species as influenced by dietary constituents: a meta-analysis. Livestock Science 162, 76–85.
| Voluntary feed intake and digestibility of four domestic ruminant species as influenced by dietary constituents: a meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Ricklefs RE, Matthew KK (1982) Chemical characteristics of the foliage of some deciduous trees in southeastern Ontario. Canadian Journal of Botany 60, 2037–2045.
| Chemical characteristics of the foliage of some deciduous trees in southeastern Ontario.Crossref | GoogleScholarGoogle Scholar |
Rosenqvist H, Laakso S (1991) Fatty acids and alkanes in leaves of frost-tolerant and frost-susceptible willows. Phytochemistry 30, 2161–2164.
| Fatty acids and alkanes in leaves of frost-tolerant and frost-susceptible willows.Crossref | GoogleScholarGoogle Scholar |
Roukos CH (2016) Seasonal and altitudinal variations in nutritional quality of kermes oak (Quercus coccifera L.) in northwest Greece and extensive goat farming. Bulgarian Journal of Agricultural Science 22, 804–814.
San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (2016) ‘European atlas of forest tree species.’ (Publications Office of the European Union: Luxembourg)
Santini FJ, Lu CD, Potchoiba MJ, Fernandez JM, Coleman SW (1992) Dietary fiber and milk yield, mastication, digestion, and rate of passage in goats fed alfalfa hay. Journal of Dairy Science 75, 209–219.
| Dietary fiber and milk yield, mastication, digestion, and rate of passage in goats fed alfalfa hay.Crossref | GoogleScholarGoogle Scholar | 1311727PubMed |
Seidavi A, Tavakoli M, Rasouli B, Corazzin M, Salem AZM (2018) Application of some trees/shrubs in ruminant feeding: a review. Agroforestry Systems 1–12.
| Application of some trees/shrubs in ruminant feeding: a review.Crossref | GoogleScholarGoogle Scholar |
Simbaya J (2002) Potential of fodder tree/shrub legumes as a feed resource for dry season supplementation of smallholder ruminant animals. In ‘Proceedings of the final review meeting of an IAEA technical co-operation regional AFRA project’, Cairo, Egypt. pp. 69–76. (International Atomic Energy Agency: Cairo, Egypt)
Škɹma M, Mikšys V, Aleinikovas M, Šilinskas B, Varnagirytɹ-Kabašinskienɹ I (2018) Biomass Structure and morphometric parameters for non-destructive biomass estimation of common forest underbrush species in Lithuania. Polish Journal of Environmental Studies 27, 325–333.
| Biomass Structure and morphometric parameters for non-destructive biomass estimation of common forest underbrush species in Lithuania.Crossref | GoogleScholarGoogle Scholar |
Tefera S, Mlambo V, Dlamini BJ, Dlamini AM, Koralagama KDN, Mould FL (2008) Chemical composition and in vitro ruminal fermentation of common tree forages in the semi-arid rangelands of Swaziland. Animal Feed Science and Technology 142, 99–110.
| Chemical composition and in vitro ruminal fermentation of common tree forages in the semi-arid rangelands of Swaziland.Crossref | GoogleScholarGoogle Scholar |
Thiebault S (2005) L’apport du fourrage d’arbre dans l’élevage depuis le Néolithique. Anthropozoologica 40, 95–108.
Uysal S, Zengin G, Aktumsek A, Karatas S (2015) Fatty acid composition, total sugar content and anti-diabetic activity of methanol and water extracts of nine different fruit tree leaves collected from Mediterranean region of Turkey. International Journal of Food Properties 18, 2268–2276.
| Fatty acid composition, total sugar content and anti-diabetic activity of methanol and water extracts of nine different fruit tree leaves collected from Mediterranean region of Turkey.Crossref | GoogleScholarGoogle Scholar |
Van Soest PJ (1994) ‘Nutritional ecology of the ruminant.’ 2nd edn. (Cornell University Press: Ithaca, NY, USA)
Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science 74, 3583–3597.
| Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition.Crossref | GoogleScholarGoogle Scholar | 1660498PubMed |
Vandermeulen S, Ramírez-Restrepo CA, Beckers Y, Claessens H, Bindelle J (2018a) Agroforestry for ruminants: a review of trees and shrubs as fodder in silvopastoral temperate and tropical production systems. Animal Production Science 58, 767–777.
| Agroforestry for ruminants: a review of trees and shrubs as fodder in silvopastoral temperate and tropical production systems.Crossref | GoogleScholarGoogle Scholar |
Vandermeulen S, Ramírez-Restrepo CA, Marche C, Decruyenaere V, Beckers Y, Bindelle J (2018b) Behaviour and browse species selectivity of heifers grazing in a temperate silvopastoral system. Agroforestry Systems 92, 705–716.
| Behaviour and browse species selectivity of heifers grazing in a temperate silvopastoral system.Crossref | GoogleScholarGoogle Scholar |
Vasta V, Bessa RJB (2012) Manipulating ruminal biohydrogenation by the use of plants bioactive compounds. In ‘Dietary phytochemicals and microbes’. pp. 263–284. (Springer: Dordrecht, The Netherlands)
Waghorn G (2008) Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production: progress and challenges. Animal Feed Science and Technology 147, 116–139.
| Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production: progress and challenges.Crossref | GoogleScholarGoogle Scholar |
Waghorn GC, McNabb WC (2003) Consequences of plant phenolic compounds for productivity and health of ruminants. The Proceedings of the Nutrition Society 62, 383–392.
| Consequences of plant phenolic compounds for productivity and health of ruminants.Crossref | GoogleScholarGoogle Scholar | 14506885PubMed |
Yayneshet T, Eik LO, Moe SR (2009) Seasonal variations in the chemical composition and dry matter degradability of exclosure forages in the semi-arid region of northern Ethiopia. Animal Feed Science and Technology 148, 12–33.
| Seasonal variations in the chemical composition and dry matter degradability of exclosure forages in the semi-arid region of northern Ethiopia.Crossref | GoogleScholarGoogle Scholar |