

The effects of Alnus viridis encroachment in mountain pastures on the growth performance, carcass and meat quality of Dexter cattle and Engadine sheep
T. Zehnder A B , M. K. Schneider B , A. Lüscher B , K. Giller A , P. Silacci C , J. Messadène-Chelali C , J. Berard A C * and M. Kreuzer A
B , A. Lüscher B , K. Giller A , P. Silacci C , J. Messadène-Chelali C , J. Berard A C * and M. Kreuzer A
A ETH Zurich, Institute of Agricultural Sciences, Eschikon 27, Lindau 8315, Switzerland.
B Agroscope, Forage Production and Grassland Systems, Reckenholzstrasse 191, Zurich 8046, Switzerland.
C Agroscope, Animal Production Systems, Food Microbial Systems and Method Development and Analytics, Route de la Tioleyre 4, Posieux 1725, Switzerland.
Animal Production Science 63(12) 1248-1260 https://doi.org/10.1071/AN22409
Submitted: 26 August 2022 Accepted: 24 April 2023 Published: 17 May 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Climate and economic changes promote the encroachment of woody plants on marginal mountain pastures. In the Alps, the main woody invader is the nitrogen-fixing shrub Alnus viridis (green alder). Robust breeds of different ruminant species may perform well on these shrub-encroached pastures.
Aims: To assess the effects of A. viridis encroachment levels of mountain pastures on the growth performance and carcass and meat characteristics of cattle and sheep.
Methods: For 8–9 weeks, 22 15-month-old growing Dexter cattle and 20 7-month-old weaned Engadine sheep were grazed rotationally on pastures in a subalpine zone with four different levels of A. viridis encroachment (average: 0.8%, 28.7%, 50.3% and 67.8%). Following slaughter, the assessed carcass and meat characteristics (longissimus thoracis and biceps femoris) included physicochemical meat quality, muscle fibre typing, fatty acid profiling (intramuscular and perirenal fat) and the sensory properties of patties.
Key results: The shrub encroachment levels had only minor effects on the animals. Interactions with the animal species influenced some sensory and fatty acid characteristics. The average daily gains were 201 g in the Dexter cattle and 104 g in the Engadine sheep. Both species achieved satisfactory carcass and meat quality and favourably low n-6/n-3 fatty acid ratios (0.63 and 0.68 in the intramuscular fat of the cattle and sheep, respectively). The meat from the Dexter cattle had a slightly higher cooking loss and shear force and a slightly lower proportion of fatty acids relevant to human health. Flavour appreciation was in favour of the burgers prepared from the beef.
Conclusions: Both robust breeds were suitable for producing meat on extensive pastures with A. viridis encroachment of up to 70%. Based on indirect evidence, it seems that neither species consumed significant quantities of leaves or bark from the trees.
Implications: Our study provides detailed information on rare robust breeds of two animal species in the context of meat production according to customer preferences. To better explain the differences between the two animal species, particularly in terms of fatty acid profiles, the role of the A. viridis understory needs to be clarified.
Keywords: beef, fatty acids, green alder, lamb, muscle fibre, ruminant, sensory quality, shrub.
Introduction
Grazing on high mountain pastures is considered beneficial with respect to meat quality, especially fatty acid (FA) composition (Ådnøy et al. 2005; Willems et al. 2014). Differences in the type and composition of forage plants have been shown to influence meat quality within such systems (Willems et al. 2013, 2014). Because of the higher energy expenditure due to the topography (Clapperton 1964), grazing at these sites usually comes at the cost of growth performance; however, body weight differences between species may influence this effect. Direct species comparisons on the same site are rare. One extant example is the study of sheep and goats performed by Giller et al. (2021), in which lamb meat was found to have a lower ultimate pH, cooking loss and shear force compared to that of goat kids. In other studies, data obtained from more than one experiment under different conditions have been compared, often with breeds of the same species. Willems et al. (2013, 2014) detected some differences in the carcass and meat quality and FA profiles of the body lipids of two different sheep breeds that had both grazed alpine pastures of different quality. The same was found for the carcass and meat quality of different beef breeds (autochthonous vs Piedmontese) obtained on pasture (Kreuzer et al. 2021).
Due to global warming and structural changes in agriculture, shrubs are increasingly encroaching on pastures worldwide, and biodiversity is thus being diminished (Wieczorkowski and Lehmann 2022). As a pioneer species, Alnus viridis (green alder) is the most important invasive shrub species in the Alps and dominates around 70% of successional shrub forests (reviewed in Zehnder et al. 2020). It is a fast-growing deciduous shrub that is widespread in the mountainous areas of the Northern Hemisphere (Caudullo et al. 2017). It forms dense interlocking stands, and little light reaches the ground. Although stands of A. viridis are densely covered with leaves, Bühlmann et al. (2016) reported leaf area indices of 3.3–4.5 m2 leaf area/m2 ground area for A. viridis stands, and the understory was not strongly shadowed because the lowest branches of the shrubs were well above the ground (cf. fig. 7d in Pauler et al. (2022)). A. viridis propagates by root suckers, which causes continuous expansion at stand edges, as well as by seeds, which leads to the colonisation of new sites. This has resulted in clusters of shrubs together in sections of the paddocks and random scatterings of shrubs throughout each paddock in other sections. A. viridis is able to fix atmospheric nitrogen, which causes problems of eutrophication (Svensk et al. 2023) and nitrate leaching (Bühlmann et al. 2016). Pollen records show that its appearance in the Alps coincided with signs of early pastoralism, which suggests that it is a native invader of open pastureland (Schwörer et al. 2014). Few studies have addressed how different livestock species and breeds deal with shrub-encroached mountain pastures. In a comparison to goats and cattle that consumed the leaves of A. viridis, Pauler et al. (2022) found that Engadine sheep, a robust mountain sheep breed, have a special preference for the bark of these shrubs. Farmers have reported that individuals among Dexter cattle, a small and robust British breed (Dexter Cattle Society 2021), seem to use woody plants as a feed resource. This breed was recently experimentally investigated (Giller et al. 2019) although not for this property. As shrub-encroached pastures are expected to provide forage of a lower density and limited nutritional quality, the growth performance and carcass quality (insufficient conformation and fat cover) of grazing animals may be impaired, whereas the FA profile of the meat lipids may shift to become particularly favourable in terms of human health.
We therefore tested the following hypotheses: (1) Increasing encroachment levels of the shrub A. viridis impair the growth and carcass characteristics of robust breeds like Dexter cattle and Engadine sheep due to decreasing biomass and forage quality. (2) Due to differences in foraging behaviours, digestion and metabolism, and growth and carcass characteristics, sheep and cattle respond differently to different levels of shrub encroachment. (3) There is an inherent species difference in meat quality between Dexter cattle and Engadine sheep even when grazing the same pasture types.
Materials and methods
Ethical statement
The experiment adhered to the national standards for animal welfare and was approved by the Committees for Animal Care and Use of the Cantons Zug and Grisons (ZG 70/15).
Experimental animals and sites
Twenty-two growing Dexter cattle (12 steers, 10 females; average age: 430 ± 4.7 days [mean ± s.e.] at the start of the experiment) and 20 weaned Engadine sheep (13 castrated males, 7 females; average age: 203 ± 2.5 days) were included in the study. For easier adaptation to the mountain environment, the lambs were accompanied by eight non-lactating adult female sheep, the data on which were not included in the present study. From 10 March 2015 onwards, the animals from each species were gathered in groups and received ad libitum access to hay and grass silage at 983 m above sea level (asl; farm site) at the AgroVet-Strickhof Research Station Früebüel, Zug, Switzerland.
On 19 June 2015, the animals were moved to four high-elevation experimental pastures with different levels of shrub encroachment. Based on the evaluated satellite photographs, the A. viridis cover (mean ± s.d.) was 0.8 ± 0.4%, 28.7 ± 2.2%, 50.3 ± 2.6% and 67.8 ± 3.2% of the total area at the different levels (n = 6). The animals were randomly split into four encroachment-level groups per species. The pastures with 0.8% or 28.7% encroachment levels were situated at 1950–2150 m asl and belonged to the AgroVet-Strickhof Research Station Weissenstein, Preda, Switzerland (46°34′54.5″N, 9°48′3.0″E). The pastures with encroachment levels of 50.3% and 67.8% were located at 1950–2150 m asl at Alp Drosa, La Punt Chamues-ch, Switzerland (46°34′53.3″, 9°54′5.8″E). The two sites were only 7 km apart. The shrub height was calculated using a drone-derived surface model (Pauler et al. 2022) and found to be between 1.5 and 5.4 m (90% confidence interval). The height (mean ± s.d.) was 3.3 ± 1.3 m and 3.0 ± 1.1 m for the paddocks grazed by the cattle and sheep, respectively.
Each of the four pastures at different encroachment levels was separated into six paddocks (24 in total). Among them, three were grazed twice in rotation by the cattle and three by the sheep. This set-up made it possible to move the animals to new paddocks every 10–12 days. The paddocks were delimited to obtain comparably intensive grazing pressure based on the biomass availability and estimated consumption. Forage availability was calculated based on the distribution of phytosociological units, as defined by Dietl (1977), and associated biomass yields depending on the elevation. Forage consumption was estimated based on the tabulated values of Agroscope (2014) for slow-growing cattle, growing lambs and adult sheep of a certain body weight. Accordingly, on average, the Dexter cattle and adult and growing Engadine sheep were assumed to consume 3.5, 1.5 and 1.0 kg forage dry matter per day, respectively. These considerations were the basis for allocating paddock sizes of between 1 and 1.5 ha to the Dexter cattle and 0.7 and 1 ha to the Engadine sheep. The total duration of grazing was set to 2 months, which was considered sufficient to discern the differences in the pasture treatments on the growth performance and carcass and meat quality of the animals. The grazing duration was 8 weeks for the cattle and 9 weeks for the sheep in line with the capacity limitations for the large number of animals that needed to be slaughtered and processed at the same time. During the entire experiment, the animals were only pasture-fed and finished on pasture without supplements.
Characterisation of the biomass
To generally determine the forage availability and quality, 30 1.2 m × 1.2 m animal exclusion cages were distributed in the main vegetation types stratified by elevation. The main vegetation types were nutrient-rich vegetation (phytosociological associations between Poion alpinae and Polygono–Trisetion), nutrient-poor vegetation (Nardion, Caricion curvulae and Seslerion) and understory vegetation, namely, the herb layer in the A. viridis stands (Alnenion viridis). The vegetation grown under each cage was cut twice towards the end of the two grazing rotations. The A. viridis leaves were not sampled, as they were assumed to make up only a small proportion of the diet. Their contribution to the diet was studied indirectly by comparing the growth performance of the animals grazing in paddocks at different levels of encroachment. The forage samples were dried for 48 h at 60°C, weighed, milled through a 1.0 mm screen and analysed by applying standard methods (AOAC International 1997). The contents of the dry matter (DM) and total ash were assessed using the thermogravimetric device TGA 701 (Leco Corporation, St Joseph, USA; AOAC Official Method 942.05). The crude protein was determined as nitrogen × 6.25 using a TruMac C/N-analyser (Leco Corporation, St. Joseph, USA; AOAC Official Method 968.06). The digestible organic matter was estimated in accordance with the method used by Tilley and Terry (1963).
Body weight assessment, slaughter, sampling and measurements performed at slaughter
Each animal was weighed on a transportable animal scale (Amman Waagen, Ermatingen, Switzerland) after each paddock change and at the end of the experiment. The cattle and sheep were slaughtered using a standard procedure on 16 and 23 August 2015, respectively. Conformation was scored by the Swiss system (cf. Giller et al. 2019) into nine classes (C, H, T–, T, T+, A, X1, X2, X3). For comparison with the existing literature, the results were translated into five classes (C, H, T, A, X). At 24 h p.m., the longissimus thoracis (LT) was sampled starting from between ribs 8 and 9 and the biceps femoris (BF) within 40 mm of its cranial extremity from the left carcass side. For later meat analysis, three 1.5 and three 3 cm slices of LT and BF were prepared and vacuum-packed. Some parts were stored at −20°C, and the remainder was aged at 2°C for 21 days. Finally, approximately 100 g of perirenal fat were dissected and frozen at −20°C. For further procedural details, see Gangnat et al. (2016).
Analysis of the physicochemical properties of the meat
The previously frozen and then homogenised meat from LT and BF were analysed for moisture, ash, protein (via the same methods as used for forage analysis) and intramuscular fat (IMF; AOAC Official Method 963.15). Further properties were determined of the intact slices of the LT and BF aged for 21 days. The colour space was measured with a Minolta Lab photometer, and drip loss was determined as the weight loss within 48 h (Honikel 1998). Cooking loss was assessed after heating 3 cm-thick aged slices for 45 min at 72°C in a water bath with subsequent cooling for 2 h at an ambient temperature. Afterwards, the shear force was determined in these slices by applying a Warner-Bratzler shear blade. Six meat cores with a diameter of 12.7 mm were obtained and sheared perpendicular to the muscle fibre direction after reaching room temperature (approximatively 20°C). For further details, see Gangnat et al. (2016).
Sensory analysis
Eight panellists were trained in four sessions on a standard protocol (ISO 2016) for the flavour and aroma attributes of burger patties. Patties of 100 g were prepared by thawing the meat at 4°C overnight, mincing and homogenising the LT and perirenal fat (ratio 80:20 g) for each individual animal using a standard burger press (diameter: 10.4 cm; thickness: 0.9 ± 0.1 cm). The burgers were grilled at 170°C for 90 s per side on a preheated aluminium plate. Each of the 42 burgers was cut into 1 cm3 cubes, covered with preheated inverted glass bowls on preheated plates and served to the panellists. The Dexter cattle and lamb burgers were evaluated in eight separate sessions for each species. Within the same session, two services of two samples from two different encroachment levels each were presented in a randomised manner. The samples were encoded using three-digit random numbers to enable double-blind testing. The panellists were served bread and tempered still water or light black tea after each sample to cleanse the palate. The common attributes determined in both species were the global flavour, liver flavour, sour flavour and global appreciation. The additional attributes were venison, rancid and blood flavour for the Dexter cattle and herb and barn flavour for the Engadine sheep. All the criteria were scored on a ten point scale ranging from 0 (lowest intensity) to 10 (highest intensity).
Muscle fibre typing
Transverse serial sections (10 μm thick) of the LT and BF were prepared using a Cryotome (Shandon Inc., Pittsburgh, USA). The muscle fibres were differentiated as slow-oxidative (type I), fast-oxidative-glycolytic (type IIA) and fast-glycolytic (type IIX/B) by their myosin adenosine triphosphatase activity based on the method of Solomon and Dunn (1988). Each section was first incubated at pH 9.4 and then at pH 4.6. Three stained sections (containing at least 100 muscle fibres) per muscle and animal were analysed. The fibres were differentiated by type and counted, and their area was measured using the software AnalySIS ver. 5.0 (Olympus Europa, Hamburg, Germany).
Analysis of the fatty acids in the intramuscular and perirenal fat
The FA profiles of the IMF (non-aged LT) and perirenal fat were analysed as per Wolf et al. (2019). Briefly, following extraction, the FA were converted into methyl esters in methanolic sodium hydroxide (NaOH) and boron triflouride. The FA quantification was performed by gas chromatography (HP 6890 with flame ionisation detector, Agilent Technologies, Wilmington, USA) using a CP7421 column (200 mm × 0.25 mm, 0.25 μm; Varian Inc., Lake Forest, USA). C11:0 triglyceride (Merck, Darmstadt, Germany) was used as the internal standard, and the peaks were identified via a 37-component FA methyl ester standard (Merck) and quantified using HP ChemStation® software (Agilent, Palo Alto, USA).
Statistical analyses
The data were subjected to analysis of variance by applying a linear mixed-effects model using R (R Core Team 2021) with individual animal as the experimental unit. The fixed effects were the level of shrub encroachment (as an ordered factor), and for the muscle fibre and meat quality traits, the animal species, the levels of shrub encroachment and the interactions of the animal species and encroachment levels. The random effects were the slaughter date and, with respect to the sensory data, the panellists. Equality of variance was inspected visually by comparing the standardised residuals to the fitted values. If heteroscedasticity was detected, the mixed model was updated with a weight argument in accordance with Pinheiro and Bates (2000, p. 177) to allow different variances for each factor level. The results were presented as arithmetic means and standard errors of the mean (s.e.m.). P < 0.05 was considered statistically significant and P < 0.10 as a trend. Multiple comparisons of the shrub encroachment levels and the animal species × shrub encroachment interaction means were performed using Tukey’s method.
Results and discussion
Forage quality
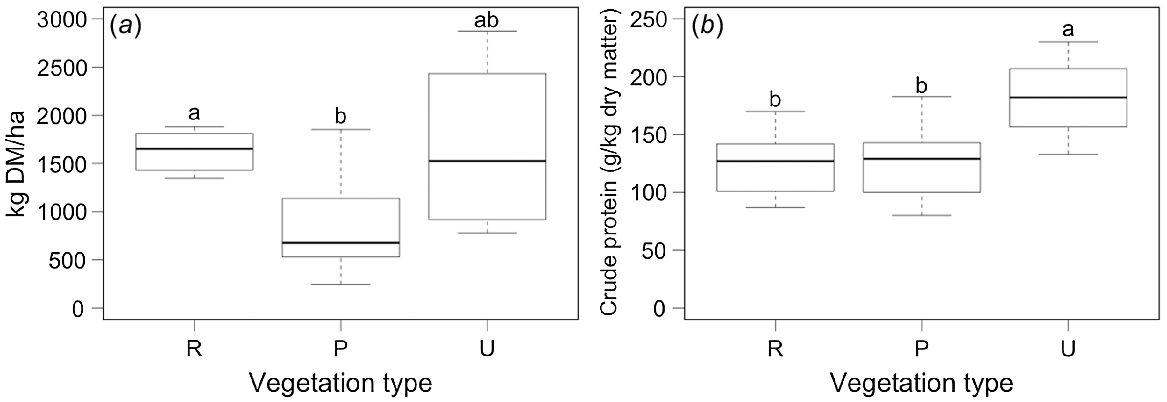
Studies on the impact of A. viridis on soil and biodiversity have previously been undertaken (cf. Zehnder et al. 2020), but little is known about the implications of its exploitation by grazing animals (Chassot and Deslandes 2009; Meisser et al. 2009). The exclusion cage data indicated that the average DM yield per ha was higher (P < 0.001) in the nutrient-rich vegetation than in the nutrient-poor vegetation but not in the shrub understory (P = 0.23) (Fig. 1a). The latter had a higher (P = 0.05) crude protein content than the other vegetation types (Fig. 1b). The content of the organic matter and digestible organic matter was not significantly different between the vegetation types (on average 931 and 527 g/kg DM, respectively). Accordingly, the A. viridis understory growing below the shrubs had quite a high and different feeding value compared to that of the open pastures. The higher crude protein content of the understory could be a result of the nitrogen (N) provided by the N2 fixing activity of A. viridis. It seems that the Dexter cattle were using this resource less intensively than the Engadine sheep, as they were found by Pauler et al. (2022) to stay in the A. viridis understory vegetation less frequently. A key to combating shrub encroachment levels could be more browsing on the leaves and bark of the shrubs, and especially of the bark. Piluzza et al. (2014) found that the high tannin content in materials from woody species deters herbivores. However, this obviously happens to a different degree in the two animal species investigated in the present study. Aside from Dexter cattle, Engadine sheep also have been found to consume both bark and leaves (Pauler et al. 2022). It was not possible to quantify the proportion of woody plant material in the diet in the present study, but we concluded from our results concerning animal performance that it did not contribute considerably to the total intake. In the case of debarking, the amount of woody material consumed does not need to constitute a significant proportion of the diet for shrubs to be severely hampered.
Dry matter yield and crude protein content of the forage determined within the exclusion cages. R: nutrient-rich vegetation (n = 6); P: nutrient-poor vegetation (n = 13); U: A. viridis understory vegetation (n = 4). Effect of vegetation type in (a): P = 0.013 and (b): P = 0.018. Boxplots with different letters within figure are significantly different at P < 0.05. Central lines describe the median, the boxes indicate first and third quartiles. Whiskers extend to the smallest and largest data points. Levels of s.e.m. for Fig. 1a: R, 91.2: P, 146; U, 476, and for Fig. 1b: R, 12.5; P, 8.9; U, 19.8.
Growth performance and carcass characteristics
The growth yield variables were not significantly influenced by shrub encroachment, even though the average daily gains differed numerically between the encroachment levels by as much as 100 g/day in the cattle (Table 1). The body weight changes in the animals during the experiment are illustrated in Supplementary Fig. S1. Even though the Dexter cattle were older than the Engadine sheep, at an average of 212 kg at slaughter, the Dexter cattle were far from reaching the anticipated mature weight of approximately 350 kg (females) to 500 kg (males; www.mutterkuh.ch/de/herdebuch/rasse/dexter). Overall, the growth data showed that the feeding value of the consumed forage was similar in the differently shrub-encroached paddocks. The dressing percentage remained fairly constant across the shrub encroachment levels for the cattle but increased in the sheep from the 0.8% shrub encroachment level to the 50.3% level. The shrub encroachment level had no effect on carcass conformation. The cattle had a lower fat cover score with the 50.3% shrub encroachment level compared to 28.7% shrub encroachment, whereas the sheep tended (P < 0.10) to have the highest fat cover score at 67.8% shrub encroachment. The differences in the responses to the shrub encroachment levels between the cattle and sheep were likely associated with differences in body and carcass weights. In general, the carcass conformation was favourable for both breeds. The conformation score of 5.3 (9-class CH-TAX system) found by Giller et al. (2019) for Dexter cattle resembled the 5.4 scored for those in our study (equivalent to 3.0 in the 5-class CH-TAX system). Chambaz et al. (2001, 2003) reported slightly higher scores (3.5 and 3.7; 5-class CH-TAX system) for beef and dual-purpose breeds (Angus or Simmental) that were fed indoors. The Engadine sheep in our study scored above the Willems et al. (2013) measurement for Engadine sheep on nutrient-rich lowland vegetation (3.3 vs 3.0, 5-class CH-TAX system). The unchanged fat cover score of approximately 2.5 in both animal species coincided with a similar dietary energy content. For the Dexter cattle, the fat cover score matched the 2.8 reported by Giller et al. (2019).
| Trait | Shrub encroachment (% of area) (E) | Average | s.e.m. | P-value | |||
|---|---|---|---|---|---|---|---|
| Animal species | 0.8 | 28.7 | 50.3 | 67.8 | E | ||
| Number of animals | |||||||
| Dexter cattle | 5 | 5 | 6 | 6 | |||
| Engadine sheep | 5 | 5 | 5 | 5 | |||
| Performance-related traits | |||||||
| Initial body weight (kg) | |||||||
| Dexter cattle | 209 | 206 | 204 | 207 | 207 | 5.32 | 0.96 |
| Engadine sheep | 34 | 36 | 35 | 34 | 35 | 1.09 | 0.92 |
| Average daily gains (g/day) | |||||||
| Dexter cattle | 188 | 243 | 230 | 139 | 201 | 21.46 | 0.36 |
| Engadine sheep | 138 | 87 | 118 | 113 | 114 | 8.13 | 0.14 |
| Slaughter-related traits | |||||||
| Age at slaughter (days) | |||||||
| Dexter cattle | 491 | 486 | 494 | 498 | 493 | 4.7 | 0.84 |
| Engadine sheep | 269 | 275 | 276 | 271 | 273 | 2.51 | 0.83 |
| Body weight at slaughter (kg) | |||||||
| Dexter cattle | 216 | 216 | 211 | 206 | 212 | 5.4 | 0.93 |
| Engadine sheep | 40 | 42 | 42 | 42 | 41 | 1.05 | 0.93 |
| Carcass weight (hot, kg) | |||||||
| Dexter cattle | 105 | 106 | 104 | 98 | 103 | 3.03 | 0.80 |
| Engadine sheep | 16 | 18 | 19 | 19 | 18 | 0.59 | 0.25 |
| Dressing percentage (%) | |||||||
| Dexter cattle | 48.8 | 49.1 | 49.0 | 47.5 | 48.6 | 0.31 | 0.19 |
| Engadine sheep | 39.7b | 43.3ab | 45.7a | 45.1a | 43.7 | 0.67 | <0.001 |
| Carcass conformation scoreA | |||||||
| Dexter cattle | 2.8 | 3.0 | 3.0 | 3.0 | 3.0 | 0.045 | 0.35 |
| Engadine sheep | 3 | 3.6 | 3.4 | 3.2 | 3.3 | 0.11 | 0.28 |
| Fat cover scoreB | |||||||
| Dexter cattle | 2.6ab | 2.8b | 2.0a | 2.5ab | 2.5 | 0.109 | 0.044 |
| Engadine sheep | 2.3 | 2.4 | 2.4 | 3.0 | 2.5 | 0.118 | 0.100 |
Means with different letters within line are significantly different at P < 0.05.
A1 = ‘very lean’ to 5 = ‘very well muscled’.
B1 = ‘not’ to 5 = ‘excessively covered with fat tissue’.
Muscle fibre type characteristics and physicochemical meat quality
The muscle fibre characteristics were not influenced by the shrub encroachment levels (Tables S1 and S2). One interaction (P = 0.029) of the animal species and shrub encroachment level occurred in the Type IIa mean fibre cross-sectional area of the BF. No significant effects of shrub encroachment were found in the physicochemical meat quality traits (Tables S3 and S4). These results indicate that the shrub encroachment levels had no substantial effect on the muscle fibre properties and physicochemical meat quality in either species.
Notwithstanding, the muscle fibre profile and physicochemical meat quality of the two species differed. The average muscle fibre number was lower (P < 0.001) in the cattle than the sheep for the LT and BF (Table 2). The mean fibre cross-sectional area (total, Type I, Type IIa and Type IIx/IIb) also differed between the species (P < 0.001) for the LT and BF, as did the proportions of the individual fibre types (at least P < 0.01), except for Type IIx/IIb in the LT. Aside from Type IIa in the LT (P = 0.83), the cross-sectional area per fibre type as a proportion of the total area differed (P < 0.001) between the species for all types and muscles. The cattle and sheep differed at least at P < 0.05 in both muscles in terms of colour (darker and less red for LT only and less yellow in the cattle than the sheep), shear force (higher in the cattle) and moisture content (higher in the BF of the cattle), protein (lower in the BF of the cattle) and ash (lower in the LT of the cattle) (Table 3). The cattle and sheep meat did not differ with respect to pH at 24 h post mortem in either muscle. Similar high cooking losses in two animal species have been reported for robust Valdostana cattle (Kreuzer et al. 2021), Dexter cattle and Charolais suckling calves (Giller et al. 2019), as well as Engadine sheep and Valaisan Black Nose sheep (Willems et al. 2013). However, in other studies carried out with different breeds under varying conditions, lower cooking losses were noted in the two muscles (Chambaz et al. 2003; Gangnat et al. 2016; Giller et al. 2021). Despite the generally higher shear force compared to the Engadine sheep meat in our study, the BF of the Dexter cattle qualified as tender (≤40 N; Branscheid et al. 2007). However, the LT of Dexter beef aged for 3 weeks had a surprisingly large shear force not previously found in this breed (Giller et al. 2019). The differences in the muscle fibre profiles between the two animal species may help interpret the differences in the water-holding capacity and tenderness. Wood et al. (1999) found higher amounts of Type I muscle fibres in tender cuts and indicated a generally higher tenderness of ‘red’ (Type I) vs ‘white’ (Type IIb) muscles. In our study, this only held true within the species, where higher proportions of Type I muscle fibres were associated with a lower shear force.
| Muscle | Longissimus thoracis | Biceps femoris | ||||||
|---|---|---|---|---|---|---|---|---|
| Animal species | Dexter cattle | Engadine sheep | s.e.m. | P-value | Dexter cattle | Engadine sheep | s.e.m. | P-value |
| Average number of fibres (n) | 178 | 337 | 15.8 | <0.001 | 160 | 298 | 13.0 | <0.001 |
| Mean fibre cross-sectional area (μm2) | ||||||||
| All | 1636 | 1046 | 118.8 | <0.001 | 1808 | 1244 | 115.8 | <0.001 |
| Type I | 1243 | 834 | 51.5 | <0.001 | 2009 | 1255 | 76.8 | <0.001 |
| Type IIa | 1947 | 964 | 89.2 | <0.001 | 1670 | 1157 | 60.1 | <0.001 |
| Type IIx/IIb | 1719 | 1336 | 62.6 | 0.003 | 1743 | 1321 | 60.2 | <0.001 |
| Fibre number (% of total fibres) | ||||||||
| Type I | 27.0 | 11.2 | 3.07 | <0.001 | 50.5 | 23.0 | 5.27 | <0.001 |
| Type IIa | 41.5 | 53.3 | 2.56 | <0.001 | 23.3 | 38.3 | 3.12 | <0.001 |
| Type IIx/IIb | 32.0 | 35.8 | 1.03 | 0.058 | 26.5 | 39.3 | 2.57 | 0.003 |
| Fibre area (% of total area) | ||||||||
| Type I | 19.3 | 8.4 | 1.05 | <0.001 | 52.9 | 22.5 | 2.65 | <0.001 |
| Type IIa | 46.6 | 46.0 | 1.04 | 0.83 | 20.3 | 34.6 | 2.04 | <0.001 |
| Type IIx/IIb | 31.6 | 42.2 | 1.33 | <0.001 | 24.1 | 39.1 | 2.06 | <0.001 |
The complete dataset concerning the individual species values grazing the pastures with increasing shrub encroachment are given in Tables S1 and S2.
| Muscle | Longissimus thoracis | Biceps femoris | ||||||
|---|---|---|---|---|---|---|---|---|
| Animal species | Dexter cattle | Engadine sheep | s.e.m. | P-value | Dexter cattle | Engadine sheep | s.e.m. | P-value |
| pH24 h post mortem | 5.77 | 5.76 | 0.029 | 0.83 | 5.73 | 5.78 | 0.026 | 0.37 |
| Colour | ||||||||
| Lightness (L*) | 34 | 37 | 0.3 | <0.001 | 32 | 37 | 0.4 | <0.001 |
| Redness (a*) | 21 | 25 | 0.4 | <0.001 | 22 | 25 | 1.1 | 0.19 |
| Yellowness (b*) | 2.0 | 4.9 | 0.30 | <0.001 | 2.2 | 3.8 | 0.18 | <0.001 |
| Water holding capacity | ||||||||
| Drip loss (%) | 1.40 | 1.64 | 0.067 | 0.070 | 1.76 | 1.27 | 0.069 | <0.001 |
| Cooking loss (%) | 28.3 | 26.0 | 0.67 | 0.097 | 30.4 | 28.9 | 0.28 | 0.012 |
| Shear force (N) | 65.6 | 40.4 | 2.26 | <0.001 | 39.5 | 31.8 | 1.18 | 0.001 |
| Chemical composition (g/100 g) | ||||||||
| Moisture | 73.4 | 73.5 | 0.18 | 0.90 | 74.9 | 74.2 | 0.17 | 0.051 |
| Protein | 21.7 | 21.5 | 0.12 | 0.33 | 20.1 | 21.8 | 0.17 | <0.001 |
| Fat | 2.38 | 2.38 | 0.136 | 0.32 | 2.73 | 2.30 | 0.147 | 0.18 |
| Total ash | 1.23 | 1.45 | 0.039 | 0.002 | 1.60 | 1.46 | 0.040 | 0.085 |
The complete dataset concerning the individual species values grazing the pastures with increasing shrub encroachment are given in Tables S3 and S4.
Our Dexter beef was darker, less red and less yellow compared to Limousin beef kept in the same vicinity (Gangnat et al. 2016), similar to the differences between the Dexter beef and Engadine sheep meat in our study. It therefore seems that the higher proportion of the ‘red’ muscle fibre type in the Dexter cattle compared to the sheep was not reflected in the redness of the meat. Nevertheless, the lightness and redness found for the Dexter cattle in the present study were more intense compared to most cuts offered to consumers by Carpenter et al. (2001). Consumers prefer beef to be a bright red rather than a darker and more purple colour (Carpenter et al. 2001) and are willing to pay more for such meat (Killinger et al. 2004). However, as grazing generally tends to produce darker meat compared to concentrate feeding, customers knowingly buying meat from such systems may even expect a darker colour (Preisinger et al. 1991; Allingham et al. 1998). This could also explain why Swiss Dexter breeders have reported the dark red colour as being particularly appealing to their customers. The LT and BF of the Engadine sheep with lightness (L*) at 37 and redness (a*) at 25 was at the lower end of what is desirable in terms of lightness but favourable in terms of redness according to the thresholds of Khliji et al. (2010).
Species comparisons of meat quality are extremely rare even though they may help in decision-making regarding the best way to preserve marginal lands and the ecosystem services they provide. The two breeds used in our study for this comparison have rarely been investigated. Giller et al. (2019) found similarities in the meat of Dexter cattle and suckling Charolais-crossbred calves with respect to water-holding capacity, shear force, nutrient composition and muscle fibre characteristics. This indicates that Dexter beef can compete with quality beef from production systems aimed at producing particularly high-quality meat. Willems et al. (2013) noted an even better quality of meat from Engadine sheep compared to Valaisian Black Nose sheep, another robust mountain breed, thus making the former likely comparable in terms of meat quality with meat-type sheep.
Sensory perception of the burger patties
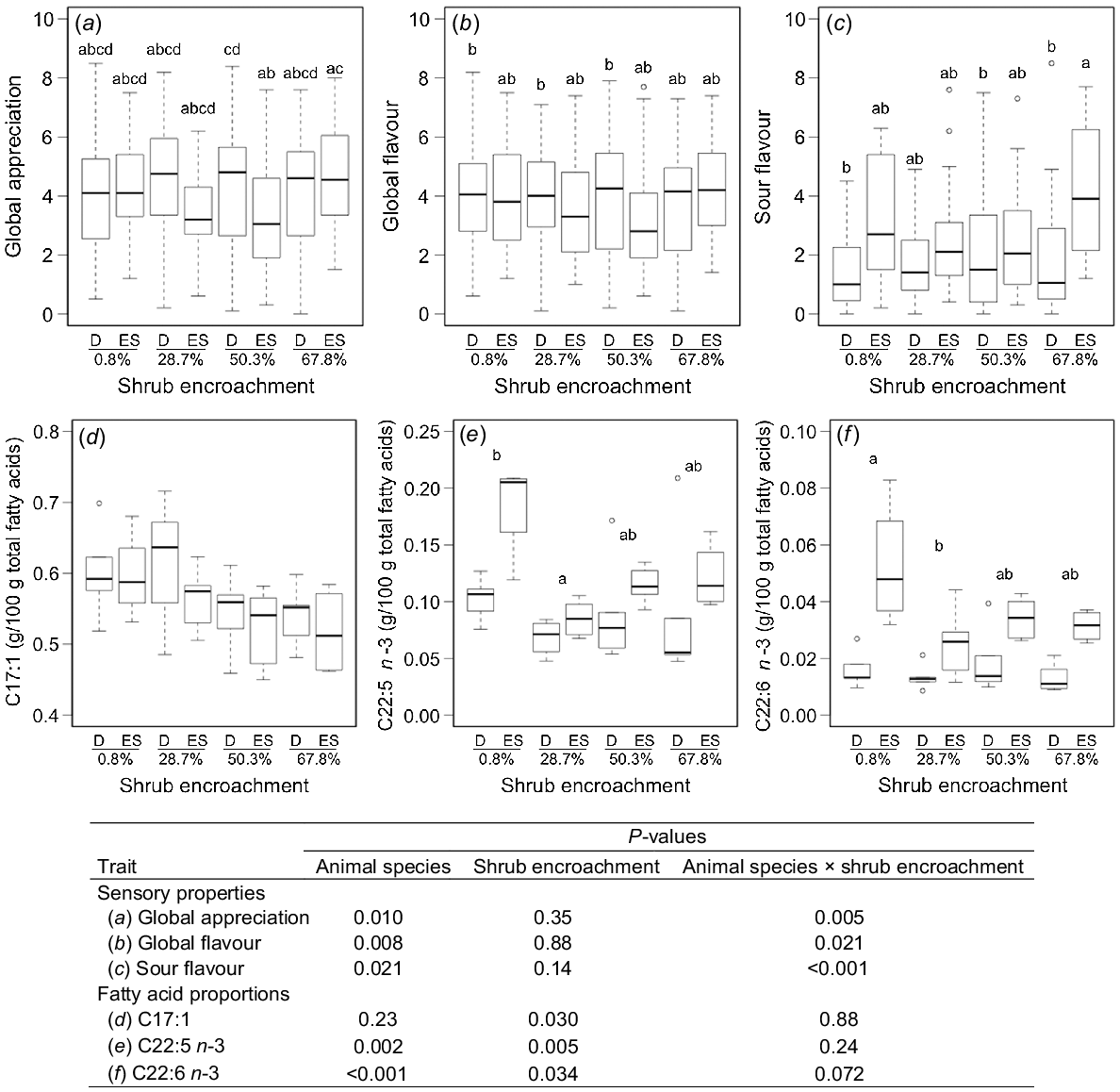
The global appreciation of the burger patties was highest at 28.7% shrub encroachment for the cattle and 67.8% for the sheep (interaction, P = 0.005; Fig. 2a). The high encroachment levels led to the lowest global flavour scores for the cattle and the highest scores for the sheep, whereas 50.3% shrub encroachment resulted in the highest scores for the cattle and the lowest scores for the sheep (interaction, P = 0.021; Fig. 2b). The sour flavour scores were lowest for the cattle and highest for the sheep at 67.8% shrub encroachment (interaction, P < 0.001; Fig. 2c). The lowest liver flavour scores for the cattle and the highest scores for the sheep coincided with 67.8% shrub encroachment (interaction, P = 0.056; Table 4). Despite these interactions, no single flavour trait was affected by the shrub encroachment levels alone across or within the species (Table 4, Fig. 2a–c). Overall, the global appreciation was higher (P = 0.01) for the cattle (4.32) than the sheep (3.87). Similarly, the global flavour scores were higher (P = 0.008) for the cattle (3.96) compared to the sheep (3.79). However, the sour flavour scores were lower (P = 0.021) for the cattle (1.71) than for the sheep (3.04). Considering the more frequent animal species effects, the shrub encroachment effects were uncertain and less important. The more pronounced sour flavour found in the patties from the Engadine sheep compared to those from the Dexter cattle could possibly explain the better global flavour and appreciation scores for the Dexter cattle patties. However, the traits indicating potential off-flavours scored very low in both species. In their breed comparison, Giller et al. (2019) also found differences in some traits with regard to the sensory perceptions of the meat from the Dexter cattle and suckling Charolais-crossbred calves. It needs to be considered, though, that the ratings of hedonic characteristics like global flavour and appreciation may be different when judged by a small number of trained panellists, as realised in our study, compared to a large number of untrained panellists from consumers of the target group.
Interaction of animal species (D, Dexter cattle; ES, Engadine sheep) and shrub encroachment level in distinct sensory properties of burger patties [(a) global appreciation, (b) global flavour, (c) sour flavour] and fatty acid proportions [(d) C17:1, (e) C22:5 n-3, (f) C22:6 n-3] in perirenal fat tissue. Central lines describe the median, the boxes indicate first and third quartiles. Whiskers extend to the data points within 1.5 times the length of the box. Values outside this range are shown as circles. Different lowercase letters indicate significant differences between interaction means (a, b, c) and shrub encroachment means (e, f)(P<0.05).
| Trait | Shrub encroachment (% of area) (E) | Average | s.e.m. | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Animal species | 0.8 | 28.7 | 50.3 | 67.8 | S | E | S × E | ||
| Liver flavour | 0.081 | 0.49 | 0.84 | 0.056 | |||||
| Dexter cattle | 1.26 | 1.50 | 1.40 | 1.16 | 1.32 | ||||
| Engadine sheep | 1.82 | 1.71 | 1.54 | 2.38 | 1.84 | ||||
| Dexter cattle only | |||||||||
| Venison flavour | 1.65 | 1.72 | 1.51 | 1.43 | 1.57 | 0.082 | ‒ | 0.53 | ‒ |
| Rancid flavour | 0.79 | 0.70 | 0.83 | 0.87 | 0.79 | 0.065 | ‒ | 0.76 | ‒ |
| Blood flavour | 0.90 | 1.23 | 1.30 | 1.11 | 1.12 | 0.075 | ‒ | 0.10 | ‒ |
| Engadine sheep only | |||||||||
| Herb flavour | 1.34 | 1.27 | 1.54 | 1.71 | 1.45 | 0.148 | ‒ | 0.69 | ‒ |
| Barn flavour | 2.68 | 2.18 | 2.18 | 2.24 | 2.31 | 0.157 | ‒ | 0.72 | ‒ |
Ranging from 0 = ‘lowest intensity’ to 10 = ‘highest intensity’.
Fatty acid profiles of the intramuscular and perirenal fat
The shrub encroachment levels did not significantly influence the FA proportions in the IMF, and only very few FA in the perirenal fat tissue (Tables S5 and S6). In the latter, the shrub encroachment level affected (P = 0.023) the C17:1 proportion (Fig. 2d). Excluding the non-significant interaction term from the model yielded trends in the C17:1 proportion that were different for the 0.8%, the 50.3% and the 67.8% shrub encroachment levels (P = 0.094 and P = 0.072, respectively). The proportions of C22:5 n-3 and C22:6 n-3 in perirenal fat were higher (P = 0.005 and P = 0.034, respectively) for the pastures with 0.8% shrub encroachment compared to 28.7% shrub encroachment (Fig. 2e, f). There was an interaction trend (P = 0.072) between the species and shrub encroachment levels in C22:6 n-3 (Fig. 2f). The consumption of woody plant material may have had an influence on the FA derived from ruminal biohydrogenation, as this process is modulated by plant secondary metabolites like tannins (Frutos et al. 2020). Accordingly, Leparmarai et al. (2021) found higher proportions of monounsaturated and polyunsaturated fatty acids (MUFA and PUFA) in camel milk compared to cattle milk when the animals were grazing wood pastures. The debarking activity of the Engadine sheep, in addition to the leaf browsing activity practiced by both animal species (Pauler et al. 2022), may therefore be of importance. Still, even the Engadine sheep apparently did not consume sufficient woody plant material to change the FA profile of their meat. Some of the long-chain PUFA (C22:5 n-3 and C22:6 n-3), which are assumed to be protected by the tannins from ruminal biohydrogenation, even decreased in proportion in the perirenal fat with increasing shrub encroachment levels. It is possible that the high crude protein content of the A. viridis understory levelled out the effects of the tannins against the ruminal biohydrogenation, as Gerson et al. (1986) showed an in vitro inhibition of the biohydrogenation of FA by the low crude protein content of the forage.
Direct comparisons of the FA composition of beef and sheep meat, as performed in the present study, are very rare. One exception is the study by Purchas et al. (2014), where cuts were obtained at the same time and analysed in the same laboratory. The FA composition data of the meat from the cattle and sheep listed in the national food composition databases provide no information about the production systems, animal feed composition, age at slaughter or other factors, and the data for the same cuts largely deviate (cf. for instance, the New Zealand Institute for Plant and Food Research Limited and Ministry of Health 2019). Overall, in the present study, the animal species that grazed the same types of pasture had a clearly greater influence on the FA profile of the IMF and perirenal fat than the shrub encroachment levels. An astonishingly high number of FA in the IMF and perirenal fat differed significantly in their proportions between the animal species. The species differences were even more pronounced in magnitude in the perirenal fat than in the IMF (Table 5). More specifically, at the same total saturated fatty acid (SFA) proportion, like that described by Purchas et al. (2014) when related to total FA, the SFA C10:0 and C18:0 were lower in proportion in the Dexter cattle than the Engadine sheep. The proportions of the medium-chain SFA, which are particularly unfavourable with respect to human health (Daley et al. 2010), were similar (C12:0) or higher (C14:0, C16:0) in the perirenal fat of the Dexter cattle. The differences were less clear for the IMF. The Dexter cattle lipids had higher proportions of MUFA and lower proportions of PUFA than those of the sheep. In contrast, the proportion of MUFA was similar between the beef and lamb meat in the study of Purchas et al. (2014); these authors found lower PUFA proportions for cattle, as was the case in the present study. The FA associated with ruminal microbial activity (uneven chain FA and trans biohydrogenation intermediates) and the n-6/n-3 ratios were either less favourable for the Dexter cattle (C18:1 t11, C18:2 n-6, C18:3 n-3, n-6/n-3 ratio) or did not differ significantly between the species (C17:1, C18:2 c9,t11). Only C15:1 was higher in the perirenal fat of the Dexter cattle. The proportions of the n-3 FA and n-6/n-3 ratios were also less favourable for the cattle than the sheep in the comparison by Purchas et al. (2014). The reasons for the species differences may be a different plant selection behaviour (the possible effect of woody plants has been discussed above), a different metabolic set-up for FA synthesis and transformation or a different rumen microbiome possibly already formed before the experiment. For both animal species, the meat n-6/n-3 ratios were well within the recommended ranges of 1:1 up to 5:1 (Simopoulos et al. 2000) and much lower than the ratios of 15:1 to 20:1 assumed to be characteristic for Western diets (Simopoulos and De Meester 2009). In other species comparisons, with the exception of a lower C18:1 c9 proportion, Giller et al. (2021) and Sheridan et al. (2003) both found a higher proportion of individual C18 FA in sheep compared to goats. The similarity of the species differences found in our study with that by Purchas et al. (2014) indicates that most of the differences between the proportions of groups and important individual FA are indeed inherent species differences and not from non-genetic factors or breed differences. The latter was confirmed by Willems et al. (2014). In their study, the FA proportions were found to differ little between the breeds Engadine sheep and Valaisian Black Nose sheep.
| Muscle | Longissimus thoracis | Perirenal fat tissue | ||||||
|---|---|---|---|---|---|---|---|---|
| Animal species | Dexter cattle | Engadine sheep | s.e.m. | P-value | Dexter cattle | Engadine sheep | s.e.m. | P-value |
| C8:0 | 0.013 | 0.013 | 0.0008 | 0.83 | 0.009 | 0.017 | 0.0009 | <0.001 |
| C10:0 | 0.047 | 0.113 | 0.0058 | <0.001 | 0.046 | 0.112 | 0.0066 | <0.001 |
| C12:0 | 0.082 | 0.139 | 0.0074 | <0.001 | 0.165 | 0.171 | 0.0115 | 0.79 |
| C14:0 | 2.65 | 2.08 | 0.098 | 0.003 | 4.72 | 2.92 | 0.216 | <0.001 |
| C15:1 | 0.179 | 0.167 | 0.0039 | 0.167 | 0.340 | 0.310 | 0.0074 | 0.047 |
| C16:0 | 24.4 | 20.2 | 0.46 | <0.001 | 26.2 | 18.3 | 0.718 | <0.001 |
| ∑ 16:1A | 3.74 | 1.96 | 0.163 | <0.001 | 2.90 | 2.02 | 0.088 | <0.001 |
| C17:0 | 1.57 | 1.95 | 0.051 | <0.001 | 1.68 | 1.98 | 0.037 | <0.001 |
| C17:1 | 0.672 | 0.638 | 0.0120 | 0.16 | 0.573 | 0.549 | 0.0103 | 0.23 |
| C18:0 | 15.4 | 19.9 | 0.44 | <0.001 | 25.2 | 34.4 | 0.96 | <0.001 |
| C18:1 c9 | 31.7 | 29.3 | 0.46 | 0.007 | 25.8 | 21.0 | 0.62 | <0.001 |
| C18:1 t11 | 1.49 | 2.66 | 0.121 | <0.001 | 1.84 | 3.76 | 0.210 | <0.001 |
| Further C18:1 isomersB | 2.37 | 2.51 | 0.046 | 0.12 | 3.58 | 5.26 | 0.200 | <0.001 |
| C18:2 n-6 | 4.86 | 6.72 | 0.291 | 0.001 | 1.20 | 2.55 | 0.118 | <0.001 |
| C18:2 c9t11 | 0.012 | 0.014 | 0.0006 | 0.14 | 0.443 | 0.452 | 0.0195 | 0.83 |
| C18:2 t11c15 | 0.286 | 0.403 | 0.0180 | <0.001 | 0.488 | 0.730 | 0.0364 | <0.001 |
| C18:3 n-3 | 2.00 | 3.30 | 0.131 | <0.001 | 0.77 | 1.85 | 0.095 | <0.001 |
| Further C18:3 isomersC | 4.84 | 6.68 | 0.293 | 0.001 | 1.18 | 2.30 | 0.099 | <0.001 |
| C20:3 n-3 | 0.039 | 0.049 | 0.0018 | 0.006 | 0.032 | 0.037 | 0.0014 | 0.067 |
| C20:4 n-6 | 1.43 | 1.48 | 0.090 | 0.78 | 0.03 | 0.04 | 0.003 | 0.075 |
| C20:5 n-3 | 0.855 | 1.040 | 0.0613 | 0.14 | 0.054 | 0.018 | 0.0035 | <0.001 |
| C22:4 n-6 | 0.013 | 0.016 | 0.0009 | 0.049 | 0.029 | 0.034 | 0.0020 | 0.17 |
| C22:5 n-3 | 0.946 | 0.916 | 0.0485 | 0.78 | 0.086 | 0.124 | 0.0072 | 0.002 |
| C22:6 n-3 | 0.124 | 0.318 | 0.0193 | <0.001 | 0.013 | 0.035 | 0.0024 | <0.001 |
| ∑ Saturated fatty acids | 44.7 | 45.0 | 0.37 | 0.77 | 59.4 | 59.1 | 0.46 | 0.75 |
| ∑ Monounsaturated fatty acids | 40.9 | 37.4 | 0.56 | <0.001 | 35.7 | 33.0 | 0.51 | 0.007 |
| ∑ Polyunsaturated fatty acids | 11.6 | 15.1 | 0.62 | 0.005 | 3.5 | 6.3 | 0.26 | <0.001 |
| ∑ n-3 | 4.22 | 5.71 | 0.231 | 0.001 | 0.99 | 2.09 | 0.099 | <0.001 |
| n-6:n-3 = x:1 | 0.625 | 0.679 | 0.0115 | 0.17 | 0.674 | 0.740 | 0.0129 | 0.007 |
The complete dataset concerning the individual species values grazing the pastures with increasing shrub encroachment are given in Tables S5 and S6.
AIncludes C16:1 c7, C16:1 c9, C16:1 t9.
BIncludes C18:1 t6-12, C18:1 c10-13, C18:1 c15.
CIncludes C18:3 n-6, C18:3 2n6c.
Conclusions
Both robust breeds reached satisfactory growth and carcass and meat quality to allow for viable meat production on pastures with shrub encroachment of up to 70% of the area, which disproved hypothesis (1). This opens up new opportunities for sustainable meat production without competing with direct human nutrition.
We found only a few significant interactions between the species and shrub encroachment levels, which thus only partly confirmed hypothesis (2).
The Engadine sheep and Dexter cattle differed in terms of many meat quality characteristics, which confirmed hypothesis (3). The Dexter cattle were superior to the sheep with respect to sensory characteristics, and in general, the beef was preferred over the lamb meat. However, the beef had a higher cooking loss and shear force compared to the lamb meat, even though these properties mostly remained in the range of what consumers deem acceptable. The fatty acids relevant to human health were slightly inferior in proportion in the Dexter cattle compared to those found in the Engadine sheep.
Demonstrating the advantages and disadvantages of the two animal alternatives provides information for producers to determine the optimal choice. As other studies have shown, Engadine sheep also have the benefit of performing debarking activity. In our study, indirect evidence suggested that neither the sheep nor the cattle consumed significant quantities of leaves or bark from the trees.
The role of the A. viridis understory in shrub-encroached areas needs to be investigated.
Acknowledgements
We are grateful to the municipality of La Punt Chamues-ch, Ralph Fluor and Jörg Pleisch for providing the site, breeders of the SwissDexters association, the Erny, Hossli and Schriber families for loaning their animals, and team of the AgroVet-Strickhof research station Alp Weissenstein for the help with animals and the hospitality. We owe thanks to Raphael Gago, Cornel Stutz and Markus Staudinger for valuable assistance in the field, the employees of the animal hospital Zürich for slaughtering and Carmen Kunz from ETH Zurich for the help in the laboratory. The great number of valuable suggestions for improving the manuscript made by the Associate Editor and the Reviewer are highly appreciated.
References
Ådnøy T, Haug A, Sørheim O, Thomassen MS, Varszegi Z, Eik LO (2005) Grazing on mountain pastures-does it affect meat quality in lambs? Livestock Production Science 94, 25-31.
| Crossref | Google Scholar |
Allingham PG, Harper GS, Hunter RA (1998) Effect of growth path on the tenderness of the semitendinosus muscle of Brahman-cross steers. Meat Science 48, 65-73.
| Crossref | Google Scholar |
Bühlmann T, Körner C, Hiltbrunner E (2016) Shrub expansion of Alnus viridis drives former montane grassland into nitrogen saturation. Ecosystems 19, 968-985.
| Crossref | Google Scholar |
Carpenter CE, Cornforth DP, Whittier D (2001) Consumer preferences for beef color and packaging did not affect eating satisfaction. Meat Science 57, 359-363.
| Crossref | Google Scholar |
Caudullo G, Welk E, San-Miguel-Ayanz J (2017) Chorological maps for the main European woody species. Data in Brief 12, 662-666.
| Crossref | Google Scholar |
Chambaz A, Morel I, Scheeder MRL, Kreuzer M, Dufey P-A (2001) Characteristics of steers of six beef breeds fattened from eight months of age and slaughtered at a target level of intramuscular fat – I. Growth Performance and carcass quality. Archives Animal Breeding 44, 395-412.
| Crossref | Google Scholar |
Chambaz A, Scheeder MRL, Kreuzer M, Dufey P-A (2003) Meat quality of Angus, Simmental, Charolais and Limousin steers compared at the same intramuscular fat content. Meat Science 63, 491-500.
| Crossref | Google Scholar |
Chassot A, Deslandes KA (2009) Nutztiere PASTO: Tierleistungen und Wirtschaftlichkeit. AGRAR Forschung 16, 308-313.
| Google Scholar |
Clapperton JL (1964) The energy metabolism of sheep walking on the level and on gradients. British Journal of Nutrition 18, 47-54.
| Crossref | Google Scholar |
Daley CA, Abbott A, Doyle PS, Nader GA, Larson S (2010) A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutrition Journal 9, 10.
| Crossref | Google Scholar |
Dexter Cattle Society (2021) Breed history. Available at https://www.dextercattle.co.uk/the-breed/breed-history [Verified 27 July 2022]
Dietl W (1977) Vegetationskunde als Grundlage der Verbesserung des Graslandes in den Alpen. In ‘Application of vegetation science to grassland husbandry’. (Ed. W Krause) pp. 405–458. (Springer Netherlands: Dordrecht) doi:10.1007/978-94-010-1315-4_9
Frutos P, Hervás G, Natalello A, Luciano G, Fondevila M, Priolo A, Toral PG (2020) Ability of tannins to modulate ruminal lipid metabolism and milk and meat fatty acid profiles. Animal Feed Science and Technology 269, 114623.
| Crossref | Google Scholar |
Gangnat IDM, Kreuzer M, Clavijo McCormick A, Leiber F, Berard J (2016) Carcass and meat quality of finished and non-finished Limousin heifers from alpine livestock systems differing in altitudinal origin of the forage. Archives of Animal Nutrition 70, 108-126.
| Crossref | Google Scholar |
Gerson T, John A, King ASD (1986) Effects of feeding ryegrass of varying maturity on the metabolism and composition of lipids in the rumen of sheep. The Journal of Agricultural Science 106, 445-448.
| Crossref | Google Scholar |
Giller K, Gangnat IDM, Silacci P, Messadene-Chelali J, Kreuzer M, Berard J (2019) Meat and carcass quality of Dexter cattle compared with that of suckler, Charolais-cross calves: a preliminary study. Animal Production Science 59, 986-992.
| Crossref | Google Scholar |
Giller K, Sinz S, Messadene-Chelali J, Marquardt S (2021) Maternal and direct dietary polyphenol supplementation affect growth, carcass and meat quality of sheep and goats. Animal 15, 100333.
| Crossref | Google Scholar |
Honikel KO (1998) Reference methods for the assessment of physical characteristics of meat. Meat Science 49, 447-457.
| Crossref | Google Scholar |
ISO(2016) Method 13299 Sensory analysis – methodology – general guidance for establishing a sensory profile. (International Organization for Standardization [ISO]: Geneva, Switzerland). Available at https://www.iso.org/standard/58042.html [Verified 8 May 2023]
Khliji S, van de Ven R, Lamb TA, Lanza M, Hopkins DL (2010) Relationship between consumer ranking of lamb colour and objective measures of colour. Meat Science 85, 224-229.
| Crossref | Google Scholar |
Killinger KM, Calkins CR, Umberger WJ, Feuz DM, Eskridge KM (2004) Consumer visual preference and value for beef steaks differing in marbling level and color. Journal of Animal Science 82, 3288-3293.
| Crossref | Google Scholar |
Kreuzer M, Pervier S, Turille G, Karpatcheva M, Julius N, Oreiller C, Berard J (2021) Beef quality in two autochthonous Valdostana breeds fattened in alpine transhumance: effect of lowland finishing and meat ageing. Italian Journal of Animal Science 20, 267-278.
| Crossref | Google Scholar |
Leparmarai PT, Kunz C, Mwangi DM, Gluecks I, Kreuzer M, Marquardt S (2021) Camels and cattle respond differently in milk phenol excretion and milk fatty acid profile to free ranging conditions in East-African rangelands. Scientific African 13, e00896.
| Crossref | Google Scholar |
Meisser M, Tarery M, Chassot A, Freléchoux F (2009) Nutztiere Weidemanagement und Verhalten der Rinder in stark verbuschtem Gelände. AGRAR Forschung 16, 408-413.
| Google Scholar |
New Zealand Institute for Plant and Food Research Limited and Ministry of Health (2019) ‘The concise New Zealand food composition tables’. 13th edn. (New Zealand Institute for Plant and Food Research Limited and Ministry of Health) Available at www.foodcomposition.co.nz/concise-tables
Pauler CM, Zehnder T, Staudinger M, Lüscher A, Kreuzer M, Berard J, Schneider MK (2022) Thinning the thickets: foraging of hardy cattle, sheep and goats in green alder shrubs. Journal of Applied Ecology 59, 1394-1405.
| Crossref | Google Scholar |
Piluzza G, Sulas L, Bullitta S (2014) Tannins in forage plants and their role in animal husbandry and environmental sustainability: a review. Grass and Forage Science 69, 32-48.
| Crossref | Google Scholar |
Preisinger R, Kalm E, Guhe M, Schmidt G, Augustini C, Henning M (1991) Is it possible to produce high quality beef with young bulls? Carcass composition and beef quality of bulls, oxen and heifers in different production systems. Tierzuechter 43, 437-440.
| Google Scholar |
Purchas RW, Wilkinson BHP, Carruthers F, Jackson F (2014) A comparison of the nutrient content of uncooked and cooked lean from New Zealand beef and lamb. Journal of Food Composition and Analysis 35, 75-82.
| Crossref | Google Scholar |
R Core Team (2021) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/
Schwörer C, Henne PD, Tinner W (2014) A model-data comparison of Holocene timberline changes in the Swiss Alps reveals past and future drivers of mountain forest dynamics. Global Change Biology 20, 1512-1526.
| Crossref | Google Scholar |
Sheridan R, Hoffman LC, Ferreira AV (2003) Meat quality of Boer goat kids and Mutton Merino lambs 2. Sensory meat evaluation. Animal Science 76, 73-79.
| Crossref | Google Scholar |
Simopoulos AP, De Meester F (Eds) (2009) ‘A balanced omega-6/omega-3 fatty acid ratio, cholesterol and coronary heart disease.’ (S. Karger AG) 10.1159/isbn.978-3-8055-9225-3
Simopoulos AP, Leaf A, Salem N (2000) Workshop on the essentiality of and recommended dietary intakes for omega-6 and omega-3 fatty acids. Food Reviews International 16, 113-117.
| Crossref | Google Scholar |
Solomon MB, Dunn MC (1988) Simultaneous histochemical determination of three fiber types in single sections of ovine, bovine and porcine skeletal muscle. Journal of Animal Science 66, 255-264.
| Crossref | Google Scholar |
Svensk M, Pittarello M, Mariotte P, Nota G, Schneider MK, Frund D, Dubois S, Allan E, Probo M (2023) Nitrogen translocation by Highland cattle grazing in Alnus viridis-encroached pastures. Nutrient Cycling in Agroecosystems 126, 127-141.
| Crossref | Google Scholar |
Tilley JMA, Terry RA (1963) A two-stage technique for the in vitro digestion of forage crops. Grass and Forage Science 18, 104-111.
| Crossref | Google Scholar |
Wieczorkowski JD, Lehmann CER (2022) Encroachment diminishes herbaceous plant diversity in grassy ecosystems worldwide. Global Change Biology 28, 5532-5546.
| Crossref | Google Scholar |
Willems H, Kreuzer M, Leiber F (2013) Vegetation-type effects on performance and meat quality of growing Engadine and Valaisian Black Nose sheep grazing alpine pastures. Livestock Science 151, 80-91.
| Crossref | Google Scholar |
Willems H, Kreuzer M, Leiber F (2014) Alpha-linolenic and linoleic acid in meat and adipose tissue of grazing lambs differ among alpine pasture types with contrasting plant species and phenolic compound composition. Small Ruminant Research 116, 153-164.
| Crossref | Google Scholar |
Wolf C, Gredig N, Ulbrich SE, Kreuzer M, Berard J, Giller K (2019) Partitioning of rumen-protected n-3 and n-6 fatty acids is organ-specific in growing Angus heifers. Lipids 54, 503-517.
| Crossref | Google Scholar |
Wood JD, Enser M, Fisher AV, Nute GR, Richardson RI, Sheard PR (1999) Manipulating meat quality and composition. Proceedings of the Nutrition Society 58, 363-370.
| Crossref | Google Scholar |
Zehnder T, Lüscher A, Ritzmann C, Pauler CM, Berard J, Kreuzer M, Schneider MK (2020) Dominant shrub species are a strong predictor of plant species diversity along subalpine pasture-shrub transects. Alpine Botany 130, 141-156.
| Crossref | Google Scholar |