

Availability to ruminants of nitrogen in senesced C4 tropical grasses
R. M. Dixon A * and R. J. Mayer B
A * and R. J. Mayer B
A The University of Queensland, QAAFI, St Lucia, Qld 4067, Australia.
B Queensland Department of Agriculture and Fisheries, Maroochy Research Facility, PO Box 5083, SCMC, Nambour, Qld 4560, Australia.
Animal Production Science 63(3) 269-278 https://doi.org/10.1071/AN22197
Submitted: 19 May 2022 Accepted: 26 September 2022 Published: 28 October 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Nutritional standards usually assume that ∼10% of the total nitrogen (TN) in forages is indigestible and hence not available to the ruminant. Senesced tropical C4 grasses often contain TN concentrations that are marginal or deficient to meet the nutrient requirements of ruminants, and low TN availability will exacerbate N deficiencies.
Aim: The aim of the study was to estimate the availability (i.e. digestibility) to ruminants of TN in mature and senesced C4 grasses, using data derived from previous experiments and published data.
Methods: In Dataset 1, forages grown in subtropical or tropical environments, including C4 (n = 143) and C3 (n = 15) grasses, were analysed for parameters including TN, acid detergent fibre (ADF), and N insoluble in ADF solution (ADIN). ADIN was used as a measure of unavailable TN. The Dataset 2 analysis included published measurements of TN and ADIN in C4 (n = 187) and C3 (n = 45) grasses.
Key results: In Dataset 1, TN averaged 9.7 and ADIN 1.45 g N/kg diet DM in C4 grasses. ADIN concentration could be predicted from TN and ADF concentrations by multiple regression (R2 = 0.50; P < 0.001). The ratio ADIN/TN averaged 170 g/kg and increased exponentially with increasing ADF concentration (R2 = 0.43; P < 0.001). Also, ADIN/TN was inversely related to TN concentration and DM digestibility. In C4 grasses containing >400 and >500 g ADF/kg DM, ADIN/TN averaged 190 and 230 g/kg, respectively, and in those containing <10 g TN/kg DM, ADIN/TN averaged 194 g/kg. In these low-quality C4 grasses only ∼80%, and as little as ∼50%, of TN was available to ruminants. Dataset 2 produced similar results, although C4 grass forages were generally of higher quality, most having been harvested during vegetative growth. ADIN/TN was much lower in C3 grasses (89 g/kg).
Conclusions: In senesced, low-quality C4 grasses containing >400 g ADF, <10 g TN or <530 g digestible DM/kg, the proportion of TN available to ruminants is substantially lower than that assumed for forages in general.
Implications: Low availability of TN in many senesced C4 grasses needs to be considered when evaluating the adequacy of dietary TN for ruminants grazing senesced tropical grass pastures.
Keywords: acid detergent insoluble N, C4 grasses, forage digestibility, forage N, low quality forages, N-deficiency, ruminants, tropical forages.
Introduction
There has been extensive investigation of ruminant digestion of the total nitrogen (TN) fractions in forages to understand their breakdown and use as substrates by rumen microbes and, following passage of some forage N to the post-ruminal tract, whether various fractions are digested or are excreted in faeces (Webster 1987; AFRC Technical Committee on Responses to Nutrients 1992). This has often been investigated through use of surgically prepared animals to partition the sites of digestion, and synthetic fibre bags to measure digestion in the rumen and post-ruminally. A laboratory approach to estimating the amounts and proportions of the TN fractions in forages has been used to determine the solubility of TN under various specified conditions (Sniffen et al. 1992; Licitra et al. 1996). There is consensus that the forage N that is insoluble in acid detergent fibre (ADF) solution (i.e. ADIN) represents the TN that is not digested in the rumen or post-ruminally, and hence provides a reliable measure of the TN that is indigestible and not available to the ruminant (Van Soest 1982; Broderick 1994). It is generally agreed that ∼90% of the TN consumed in forages is digested and thus available in ruminants (Webster et al. 1982; AFRC Technical Committee on Responses to Nutrients 1992; Sniffen et al. 1992). These approaches have, with some variations, been incorporated into evaluations of the nutritional value of feeds and feeding standards (AFRC Technical Committee on Responses to Nutrients 1992; CSIRO 2007; National Academies of Sciences, Engineering and Medicine 2016).
Investigation of the digestion of forage TN fractions has focused on improved and sown species of grasses and legumes during vegetative growth, or soon after physiological maturity, when forages are usually grazed or harvested in higher input ruminant production systems (Wilson and Strachan 1981; Waters et al. 1992; Wales et al. 1999; National Academies of Sciences, Engineering and Medicine 2016). Relatively little information is available on the digestion of TN fractions in mature, senesced and weathered grasses, particularly C4 grasses, that usually have low TN concentrations and are highly fibrous. Nevertheless, this is important in some low-input rangeland systems, and crop–livestock systems in low-income countries, where ruminants are highly dependent on senesced C4 grasses and stovers of low nutritional value as feeds during some seasons of the annual cycle. Also, there is evidence that the N availability in at least some C4 grass hays is lower than in C3 grass hays of comparable low nutritional value (Hogan et al. 1989; Hogan 1996). Because senesced forages, particularly C4 grasses, are usually marginal or deficient in TN for ruminants, it is important to understand the availability of this TN to the ruminant.
The present study examined the concentrations of TN, N insoluble in neutral detergent fibre (NDF) solution (i.e. NDIN), ADIN, and fibre components in C4 grasses, as well as in some C3 grasses and legumes, grown in subtropical or tropical environments of northern Australia. Forages sampled from a wide range of seasons and sites were used to examine the hypothesis that a substantial proportion of the TN in low-quality tropical C4 grasses is present as indigestible ADIN. In addition, the study collated published measurements of TN and ADIN in C4 grasses grown in the subtropics or tropics.
Materials and methods
Dataset 1: forages analysed for N fractions
The forages analysed (n = 194) were obtained from previous experiments. Most (n = 184) had been fed to cattle in individual pens during the development of faecal NIRS calibration equations (Coates and Dixon 2011) or in other, later pen experiments (Kennedy and Charmley 2012; McLennan 2014; RM Dixon and DB Coates, unpublished data) where both the forage and faeces were sampled. The forages were all mechanically harvested from pastures growing in the subtropics or tropics. The forages were considered as classes comprising native or improved species of C4 grasses, improved species of C3 grasses, tropical legumes and temperate legumes (Table 1). Subclasses were forage species represented by ≥10 samples, or combinations of several species when there were fewer samples. The native species of C4 grass forages were generally cut with a forage harvester from swards of native pastures and classified into three subclasses: (1) dominated by Heteropogon contortus (black speargrass), (2) dominated by Astrebla spp. (Mitchell grasses), or (3) ‘mixed native C4 grasses’. The mixtures of species included H. contortus, Bothriochloa pertusa (Indian couch) and Chrysopogon fallax (golden beard grass), and sometimes Urochloa mosambicensis that had invaded native grass paddocks. Hays of the introduced, sown C4 grass species Cenchrus ciliaris and Chloris gayana were obtained from commercial farms, and the U. mosambicensis was harvested from swards dominated by this species. The subclass ‘Other improved C4 grasses’ combined such grasses where there were <10 samples of a species; these included Pennisetum spp. (millets), Setaria spp., Brachiaria mutica (para grass), Cynodon dactylon (couch) and Stenotaphrum secundatum (buffalo couch). The C3 grass forages were the cereal crop species Avena sativa (oats, n = 6), Triticum spp. (wheat, n = 3) and Hordeum vulgare (barley; n = 1), all in the vegetative growth stage. The tropical legume Stylosanthes included both Stylosanthes scabra and S. hamata, and ‘Other herbaceous legumes’ included Lablab purpureus (dolichos), Neonotonia wightii (glycine), Centrosema pascuorum (cavalcade), Clitoria ternatea (butterfly pea), Macroptilium bracteatum (burgundy bean) and Arachis pintoi (peanut). The temperate legume was Medicago sativa (lucerne). The C3 grasses and the legumes had been cut as hays on commercial farms.

|
The forage samples were analysed for TN by Kjeldahl analyses. NDF and ADF contents, and NDIN and ADIN, were analysed as described by Pichard and van Soest (1977) and Licitra et al. (1996). The dry matter digestibility (DMD) of the forages was measured by near-infrared reflectance spectroscopy of faeces (F.NIRS) as described by Coates and Dixon (2011).
Dataset 2: collation of published results reporting ADIN content of C4 grasses
Data were collated from published papers reporting measurements of the TN and ADIN fractions in C4 grasses, or mixtures containing predominantly C4 grasses (n = 187), grown in the subtropics or tropics. Where a study also reported measurements in C3 grasses (n = 45) or tropical herbaceous legumes expected not to contain tannins (n = 51) and grown in the same environment, those results were included. Measurements of wheat or rice straws as senesced C3 grasses in the subtropics were also included for comparison. One additional study with C4 grasses (Buckner et al. 2013), in which ‘total tract indigestible protein’ was measured using disappearance from synthetic fibre bags incubated in the rumen and during passage through the post-ruminal gastrointestinal tract, was also considered. Some of the papers also reported ADF, NDF and NDIN concentrations (105, 120 and 77 of the 187 C4 grass forage samples, respectively). Samples from treatments within experiments where the pasture had been fertilised with N were excluded. The papers had been published by research groups in North America (n = 11), South America (n = 4), India (n = 7), Africa (n = 4), and elsewhere (n = 5), and often the C4 grass forages had been measured at various maturity stages during the summer growing season. Genera most often reported and the number of samples were Cynodon (31), Cenchrus (21), Panicum (20), Andropogon (16), Digitaria (12), Pennisetum (11) Brachiaria, (9), Zea mays (8), Eragrostis (7), Paspalum (8), Setaria (5) and Sorghum (5). The dataset included one experiment where C. dactylon pasture had been ‘stockpiled’ after physiological maturity and then sampled several times over 3 months (Scarbrough et al. 2002), and another experiment with hay made from mature Andropogon spp. (‘bluestem range mixed forage’) (Hannah et al. 1991). One paper reported the changes in two C4 and two C3 grass species in the same environment through the summer growth period (Mitchell et al. 1997). Another study reported measurements in a range of grass species during the dry season in a southern African semi-arid environment (Gemeda and Hassen 2014). Nine papers included herbaceous tropical legumes (23 species and 51 samples). The forages were considered as groups comprising C4 grasses, C3 grasses, tropical legumes and temperate legumes.
Calculations and statistical analyses
Each class of forage within each dataset was divided into six categories of ADIN concentration (<1.00, 1.00–1.49, 1.50–1.99, 2.00–2.99, 3.00–3.99 and >4.00 g/kg DM) and six categories of ADIN/TN (<100, 100–149, 150–199, 200–299, 300–399 and >400 g/kg). In Dataset 1, the attributes of the various subclasses of forages were compared by one way analysis of variance. The relationships between the variables were examined by regression. The statistical package used was Genstat Release 16.1 (VSN International, Hemel Hempstead, UK).
Results
Dataset 1: measured concentrations of N fractions and DMD
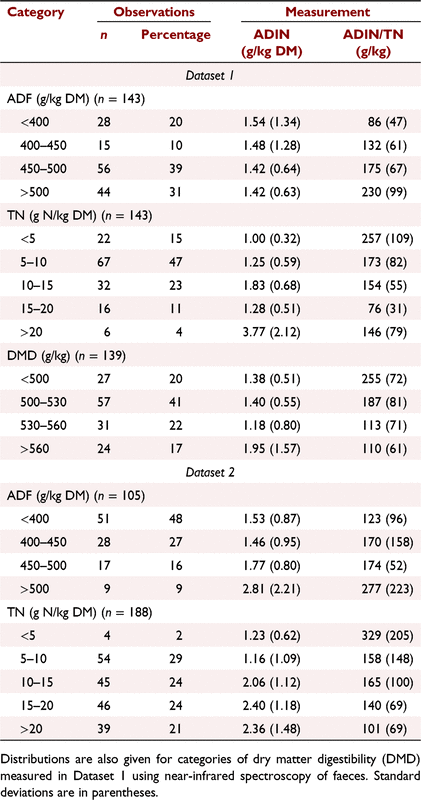
The measured forage attributes generally did not differ (P > 0.05) among subclasses within the C4 native and C4 introduced grass species, or between these classes, and there was no consistent pattern (Table 1). Overall, the C4 grasses (n = 143) averaged 9.7 g TN/kg DM, 1.45 g ADIN/kg DM, 170 g ADIN/kg TN, and (n = 139) 526 g digestible DM/kg, but each of these attributes had large variation and range. The C3 grass and both legume classes were generally much higher in TN and ADIN concentrations, but they had lower ADIN/TN ratios, than the C4 grasses. However, due to the small number of samples and few species, the C3 grasses and temperate legumes did not broadly represent these classes of forages. The tropical legumes had high concentrations of TN and ADIN. The distributions of the categories of ADIN and ADIN/TN ratio in the forage classes in Dataset 1 are shown in Fig. 1. Among all C4 grasses, 44% contained ≥1.50 g ADIN/kg DM and 62% had a ratio of ≥150 g ADIN/kg TN; among C4 grasses with >400 g ADF/kg DM, 46% contained ≥1.50 g ADIN/kg DM and 74% had a ratio of ≥150 g ADIN/kg TN.

|
Many of the measured forage attributes were correlated (P < 0.01), but the correlation coefficients (r) were generally <0.7 and did not allow useful prediction of one variable from another (Table 2). TN was negatively correlated with NDF and ADF (r = −0.81 and −0.77), and was positively correlated with DMD (r = 0.85). ADIN concentration was not correlated with either NDF or ADF content (both P > 0.05), and was only poorly correlated with TN and NDIN (r = 0.42 and 0.65, respectively).

|
Among the C4 grass forages, the ADIN/TN ratio increased exponentially with increasing ADF content (Fig. 2a). Forages containing <400, 400–450, 450–500 and >500 g ADF/kg DM had, on average, ratios of 86, 132, 175 and 230 g ADIN/kg TN, respectively (Table 3). Conversely, increasing TN concentration or DMD was associated with decreasing ADIN/TN ratio (Fig. 2b, c). In forages containing <5 g TN/kg DM, the ADIN/TN ratio averaged 257 (s.d. 109) g/kg, and in forages containing 5–10 g TN/kg DM, the ADIN/TN ratio averaged 173 (s.d. 82) g/kg (Table 3). As the TN concentration in forages increased to 20 g TN/kg DM, the ADIN/TN ratio decreased to 154 and then 76 g/kg. There was very large variation in the ADIN/TN ratio within each of these categories, especially at low forage TN concentrations. Similar large variation and changes between categories were also observed for four categories of DMD (<500, 500–530, 530–560 and >560 g/kg) (Table 3). In summary, these relationships indicated that the ADIN/TN ratio was increasing, and thus availability of the TN to the ruminant was decreasing, with decreasing nutritional value of the C4 grass forages. ADIN (g/kg DM) and ADIN/TN ratio (g/kg) could be estimated using multiple regression models:
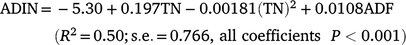
and:


|
|
Dataset 2: collation of published results
The distributions of the categories of ADIN concentration and ADIN/TN ratio in the various forage classes of Dataset 2 are shown in Fig. 3, and the ADIN concentrations and ADIN/TN ratios in Table 3. On average, the C4 grasses (n = 187) contained 1.93 g ADIN/kg DM and had a ratio of 147 g ADIN/kg TN. The more fibrous grasses with >400 g ADF/kg DM contained, on average, 1.78 g ADIN/kg DM and had a ratio of 189 g ADIN/kg TN, with 55% recording a ratio of ≥150 g ADIN/kg TN. The C4 grasses containing <400 g ADF/kg DM averaged 1.53 g ADIN/kg DM and 123 g ADIN/kg TN. By comparison, 43% of C4 grasses with <10 g N/kg DM had a ratio of ≥150 g ADIN/kg TN. In the entire sample set, the concentrations of ADIN were comparable in C4 grasses, C3 grasses and tropical legumes (1.93, 1.78 and 2.63 g ADIN/kg DM, respectively), but because the TN concentration was usually lower in the C4 grasses, the proportion of forages with ratios ≥150 g ADIN/kg TN was higher among C4 grasses (41%) than C3 grasses (19%) or tropical legumes (17%). In summary, the profiles of forage constituents in this dataset derived from published measurements by many research groups showed concentrations and trends similar to those observed in Dataset 1.

Discussion
Concentrations and proportions of ADIN as indigestible N
There is consensus that, in grass and herbaceous legume forages, ADIN represents the N that is bound to plant cell walls, is indigestible and not available to the ruminant, and that it typically comprises ∼10% of the TN (Van Soest 1982; Webster et al. 1988; AFRC Technical Committee on Responses to Nutrients 1992; Broderick 1994). Dataset 1 in the present study clearly showed that in low-quality, senesced C4 grasses the ADIN/TN ratio was substantial, averaging 190 g ADIN/kg TN in those with >400 g ADF/kg, and similar ratios in those with ≤10 g TN/kg or ≤560 g digestible DM/kg. Importantly, the ADIN/TN ratio in these groups was highly variable, ranging from negligible up to 495 g ADIN/kg TN, and was more closely correlated with ADF than with TN or DMD (Table 2). This is in agreement with associations between ADIN concentration and ADF content reported by Webster et al. (1982), AFRC Technical Committee on Responses to Nutrients (1992) and Waters et al. (1992). The efficacy of low TN concentrations or low DMD for identifying samples with high dietary ADIN/TN ratio was likely due primarily to the associations among these attributes.
The samples collated from published results as Dataset 2 (Table 3) generally included higher quality C4 grass forages than Dataset 1 (Table 3). This was most obviously because the forages in Dataset 2 were largely harvested during their vegetative growth rather than after senescence. However, among the C4 grasses in Dataset 2 that contained >400 g ADF/kg or <10 g TN/kg, the ADIN/TN ratio was comparable to the same classes of C4 grasses in Dataset 1.
The results are in accord with other evidence of low rumen availability of the TN in some low-quality C4 grasses, from observations of expectedly low rumen ammonia concentrations in sheep fed such grasses (Hogan et al. 1989; Hogan 1996). In summary, there is considerable evidence to support the hypothesis that a substantial proportion of the TN in low-quality C4 grasses is present as ADIN and therefore indigestible in ruminants.
Variation in ADIN concentration and ADIN/TN ratio among the forages
From the present study and other published studies, it is not possible to separate the effects of factors such as genotype, growth environment and stage of growth of the grasses on ADIN concentration or ADIN/TN ratio. Both Blasi et al. (1991) and Redfearn et al. (1995) reported that during vegetative growth both ADIN and ADIN/TN were consistently higher in two C4 grasses (Andropogon gerardii and Panicum virgatum) than in a C3 grass (Bromus inermis), and the former study also found that these attributes could vary widely between years. However, contrasting results were reported by Mitchell et al. (1997), who studied two C3 grasses and two C4 grasses grown in the same environment during one summer. First, values of ADIN/kg DM and ADIN/TN were generally higher than reported in the two studies cited above, and also increased with plant maturity; at the greatest maturity examined, there was a range from 226 to 408 g ADIN/kg TN among these C3 and C4 grasses. Second, there were large differences among the grass species such that values of ADIN/kg DM and ADIN/TN ranked in the order: A. gerardii (C4) > P. virgatum (C4) ≈ B. inermis (C3) > Thinopyrum intermedium (C3). In this experiment the differences between the two grass species within each of the C3 and C4 classes could be as great as differences between C4 and C3 classes. There is evidence that higher ADIN concentration and ADIN/TN ratio occur with low soil fertility and/or unfavourable environmental conditions for grass growth; Johnson et al. (2001) reported that the ADIN/TN ratio was generally low (<120 g N/kg TN) in C4 grasses (C. dactylon, Cynodon nlemfuensis and Paspalum notatum) grown in a favourable environment. Differences between C3 and C4 grasses and changes with increasing plant maturity may be associated with the different anatomical structures and greater proportions of parenchymal bundle sheaths and epidermis in both the leaf and stem of the C4 grasses (Wilson 1994), and with the generally higher content of ADF in C4 grasses (Laetsch 1974; Akin and Burdick 1975; Akin 1989). It is clearly important to understand the circumstances under which the ADIN/TN ratio is high and TN is low in C4 grass forages, but further investigation is needed to understand the reasons for the variation in the ADIN/TN ratio.
Limitations of the present study for examining the relationships between ADIN/TN ratio and other forage attributes were that neither sample set was designed for that purpose, and that the reported measurements were limited. The forages in Dataset 1 were harvested from pasture swards that usually contained a mix of C4 perennial grass species in various stages of regrowth, or were hays obtained from commercial farms. The growth and physiological maturity of the grasses sampled varied with the grass species and with interactions such as with soils and rainfall events to provide a broad array of forages as required for the development of the F.NIRS calibrations. Similarly, Dataset 2 collating results for 305 forages from 30 published papers did not provide a rigorous or balanced sample set. Nevertheless the present study showed that, on average, lower quality C4 grass forages containing ∼>400 g ADF/kg, ∼<10 g TN/kg or ∼<530 g digestible DM/kg had much higher ADIN/TN ratios than generally found in higher quality C4 grass forages, C3 grass forages or legumes. It was also clear that among the low-quality C4 grasses there was great variability in the ADIN/TN ratio, ranging from 11 to 495 g ADIN/kg TN. Wilson (1982, 1994) has provided excellent qualitative descriptions of the structure and anatomy of C4 grasses, C3 grasses and dicotyledonous forages and how these are likely to be influenced by plant part, age of individual leaves and stem, water stress and temperature. However, it was not possible to relate such factors to the species, growing conditions or anatomical structure of the grasses in Datasets 1 and 2. High soil N availability during grass growth would be expected to increase the TN concentration and proportion of soluble N; however, as stated in the Materials and methods, the forages in Dataset 1 were generally not from fertilised soils. Also Dataset 2 did not include samples from treatments where the grass had been fertilised with N. It seems likely that better understanding of effects of forage species and environment will explain at least some of the variation in ADIN/TN ratio among C4 grasses and provide the reasons why this ratio is high in highly fibrous C4 grasses. This requires further investigation. Until further information is available it seems reasonable that for senesced C4 grasses, defined, for example, as containing >400 g ADF/kg, <10 g TN/kg or <530 g digestible DM/kg, nutritional recommendations should assume that 20–30% of the TN is not available to the ruminant.
Importance of indigestible N in forages for ruminant production systems
The finding that a substantial proportion of the TN in mature C4 grasses is indigestible will be most important in circumstances where diets are based on such forages that are marginal or deficient in N. One such situation is in low-input, low-output tropical production systems where ruminants are highly dependent on physiologically mature or senesced C4 grass forages for many months of the annual cycle. This often occurs in rangeland production systems in the seasonally dry tropics of the northern Australian, African and South American rangelands where the only pastures available during the dry season are physiologically mature or senesced and weathered (Winks 1984; Hogan 1996). Many of these C4 grass forages, such as those in the present Dataset 1 and in the database of Norton (1982), contain <10 g N/kg DM and have low DMD, so that the ADIN/TN ratio is expected to be high. In addition, Aumont et al. (1995) reported that in a large, normally distributed database of forages from the humid tropics in the Caribbean and La Reunion (n = 1313), the means and standard deviations indicated that ∼30% of the forages contained >400 g ADF/kg DM or <11 g N/kg DM. High ADIN/TN ratios are also likely to occur in C4 crop residues (e.g. maize, sorghum and millet stovers), which are important feedstuffs in many African crop–livestock systems (Duncan et al. 2013, 2016; Dejene et al. 2022). Senesced tropical pastures or crop residues are usually deficient in TN for ruminants (Winks et al. 1979; Hennessy and Williamson 1990; Kennedy et al. 1992; Coates and Dixon 2008; Dixon and Coates 2010). Lower than expected availability of the TN in such forages will require reconsideration of the need for, and responses of ruminants to, supplementary rumen-degradable N. In low-input systems such as those described above, the provision of N supplements usually incurs substantial increases in input costs and necessary management skills, and especially where urea non-protein N is used to provide supplementary N. The development and availability of F.NIRS in ruminants to measure diet attributes (Stuth et al. 1999; Coates 2004; Dixon and Coates 2009, 2015) provides opportunity for commercial farms to routinely monitor the ADF, TN, DMD and metabolisable energy of the diet of grazing cattle, and to use these measurements to estimate the proportion of indigestible N.
Conclusions
The study established that in highly fibrous (>400 g ADF/kg DM) and low TN (<10 g TN/kg DM) C4 grass forages, the ADIN/TN ratio was highly variable, ranging from negligible to ∼50%, but averaging ∼20%. This greater than expected proportion of indigestible forage TN bound to plant cell walls requires consideration when evaluating the adequacy of TN in the diets of ruminants depending on such forages. Further investigation is needed to identify the grass species, environmental conditions and circumstances where the proportion of indigestible ADIN in TN in such forages is of practical nutritional importance.
Data availability
The data that support this study may be shared upon reasonable request to the corresponding author if appropriate.
Conflicts of interest
The authors declare no conflict of interest. Dr R M Dixon is an Associate Editor of Animal Production Science but had no role in the review or evaluation of the manuscript.
Declaration of funding
The research did not receive any specific funding.
Acknowledgements
We thank Mr David Coates, Dr Peter Kennedy and Dr Stuart McLennan for making available forage samples from their experiments. We also thank Mr Michael Gravel and DAF technical staff for undertaking the laboratory analyses.
References
AFRC Technical Committee on Responses to Nutrients (1992) Report No. 9. Nutritive requirements of ruminant animals: Protein. Nutrition Abstracts and Reviews (Series B) 62, 787–835.Akin DE (1989) Histological and physical factors affecting digestibility of forages. Agronomy Journal 81, 17–25.
| Histological and physical factors affecting digestibility of forages.Crossref | GoogleScholarGoogle Scholar |
Akin DE, Burdick D (1975) Percentage of tissue types in tropical and temperate grass leaf blades and degradation of tissues by rumen microorganisms. Crop Science 15, 661–668.
| Percentage of tissue types in tropical and temperate grass leaf blades and degradation of tissues by rumen microorganisms.Crossref | GoogleScholarGoogle Scholar |
Aumont G, Caudron I, Saminadin G, Xande A (1995) Sources of variation in nutritive values of tropical forages from the Caribbean. Animal Feed Science and Technology 51, 1–13.
| Sources of variation in nutritive values of tropical forages from the Caribbean.Crossref | GoogleScholarGoogle Scholar |
Blasi DA, Ward JK, Klopfenstein TJ, Britton RA (1991) Escape protein for beef cows: III. Performance of lactating beef cows grazing smooth brome or big bluestem. Journal of Animal Science 69, 2294–2302.
| Escape protein for beef cows: III. Performance of lactating beef cows grazing smooth brome or big bluestem.Crossref | GoogleScholarGoogle Scholar |
Bowen MK (2004) Efficiency of microbial protein production in cattle grazing tropical pastures. PhD Thesis, The University of Queensland, St Lucia, Qld, Australia.
Broderick GA (1994) Quantifying forage protein quality. In ‘Forage quality, evaluation and utilization’. (Ed. GC Fahey) pp. 200–228. (American Society of Agronomy: Madison, WI, USA)
Brown WF, Pitman WD (1991) Concentration and degradation of nitrogen and fibre fractions in selected tropical grasses and legumes. Tropical Grasslands 25, 305–312.
Buckner CD, Klopfenstein TJ, Rolfe KM, Griffin WA, Lamothe MJ, Watson AK, MacDonald JC, Schacht WH, Schroeder P (2013) Ruminally undegradable protein content and digestibility for forages using the mobile bag in situ technique. Journal of Animal Science 91, 2812–2822.
| Ruminally undegradable protein content and digestibility for forages using the mobile bag in situ technique.Crossref | GoogleScholarGoogle Scholar |
Chaurasia M, Kundu SS, Singh S, Misra AK (2006) Cornell net carbohydrate and protein system for nutritional evaluation of tree leaves, shrubs and grasses. Indian Journal of Animal Science 76, 81–87.
Coates DB (2004) Improving reliability of faecal NIRS calibration equations. Final Report of Project NAP3.121. Meat and Livestock Australia, Sydney.
Coates DB, Dixon RM (2008) Faecal near infrared reflectance spectroscopy estimates of diet quality and responses to nitrogen supplements by cattle grazing Bothriochloa pertusa pastures. Australian Journal of Experimental Agriculture 48, 829–834.
| Faecal near infrared reflectance spectroscopy estimates of diet quality and responses to nitrogen supplements by cattle grazing Bothriochloa pertusa pastures.Crossref | GoogleScholarGoogle Scholar |
Coates DB, Dixon RM (2011) Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages. Journal of Near Infrared Spectroscopy 19, 507–519.
| Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages.Crossref | GoogleScholarGoogle Scholar |
Coblentz WK, Turner JE, Ogden RK, Coffey KP, Pohlman FW, Brown AH, Daniels MB, Gunsaulis JL, Thomas ML, Wells CA, Morrow RE (2004) Case study: estimating degradable intake protein in warm- and cool-season forages grown on producer farms in northern Arkansas. The Professional Animal Scientist 20, 443–452.
| Case study: estimating degradable intake protein in warm- and cool-season forages grown on producer farms in northern Arkansas.Crossref | GoogleScholarGoogle Scholar |
CSIRO (2007) ‘Nutrient requirements of domesticated ruminants.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Das LK, Kundu SS, Kumar D, Datt C (2015) Fractionation of carbohydrate and protein content of some forage feeds of ruminants for nutritive evaluation. Veterinary World 8, 197–202.
| Fractionation of carbohydrate and protein content of some forage feeds of ruminants for nutritive evaluation.Crossref | GoogleScholarGoogle Scholar |
da Silva SP, Rodrigues MT, Vieira RAM, da Silva MMC (2013) In vitro degradation kinetics of protein and carbohydrate fractions of selected tropical forages. Bioscience Journal 29, 1300–1310.
Dejene M, Dixon RM, Walsh KB, McNeill D, Seyoum S, Duncan AJ (2022) High-cut harvesting of maize stover and genotype choice can provide improved feed for ruminants and stubble for conservation agriculture. Agronomy Journal 114, 187–200.
| High-cut harvesting of maize stover and genotype choice can provide improved feed for ruminants and stubble for conservation agriculture.Crossref | GoogleScholarGoogle Scholar |
Dixon R, Coates D (2009) Review: Near infrared spectroscopy of faeces to evaluate the nutrition and physiology of herbivores. Journal of Near Infrared Spectroscopy 17, 1–31.
| Review: Near infrared spectroscopy of faeces to evaluate the nutrition and physiology of herbivores.Crossref | GoogleScholarGoogle Scholar |
Dixon RM, Coates DB (2010) Diet quality estimated with faecal near infrared reflectance spectroscopy and responses to N supplementation by cattle grazing buffel grass pastures. Animal Feed Science and Technology 158, 115–125.
| Diet quality estimated with faecal near infrared reflectance spectroscopy and responses to N supplementation by cattle grazing buffel grass pastures.Crossref | GoogleScholarGoogle Scholar |
Dixon RM, Coates DB (2015) Application of faecal near infrared spectroscopy to manage the nutrition and productivity of grazing ruminants. In ‘Proceedings of the first international conference on forages in warm climates’. (Eds AR Evangelista, CLS Avila, DR Casagrande, MAS Lara, TF Bernardes) pp. 207–230. (Universidade Federal de Lavras: Lavras, MG, Brazil)
Duncan AJ, Teufel N, Mekonnen K, Singh VK, Bitew A, Gebremedhin B (2013) Dairy intensification in developing countries: effects of market quality on farm-level feeding and breeding practices. Animal 7, 2054–2062.
| Dairy intensification in developing countries: effects of market quality on farm-level feeding and breeding practices.Crossref | GoogleScholarGoogle Scholar |
Duncan AJ, Bachewe F, Mekonnen K, Valbuena D, Rachier G, Lule D, Bahta M, Erenstein O (2016) Crop residue allocation to livestock feed, soil improvement and other uses along a productivity gradient in Eastern Africa. Agriculture, Ecosystems & Environment 228, 101–110.
| Crop residue allocation to livestock feed, soil improvement and other uses along a productivity gradient in Eastern Africa.Crossref | GoogleScholarGoogle Scholar |
Erasmus LJ, Prinsloo J, Botha PM, Meissner HH (1990) Establishment of a ruminal protein degradation data base for dairy cattle using the in situ polyester bag technique. South African Journal of Animal Science 20, 130–135.
Fondevila M, Nogueira-Filho JCM, Barrios-Urdaneta A (2002) In vitro microbial fermentation and protein utilisation of tropical forage legumes grown during the dry season. Animal Feed Science and Technology 95, 1–14.
| In vitro microbial fermentation and protein utilisation of tropical forage legumes grown during the dry season.Crossref | GoogleScholarGoogle Scholar |
Foster JL, Lamb GC, Tillman BL, Marois JJ, Wright DL, Maddox MK (2012) In sacco degradation kinetics of fresh and field-cured peanut (Arachis hypogaea L.) forage harvested at different maturities. Animal Feed Science and Technology 171, 52–59.
| In sacco degradation kinetics of fresh and field-cured peanut (Arachis hypogaea L.) forage harvested at different maturities.Crossref | GoogleScholarGoogle Scholar |
Gemeda BS, Hassen A (2014) In vitro fermentation, digestibility and methane production of tropical perennial grass species. Crop and Pasture Science 65, 479–488.
| In vitro fermentation, digestibility and methane production of tropical perennial grass species.Crossref | GoogleScholarGoogle Scholar |
Gupta A, Singh S, Kundu SS, Jha N (2011) Evaluation of tropical feedstuffs for carbohydrate and protein fractions by CNCP system. Indian Journal of Animal Science 81, 1154–1160.
Hannah SM, Cochran RC, Vanzant ES, Harmon DL (1991) Influence of protein supplementation on site and extent of digestion, forage intake, and nutrient flow characteristics in steers consuming dormant bluestem-range forage. Journal of Animal Science 69, 2624–2633.
| Influence of protein supplementation on site and extent of digestion, forage intake, and nutrient flow characteristics in steers consuming dormant bluestem-range forage.Crossref | GoogleScholarGoogle Scholar |
Hennessy DW, Williamson PJ (1990) Feed intake and liveweight of cattle on subtropical native pasture hays. 1. The effect of urea. Australian Journal of Agricultural Research 41, 1169–1177.
| Feed intake and liveweight of cattle on subtropical native pasture hays. 1. The effect of urea.Crossref | GoogleScholarGoogle Scholar |
Hogan JP (1996) Options for manipulating nutrition if feed supply is immutable. Australian Journal of Agricultural Research 47, 289–305.
| Options for manipulating nutrition if feed supply is immutable.Crossref | GoogleScholarGoogle Scholar |
Hogan JP, Kennedy PM, McSweeney CS, Schlink AC (1989) Quantitative studies of the digestion of tropical and temperate forages by sheep. Australian Journal of Experimental Agriculture 29, 333–337.
| Quantitative studies of the digestion of tropical and temperate forages by sheep.Crossref | GoogleScholarGoogle Scholar |
Johnson CR, Reiling BA, Mislevy P, Hall MB (2001) Effects of nitrogen fertilization and harvest date on yield, digestibility, fiber, and protein fractions of tropical grasses. Journal of Animal Science 79, 2439–2448.
| Effects of nitrogen fertilization and harvest date on yield, digestibility, fiber, and protein fractions of tropical grasses.Crossref | GoogleScholarGoogle Scholar |
Juarez Lagunes FI, Fox DG, Blake RW, Pell AN (1999) Evaluation of tropical grasses for milk production by dual-purpose cows in tropical Mexico. Journal of Dairy Science 82, 2136–2145.
| Evaluation of tropical grasses for milk production by dual-purpose cows in tropical Mexico.Crossref | GoogleScholarGoogle Scholar |
Kabuga JD, Darko CA (1993) In sacco degradation of dry matter and nitrogen in oven dried and fresh tropical grasses and some relationships to in vitro dry matter digestibility. Animal Feed Science and Technology 40, 191–205.
| In sacco degradation of dry matter and nitrogen in oven dried and fresh tropical grasses and some relationships to in vitro dry matter digestibility.Crossref | GoogleScholarGoogle Scholar |
Kennedy PM, Charmley E (2012) Methane yields from Brahman cattle fed tropical grasses and legumes. Animal Production Science 52, 225–239.
| Methane yields from Brahman cattle fed tropical grasses and legumes.Crossref | GoogleScholarGoogle Scholar |
Kennedy PM, Boniface AN, Liang ZJ, Muller D, Murray RM (1992) Intake and digestion in swamp buffaloes and cattle. 2. The comparative response to urea supplements in animals fed tropical grasses. The Journal of Agricultural Science 119, 243–254.
| Intake and digestion in swamp buffaloes and cattle. 2. The comparative response to urea supplements in animals fed tropical grasses.Crossref | GoogleScholarGoogle Scholar |
Laetsch WM (1974) The C4 syndrome: a structural analysis. Annual Review of Plant Physiology 25, 27–52.
| The C4 syndrome: a structural analysis.Crossref | GoogleScholarGoogle Scholar |
Licitra G, Hernandez TM, Van Soest PJ (1996) Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology 57, 347–358.
| Standardization of procedures for nitrogen fractionation of ruminant feeds.Crossref | GoogleScholarGoogle Scholar |
McLennan SR (1997) Developing profitable strategies for increasing growth rates of cattle grazing tropical pastures. Final Report Project DAQ.100. Meat Research Corporation, Sydney, NSW, Australia.
McLennan SR (2014) Optimizing growth paths of beef cattle in northern Australia for increased profitability. Final Report Project B.NBP.0391. Meat & Livestock Australia, Sydney.
Mitchell RB, Redfearn DD, Moser LE, Grant RJ, Moore KJ, Kirch BH (1997) Relationships between in situ protein degradability and grass developmental morphology. Journal of Dairy Science 80, 1143–1149.
| Relationships between in situ protein degradability and grass developmental morphology.Crossref | GoogleScholarGoogle Scholar |
Mupangwa JF, Ngongoni NT, Hamudikuwanda H (2003) Effects of stage of maturity and method of drying on in situ nitrogen degradability of fresh herbage of Cassia rotundifolia, Lablab purpureus and Macroptilium atropurpureum. Livestock Research for Rural Development 15,
National Academies of Sciences, Engineering and Medicine (2016) ‘Nutrient requirements of beef cattle.’ 8th revised edn. (The National Academies Press: Washington, DC, USA)
Negi SS, Singh B, Makkar HPS (1988a) Rumen degradability of nitrogen in typical cultivated grasses and leguminous fodders. Animal Feed Science and Technology 22, 79–89.
| Rumen degradability of nitrogen in typical cultivated grasses and leguminous fodders.Crossref | GoogleScholarGoogle Scholar |
Negi SS, Singh B, Makkar HPS (1988b) An approach to the determination of rumen degradability of nitrogen in low-grade roughages and partition of nitrogen therein. The Journal of Agricultural Science 111, 487–494.
| An approach to the determination of rumen degradability of nitrogen in low-grade roughages and partition of nitrogen therein.Crossref | GoogleScholarGoogle Scholar |
Nogueira Filho JCM, Fondevila M, Barrios Urdaneta A, Gonzalez Ronquillo M (2000) In vitro microbial fermentation of tropical grasses at an advanced maturity stage. Animal Feed Science and Technology 83, 145–157.
| In vitro microbial fermentation of tropical grasses at an advanced maturity stage.Crossref | GoogleScholarGoogle Scholar |
Norton BW (1982) Differences between species in forage quality. In ‘Nutritional limits to animal production from pastures’. (Ed. JB Hacker) pp. 89–110. (Commonwealth Agricultural Bureaux: Farnham Royal, UK)
Ogden RK, Coblentz WK, Coffey KP, Turner JE, Scarbrough DA, Jennings JA, Richardson MD (2006) Ruminal in situ disappearance kinetics of nitrogen and neutral detergent insoluble nitrogen from common crabgrass forages sampled on seven dates in northern Arkansas. Journal of Animal Science 84, 669–677.
| Ruminal in situ disappearance kinetics of nitrogen and neutral detergent insoluble nitrogen from common crabgrass forages sampled on seven dates in northern Arkansas.Crossref | GoogleScholarGoogle Scholar |
Peripoli V, Barcellos JOJ, Prates ER, McManus C, da Silva LP, Stella LA, Costa Junior JBG, Lopes RB (2016) Nutritional value of baled rice straw for ruminant feed. Revista Brasileira de Zootecnia 45, 392–399.
| Nutritional value of baled rice straw for ruminant feed.Crossref | GoogleScholarGoogle Scholar |
Pichard G, van Soest PJ (1977) Protein solubility of ruminant feeds. In ‘Proceedings of the nutrition conference’. pp. 91–98. (Department of Animal Science, Cornell University: Ithaca, NY, USA)
Prakash JR, Kundu SS, Das MM (2003) Carbohydrate and nitrogen fractionation of certain feeds and their utilization in growing buffaloes fed on total mixed ration. Indian Journal of Animal Science 73, 432–436.
Redfearn DD, Moser LE, Waller SS, Klopfenstein TJ (1995) Ruminal degradation of switchgrass, big bluestem, and smooth bromegrass leaf proteins. Journal of Animal Science 73, 598–605.
| Ruminal degradation of switchgrass, big bluestem, and smooth bromegrass leaf proteins.Crossref | GoogleScholarGoogle Scholar |
Salazar-Cubillas KC, Dickhoefer U (2021) Evaluating the protein value of fresh tropical forage grasses and forage legumes using in vitro and chemical fractionation methods. Animals 11, 2853
| Evaluating the protein value of fresh tropical forage grasses and forage legumes using in vitro and chemical fractionation methods.Crossref | GoogleScholarGoogle Scholar |
Scarbrough DA, Coblentz WK, Coffey KP, Turner JE, Davis GV, Kellogg DW, Hellwig DH (2002) Effects of summer management and fall harvest date on ruminal in situ degradation of crude protein in stockpiled bermudagrass. Animal Feed Science and Technology 96, 119–133.
| Effects of summer management and fall harvest date on ruminal in situ degradation of crude protein in stockpiled bermudagrass.Crossref | GoogleScholarGoogle Scholar |
Singh KK, Das MM, Samanta AK, Kundu SS, Sharma SD (2002) Evaluation of certain feed resources for carbohydrate and protein fractions and in situ digestion characteristics. Indian Journal of Animal Science 72, 794–797.
Singh S, Kushwaha BP, Nag SK, Mishra AK, Singh A, Anele UY (2012) In vitro ruminal fermentation, protein and carbohydrate fractionation, methane production and prediction of twelve commonly used Indian green forages. Animal Feed Science and Technology 178, 2–11.
| In vitro ruminal fermentation, protein and carbohydrate fractionation, methane production and prediction of twelve commonly used Indian green forages.Crossref | GoogleScholarGoogle Scholar |
Sniffen CJ, O’Connor JD, Van Soest PJ, Fox DG, Russell JB (1992) A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. Journal of Animal Science 70, 3562–3577.
| A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability.Crossref | GoogleScholarGoogle Scholar |
Stuth JW, Freer M, Dove H, Lyons RK (1999) Nutritional management of free-ranging livestock. In ‘Nutritional ecology of herbivores’. (Eds H-JG Jung, GC Fahey) pp. 696–751. (American Society of Animal Science: Savoy, IL, USA)
Van Soest PJ (1982) ‘Nutritional ecology of the ruminant.’ (O & B Books: Corvallis, OR, USA)
Wales WJ, Dellow DW Wales WJ, Dellow DW (1999) Degradabilities of dry matter and crude protein from perennial herbage and supplements used in dairy production systems in Victoria. Australian Journal of Experimental Agriculture 39, 645–656.
| Degradabilities of dry matter and crude protein from perennial herbage and supplements used in dairy production systems in Victoria.Crossref | GoogleScholarGoogle Scholar |
Wang Y, Zhang YG, Liu X, Kopparapu NK, Xin H, Liu J, Guo J (2015) Measurement of the intestinal digestibility of rumen undegraded protein using different methods and correlation analysis. Asian-Australasian Journal of Animal Sciences 28, 1454–1464.
| Measurement of the intestinal digestibility of rumen undegraded protein using different methods and correlation analysis.Crossref | GoogleScholarGoogle Scholar |
Waters CJ, Kitcherside MA, Webster AJF (1992) Problems associated with estimating the digestibility of undegraded dietary nitrogen from acid-detergent insoluble nitrogen. Animal Feed Science and Technology 39, 279–291.
| Problems associated with estimating the digestibility of undegraded dietary nitrogen from acid-detergent insoluble nitrogen.Crossref | GoogleScholarGoogle Scholar |
Webster AJF (1987) Metabolizable protein – the UK approach. In ‘Feed evaluation and protein requirement systems for ruminants’. (Eds R Jarrige, G Alderman) pp. 47–53. (Office of the Official Publications of the European Communities: Luxemburg)
Webster AJF, Simmons IP, Kitcherside MA (1982) Forage protein in the performance and health of the dairy cow. In ‘Forage protein in ruminant animal production’. (Eds DJ Thomson, DE Beever, RG Gunn) pp. 89–95. Occasional Publication No 6. (British Society of Animal Production: Glenrothes, UK)
Webster AJF, Dewhurst RJ, Waters CJ (1988) Alternative approaches to the characterization of feedstuffs for ruminants. In ‘Recent advances in animal nutrition’. (Eds W Haresign, DJA Cole) pp. 167–191. (Butterworths: London, UK)
Wilson JR (1982) Environmental and nutritional factors affecting herbage quality. In ‘Nutritional limits to animal production from pastures’. (Ed. JB Hacker) pp. 111–131. (Commonwealth Agricultural Bureaux: Farnham Royal, UK)
Wilson JR (1994) Cell wall characteristics in relation to forage digestion by ruminants. The Journal of Agricultural Science 122, 173–182.
| Cell wall characteristics in relation to forage digestion by ruminants.Crossref | GoogleScholarGoogle Scholar |
Wilson PN, Strachan PJ (1981) The contribution of undegraded protein to the protein requirements of dairy cows. In ‘Recent advances in animal nutrition – 1980’. (Ed. W Haresign) (Butterworths: London, UK)
Winks L (1984) ‘Cattle growth in the dry tropics of Australia. Review number 45.’ (Australian Meat Research Committee: Sydney, NSW, Australia)
Winks L, Laing AR, O’Rourke PK, Wright GS (1979) Factors affecting response to urea-molasses supplements by yearling cattle in tropical Queensland. Australian Journal of Experimental Agriculture and Animal Husbandry 19, 522–529.
| Factors affecting response to urea-molasses supplements by yearling cattle in tropical Queensland.Crossref | GoogleScholarGoogle Scholar |
Zhao GY, Cao JE (2004) Relationship between the in vitro-estimated utilizable crude protein and the Cornell net carbohydrate and protein system crude protein fractions in feeds for ruminants. Journal of Animal Physiology and Animal Nutrition 88, 301–310.
| Relationship between the in vitro-estimated utilizable crude protein and the Cornell net carbohydrate and protein system crude protein fractions in feeds for ruminants.Crossref | GoogleScholarGoogle Scholar |