

Can measurements of foraging behaviour predict variation in weight gains of free-ranging cattle?
David J. Augustine A * , Edward J. Raynor A , Sean P. Kearney A and Justin D. Derner B
A * , Edward J. Raynor A , Sean P. Kearney A and Justin D. Derner B
A United States Department of Agriculture, Agricultural Research Service, Rangeland Resources and Systems Research Unit, Fort Collins, CO 80526, USA.
B United States Department of Agriculture, Agricultural Research Service, Rangeland Resources and Systems Research Unit, Cheyenne, WY 82009, USA.
Animal Production Science - https://doi.org/10.1071/AN21560
Submitted: 31 October 2021 Accepted: 3 March 2022 Published online: 20 May 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Technologies are now available to continuously monitor livestock foraging behaviours, but it remains unclear whether such measurements can meaningfully inform livestock grazing management decisions. Empirical studies in extensive rangelands are needed to quantify relationships between short-term foraging behaviours (e.g. minutes to days) and longer-term measures of animal performance.
Aims: The objective of this study was to examine whether four different ways of measuring daily foraging behaviour (grazing-bout duration, grazing time per day, velocity while grazing, and turn angle while grazing) were related to weight gain by free-ranging yearling steers grazing semiarid rangeland.
Methods: Yearling steers were fitted with neck collars supporting a solar-powered device that measured GPS locations at 5 min intervals and used an accelerometer to predict grazing activity at 4 s intervals. These devices were used to monitor steers in four different paddocks that varied in forage biomass, and across two grazing seasons encompassing a wide range of forage conditions. Steer weight gain (kg/steer.day) was measured in each paddock during each of three ∼60 day time intervals, and daily foraging behaviours were measured during 15–21 days in each interval.
Results: A model based on only two daily measurements of foraging behaviour, mean grazing bout duration (calculated at a 5 min resolution) and mean velocity while grazing explained 62% of the variation in animal weight gain.
Conclusions: Daily measurements of foraging behaviour vary substantially in response to varying foraging conditions in space and time, and can effectively serve as indicators of variation in cattle weight gain.
Implications: On-animal sensors that monitor foraging behaviour have the potential to transmit indicators to livestock managers in real time (e.g. daily) to help inform decisions such as when to move animals among paddocks, or when to sell or transition animals from rangeland to confined feeding operations.
Keywords: accelerometer, average daily gain, forage limitation, grazing bout duration, grazing velocity, semiarid rangeland, shortgrass steppe, weight gain.
Introduction
Free-ranging livestock in extensive rangelands often experience substantial spatiotemporal variability in the biomass and quality of available forage. In response, animals adjust their foraging behaviours at multiple temporal scales, ranging from bite formation and rate, to the length of foraging bouts, and the amount of time spent foraging within a day, week or season (e.g. Brosh et al. 2006; Laca 2008; Braghieri et al. 2011). Understanding the magnitude of change in daily foraging behaviours in response to forage quality and daily intake may enable forage behavior measurements to be used as indicators of animal performance (Searle et al. 2007; Carvalho et al. 2015). Over the past two decades, the ability to measure subdaily to daily foraging behaviours has advanced rapidly through development of technologies to remotely monitor animal movement pathways via GPS collars, and simultaneously monitor other types of activity (e.g. heart rate and movements associated with grazing activity) via sensors (Brosh et al. 2006; Ungar and Rutter 2006; Ungar et al. 2011; Augustine and Derner 2013; Barwick et al. 2020; Raynor et al. 2021). One key advance has been the ability to monitor and classify diurnal animal location and foraging activity (e.g. Raynor et al. 2021; Suparwito et al. 2021). If daily foraging behaviours can provide reliable indicators of animal performance across paddocks and management systems, they could be used by managers for timing herd rotations among paddocks, assessing optimal stocking densities on the basis of current forage conditions, and determining supplement needs or ideal timing to move livestock among types of extensive rangelands or to confined finishing operations (Peel 2003; Da Trindade et al. 2012; Imaz et al. 2020).
Recent studies of diurnal foraging activity have begun to quantify variation in time spent grazing and distance travelled per day for different cattle breeds using the same forage conditions, as well as variation within a breed between wet and dry seasons (Aharoni et al. 2013; Spiegal et al. 2019). In some cases, shifts in behaviour or differences among breeds may be expressed only under conditions when forage biomass and/or quality is low (Braghieri et al. 2011; Aharoni et al. 2013). However, the direction and magnitude of these changes may be difficult to predict. During periods of higher forage quality and/or biomass, animals may be able to forage more selectively, and that might change the length of grazing bouts, short-term intake rate (e.g. g per minute), and grazing time per day (Carvalho et al. 2015). However, increased diet quality could also be achieved with slower, more selective searching and prehension bites (i.e. area-restricted search; Kareiva and Odell 1987), with little change in short-term intake, but a reduction in time spent ruminating and increased grazing time per day. For example, cattle grazing desert grasslands substantially increased grazing time per day when grazing lush, green herbage, as compared with dry-season forage (Brosh et al. 2006). Empirical studies in extensive rangelands are needed to unravel the complex interactions among foraging behaviours expressed over minutes to days, and how they relate to measures of animal performance. Improved understanding of which behavioural measurements are most sensitive to changing forage conditions could additionally assist development of on-animal sensors that could detect impending changes in animal performance and transmit these indicators to livestock managers in real time.
We examined the use of a GPS tracking collar combined with an accelerometer to quantify grazing behaviours that may be associated with measures of animal weight gain. Our first objective was to examine the degree to which grazing time per day varies in relation to the temporal resolution of the calculation of grazing activity, as well as in response to spatiotemporal variation in forage conditions both early in the growing season when vegetation was green and actively growing, and later when vegetation was senescing. Our second objective was to examine how additional measurements of daily foraging behaviour, such as movement rate, complexity of the foraging pathways, and length of foraging bouts could be used along with grazing time per day to predict variation in cattle weight gain. We hypothesised that behavioural measurements of foraging selectivity could potentially serve as better indicators of forage limitation and/or animal performance than would time spent grazing per day.
Materials and methods
Study area
All research was conducted at the United States Department of Agriculture (USDA)–Agricultural Research Service’s Central Plains Experimental Range (CPER), a Long-Term Agroecosystem Research network site that encompasses approximately 6400 ha of shortgrass steppe in north-eastern Colorado (40°50′N, 104°43′W). Mean annual precipitation was 340 mm and mean growing-season precipitation (April–August) was 241 mm. The CPER was subdivided into paddocks that vary from 65 to 390 ha in size, and they have been grazed by cattle since the station was established in 1939. Topography consisted of gently undulating plains at a mean elevation of 1640 m. Soils consisted of deep, well drained, fine sandy loams to loamy sands on alluvial flats and upland plains (Kelly et al. 2008). All research followed the Institutional Animal Care and Use Committee protocol (#CPER-4) approved March 2019 by the USDA–Agricultural Research Service in Fort Collins, CO, USA.
Cattle foraging behaviour was studied during 2019 and 2020 in four 130-ha paddocks that were selected to represent a gradient from high to low standing forage biomass and total herbaceous productivity during the growing season (Table 1, Fig. 1). Paddock 1 contained soils associated with the Sandy Plains ecological site (USDA 2007a) and was dominated by C3 perennial grasses (Pascopyrum smithii and Hesperostipa comata) combined with an understorey of C4 shortgrasses (Bouteloua gracilis and B. dactyloides) that had been grazed at a moderate growing-season stocking rate for the past 30 years. Prior measurements during near-average production years of 2016–2018 (Augustine et al. 2020) found that this soil type and plant community yielded mean herbaceous vegetation production of 1050 kg DM/ha.

|

|
The remaining three paddocks had soils associated with the Loamy Plains ecological site (USDA 2007b). Paddocks 2 and 3 were consistently grazed at moderate and heavy growing-season stocking rates respectively, since 1939, as part of a long-term stocking-rate experiment (Porensky et al. 2017). Paddock 2 was dominated by C4 shortgrasses coexisting with a subdominant layer of C3 perennial grasses. During 2016–2018, this soil and plant community yielded net herbaceous production of 770 kg DM/ha (Augustine et al. 2020). In Paddock 3, the history of heavy grazing (∼50% greater than in Paddock 2) has maintained a C4-dominated plant community where C3 perennial grasses are rare (Porensky et al. 2017). During 2003–2013, this history of heavy grazing reduced herbaceous production, on average, by 25% compared with moderately grazed paddocks (Irisarri et al. 2016). Paddock 4 was grazed by cattle at a moderate stocking rate for the past 30 years, and was dominated by C4 shortgrasses with subdominant C3 perennial grasses (as in Paddock 2). In addition, Paddock 4 had been grazed by black-tailed prairie dogs (Cynomys ludovicianus), which are a colonial, burrowing rodent that consumes a similar diet as does cattle. The proportion of Paddock 4 occupied by prairie dog colonies has fluctuated since the mid-1990s in response to disease outbreaks (Augustine and Derner 2021). During 2018, prairie dog colonies occupied 48% of Paddock 4, which has been shown to substantially reduce standing forage biomass when compared with areas not occupied by colonies during average-precipitation years (Augustine and Springer 2013). It was therefore expected that the daily standing forage biomass over the course of the growing season in the four study paddocks would follow the order of 1 > 2 > 3 = 4.
Forage conditions
Kearney et al. (2022) recently calibrated a model that uses the Harmonized Landsat–Sentinel dataset (Claverie et al. 2018) to generate smoothed daily estimates of standing herbaceous biomass at a 30 × 30 m pixel resolution for this site. The model was calibrated specifically to the vegetation, soils, and weather patterns in our study area by using 8 years of previous ground-based vegetation biomass measurements (Kearney et al. 2022). This model was used to estimate daily standing herbaceous biomass for each of the four study paddocks during the 2019 and 2020 growing season. The model estimates standing biomass (including live and dead) of grasses and forbs only and does not include cactus or subshrubs, which are generally not eaten by cattle.
Forage quality was measured by conducting direct observations of grazing steers in Paddocks 1 and 2 during 10 min grazing bouts, as part of a continuous bite-monitoring study described by Raynor et al. (2021). At the conclusion of each day of direct observations conducted in a paddock, technicians collected hand-plucked samples of each of the types of bites that they observed, where bite types were classified by plant functional group and bite depth, following Agreil and Meuret (2004) and Bonnet et al. (2015). After the hand-plucked samples were oven dried, a composite forage-intake sample was generated by weighting a pooled sample of bite types by the number of bites collectively observed for that type during a given observation day. Six of these weighted daily forage samples were obtained for each paddock and study period combination. Samples were analysed for crude protein and total digestible nutrient content by near-infrared spectroscopy (NIRS) at Ward Laboratories (Kearney, NE, USA), using calibrations developed by the NIRS Forage and Feed Testing Consortium (www.nirsconsortium.com). Due to the labour-intensive nature of forage-sample collection method, forage-quality samples were not collected in Paddocks 3 and 4.
Cattle stocking rates and measurements of liveweight and foraging behaviour
Study paddocks were stocked with yearling steers that entered in mid-May and grazed until early October each year. Steers were mixed-European breeds provided by local ranchers, and were weighed individually at the beginning and end of the grazing season. Steers were weighed without overnight restriction of water or food; however, weights were corrected for estimated overnight shrinkage following Derner et al. (2016). Steers were additionally weighed 53 days before the end of the 2019 grazing season, 59 days after the start of the 2020 grazing season, and 60 days prior to the end of the 2020 season, to obtain seasonal estimates of average daily weight gain (ADG) that corresponded to the time periods when cattle foraging behaviour was measured (see below and Table 1). Each year, stocking rates were set prior to the grazing season by a group of stakeholders that included local ranchers, land managers, and representatives of non-governmental organisations (Wilmer et al. 2018). Generally, yearling steers at our study site gain ∼125–130 kg liveweight during the May–October grazing season; hence, it was assumed that one yearling steer = 1.03 animal equivalents (AE), as recommended by McLennan et al. (2020) for livestock of moderate productivity. On the basis of stakeholders’ recommendation, the moderate stocking rate on Paddock 1 (Sandy Plains) was 0.21 AE/ha, and on Paddocks 2 and 4 (Loamy Plains), it was 0.19 AE/ha over 136 and 137 day periods in 2019 and 2020 respectively. The heavy stocking rate for Paddock 3 (Loamy Plains) was 0.27 AE/ha, or ∼50% above moderate.
In 2019, MOOnitor GPS collars (www.moonitorcows.com) were deployed on four randomly selected steers in each study paddock from 5 to 30 September, which corresponded to a period of low and senescent forage biomass, when it was anticipated that cattle weight gain may vary substantially among paddocks. Collars were set to collect GPS fixes every 5 min, and additionally recorded the steer’s predicted activity state (resting, grazing, or walking) at 4 s intervals derived from a proprietary algorithm applied to measurements recorded by a three-axis accelerometer. In 2020, MOOnitor collars were deployed on four randomly selected steers in each paddock at the beginning of the grazing season, when vegetation was green and increasing in biomass (albeit at rates lower than during average precipitation years), and again at the end of the grazing season, when vegetation was senescent and at unusually low biomass in Paddocks 3 and 4 (<400 kg DM/ha; Fig. 1). In 2019, some collars failed to collect data (including all collars in Paddock 2) because problems with the connection of the neck straps to the solar panels and GPS receiver caused them to shift sideways rather than being centred on the steer’s neck (Supplementary Fig. S1), which prevented sufficient daily charging. This strap connection was modified in 2020, but some collars still occasionally shifted to where they were not centred on the neck, resulting in incomplete charging on some days, depending on the steer’s orientation to the sun and weather conditions. Such days were not included in analyses.
To assess the validity of predictions generated by the activity sensors, direct visual observations of steer behaviours were conducted during each of the three deployment intervals (late summer of 2019, early and late summer of 2020). Following Augustine and Derner (2013), visual observations occurred during daylight hours, typically beginning during morning hours and continuing until early afternoon, with the goal of encompassing bouts of both grazing and resting within a given day. Cattle activity was classified into categories consisting of grazing (including grazing while walking), travelling (walking without grazing), standing, bedding, grooming, drinking, or consuming mineral supplements. Activity timing and duration were recorded every 30 s. When an animal changed activity, observers noted the precise transition time. If the new activity persisted for >15 s, then the transition was recorded on the field data sheet. If the animal resumed prior activity in <15 s, the interlude was ignored.
Data analyses
To evaluate concordance between activity sensor predictions and direct observations of grazing steers, both data types were summarised at 1 min intervals and were linked on the basis of time stamps. For direct observations, if the minute interval included only one 30 s grazing period, it was still classified as grazing. For activity sensor output, each minute was classified as ‘grazing’ when >50% of the 4 s intervals in that minute were classified as grazing.
To examine daily grazing behaviour, the activity data recorded at 4 s intervals were used to calculate the time spent grazing each day in hours (GHrs4sec, in hours) and the mean grazing bout duration (GBD4sec, in seconds) each day, where a grazing bout was defined as a continuous string of 4 s intervals of grazing. For example, if the sensor measured that an animal was grazing for 20 consecutive 4 s intervals, then was not grazing for one or more subsequent 4 s intervals, the bout duration was 80 s. In addition, the 4 s records of grazing activity were used to classify each 5 min interval between the GPS fixes as either a grazing interval (>50% of the 5 min interval spent grazing) or non-grazing interval (<50% spent grazing). A 5 min resolution version of grazing time per day (GHrs5min, in hours) was calculated as the sum of 5 min grazing intervals, and a 5 min resolution of the mean grazing bout duration (GBD5min, in minutes) was calculated where a bout was defined as a continuous string of 5 min intervals of grazing, separated from other grazing bouts by at least one 5 min interval of non-grazing activity. This method of calculating the grazing-bout length corresponds closely to that used by Orr et al. (2001), who defined the end of a grazing bout as when an animal stopped grazing for 6 min or more, and demonstrated that grazing-bout duration of sheep varied inversely with forage quality. For all analyses, data were used only from days when the MOOnitor device successfully collected GPS and activity data for ≥99% of the 4 s intervals.
The GPS fix data were used to calculate the mean velocity for each 5 min interval during which the majority of the 4 s intervals were classified as grazing (VG, in m/s). In addition, for each series of three consecutive GPS fixes where the animal was classified as grazing for the entire 10 min time period, the angle between the vector connecting the first and second fix, and the vector connecting the second and third fix was calculated. This value was then subtracted from 180. Hereafter, this is referred to as the turn angle while grazing (TAG, deviation from a straight line in degrees). An animal grazing in a straight line would have a turn angle of 0°, while an animal whose first vector is perpendicular to its second vector would have a turn angle of 90°.
The weight gain of each collared steer was calculated on the basis of liveweight measured at the start and end of each study period (Table 1). Each weight-gain interval was approximately 60 days, which encompassed the collar deployment intervals (Table 1). Models of cattle weight gain were fit as a function of all possible combinations of the foraging behaviour measurements, and an information-theoretic approach was used for model selection (Anderson 2008), where Akaike’s information criterion with a correction for a small sample size (AICc) was used. All statistical analyses were performed using JMP v13.2.1 from the SAS Institute, Inc. (Cary, NC, USA).
Results
Forage conditions
In 2019, peak biomass varied among paddocks from ∼700 to 1100 kg DM/ha in the middle of the growing season, and then senesced rapidly during the second half due to dry conditions. Paddocks 1 and 2 contained substantially more standing forage biomass than did Paddocks 3 and 4 during late summer, when measurements of cattle weight gains and foraging behaviour were conducted (Fig. 1a). Forage quality in Paddocks 1 and 2 (not measured in Paddocks 3 and 4) was especially low in this study period, compared with early summer of 2020 (Table 2). In 2020, biomass increased marginally during the first 20 days and remained green and highly digestible throughout the early summer study period, reaching a peak of 500–600 kg DM/ha, with Paddocks 1 and 2 supporting ∼100 kg DM/ha more biomass than did Paddocks 3 and 4 (Fig. 1b). Due to lower rainfall and forage growth in 2020, study paddocks had especially low forage biomass during the late-summer study period (Fig. 1b), with levels in Paddocks 3 and 4 approaching conditions at which supplemental feed is recommended to maintain cattle weight gain in our study system (Bement 1969). Forage quality in Paddocks 1 and 2 at this time was much reduced compared with that in early summer (Table 2).

|
Collar operation and activity sensor validation
The collars successfully collected GPS fix and activity sensor data for nine steers in late summer of 2019, 14 steers in early summer of 2020, and 13 steers in late summer of 2020 (Table 1).
Concurrent visual observations and collar-based measurements of grazing activity were obtained for three, six, and five steers in late summer 2019, early summer 2020, and late summer 2020 respectively, to determine the reliability of the collar-based measurements. Two of these direct observation periods were only 40 min long. The remaining 12 periods ranged from 203 to 345 min (mean = 297 min), for a total of 3648 min of concurrent measurements. Of these minutes, 2173 min were correctly classified as a grazing interval, 1160 min were correctly classified as a non-grazing interval, 203 min were falsely classified as grazing, and 112 min were falsely classified as non-grazing intervals. This yielded a false positive rate of 8.5% (95% CI = 4.3%), false negative rate of 8.8% (95% CI = 7.2%), and overall error rate of 8.6% (95% CI = 3.6%), which is slightly lower than error rates reported for other grazing activity sensors (Ungar et al. 2011; Augustine and Derner 2013).
Daily foraging behaviour measurements versus ADG
We found that GHrs4sec and GHrs5min were closely correlated (r2 = 0.91, P < 0.001; Fig. S2), with similar means of 10.7 h/day and 10.6 h/day respectively. However, estimates from 5 min intervals varied more widely, from 6.1 to 15.9 h/day, while estimates from the 4 s intervals varied only from 7.2 to 14.9 h/day. The regression largely followed the 1:1 line for values from 8 to 13 h, but the 5 min estimates were greater than expected (above 1:1 line) for days with grazing times greater than 13 h, and lower than expected (below 1:1 line) for days with grazing times less than 8 h. This result indicated that GHrs4sec was more accurate than GHrs5min; hence, we considered only GHrs4sec in all models of weight gain. GHrs4sec varied significantly among the three study periods, with increased daily grazing time in early summer of 2020 with green, highly digestible vegetation (11.7 ± 0.18 h; mean ± s.e.), compared with late summer of 2019 (10.0 ± 0.21 h) and 2020 (10.4 ± 0.16 h) with senescent vegetation.
We also found that GBD4sec and GBD5min were correlated, but much less strongly than was grazing time (r2 = 0.31, P < 0.001). The lower correlation reflects that they are measuring two different but related behaviours, where the 4 s estimate is measuring how frequently the animal stops taking bites, typically to search for its next set of bites. The 5 min estimate is primarily a measure of how frequently the animal stops a grazing bout entirely, often arising from a combination of the animal raising the head to survey its environment, searching for or walking to a new area in which to graze, or stopping to rest and ruminate. Because of their low correlation, we considered models with both estimates, but only included one or the other in any given model.
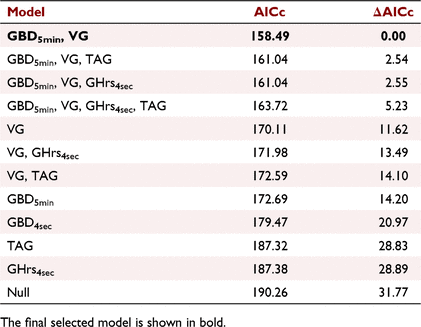
Models of ADG based on the two different procedures to measure duration showed that GBD5min performed better than GBD4sec (Table 3). In addition, single-predictor models based on TAG, GHrs4sec and VG all performed better than did the null model (reducing AICc by 2.88, 2.94, and 20.16 respectively, compared with the null; Table 3). When we fit all possible models on the basis of the main effects, including all possible combinations of two or more of these four behavioural measurements (GBD5min, TAG, GHrs4sec, and VG), the model based only on GBD5min and VG provided the best fit to the data (Table 2). These two measurements of daily grazing behaviour were able to predict 62% of the variation in ADG (Fig. 2), where
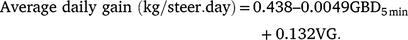
|

|
The standard error for the estimate of the intercept was 0.288, and for the estimates of the coefficients for GBD5min and VG were 0.0012 and 0.029 respectively.
Discussion
There are several technologies available that may have application for summarising daily foraging behaviour measurements and transmitting them to smart devices through satellite transmissions (e.g. www.moonitor.com), cell phones (in areas with adequate coverage), or through non-cellular, long-range and low-power wireless, wide-area networks (employed by companies developing virtual fence technologies in Australia, Europe, and North America). These technologies create the potential to monitor changes in forage conditions within paddocks and/or animal performance via changes in foraging behaviour. Here, we examined how four measurements of daily grazing behaviour varied across paddocks with varying forage biomass and sward composition, and across two grazing seasons within a semiarid, temperate rangeland.
First, for daily time spent grazing, we found the temporal resolution at which grazing time is calculated to be an important factor. Using an activity sensor that classified each 4 s interval as grazing or non-grazing, our estimates of daily grazing time rarely exceeded 14 h or were less than 7 h. In contrast, when each 5 min interval was classified as grazing or non-grazing, the frequency of daily grazing times above or below these extremes increased. This result suggests that using accelerometers to classify behaviour at subminute time scales may provide more robust estimates of grazing time than do measurements conducted at 5 min or 10 min intervals (e.g. as estimated by 2-axis mechanical sensors; Augustine and Derner 2013). At the 4 s resolution, daily grazing time varied significantly among the study periods, and explained more variation in animal weight gain than did a null model (Table 2). When cattle are grazing a sward dominated by green leaves, initial depletion of the sward can lead to an increase in daily grazing time as the animals compensate for declining bite size (Benvenutti et al. 2016). However, as the quality of available forage declines due to plant maturation and senescence, grazing time per day can decline substantially as intake is restricted by digestion rate. In this study, we found that grazing time per day declined by ∼1.5 h when steers were grazing senescent, low-quality vegetation late in the growing season. Brosh et al. (2006) reported an even larger decline in daily grazing time of ∼5–6 h between growing and dormant seasons in arid Israeli rangelands. Daily grazing time did not improve predictions of animal weight gain when added to a model that already included grazing bout duration and velocity while grazing. Thus, the importance of shifts in daily grazing time as an indicator of animal performance may depend on a variety of factors such as the magnitude of the change in forage conditions, diversity and composition of the rangeland, and breed or genetic strain of the animals (Aharoni et al. 2013; Spiegal et al. 2019).
Second, grazing-bout duration provided a valuable indicator of differences in animal weight gain. We calculated bout duration at two different temporal resolutions because we expected that they would represent two different levels of foraging selectivity. If the start and end of a bout is defined as a switch between grazing and non-grazing activity for just a 4 s interval, then ‘bouts’ can become longer when the animal rarely pauses between bites or feeding stations to search for the next bite or station. In contrast, if a ‘bout’ ends when the animal stops grazing for 5 min or more, then bouts can become longer as the animal engages in a few long, continuous periods of intense grazing each day (often an hour or more), rather than several bouts shorter than an hour, which are punctuated by longer searches for the next feeding patch (e.g. collection of feeding stations). We found that GBD5min was a better predictor of animal weight gain than was any other foraging-behaviour measurement. One potential explanation is that when animals are given access to swards with both larger and more digestible bites, they can increase diet quality by being more selective in which feeding patches they choose, with an associated increase in time periods of 5 min or more spent searching for the next feeding patch. In addition, animals with access to larger and more digestible bites can increase short-term intake rate and thereby reach satiety and gut fill more rapidly, thereby reducing grazing bout length (Orr et al. 2001). Although our findings are specific to the shortgrass rangeland grazed by these study steers, they suggest grazing bout duration be examined more widely as a key indicator of changes in animal performance.
Third, we found that animal velocity while grazing was an important indicator of variation in animal weight gain. When vegetation was green and growing early in 2020, grazing velocities were consistently ≥7.2 m/min, but during late-summer conditions with senescent, low-quality forage, velocities were reduced to <6.5 m/min (Fig. 2b). Steers may have increased their velocity while grazing high-quality vegetation, so as to increase encounter rates with high-quality bites, and thereby increase diet quality without a reduction in short-term intake rate. To our knowledge, this is the first demonstration that grazing velocity, when used in combination with grazing bout duration, can be a second key indicator of animal performance. Given the increasing prevalence of GPS tracking studies of cattle at 5 min or shorter time steps, we also encourage evaluation of this indicator across a broad spectrum of rangelands and livestock types.
Fourth, we hypothesised that mean turning angle while grazing might increase with improved weight gain, because animals that are foraging more selectively (and hence acquiring higher-quality diets) may move in more convoluted pathways, which could be reflected in the turn angle. Under the conditions of our study, we found no strong correlation between turn angle and animal performance. However, our study examined only steers grazing at relatively low stock densities (3.8–5.8 ha/animal), and we suggest that turn angle while grazing could still potentially be sensitive to management approaches that employ higher stock densities, or paddocks with greater diversity of forage patch types. We also note that our turn angles were calculated at a 5 min time step, which may be too coarse to detect how foraging pathway tortuosity co-varies with diet quality. We encourage future studies to still consider turn angle while grazing as a potential indicator of animal performance, and, in particular, to continue to evaluate how grazing pathway complexity may be related to changes in stock density and vegetation diversity.
Overall, our findings support the idea that foraging behaviour measurements could serve as an indicator of changes in animal performance, but substantially more evaluation is needed to test this idea and develop weight gain predictions in varying types of rangelands, and across varying combinations of circumstances. We acknowledge that other technologies to monitor animal performance, such as walk-over weigh scales at water locations, remote sensing of forage biomass and quality, and faecal analyses of diet quality may be more effective under certain conditions. At the same time, the use of multiple indicators of cattle performance, including foraging behaviour measurements, could increase manager confidence and timeliness in their decision-making, as well as provide insights as to why animal performance is declining during the grazing season. Additionally, for applications based on foraging behaviour measurements to be useful and economical, the economic benefits of changes in management decisions would need to exceed costs of the technology. Such measurements may be most economical when used in combination with other emerging technologies, such as virtual fencing, which already require that the livestock wear a collar designed to communicate remotely with livestock managers. Behavioural measurements may also be most valuable under extensive rangeland conditions where livestock are not visited or observed frequently by managers. We encourage future studies to evaluate the measurements presented here across diverse types of rangelands worldwide, using research designs that encompass spatial variation in forage conditions associated with differing soils, plant communities (of varying heights and plant densities), and grazing management strategies, as well as temporal variation among seasons and years that differ in plant phenology and production levels.
Conclusions
We showed that daily measurements of cattle foraging behaviour can serve as indicators of changes in cattle weight gains associated with spatial and temporal variation in forage conditions. In the case of yearling steers grazing shortgrass rangeland in central North America, mean daily grazing-bout duration and velocity while grazing were the most valuable predictors of variation in individual weight gain, whereas grazing time per day and mean turn angle while grazing did not additionally improve predictions of weight gain. Given ongoing technological advances in on-animal sensors that can monitor animal foraging behaviour, these findings provide a pathway to provide cattle managers with daily updates of indicators of animal performance.
Supplementary material
Supplementary material is available online.
Data availability
Data supporting this publication are available online at data.nal.usda.gov/dataset/data-from-can-measurements-of-foraging-behaviour-predict-variation-in-weight-gains-of-free-ranging-cattle (https://doi.org/10.15482/USDA.ADC/1524788).
Conflicts of interest
MOOnitor provided ongoing support during this study but had no role in the interpretation of results or decision to publish. Mention of trade names or commercial products in this report is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Declaration of funding
Funding was provided by the USDA, National Institute for Agriculture (Award: 2018-68008-28090). This research was also a contribution from the Long-Term Agroecosystem Research network, which is funded by the USDA.
Acknowledgements
We thank N. Kaplan for assistance with collar data management, and M. Johnston, J. Thomas, J. Holtman, and T. Jorns for weighing and managing cattle. We thank A. Brosh for assistance in troubleshooting collar operation.
References
Agreil C, Meuret M (2004) An improved method for quantifying intake rate and ingestive behaviour of ruminants in diverse and variable habitats using direct observation. Small Ruminant Research 54, 99–113.| An improved method for quantifying intake rate and ingestive behaviour of ruminants in diverse and variable habitats using direct observation.Crossref | GoogleScholarGoogle Scholar |
Aharoni Y, Dolev A, Henkin Z, Yehuda Y, Ezra A, Ungar ED, Shabtay A, Brosh A (2013) Foraging behavior of two cattle breeds, a whole-year study: I. Heat production, activity and energy costs. Journal of Animal Science 91, 1381–1390.
| Foraging behavior of two cattle breeds, a whole-year study: I. Heat production, activity and energy costs.Crossref | GoogleScholarGoogle Scholar | 23348687PubMed |
Anderson DR (2008) ‘Model based inference in the life sciences: a primer on evidence.’ (Springer: New York, NY, USA)
Augustine DJ, Derner JD (2013) Assessing herbivore foraging behavior with GPS collars in a semiarid grassland. Sensors 13, 3711–3723.
| Assessing herbivore foraging behavior with GPS collars in a semiarid grassland.Crossref | GoogleScholarGoogle Scholar | 23503296PubMed |
Augustine DJ, Derner JD (2021) Long-term effects of black-tailed prairie dogs on livestock grazing distribution and mass gain. The Journal of Wildlife Management 85, 1332–1343.
| Long-term effects of black-tailed prairie dogs on livestock grazing distribution and mass gain.Crossref | GoogleScholarGoogle Scholar |
Augustine DJ, Derner JD, Fernández-Giménez ME, Porensky LM, Wilmer H, Briske DD (2020) Adaptive, multipaddock rotational grazing management: a ranch-scale assessment of effects on vegetation and livestock performance in semiarid rangeland. Rangeland Ecology & Management 73, 796–810.
| Adaptive, multipaddock rotational grazing management: a ranch-scale assessment of effects on vegetation and livestock performance in semiarid rangeland.Crossref | GoogleScholarGoogle Scholar |
Augustine DJ, Springer TL (2013) Competition and facilitation between a native and a domestic herbivore: trade-offs between forage quantity and quality. Ecological Applications 23, 850–863.
| Competition and facilitation between a native and a domestic herbivore: trade-offs between forage quantity and quality.Crossref | GoogleScholarGoogle Scholar | 23865235PubMed |
Barwick J, Lamb DW, Dobos R, Welch M, Schneider D, Trotter M (2020) Identifying sheep activity from tri-axial acceleration signals using a moving window classification model. Remote Sensing 12, 646
| Identifying sheep activity from tri-axial acceleration signals using a moving window classification model.Crossref | GoogleScholarGoogle Scholar |
Bement RE (1969) A stocking-rate guide for beef production on blue-grama range. Journal of Range Management 22, 83–86.
| A stocking-rate guide for beef production on blue-grama range.Crossref | GoogleScholarGoogle Scholar |
Benvenutti MA, Pavetti DR, Poppi DP, Gordon IJ, Cangiano CA (2016) Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle. Grass and Forage Science 71, 424–436.
| Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle.Crossref | GoogleScholarGoogle Scholar |
Bonnet OJF, Meuret M, Tischler MR, Cezimbra IM, Azambuja JCR, Carvalho PCF (2015) Continuous bite monitoring: a method to assess the foraging dynamics of herbivores in natural grazing conditions. Animal Production Science 55, 339–349.
| Continuous bite monitoring: a method to assess the foraging dynamics of herbivores in natural grazing conditions.Crossref | GoogleScholarGoogle Scholar |
Braghieri A, Pacelli C, Girolami A, Napolitano F (2011) Time budget, social and ingestive behaviours expressed by native beef cows in Mediterranean conditions. Livestock Science 141, 47–52.
| Time budget, social and ingestive behaviours expressed by native beef cows in Mediterranean conditions.Crossref | GoogleScholarGoogle Scholar |
Brosh A, Henkin Z, Ungar ED, Dolev A, Orlov A, Yehuda Y, Aharoni Y (2006) Energy cost of cows’ grazing activity: use of the heart rate method and the Global Positioning System for direct field estimation. Journal of Animal Science 84, 1951–1967.
| Energy cost of cows’ grazing activity: use of the heart rate method and the Global Positioning System for direct field estimation.Crossref | GoogleScholarGoogle Scholar | 16775080PubMed |
Carvalho PCF, Bremm C, Mezzalira JC, Fonseca L, da Trindade JK, Bonnet OJF, Tischler M, Genro TCM, Nabinger C, Laca EA (2015) Can animal performance be predicted from short-term grazing processes? Animal Production Science 55, 319–327.
| Can animal performance be predicted from short-term grazing processes?Crossref | GoogleScholarGoogle Scholar |
Claverie M, Ju J, Masek JG, Dungan JL, Vermote EF, Roger J-C, Skakun SV, Justice C (2018) The Harmonized Landsat and Sentinel-2 surface reflectance data set. Remote Sensing of Environment 219, 145–161.
| The Harmonized Landsat and Sentinel-2 surface reflectance data set.Crossref | GoogleScholarGoogle Scholar |
Da Trindade JK, Pinto CE, Neves FP, Mezzalira JC, Bremm C, Genro TCM, Tischler MR, Nabinger C, Gonda HL, Carvalho PCF (2012) Forage allowance as a target of grazing management: implications on grazing time and forage searching. Rangeland Ecology & Management 65, 382–393.
| Forage allowance as a target of grazing management: implications on grazing time and forage searching.Crossref | GoogleScholarGoogle Scholar |
Derner JD, Reeves JL, Mortenson MC, West M, Gonzalo Irisarri J, Durante M (2016) Estimating overnight weight loss of corralled yearling steers in semiarid rangeland. Rangelands 38, 101–104.
| Estimating overnight weight loss of corralled yearling steers in semiarid rangeland.Crossref | GoogleScholarGoogle Scholar |
Imaz JA, Garcia S, González LA (2020) Using automated in-paddock weighing to evaluate the impact of intervals between liveweight measures on growth rate calculations in grazing beef cattle. Computers and Electronics in Agriculture 178, 105729
| Using automated in-paddock weighing to evaluate the impact of intervals between liveweight measures on growth rate calculations in grazing beef cattle.Crossref | GoogleScholarGoogle Scholar |
Irisarri JGN, Derner JD, Porensky LM, Augustine DJ, Reeves JL, Mueller KE (2016)) Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecological Applications 26, 1370–1380.
Kareiva P, Odell G (1987) Swarms of predators exhibit ‘preytaxis’ if individual predators use area-restricted search. The American Naturalist 130, 233–270.
| Swarms of predators exhibit ‘preytaxis’ if individual predators use area-restricted search.Crossref | GoogleScholarGoogle Scholar |
Kearney SP, Porensky LM, Augustine DJ, Gaffney R, Derner JD (2022) Monitoring standing herbaceous biomass and thresholds in semiarid rangelands from harmonized Landsat 8 and Sentinel-2 imagery to support within-season adaptive management. Remote Sensing of Environment 271, 112907
| Monitoring standing herbaceous biomass and thresholds in semiarid rangelands from harmonized Landsat 8 and Sentinel-2 imagery to support within-season adaptive management.Crossref | GoogleScholarGoogle Scholar |
Kelly EF, Yonkers CM, Blecker SW, Olson CG (2008) Soil development and distribution in the shortgrass steppe ecosystem. In ‘Ecology of the shortgrass steppe: a long-term perspective’. (Eds W Lauenroth, IC Burke) pp. 30–54. (Oxford University Press: New York, NY, USA)
Laca EA (2008) Foraging in a heterogeneous environment: intake and diet selection. In ‘Resource ecology: spatial and temporal dynamics of foraging’. (Eds HHT Prins, F Van Langevelde) pp. 81–100. (CAB International: Wageninger, The Netherlands)
McLennan S, McLean I, Paton C (2020) Re-defining the animal unit equivalence (AE) for grazing ruminants and its application for determining forage intake, with particular relevance to the northern Australian grazing industries. Meat and Livestock Australia Limited, Sydney, NSW, Australia.
Orr RJ, Penning PD, Rutter SM, Champion RA, Harvey A, Rook AJ (2001) Intake rate during meals and meal duration for sheep in different hunger states, grazing grass or white clover swards. Applied Animal Behaviour Science 75, 33–45.
| Intake rate during meals and meal duration for sheep in different hunger states, grazing grass or white clover swards.Crossref | GoogleScholarGoogle Scholar |
Peel DS (2003) Beef cattle growing and backgrounding programs. Veterinary Clinics of North America: Food Animal Practice 19, 365–385.
| Beef cattle growing and backgrounding programs.Crossref | GoogleScholarGoogle Scholar |
Porensky LM, Derner JD, Augustine DJ, Milchunas DG (2017) Plant community composition after 75 yr of sustained grazing intensity treatments in shortgrass steppe. Rangeland Ecology & Management 70, 456–464.
| Plant community composition after 75 yr of sustained grazing intensity treatments in shortgrass steppe.Crossref | GoogleScholarGoogle Scholar |
Raynor EJ, Derner JD, Soder KJ, Augustine DJ (2021) Noseband sensor validation and behavioural indicators for assessing beef cattle grazing on extensive pastures. Applied Animal Behaviour Science 242, 105402
| Noseband sensor validation and behavioural indicators for assessing beef cattle grazing on extensive pastures.Crossref | GoogleScholarGoogle Scholar |
Searle KR, Hobbs NT, Gordon IJ (2007) It’s the ‘foodscape’, not the landscape: using foraging behaviour to make functional assessments of landscape condition. Israel Journal of Ecology and Evolution 53, 297–316.
| It’s the ‘foodscape’, not the landscape: using foraging behaviour to make functional assessments of landscape condition.Crossref | GoogleScholarGoogle Scholar |
Spiegal S, Estell RE, Cibils AF, James DK, Peinetti HR, Browning DM, Romig KB, Gonzalez AL, Lyons AJ, Bestelmeyer BT (2019) Seasonal divergence of landscape use by heritage and conventional cattle on desert rangeland. Rangeland Ecology & Management 72, 590–601.
| Seasonal divergence of landscape use by heritage and conventional cattle on desert rangeland.Crossref | GoogleScholarGoogle Scholar |
Suparwito H, Thomas DT, Wong KW, Xie H, Rai S (2021) The use of animal sensor data for predicting sheep metabolisable energy intake using machine learning. Information Processing in Agriculture 8, 494–504.
| The use of animal sensor data for predicting sheep metabolisable energy intake using machine learning.Crossref | GoogleScholarGoogle Scholar |
Ungar ED, Rutter SM (2006) Classifying cattle jaw movements: comparing IGER behaviour recorder and acoustic techniques. Applied Animal Behaviour Science 98, 11–27.
| Classifying cattle jaw movements: comparing IGER behaviour recorder and acoustic techniques.Crossref | GoogleScholarGoogle Scholar |
Ungar ED, Schoenbaum I, Henkin Z, Dolev A, Yehuda Y, Brosh A (2011) Inference of the activity timeline of cattle foraging on a Mediterranean woodland using GPS and pedometry. Sensors 11, 362–383.
| Inference of the activity timeline of cattle foraging on a Mediterranean woodland using GPS and pedometry.Crossref | GoogleScholarGoogle Scholar | 22346582PubMed |
USDA (2007a) Ecological site description for Loamy Plains (R067BY002CO). Available at https://esis.sc.egov.usda.gov/ESDReport/fsReport.aspx?approved=yes&id=R067BY002CO
USDA (2007b) Ecological site description for Sandy Plains (R067BY024CO). Available at https://esis.sc.egov.usda.gov/ESDReport/fsReport.aspx?approved=yes&id=R067BY024CO
Wilmer H, Derner JD, Fernández-Giménez ME, Briske DD, Augustine DJ, Porensky LM (2018)) Collaborative adaptive rangeland management fosters management-science partnerships. Rangeland Ecology & Management 71, 646–657.