

Rumen temperature is a reliable proxy of core body temperature in sheep (Ovis aries)
Kristine Vesterdorf A * , David T. Beatty B , Anne Barnes C and Shane K. Maloney A
A * , David T. Beatty B , Anne Barnes C and Shane K. Maloney A
A School of Human Sciences, The University of Western Australia, Crawley, WA, Australia.
B Meat and Livestock Australia, North Sydney, NSW, Australia.
C School of Veterinary and Life Sciences, Murdoch University, Perth, WA, Australia.
Animal Production Science 62(17) 1671-1682 https://doi.org/10.1071/AN21490
Submitted: 23 September 2021 Accepted: 3 June 2022 Published: 8 July 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: In ruminant animals, such as sheep, the maintenance of a stable core body temperature is achieved through the controlled process of thermoregulation. The live export of sheep from Australia can expose sheep to heat stress, especially when vessels near, or cross, the equator, that can impact on animal welfare by causing hyperthermia.
Aim: The use of thermologgers located in the rumen of sheep could provide a direct, real-time method to monitor the thermal status of sheep during live export without the need to handle animals during shipping. We determined the relationship between core temperature and rumen temperature during changes in various factors that impact on an animal’s heat balance.
Methods: Sixteen wethers were implanted with thermologgers in the rumen to measure rumen temperature and in the abdominal cavity to measure core temperature. The sheep were exposed to environmental conditions similar to those experienced by sheep on live export vessels. The environment was assessed using the wet-bulb temperature. In the first trial, eight Merinos were shorn while the remaining eight retained their fleece. In the second trial all sheep were shorn, and eight sheep were offered a chaff diet and eight a pelletised diet while the wet bulb temperature increased. The rumen and core temperatures were compared.
Key results: We show that when core temperature increased during exposure to heat load, so did rumen temperature and, despite relatively large changes in overall body temperature, the difference between these temperatures remained stable. The daily average relationship between rumen and core temperatures was 0.5 ± 0.1°C and that difference was not affected by water ingestion, length of fleece, diet composition, or exposure to hot and humid conditions similar to those reported on live export vessels.
Conclusion and implications: By subtraction of 0.5°C from the measured rumen temperature in live export sheep, the use of rumen thermologgers would provide a reliable measure of core temperature and hence an indication of the thermal status of the sheep throughout the export journey.
Keywords: animal welfare, biologging, heat stress, live sheep export, nutrition, rumen temperature, thermoregulation, wool.
Introduction
In mammals, the maintenance of a stable core body temperature (Tcore) is achieved through the homeostatically controlled process of thermoregulation. The Tcore varies with time of day in a circadian rhythm, and variations in the characteristics of that rhythm can provide an indication of pathology (Maloney et al. 2013, 2017, 2019). Variations in the circadian rhythm can occur during heat exposure, malnutrition, pregnancy, or infection (Hales 1973; Marai et al. 2007, 2008; Maloney et al. 2019).
Sheep that are being exported live from Australia can be exposed to thermal stress, especially when they cross the equator (Stockman et al. 2011). When the environmental conditions exceed the prescriptive zone for sheep (Mitchell et al. 2018), they cannot achieve heat balance and the Tcore increases (Marai et al. 2007; Beatty et al. 2008b; Stockman et al. 2011; Koltes et al. 2018). Welfare in an animal can be defined by alterations in body temperature, hormone levels and animal behaviour. If an animal has difficulties coping in a challenging environment, poor welfare will ensue (Silanikove 2000). Monitoring the Tcore of live export sheep could therefore provide a useful indicator of the physiological responses of the animals during transport, as an indication of their welfare.
The measurement of Tcore over long periods of time in live sheep is, however, not a straightforward task. The gold standard measurement of body core temperature is the temperature of the blood in the pulmonary artery, but the placement of a sensor into a patent artery involves complex surgery (Lim et al. 2008; Fuller et al. 2011; Vesterdorf et al. 2011). Other sites within the body are easier to access and can correlate reasonably well with the arterial blood temperature, such as within the abdominal cavity (Beatty et al. 2008a, 2008b; Adam et al. 2016). The insertion of a sensor into an artery or into the abdominal cavity requires general anaesthesia and surgery for implantation. If the sensor is connected to a telemetry unit, data can be read in real-time with the appropriate infrastructure (Beatty et al. 2006). More commonly the sensor is connected to a logger that has to be retrieved to access the stored data (Beatty et al. 2008a, 2008b; Fuller et al. 2011; Vesterdorf et al. 2011). Directly measuring Tcore is therefore difficult and impractical when large numbers of animals are involved. The handling of an animal for recording of rectal temperature (Trec) using handheld devices may increase body temperature, thus confounding the results. True body temperature must be collected in unrestrained animals at frequent intervals (Sherwin and Johnson 1990). An alternative is to measure at a site that is more easily accessible, such as the gastrointestinal tract. In monogastric animals, including humans, a device can be swallowed. That device passes through the gastrointestinal tract, and can provide one to 4 days of temperature recordings (Byrne and Lim 2007). In ruminants, such as sheep and cattle, devices inserted into the esophagus will settle in the rumen and can provide long-term recordings of rumen temperature (Trum) (Crock et al. 1992; Beatty et al. 2008a, 2008b; Cooper-Prado et al. 2011; Gonzales-Rivas et al. 2018; Lees et al. 2018, 2019, 2020). If the animals are destined for market, the loggers can be retrieved at slaughter, eliminating the need for surgery to retrieve the data.
The technique of measuring Trum involves the insertion of a rumen capsule that is fitted either with a thermologger that records, logs, and stores measurements (Beatty et al. 2008a, 2008b; Cooper-Prado et al. 2011), or a thermo-transmitter that senses temperature and sends a signal to a data receiving unit outside the animal in real time (Beatty et al. 2006; Signer et al. 2010; AlZahal et al. 2011; Turbill et al. 2011; Lees et al. 2018, 2019, 2020). As opposed to about an hour per animal for surgery to implant thermologgers into the abdominal cavity, the use of rumen capsules makes it possible to orally insert thermologgers into ∼30 sheep per hour using an applicator. As a non-invasive approach, the technique is time- and cost-effective, as well as imposing considerably less stress on the animal.
However, to be useful as a proxy for Tcore, Trum should vary in a consistent fashion with Tcore, regardless of changes in the various environmental factors that impact on an animal’s heat balance. Lees et al. (2018) found a distinct effect of time of day in the Trum of steers. Because variation in Tcore across the day/night cycle is important, it needs to be established whether the parameters of the circadian rhythm of Tcore are reflected in measures of Trum.
The heat of fermentation in the rumen accounts for approximately 8% of the total heat production in a ruminant (Czerkawski 1980). Due to that local heat production in the rumen, Trum has been reported to be approximately 1.0°C higher than Tcore or Trec in cattle (Beatty et al. 2008b), and approximately 0.5°C higher than Tcore in sheep (Beatty et al. 2008a). The difference between Tcore and Trum depends on the rate of local heat production in the rumen, and the rate of heat loss from the rumen, which will depend on the perfusion of the rumen and therefore on the rate of blood flow to the rumen. Beatty et al. (2008a) showed that the difference between Tcore and Trum remained quite consistent in sheep that were exposed to thermoneutrality and moderate heat (33°C dry-bulb temperature [Tdb, °C] and 25°C wet-bulb temperature [Twb, °C]). But whether the relationship remains the same at higher heat load, or when the composition of the diet changes, is unknown. Under heat load, an increase in peripheral blood flow is important for heat loss, and so any compensatory changes in rumen blood flow would be expected to impact on the heat balance of the rumen, and so on the rumen to core temperature difference.
We here test the hypothesis that, in sheep, the rumen to core temperature difference (designated Trum-core) is consistent when sheep are exposed to conditions that induce an increase in peripheral blood flow (Trial 1) or rumen heat production (Trial 2). We use data from 15 sheep that were fitted with loggers in the abdominal cavity (Tcore) and in the rumen (Trum) that measured temperature simultaneously, every 12 min. The sheep were exposed to Twb up to 26.8°C with a Tdb that exceeded the body temperature of the sheep (∼38.0°C; Lee 1950). In the first part of the experiment the sheep were either fleeced or recently shorn to determine the effect of fleece when ambient temperature (Ta; °C) is higher than body temperature. Being fleeced should insulate the sheep against heat gain from the environment under such conditions (Thwaites 1966), and so it might be expected that fleeced sheep would achieve heat balance with smaller redistribution of blood flow than shorn sheep.
In the second part of the experiment all of the sheep were shorn, and half received a change in diet from chaff to concentrated pellets, which will change the fermentation time and bacterial activity of the rumen, and thereby potentially change rumen heat production.
Materials and methods
The present work consisted of two separate trials, using the same animals in the two trials.
Animals
All experimental procedures were conducted in accordance with the guidelines in the Australian Code for the Care and Use of Animals for Scientific Purposes (National Health and Medical Research Council 2013) and were reviewed and approved by the animal ethics committee at Murdoch University (AEC approval number R2004/06).
Sixteen 2-year old Australia Merino-cross wethers (64.1 ± 6.7 kg; fleece length: ∼85 mm) were used. Most sheep exported from Australia are wethers. The use of wethers also removed any confounding effect of hormonal cycles. The sheep were obtained in the Southern Hemisphere spring, and all animals had spent the winter in the southern part of Western Australia. One week before the start of the experiments, eight of the sheep were randomly selected and shorn at the Murdoch University farm, Perth, Australia according to Australian industry standards using mechanical shears. The remaining eight sheep were kept with their fleece intact. The animals were then trucked (1 h drive) to the large animal facility at Murdoch University where they were housed for the duration of the experiment.
During the first part of the experiment (termed ‘Trial 1’; see below) the sheep were fed a commercial cube offered at 3% of body mass per day (on a dry matter basis) twice daily at 08:00 hours and 13:00 hours. The dietary regimen was chosen because it is similar to the diet that sheep consume on livestock vessels. The sheep were given one week on the diet prior to experimentation. A description of the feed administered during the second part of the experiment (termed ‘Trial 2’) is given below. Water was available ad libitum throughout both trials, and the water temperature never exceeded 26°C, which was comparable to water in troughs on livestock vessels (Beatty et al. 2007).
Climate control chambers
The 16 sheep were individually penned, with four shorn and four fleeced sheep in each of two climate chambers. The sheep had individual feed and water buckets and had room to turn around and lie down in the pens. Each pen was 0.88 m2, which is equal to the space per head on the livestock vessels. The sheep could not take food or water from neighbouring animals. Sawdust was used as bedding in all pens and was replaced or topped up as necessary.
The temperature and humidity in each chamber was controlled and adjusted to expose the sheep to gradually hotter conditions over each of the trials. The relative humidity (RH) was maintained at 55–60% and the Tdb was adjusted to achieve a given Twb in the rooms. The settings were adjusted at 08:00 hours each morning, and so for statistical purposes, each experimental day began at 08:00 hours and ended the next day at 07:59 hours. A low level of artificial lighting remained on for the duration of the experiment, as occurs on board livestock vessels (Beatty et al. 2007).
Environmental data loggers
An environmental data logger (HOBO Pro H08-032-08, Onset Computer Corp, Pocasset, MA, USA) was suspended in the centre of each climate chamber that measured Tdb and RH. The data loggers were set to log at 10 min intervals. At the end of the experiment, the data were downloaded from the two data loggers onto a personal computer (PC).
The Twb can be measured directly with a sling psychrometer, but can also be derived from electronic measures of Tdb and RH. The standard formulae that are available to derive Twb from Tdb and RH come with caveats that the formulae provide approximations only, and therefore are inaccurate under some conditions. The RH is easily derived from measures of Tdb and Twb, but working back the other way from Tdb and RH to Twb is not possible algebraically, and so deriving Twb from Tdb and RH requires an iterative process. We developed a macro in Excel using the ‘Solver’ function to perform this process. The iterative approach uses the following formulae, each taken from Barenbrug (1974):
The saturation water vapour pressure in kPa (esat) is calculated as
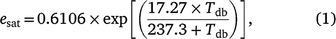
where exp is the base of natural logarithms (e) to the power of the number in the square brackets.
The water vapour pressure in an unsaturated atmosphere in kPa (e) is therefore (where RH is expressed as a proportion)

The relationship between e and the Tdb and Twb is given by
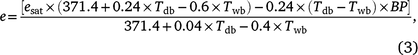
where BP is the barometric pressure in kPa. In the iterative process, e is obtained using formulae (1) and (2), and then formula (3) is solved iteratively to obtain Twb.
Body temperature loggers
Temperature loggers (iButton, DS1922L, Maximum Dallas, CA, USA) were implanted into the rumen and abdomen of all 16 sheep, one week before the start of the experiment. The temperature loggers remained in place until they were retrieved at slaughter.
Rumen temperature loggers
To measure rumen temperature, an iButton temperature logger was coated in biologically inert wax (EXP987, Sasol Wax Pty Ltd., South Africa) and inserted into an emptied control release capsule (CAPTEC, Nufarm Health and Sciences, NZ). The rumen temperature loggers were orally administered.
Abdominal temperature loggers
To measure abdominal temperature, an iButton temperature logger was coated in biologically inert wax. The loggers were surgically implanted into the peritoneal cavity via an incision in the region of the right paralumbar fossa. Each temperature logger was suspended proximal to the peritoneal wall with a short length of 0.40 mm non-absorbable suture (Vetafil, Bengen, Germany) that was incorporated into the outer layers of the wax coating of the logger and sutured into the peritoneal muscle layer. Details of the surgery, post-operative care and medication are given in Beatty et al. (2008a). Upon retrieval of the abdominal temperature loggers, it was noted that no scar tissue had formed around any of the loggers.
All of the body, rumen and abdominal temperature loggers were calibrated before, and again after, the experiment against a certified mercury-in-glass thermometer (certified by the National Association of Testing Authorities, Australia) with a resolution of 0.05°C in an insulated water bath at five temperatures over the range of 34–45°C. The loggers were programmed to log the temperature every 12 min. After retrieval from the sheep and calibration, data from the temperature loggers were downloaded onto a PC and each was adjusted according to its unique calibration. No drift between the initial and final calibration was observed in any of the loggers.
Experimental design
The aim of the experiment was to determine the relationship between Tcore and Trum under thermoneutral and hot and humid environmental conditions (high heat load, as may be experienced on board live export vessels; Stockman et al. 2011) in sheep that were shorn versus fleeced (‘Trial 1’), and in sheep that received a diet of either chaff or of pellets, with the same ME (‘Trial 2’).
Trial 1
The climate chambers were set to the prevailing ambient conditions for the first 3 days of the trial (thermoneutral; Day 1–3). During this time, the average daily Twb in each room was 14.9 ± 1.1°C. The Twb was then gradually increased from Days 4–7 (moderate heat load), and was then maintained at an average daily Twb of 25.4 ± 0.3°C from Day 8 to the end of the trial on Day 13 (high heat load). One shorn sheep was removed before the experiment began due to illness. On Day 11, all of the sheep were loaded onto a truck in the morning and driven 1 h to the University farm for shearing. The sheep that had been shorn prior to the start of the experiment were sham shorn to mimic the effect of shearing, while the remaining sheep were shorn. All of the sheep were then returned to their pens in the climate chambers where they were exposed to Twb of ∼25°C on Days 12 and 13. After Day 13 the sheep were brought back to the University farm where they remained from Day 14–16. During this time the sheep were exposed to the prevailing environmental conditions. The sheep were then returned to the climate chambers for Trial 2 early on the morning of Day 17.
Trial 2
Each sheep was returned to the same pen as Trial 1 in the climate chamber for the second trial, and randomly assigned to receive either a chaff based diet or a pellet based diet, irrespective of their treatment in the first trial. Because one sheep had been removed prior to the start of the experiment, eight sheep were fed pellets and the remaining seven sheep were fed chaff, twice daily at 08:00 hours and 13:00 hours. The diets did not differ in ME content and were offered at 3% of body mass on a dry matter basis per day. The diets were offered from Day 17.
For the initial 3 days of the trial (Day 17–19) the climate chambers were set to the prevailing ambient conditions. During this time, the average daily Twb was 12.9 ± 0.1°C. Then the Twb was increased gradually, from Day 20–23, to reach a maximum on Day 24 and Day 25 of 26.8 ± 0.1°C. The animals were brought back to the University farm on the morning of Day 26.
Analyses
Smoothing Trum
The difference between Trum and Tcore measured from the first part of the experiment was reported by Beatty et al. (2008a). Rapid, but transient, decreases in Trum were observed in all of the sheep and were directly related to water intake (Beatty et al. 2008a), as was previously found in cows (Cantor et al. 2018), potentially confounding the use of Trum to predict Tcore. We therefore tested whether the removal of the effect of water intake on the Trum made a difference to the analysis of Trum. The raw Trum for each individual sheep was run through a custom-written macro to identify rapid decreases in Trum that were associated with drinking. We looked initially at the distribution of changes in Trum between two subsequent measurements. The vast majority (95.2 ± 1.9%; average ± s.d.) were between −0.3 and +0.3°C. Changes larger than 0.4°C were always associated with a characteristic rapid decrease and then a logarithmic increase back to an asymptote over the subsequent ∼96 min. In the 15 experimental sheep, a decrease of more than 0.4°C in Trum was observed in 3.1 ± 0.8% of the collected data points, corresponding to a drinking bout on average every ∼ 6.5 h. A macro was then written to identify each incidence of a point-to-point decrease in Trum of >0.4°C, and the data points on either side of the rapid decrease, and gradual increase back to the prevailing Trum over the subsequent ∼96 min, were smoothed by interpolation. The smoothed Trum was termed Trum,smooth (°C) in further data analyses. The difference between Trum,smooth and Tcore was termed the Trum,smooth-core (°C).
Cosinor analyses over periods
We used a cosinor model (Nelson et al. 1979) to derive the parameters of the circadian rhythm (daily mesor, amplitude, and acrophase) of Tcore, Trum, Trum,smooth, Trum-core, and the Trum,smooth-core on selected days during each trial. The cosinor function was used to quantify the circadian temperature rhythm and remove the undue influence of single points that can occur when alternative approaches are used, such as analysing the ‘range’ of daily temperatures (Piccione and Caola 2003). For both trials the periods were: (1) thermoneutral, (2) moderate heat load, (3) high heat load.
Statistics
Daily means ± s.d. of the mesor, the amplitude of the Tcore, Trum, Trum,smooth, and the differences between the Trum and the Tcore and between Trum,smooth and the Tcore were calculated for each individual in each of the two trials. To test whether the mesor or the amplitude of the Trum changed with smoothing, two-way repeated measures (RM) ANOVAs were performed to compare the mesor and the amplitude, respectively, of the Trum against the Trum,smooth within individuals, regardless of treatment. Because there was no significant effect of smoothing on the mesor or on the amplitude of the Trum (see Results), all subsequent analyses were performed on the Trum. The means of the mesor and the amplitude of the Tcore and of the difference between Trum and Tcore were compared separately for Trial 1, using two-way RM ANOVAs with heat load in Periods 1, 2, and 3 (as defined above) as the repeated measure, and with the Greenhouse–Geiser correction. We used ‘shorn’ versus ‘fleeced’ as a fixed variable in the two-way RM ANOVAs.
Data from Trial 2 were separated and analysed in a similar way to those of Trial 1, except that we used repeated measures MANCOVA with ‘chaff’ versus ‘pellets’ as the fixed variable, and Periods 1, 2, and 3 as repeated measures.
Because the same sheep were used in the two trials we also tested for any carry-over effect of having been shorn early or late in Trial 1, using ‘shorn’ and ‘fleeced’ as variables in the RM MANCOVA.
For a detailed analysis of the relationship between Tcore and Trum, we took each simultaneous measurement of the two temperatures for each sheep, calculated the difference between the two temperatures (i.e. Trum − Tcore) and binned those differences into 0.1°C categories of Tcore. For each animal we then calculated the number at each category of Tcore, and the average ± s.d., minimum, and maximum of Trum − Tcore at each category of Tcore. We then did the same across sheep to determine if the difference between the temperatures remained constant across Tcore, and how common were differences that deviated from the general relationship. A Broken-Stick analysis was used to determine inflection points in the relationship between Trum-core and Tcore.
Statistical analyses for Trial 1 were performed using InStat Prism (7.0, GraphPad Software Inc., CA, USA), while analyses for Trial 2 were performed using Statistica (9.0, StatSoft Inc., Tulsa, OK, USA). Where appropriate, post hoc tests were performed using Tukey’s HSD test. Results were considered statistically significant when P < 0.05.
Results
The effect of correcting data for water intake
The effect of water intake on the circadian rhythm of Trum was obvious (Fig. 1, upper panel), and the macro that averaged out rapid decreases successfully removed those episodes (Fig. 1, lower panel). Nevertheless, correcting Trum for the effect of water intake had no effect on the parameters of the circadian rhythm of Trum. The average difference between the mesor of Trum and that of Trum-smooth was −0.07 ± 0.02°C (F1,28 = 0.5; P = 0.50), while the average difference between the daily amplitude of Trum and the daily amplitude of Trum-smooth was 0.01 ± 0.02°C (F1,28 = 0.8; P = 0.376). Similarly, there was no effect of smoothing on the estimate of the difference between Trum and Tcore, with the difference in mesor being −0.07 ± 0.03°C (F1,28 = 4.1; P = 0.053) and the difference in amplitude being 0.02 ± 0.01°C (F1,28 = 3.8; P = 0.060). Further analyses were therefore run using the original Trum.

|
Trial 1
In both treatment groups the mesor of Tcore increased with the increase in Twb (Fig. 2 – upper panel; F1.7,22.0 = 60.8, P < 0.0001), and the mesor of Tcore of the fleeced sheep was 0.6 ± 0.1°C higher than that of the shorn sheep during Periods 1 and 2 (F1,13 = 21.9, P = 0.0004). When the sheep were exposed to the maximum heat load conditions during Period 3, however, there was no effect of fleece on the Tcore of the animals. As such, while the Tcore of the fleeced animals did not increase from Period 2 to Period 3, the Tcore in the shorn sheep did (P = 0.04). The daily amplitude of the Tcore was not different between periods (F1.8,23.6 = 2.0, P = 0.16) and was not different between the fleeced and shorn animals at any period (F1,13 = 2.5, P = 0.14).

|
Like Tcore, the mesor of Trum also increased in both groups with each increase in Twb (data not shown; F1.7,21.5 = 55.4, P < 0.0001). The average mesor of Trum of the fleeced sheep was 0.6 ± 0.1°C higher than that of the shorn sheep (F1,13 = 15.8, P = 0.0016). The amplitude of the circadian rhythm of the Trum was not different between periods (F2.0,25.8 = 0.27, P = 0.77) or affected by fleece (F1,13 = 0.54, P = 0.48).
The increase in heat load between the periods had no effect on the difference between Trum and Tcore, despite the increase in Tcore with increasing heat load (Fig. 2 – lower panel; mesor Trum-core: F1.2,14.6 = 0.44, P = 0.54; amplitude: F1.5,19.5 = 0.80, P = 0.43). Being fleeced or shorn also had no affect on the difference between Trum and Tcore (Fig. 2 – lower panel; mesor Trum-core: F1,13 = 3.95, P = 0.07; amplitude: F1,13 = 0.04, P = 0.84). The Trum-core remained stable despite increasing heat load. The grand mean of the mesor of the Trum-core in the sheep was 0.5 ± 0.1°C.
Trial 2
The mesor of the Tcore was different across the periods (Fig. 3; F2,22 = 79.3, P < 0.0001), but was not affected by diet (F1,11 = 1.6, P = 0.23) or by the treatment in Trial 1 (F1,11 = 0.22, P = 0.65). The mesor of Tcore was higher during Period 2 than during Period 1 (P = 0.0001), and was lower in Period 3 than during Period 2 (P = 0.0002).

|
The amplitude of the circadian rhythm of Tcore also increased with increased heat load (data not shown); F2,22 = 4.9, P = 0.02), and was affected by diet (F1,11 = 11.3; P = 0.006), but not by previous wool treatment (F1,11 = 0.23, P = 0.64). Although the ANOVA detected an effect of diet on the amplitude of Tcore, post hoc Tukey’s HSD testing failed to detect any difference in the amplitude of Tcore between the diets within any of the three periods (F2,22 = 2.5; P = 0.1).
The mesor of Trum was affected by period (F1.7,22.3 = 35.3, P < 0.0001), and was not affected by diet (F1,13 = 3.70, P = 0.08). The amplitude of the circadian rhythm of Trum also increased with increased heat load (F1.9,25.2 = 7.72, P = 0.003), and was not affected by diet (F1,13 = 0.021, P = 0.88).
The difference between Trum and Tcore was different between the periods (Fig. 3 – lower panel; F1.8,23.6 = 9.7, P = 0.001), but there was no effect of diet (F1,13 = 1.3, P = 0.28) or previous wool treatment (F1,11 = 2.4, P = 0.16). A post hoc Tukey’s HSD test revealed that the difference between the Trum and the Tcore was higher during Period 1 than during Period 2 by 0.2 ± 0.03°C (P = 0.0002). The difference between Trum and Tcore during Period 3 was not different from Period 1 or Period 2.
There was no effect of increasing heat load on the amplitude of Trum-core (F1.6,21.0 = 1.118, P = 0.33), or of diet (F1,13 = 4.730, P = 0.05).
The range of the difference between Trum and Tcore during the Trial 2 was 0.3–1.0°C, with a grand mean of 0.5 ± 0.2°C, irrespective of the effect of diet or previous wool treatment.
The Trum-core difference during the entire experiment
Because there was no effect of either the fleece or diet on the difference between the Trum and the Tcore in the sheep, we then calculated the overall average and standard deviation for each individual sheep during the duration of the entire experimental period (25 days; Fig. 4). The grand average Trum-core difference in all sheep was 0.5 ± 0.1°C during the entire experimental period.

|
The Trum-core difference at categories of Tcore
A general relationship emerged with the difference between Trum and Tcore being higher at the lower Tcore (P < 0.05; Fig. 5), decreasing to a plateau of a difference of 0.5 ± 0.1°C in the range of Tcore higher than 38.5°C and lower than 41.1°C. The vast majority of observations (92%) occurred when Tcore was between 38.5°C and 41.1°C, with 7.9% below 38.4°C, and 0.03% above 41.1°C. The Trum was therefore consistently 0.5°C higher than the Tcore across the normal range of Tcore. The Trum-core difference was smaller than 0.5°C when Tcore exceeded 41.1°C (P < 0.05; Fig. 5).

|
Discussion
This study was conducted to determine the relationship between Tcore and Trum in sheep. Because the measurement of Trum is less invasive than the measurement of Tcore, which requires surgery for the placement of a logger in the abdominal cavity or an artery, there are advantages to using Trum. The live export of sheep from Australia to destinations around the world can expose sheep to heat stress when vessels near or cross the equator (Stockman et al. 2011). The routine measurement of Trum is a feasible strategy to monitor the physiological/thermal responses of sheep during voyages, while the measurement of Tcore would require surgery on each animal. However, before the measurement of Trum is acceptable, we need to assess if Trum can be used as a proxy for Tcore in sheep.
There are some studies in cattle suggesting that Trum is a useful proxy for Trec (Lees et al. 2018, 2019, 2020). Presumably the temperature in the rectum will be subject to most of the same factors as will temperature in the rumen, especially changes in the distribution of blood flow to the gastrointestinal tract when blood is diverted to the periphery during exposure to high Ta, and so while the temperature measured in those two locations might be well correlated, it does not mean that they will be correlated with Tcore. For example, the use of Trec as a proxy for Tcore in sheep can result in an overestimation of the use of selective brain cooling because Trec always exceeded arterial blood temperature (Maloney et al. 2001).
We show here that when Tcore increased during exposure to heat load, so did the Trum and so, despite relatively large changes in overall body temperature, the Trum-core difference did not change. The grand mean of the difference between Trum and Tcore was 0.5 ± 0.1°C across a range of individuals and environmental conditions similar to those experienced by sheep on live export vessels, and irrespective of length of wool or different diet composition.
The Twb that our sheep were exposed to was higher than the 25.5°C threshold for an increase in morbidity and mortality identified during live export but lower than the Twb that is often experienced near the Equator (Maunsell Australia 2003). During the days of high heat load the body temperature of the sheep increased relative to thermoneutral conditions, indicating that the conditions exceeded the thermoneutral zone (Mitchell et al. 2018). The central question in this study is whether the state of the animal’s body temperature can be inferred from measurements of Trum.
The applicability of Trum as a proxy for Tcore
To establish Trum as a valid proxy for Tcore in sheep, the relationship between the two needed to be validated over a range of conditions. Any change in Ta stimulates various physiological responses that allow mammals to maintain thermal homeostasis in the altered environment. These responses include changes in circulatory (Hales 1973; Von Engelhardt and Hales 1977) and respiratory (Hales 1973; Vesterdorf et al. 2011) function in ruminants. As such, rumen pH and Trum changes have been observed in response to heat stress in cattle (e.g. Beatty et al. 2006, 2008b; Baumgard and Rhoads 2013), goats (e.g. Salama et al. 2014; Castro-Costa et al. 2015), and sheep (e.g. Hales 1973; Von Engelhardt and Hales 1977; Beatty et al. 2008a; Vesterdorf et al. 2011).
The effect of drinking
The drinking of water at room temperature by our sheep resulted in a sharp but transient decrease in Trum. These decreases in Trum were always larger than 0.4°C and of approximately 90 min duration, from the initial rapid decrease and back to an asymptote. The Trum in heat stressed goats that were provided with water for only for 30 min per day decreased rapidly when water was provided and then increased back to the normal range over the subsequent 2 h (Castro-Costa et al. 2015). The logical explanation for the temperature decrease is that, because the water was cooler than the body and rumen, heat was transferred from the rumen contents to the ingested water, resulting in the rapid decrease in temperature. Because the Trum would then have been lower than temperature in the surrounding body, heat would diffuse into the rumen and cause the gradual asymptote as the ruminal contents re-established a thermal steady state with the body. Similar observations have been reported in previous studies in cattle (Bianca 1964; Beatty et al. 2008b; Cooper-Prado et al. 2011) and goats (Castro-Costa et al. 2015).
While there was a clear effect on Trum when a sheep consumed water, the transient nature of the temperature change had no impact on the estimate of any of the parameters of the circadian rhythm of Trum. Beatty et al. (2008a) found ‘spike-like’ decreases in average hourly Trum that explained 10–20% of the variability in hourly Tcore in sheep. By analysis of Trum collected at 12-min intervals, instead of hourly averages, we discerned more clearly the timing, length, and range of these ‘spike-like’ decreases. We compared the effect on the circadian rhythm of Trum of ‘smoothing’ the Trum, by eliminating those spike-like decreases from the data set, against that of the raw, ‘unsmoothed’, Trum. We found that there was no effect of drinking on the circadian rhythm of Trum or indeed on the difference between Trum and Tcore.
Rumen heat balance
Exposure to heat stress affects rumen function in sheep through changes in rumen blood flow and Trum (Hales 1973; Von Engelhardt and Hales 1977; Beatty et al. 2008a; Lees et al. 2020), as well as decreases in digestibility, motility, bacterial activity, and pH (Bernabucci et al. 2009). In cattle, exposure to heat load conditions resulted in increased Trum and lower feed intake, which is often interpreted as a thermoregulatory adaptation that reduces the heat increment of feeding (Mendel and Raghavan 1964; Beatty et al. 2006, 2008b; Gonzales-Rivas et al. 2018; Lees et al. 2018, 2019). In our study, however, increased heat load did not affect feed intake in the sheep during either of the trials. Similar observations were made in goats, also fed to maintenance level and exposed to a similar heat load as that used in our study (Castro-Costa et al. 2015).
Exposure to heat load initiates a series of thermoregulatory responses, including a redistribution of blood flow to the periphery that facilitates heat loss across the skin and mucosal membranes (Hales 1973; Von Engelhardt and Hales 1977) and increased respiratory frequency that facilitates respiratory evaporative heat loss (Vesterdorf et al. 2011). Rumen temperature likely depends on the rate of local heat production in the rumen by fermentation and the rate of heat loss from the rumen. With a change in ruminal blood flow during heat exposure, heat produced from fermentation is less readily removed from the rumen, resulting in higher Trum. The temperature of the rumen therefore depends on the level of blood flow to and from the rumen relative to the heat production (Maloney et al. 2001). When blood flow is redirected away from the rumen during heat exposure, the Trum should increase because heat exchange between the rumen and the surrounding body will decrease. That increase in Trum will then affect the relationship between Trum and Tcore.
Effect of fleece
When Ta is lower than the body temperature (Tb; °C) the presence of fleece will reduce heat flow from the skin to the environment, but when Ta exceeds Tb that fleece will reduce heat flow from the environment into the skin (Robertshaw 1968; Von Engelhardt and Hales 1977). A fully fleeced sheep should therefore rely more on respiratory evaporative heat loss (REHL) at thermoneutrality than a shorn sheep (Hales 1973; Vesterdorf et al. 2011). But when the Ta is higher than Tb, a fleeced sheep should have a lower heat load than a shorn sheep, and thus require smaller REHL to achieve heat balance. Comparing the Trum-core difference in fleeced versus shorn sheep when they are exposed to heat stress should reveal whether differences in blood flow to the rumen result in changes in Trum relative to Tcore. In our study, the Tcore of shorn sheep was 0.6 ± 0.1°C lower than that of fleeced sheep during the thermoneutral Period 1 and the moderate heat load in Period 2, reflecting a relatively higher degree of heat loss from the body of the shorn sheep that clearly was not compensated for by higher REHL in the fleeced sheep. During exposure to high heat load, the Tcore of shorn and fleeced sheep was the same, and there was no difference in the Trum-core.
Effect of diet
Different composition of a diet has the potential to affect the capacity to cope with heat stress. Particle size can affect the extent of rumen fermentation and retention time in the rumen (Kaske et al. 1992). As such, changes in particle size in the diet may cause changes to the bacterial activity of the rumen, along with fermentation, digestibility, and ruminal flow-through rate, which in turn would affect the heat production of the rumen and hence rumen heat balance (Silanikove 1985). There was, however, no effect of a change in dietary composition on the Trum or indeed the difference between the Trum and the Tcore in the sheep in the present study. Our findings are corroborated by studies in goats kept on either high or low roughage diets (Castro-Costa et al. 2015) and in cattle kept on either completely pelletised or mixed roughage diets (Bonfante et al. 2016) where no change in Trum was detected.
The Trum-core difference
The difference between Trum and Tcore remained constant at 0.5°C despite increasing heat load in sheep during the fleece experiment in Trial 1. During Trial 2, while there was no effect of diet on the Trum-core difference, that difference decreased by 0.2°C from Period 1 to Period 2, and then did not change into Period 3. Because the Trum-core difference in Period 2 and 3 of Trial 2 was similar to the Trum-core measured during all of Trial 1, it seems that the anomaly was the larger Trum-core difference in Period 1 of Trial 2. During Period 1 of Trial 2 the Tcore was lower than it had been in Trial 1 and was in the other periods of Trial 2. As a result the data from Period 1 of Trial 2 were all toward the left side of Fig. 5, and we do not think that the data reflect any impact of the diet, per se, on the Trum-core difference, but rather reflect the relatively low Tcore.
It was apparent that sheep in both Trial 1 and 2 adjusted to the high heat load with the onset of thermoregulatory mechanisms, including increased respiratory frequency that allowed the sheep to lose heat via REHL (Vesterdorf et al. 2011). That the sheep were affected by heat load was evident from the higher Tcore and Trum, and the increased daily amplitude of both, irrespective of fleece length or diet.
For the vast majority of simultaneous observations of Trum and Tcore when Tcore was within the normal range for sheep (that is, in 92% of observations when Tcore was between 38.5°C and 41.1°C) in our study, the difference between Trum and Tcore was within the 0.45–0.55°C range. At the few observations of high Tcore (0.03%; higher than 41.1°C), the Trum-core difference was smaller than 0.5°C. Similarly, for the few observations at low Tcore (8%; lower than 38.5°C), the Trum-core difference was higher than 0.5°C. The low and the high Tcore observations were associated with exposure to low and high Twb exposure, respectively.
Above a Tcore of 41.1°C, the average Trum-core decreased, suggesting either that heat production in the rumen decreased or heat loss from the rumen to the general body increased. The use of Trum to infer Tcore at these high body temperatures would lead to an overestimation of Tcore. While a small proportion of our data (0.03%) were in that category, the Twb onboard livestock vessels can exceed 27°C when ships are near the equator (Maunsell Australia 2003), and we do not know whether the Trum-core difference continues to decline as Twb increases further. Conversely, at Tcore lower than 38.5°C, the Trum-core difference increased, suggesting either that heat production in the rumen increased or that heat loss from the rumen to the general body decreased. The use of Trum to infer Tcore at these low body temperatures would lead to underestimation of Tcore.
Because the major welfare concern during live export is heat stress (Stockman et al. 2011), and the associated high body temperatures, it is unlikely that the underestimation of Tcore at low body temperature would be an important issue with the use of Trum during live export.
Conclusion
The use of thermo-transmitters located in the rumen of sheep would provide a direct, real-time method for the continuous monitoring of sheep during transport onboard live export vessels. We show here that the daily average relationship between Trum and Tcore is not affected by water ingestion, length of fleece, diet composition, or heat load. Management could directly monitor values of the Trum and could, by subtraction of the 0.5°C difference shown in this study, obtain a reliable measure of the Tcore of the sheep at all times of day, providing a live and reliable indication of the thermal status of the animals.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This research did not receive any specific funding.
Acknowledgements
The authors wish to thank Malcolm Boyce, Donald Hook, Eric Taylor, and Kim Thomas for their assistance during the experimental work in the laboratory and on the Murdoch University farm.
References
Adam D, Johnston SD, Beard L, Nicholson V, Lisle A, Gaughan J, Larkin R, Theilemann P, McKinnon A, Ellis W (2016) Surgical implantation of temperature-sensitive transmitters and data-loggers to record body temperature in koala (Phascolarctos cinereus). Australian Veterinary Journal 94, 42–47.| Surgical implantation of temperature-sensitive transmitters and data-loggers to record body temperature in koala (Phascolarctos cinereus).Crossref | GoogleScholarGoogle Scholar | 26781067PubMed |
AlZahal O, AlZahal H, Steele MA, Van Schaik M, Kyriazakis I, Duffield TF, McBride BW (2011) The use of a radiotelemetric ruminal bolus to detect body temperature changes in lactating dairy cattle. Journal of Dairy Science 94, 3568–3574.
| The use of a radiotelemetric ruminal bolus to detect body temperature changes in lactating dairy cattle.Crossref | GoogleScholarGoogle Scholar | 21700044PubMed |
Barenbrug AWT (1974) ‘Psychrometry and psychrometric charts.’ 3rd edn. (Chamber of Mines of South Africa: Johannesburg, South Africa)
Baumgard LH, Rhoads RP (2013) Effects of heat stress on postabsorptive metabolism and energetics. Annual Review of Animal Biosciences 1, 311–337.
| Effects of heat stress on postabsorptive metabolism and energetics.Crossref | GoogleScholarGoogle Scholar | 25387022PubMed |
Beatty DT, Barnes A, Taylor E, Pethick D, McCarthy M, Maloney SK (2006) Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity. Journal of Animal Science 84, 972–985.
| Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity.Crossref | GoogleScholarGoogle Scholar | 16543576PubMed |
Beatty DT, Barnes A, Taplin R, McCarthy M, Maloney SK (2007) Electrolyte supplementation of live export cattle to the Middle East. Australian Journal of Experimental Agriculture 47, 119–124.
| Electrolyte supplementation of live export cattle to the Middle East.Crossref | GoogleScholarGoogle Scholar |
Beatty DT, Barnes A, Fleming PA, Taylor E, Maloney SK (2008a) The effect of fleece on core and rumen temperature in sheep. Journal of Thermal Biology 33, 437–443.
| The effect of fleece on core and rumen temperature in sheep.Crossref | GoogleScholarGoogle Scholar |
Beatty DT, Barnes A, Taylor E, Maloney SK (2008b) Do changes in feed intake or ambient temperature cause changes in cattle rumen temperature relative to core temperature? Journal of Thermal Biology 33, 12–19.
| Do changes in feed intake or ambient temperature cause changes in cattle rumen temperature relative to core temperature?Crossref | GoogleScholarGoogle Scholar |
Bernabucci U, Lacetera N, Danieli PP, Bani P, Nardone A, Ronchi B (2009) Influence of different periods of exposure to hot environment on rumen function and diet digestibility in sheep. International Journal of Biometeorology 53, 387–395.
| Influence of different periods of exposure to hot environment on rumen function and diet digestibility in sheep.Crossref | GoogleScholarGoogle Scholar | 19370363PubMed |
Bianca W (1964) Thermoregulatory responses of the dehydrated ox to drinking cold and warm water in a warm environment. Research in Veterinary Science 5, 75–80.
| Thermoregulatory responses of the dehydrated ox to drinking cold and warm water in a warm environment.Crossref | GoogleScholarGoogle Scholar |
Bonfante E, Palmonari A, Mammi L, Canestrari G, Fustini M, Formigoni A (2016) Effects of a completely pelleted diet on growth performance in Holstein heifers. Journal of Dairy Science 99, 9724–9731.
| Effects of a completely pelleted diet on growth performance in Holstein heifers.Crossref | GoogleScholarGoogle Scholar | 27692717PubMed |
Byrne C, Lim CL (2007) The ingestible telemetric body core temperature sensor: a review of validity and exercise applications. British Journal of Sports Medicine 41, 126–133.
| The ingestible telemetric body core temperature sensor: a review of validity and exercise applications.Crossref | GoogleScholarGoogle Scholar | 17178778PubMed |
Cantor MC, Costa JHC, Bewley JM (2018) Impact of observed and controlled water intake on reticulorumen temperature in lactating dairy cattle. Animals 8, 194
| Impact of observed and controlled water intake on reticulorumen temperature in lactating dairy cattle.Crossref | GoogleScholarGoogle Scholar |
Castro-Costa A, Salama AAK, Moll X, Aguilo J, Caja G (2015) Using wireless rumen sensors for evaluating the effects of diet and ambient temperature in nonlactating dairy goats. Journal of Dairy Science 98, 4646–4658.
| Using wireless rumen sensors for evaluating the effects of diet and ambient temperature in nonlactating dairy goats.Crossref | GoogleScholarGoogle Scholar | 25958285PubMed |
Cooper-Prado MJ, Long NM, Wright EC, Goad CL, Wettemann RP (2011) Relationship of ruminal temperature with parturition and estrus of beef cows. Journal of Animal Science 89, 1020–1027.
| Relationship of ruminal temperature with parturition and estrus of beef cows.Crossref | GoogleScholarGoogle Scholar | 21169512PubMed |
Crock PL, Bassingthwaighte J, Hasker P, Lennard C, Ellis KJ (1992) Use of electronic ID transponders to test long term retention of intra-ruminal controlled release devices. In ‘Proceedings of the International Symposium on Controlled Release of Bioactive Materials’. vol. 19, pp. 437–440. (Controlled Release Society Inc.)
Czerkawski JW (1980) A novel estimate of the magnitude of heat produced in the rumen. British Journal of Nutrition 43, 239–243.
| A novel estimate of the magnitude of heat produced in the rumen.Crossref | GoogleScholarGoogle Scholar | 6768380PubMed |
Fuller A, Hetem RS, Meyer LCR, Maloney SK (2011) Angularis oculi vein blood flow modulates the magnitude but not the control of selective brain cooling in sheep. American Journal of Physiology Regulatory, Integrative and Comparative Physiology 300, R1409–R1417.
| Angularis oculi vein blood flow modulates the magnitude but not the control of selective brain cooling in sheep.Crossref | GoogleScholarGoogle Scholar | 21368272PubMed |
Gonzales-Rivas PA, Sullivan M, Cottrell JJ, Leury BJ, Gaughan JB, Dunshea FR (2018) Effect of feeding slowly fermentable grains on productive variables and amelioration of heat stress in lactating dairy cows in a sub-tropical summer. Tropical Animal Health and Production 50, 1763–1769.
| Effect of feeding slowly fermentable grains on productive variables and amelioration of heat stress in lactating dairy cows in a sub-tropical summer.Crossref | GoogleScholarGoogle Scholar |
Hales JRS (1973) Effects of exposure to hot environments on the regional distribution of blood flow and on cardiorespiratory function in sheep. Pflügers Archiv 344, 133–148.
| Effects of exposure to hot environments on the regional distribution of blood flow and on cardiorespiratory function in sheep.Crossref | GoogleScholarGoogle Scholar |
Kaske M, Hatiboglu S, Engelhardt WV (1992) The influence of density and size of particles on rumination and passage from the reticulo-rumen of sheep. British Journal of Nutrition 67, 235–244.
| The influence of density and size of particles on rumination and passage from the reticulo-rumen of sheep.Crossref | GoogleScholarGoogle Scholar | 1317720PubMed |
Koltes JE, Koltes DA, Mote BE, Tucker J, Hubbell DS (2018) Automated collection of heat stress data in livestock: new technologies and opportunities. Translational Animal Science 2, 319–323.
| Automated collection of heat stress data in livestock: new technologies and opportunities.Crossref | GoogleScholarGoogle Scholar | 32704715PubMed |
Lee DHK (1950) Studies of heat regulation in the sheep, with special reference to the merino. Australian Journal of Agricultural Research 1, 200–216.
| Studies of heat regulation in the sheep, with special reference to the merino.Crossref | GoogleScholarGoogle Scholar |
Lees AM, Lees JC, Lisle AT, Sullivan ML, Gaughan JB (2018) Effect of heat stress on rumen temperature of three breeds of cattle. International Journal of Biometeorology 62, 207–215.
| Effect of heat stress on rumen temperature of three breeds of cattle.Crossref | GoogleScholarGoogle Scholar | 28918576PubMed |
Lees AM, Sejian V, Lees JC, Sullivan ML, Lisle AT, Gaughan JB (2019) Evaluating rumen temperature as an estimate of core body temperature in Angus feedlot cattle during summer. International Journal of Biometeorology 63, 939–947.
| Evaluating rumen temperature as an estimate of core body temperature in Angus feedlot cattle during summer.Crossref | GoogleScholarGoogle Scholar | 30868342PubMed |
Lees AM, Sullivan ML, Olm JCW, Cawdell-Smith AJ, Gaughan JB (2020) The influence of heat load on Merino sheep. 2. Body temperature, wool surface temperature and respiratory dynamics. Animal Production Science 60, 1932–1939.
| The influence of heat load on Merino sheep. 2. Body temperature, wool surface temperature and respiratory dynamics.Crossref | GoogleScholarGoogle Scholar |
Lim CL, Byrne C, Lee JKW (2008) Human thermoregulation and measurement of body temperature in exercise and clinical settings. Annals Academy of Medicine Singapore 37, 347–353.
Maloney SK, Fuller A, Mitchell G, Mitchell D (2001) Rectal temperature measurement results in artifactual evidence of selective brain cooling. American Journal of Physiology Regulatory, Integrative and Comparative Physiology 281, R108–R114.
| Rectal temperature measurement results in artifactual evidence of selective brain cooling.Crossref | GoogleScholarGoogle Scholar | 11404284PubMed |
Maloney SK, Meyer LCR, Blache D, Fuller A (2013) Energy intake and the circadian rhythm of core body temperature in sheep. Physiological Reports 1, e00118
| Energy intake and the circadian rhythm of core body temperature in sheep.Crossref | GoogleScholarGoogle Scholar | 24303185PubMed |
Maloney SK, Marsh MK, McLeod SR, Fuller A (2017) Heterothermy is associated with reduced fitness in wild rabbits. Biology Letters 13, 20170521
| Heterothermy is associated with reduced fitness in wild rabbits.Crossref | GoogleScholarGoogle Scholar | 29212751PubMed |
Maloney SK, Goh G, Fuller A, Vesterdorf K, Blache D (2019) Amplitude of the circadian rhythm of temperature in homeotherms. CAB Reviews 14, 1–30.
| Amplitude of the circadian rhythm of temperature in homeotherms.Crossref | GoogleScholarGoogle Scholar |
Marai IFM, El-Darawany AA, Fadiel A, Abdel-Hafez MAM (2007) Physiological traits as affected by heat stress in sheep – a review. Small Ruminant Research 71, 1–12.
| Physiological traits as affected by heat stress in sheep – a review.Crossref | GoogleScholarGoogle Scholar |
Marai IFM, El-Darawany AA, Fadiel A, Abdel-Hafez MAM (2008) Reproductive performance traits as affected by heat stress and its alleviation in sheep. Tropical and Subtropical Agroecosystems 8, 209–234.
Maunsell Australia (2003) ‘Development of a heat stress risk management model.’ (Meat and Livestock Australia Ltd., Australia)
Mendel VE, Raghavan GV (1964) A study in diurnal temperature patterns in sheep. Journal of Physiology 174, 206–216.
| A study in diurnal temperature patterns in sheep.Crossref | GoogleScholarGoogle Scholar | 14244118PubMed |
Mitchell D, Snelling EP, Hetem RS, Maloney SK, Strauss WM, Fuller A (2018) Revisiting concepts of thermal physiology: predicting responses of mammals to climate change. Journal of Animal Ecology 87, 956–973.
| Revisiting concepts of thermal physiology: predicting responses of mammals to climate change.Crossref | GoogleScholarGoogle Scholar | 29479693PubMed |
National Health and Medical Research Council (2013) ‘Australian code for the care and use of animals for scientific purposes.’ 8th edn. (National Health and Medical Research Council: Canberra)
Nelson W, Tong YL, Lee J-K, Halberg F (1979) Methods for cosinor-rhythmometry. Chronobiologica 6, 305–323.
Piccione G, Caola G (2003) Influence of shearing on the circadian rhythm of body temperature in the sheep. Journal of Veterinary Medicine. A, Physiology, Pathology, Clinical Medicine 50, 235–240.
| Influence of shearing on the circadian rhythm of body temperature in the sheep.Crossref | GoogleScholarGoogle Scholar | 14567509PubMed |
Robertshaw D (1968) The pattern and control of sweating in the sheep and goat. Journal of Physiology 198, 531–539.
| The pattern and control of sweating in the sheep and goat.Crossref | GoogleScholarGoogle Scholar | 5685286PubMed |
Salama AAK, Caja G, Hamzaoui S, Badaoui B, Castro-Costa A, Facanha DAE, Guilhermino MM, Bozzi R (2014) Different levels of response to heat stress in dairy goats. Small Ruminant Research 121, 73–79.
| Different levels of response to heat stress in dairy goats.Crossref | GoogleScholarGoogle Scholar |
Sherwin CM, Johnson KG (1990) Skin and abdominal temperatures recorded by data loggers attached to Merino sheep voluntarily staying out of shade. Australian Journal of Agricultural Research 41, 781–790.
| Skin and abdominal temperatures recorded by data loggers attached to Merino sheep voluntarily staying out of shade.Crossref | GoogleScholarGoogle Scholar |
Signer C, Ruf T, Schober F, Fluch G, Paumann T, Arnold W (2010) A versatile telemetry system for continuous measurement of heart rate, body temperature and locomotor activity in free-ranging ruminants. Methods in Ecology and Evolution 1, 75–85.
| A versatile telemetry system for continuous measurement of heart rate, body temperature and locomotor activity in free-ranging ruminants.Crossref | GoogleScholarGoogle Scholar | 22428081PubMed |
Silanikove N (1985) Effect of dehydration on feed intake and dry matter digestibility in desert (black Bedouin) and non-desert (Swiss saanen) goats fed on lucerne hay. Comparative Biochemistry and Physiology Part A: Physiology 80, 449–452.
| Effect of dehydration on feed intake and dry matter digestibility in desert (black Bedouin) and non-desert (Swiss saanen) goats fed on lucerne hay.Crossref | GoogleScholarGoogle Scholar |
Silanikove N (2000) Effects of heat stress on the welfare of extensively managed domestic ruminants. Livestock Production Science 67, 1–18.
| Effects of heat stress on the welfare of extensively managed domestic ruminants.Crossref | GoogleScholarGoogle Scholar |
Stockman CA, Barnes AL, Maloney SK, Taylor E, McCarthy M, Pethick P (2011) Effect of prolonged exposure to continuous heat and humidity similar to long haul live export voyages in Merino wethers. Animal Production Science 51, 135–143.
| Effect of prolonged exposure to continuous heat and humidity similar to long haul live export voyages in Merino wethers.Crossref | GoogleScholarGoogle Scholar |
Thwaites CJ (1966) Fleece length and the reactions of sheep to wet and dry heat. Nature 211, 997–998.
| Fleece length and the reactions of sheep to wet and dry heat.Crossref | GoogleScholarGoogle Scholar | 5968327PubMed |
Turbill C, Ruf T, Mang T, Arnold W (2011) Regulation of heart rate and rumen temperature in red deer: effects of season and food intake. Journal of Experimental Biology 214, 963–970.
| Regulation of heart rate and rumen temperature in red deer: effects of season and food intake.Crossref | GoogleScholarGoogle Scholar | 21346124PubMed |
Vesterdorf K, Blache D, Maloney SK (2011) The cranial arterio-venous temperature difference is related to respiratory evaporative heat loss in a panting species, the sheep (Ovis aries). Journal of Comparative Physiology B 181, 277–288.
| The cranial arterio-venous temperature difference is related to respiratory evaporative heat loss in a panting species, the sheep (Ovis aries).Crossref | GoogleScholarGoogle Scholar |
Von Engelhardt W, Hales JR (1977) Partition of capillary blood flow in rumen, reticulum, and omasum of sheep. American Journal of Physiology 232, E53–E56.
| Partition of capillary blood flow in rumen, reticulum, and omasum of sheep.Crossref | GoogleScholarGoogle Scholar | 835704PubMed |