

Acute stress induced by the preslaughter use of electric prodders causes tougher beef meat
R. D. Warner A B E , D. M. Ferguson C , J. J. Cottrell B and B. W. Knee DA Cooperative Research Centre for Cattle and Beef Quality.
B Department of Primary Industries, 600 Sneydes Road, Werribee, Vic. 3030, Australia.
C CSIRO Livestock Industries, F.D. McMaster Laboratory, Chiswick, Armidale, NSW 2350, Australia.
D Department of Primary Industries, Mount Napier Road, Hamilton, Vic. 3300, Australia.
E Corresponding author. Email: robyn.warner@dpi.vic.gov.au
Australian Journal of Experimental Agriculture 47(7) 782-788 https://doi.org/10.1071/EA05155
Submitted: 10 June 2005 Accepted: 28 March 2007 Published: 2 July 2007
Abstract
Adrenergic activation and hormone release preslaughter is an inevitable outcome of the systems used to move cattle to slaughter. The aim of this experiment was to investigate the effects of acute preslaughter stress in beef cattle on postmortem muscle metabolism and the meat quality, including consumer-assessed eating quality. Eighty-four cattle were used on three separate days, with ‘mobs’ of four cattle allocated to either a ‘control’ (no electric goads used preslaughter) or a ‘stress’ (six prods given with an electric goad over 5–10 min) treatment at 15 min preslaughter. Cattle undergoing the ‘stress’ treatment had higher plasma lactate at slaughter. The prerigor pH and temperature, ultimate pH and temperature at rigor of the longissimus thoracis muscle were similar between treatments (P > 0.05 for all). The water-holding capacity of the longissimus lumborum was reduced by the ‘stress’ treatment, as indicated by higher levels of water lost during suspension (drip loss), storage (purge) for 21 days and cooking (cooking loss at 1 day postslaughter) (P < 0.05 for all). ‘Stress’ cattle produced longissimus lumborum muscle with similar sarcomere lengths and Warner–Bratzler shear force at 2, 6 and 21 days, compared to ‘control’ cattle (P < 0.05 for all). The longissimus lumborum muscle of cattle undergoing the ‘stress’ treatment was rated less tender, less juicy, with a less acceptable flavour, a lower ‘liking’ and a lower MQ4 score (P < 0.05 for all). The ‘bloomed’ surface colour (CIE L*, a*, b*) of the longissimus lumborum muscle at 2, 6 and 21 days postslaughter was similar between the ‘stress’ and ‘control’ treatments (P > 0.05 for all). In conclusion, cattle subjected to acute preslaughter stress using electric goads produced meat which the consumer rated as tougher with inferior quality. The inferior quality induced by the acute stress treatment was associated with reduced water-holding capacity but was independent of muscle pH and temperature.
Introduction
Adrenergic activation and hormone release is an inevitable outcome during the preslaughter handling of cattle. Cattle must arrive in a continuous stream at the stunning point (Gregory 1998) and most animals should walk calmly into a stunning pen or restrainer, preferably without the use of an electric goad (Grandin 1998). Although it is desirable to minimise the use of electric goads during the unloading and loading of trucks and preslaughter handling, inevitably they are still applied to varying degrees in many Australian beef abattoirs.
The negative effect of stress on beef quality has previously been attributed to the mobilisation of muscle glycogen during stress, the lack of acidification postmortem and the occurrence of ‘dark-cutting’ or high ultimate pH beef. Warriss (1990) states ‘the major influence of preslaughter handling on lean meat quality is through the potential effect on muscle glycogen stores’. Aside from the unacceptably dark colour of dark-cutting meat, it also has reduced eating quality (bland flavour, variable tenderness) in addition to reduced keeping quality (Tarrant 1989a). For these reasons, meat with an ultimate pH > 5.7 is excluded from the Meat Standards Australia (MSA) grading system (Ferguson et al. 1999).
Whilst the association between preslaughter stress and muscle glycogen depletion has been extensively studied in ruminants, the same cannot be said for the association between stress preslaughter and postmortem glycolytic rate. This is in contrast with the large body of recent research in pigs (e.g. Klont and Lambooy 1995; Warriss et al. 1995; Channon et al. 2000; Stoier et al. 2001). The general finding across these pig studies was that stress just before stunning resulted in lower initial muscle pH, higher initial muscle temperature and a faster rate of pH decline in the first hour after death, although similar rates were observed beyond that. There is a paucity of similar data in ruminants, but it is highly likely that acute stress preslaughter may alter the rate of postmortem pH fall and thus change the carcass response to electrical stimulation. Electrical stimulation is used to ensure that the optimum muscle pH–temperature window, as defined within the MSA grading system (Ferguson et al. 1999), is achieved. It is known that, within a consignment of cattle, the rate of pH fall in the longissimus can vary widely (O’Halloran et al. 1997). Consequently, this can affect the proportion of carcasses that achieve the optimum pH–temperature window. Furthermore, if electrical stimulation is applied to the carcass of an animal that has undergone acute stress preslaughter, the resulting rate of pH fall may be so rapid that the muscle proteins denature and produce pale, watery beef. Pale, watery meat has a reduced ability to tenderise during aging (pork; Channon et al. 2000) and has high water losses (beef; purge in vacuum bags) (Tarrant and Mothersill 1977), which potentially compromises juiciness of the product.
The aim of this experiment was to investigate the effects of acute preslaughter stress in beef cattle on postmortem muscle pH and temperature fall and beef eating quality.
Materials and methods
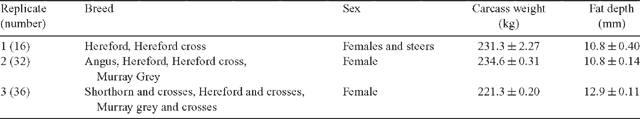
A total of 84 feedlot cattle destined for the domestic market were used in this study. The cattle were slaughtered in three groups (n = 16, 32 and 36) on three separate days. The cattle in the third slaughter group were subjected to an on-farm measurement of flight timespeed test (Burrow et al. 1988). On each slaughter day, the cattle were split into groups of four and randomly allocated to a ‘control’ or ‘stress’ treatment. The breeds, sex and average carcass weight and fat depth of each replicate (i.e. slaughter group) are described in Table 1. All animals had either zero or two permanent incisors and were all finished in a feedlot for 80–120 days before dispatch for slaughter. For replicates 1 and 2, cattle were selected from a larger mob on the day of slaughter and randomly allocated to a treatment subgroup. Whereas for replicate 3, cattle were weighed at the feedlot, given a flight timespeed test and randomly allocated to treatment within ‘low’ and ‘high’ flight speed blocks. Cattle in replicates 1 and 2 were trucked for 4 h from feedlot to abattoir and cattle in replicate 3 had a 7 h transport to the abattoir. All cattle arrived at the abattoir on the day before slaughter. The cattle were held overnight in their groups in lairage pens with access to water.
|
The two preslaughter treatments were applied at 15 min preslaughter and are described below.
-
Control (i.e. no acute preslaughter stress applied). Each animal was quietly moved from the lairage pen to the knocking box with a minimum of coercion. No electric prodders or other forcing implements were used. These cattle had no visual, auditory or physical contact with the ‘stress’ treatment cattle during application of prodding.
-
Stress treatment. Groups of four cattle entered the circular forcing pen before the race leading to the knocking box. An electric cattle prodder (Hot Shot HS 2000, 4 × 1.5 V batteries; Savage, Minnesota, distributed in Australia by Leader products Pty Ltd) was used to elicit an acute stress response in the animals. Six electric shocks were given over 5–10 min to each animal in the groups.
After application of the stress and control treatment, cattle were then moved into the knocking box, stunned using a captive bolt pistol and exsanguinated. Blood samples were collected from the jugular during exsanguination into heparinised collection tubes and immediately placed on ice. The samples were centrifuged within 30 min of collection at 1500g for 20 min. The plasma was aliquoted, frozen and stored at –20°C until analysis of plasma lactate.
AUS-MEAT carcass measurements including hot standard carcass weight, P8 fat depth and dentition were recorded. Samples of the M. longissimus thoracis (LT) from between the 12th and 13th rib were removed at 60 min postslaughter. The samples were trimmed of fat and connective tissue, immediately frozen in liquid nitrogen and stored at –80°C until analysis of lactate concentration. The pH and temperature of the LT muscle was measured in duplicate between the 10th and 11th ribs at 1 h postslaughter and every hour until 5 h postslaughter. Meat pH and temperature was measured using a Micrometer pH Vision Model 6007 (Jenco Instruments, San Diego, CA) with a direct pH probe (Ionode Model No IJ42).
At 2 days postslaughter, the striploins (including longissimus lumborum, LL) were removed from both sides of the carcass, vacuum packed and stored at 0–2°C. The striploin from the left side was despatched for MSA consumer panel assessment. The LL from the right side was used for meat quality and protein biochemistry assessment on fresh and aged samples. The striploins from the right side were cut into three portions and the portions randomly allocated to 0, 4 or 19 days aging in a vacuum bag (subsequently called 2-, 6- or 21-day postslaughter samples) at 2°C. Samples were removed from the 2-day postslaughter portion for measurement of Warner–Bratzler peak shear force (WBPSF), sarcomere length, cooking loss, drip loss, total water content, ultimate pH and surface colour. The aged samples were removed from packs at 6 and 21 days postslaughter and purge, WBPSF, cooking loss and surface colour were measured.
Consumer panel
Detailed descriptions of the development and methodology of the sensory evaluation protocol are provided by Polkinghorne et al. (1999). Briefly, at the completion of the 19 days of aging, the striploin samples were cut into five 25-mm steaks, allocated a unique code number and frozen and stored at –20°C. Steaks were thawed (2–5°C) for 24 h before cooking on a Silex griller. Steaks were cooked to an internal temperature of 70°C, halved and allocated to panellists. Untrained panellists who prefer meat cooked to a medium degree of doneness and who consume beef at least once each week were used in the study. Consumers were allocated seven half steaks and were asked to score tenderness, juiciness, flavour and overall liking on a 100-mm line scale (higher = more acceptable). These scores (1–100) were weighted to derive the overall acceptability of each steak, which was defined as the MQ4 score. Two panellists evaluated each steak. This meant that 10 consumers assessed the five steaks.
Plasma and muscle metabolites
Plasma lactate concentrations were measured using a commercially available kit as per the manufacturer’s instructions (catalogue 735-10, Sigma-Aldrich, USA). Plasma (3 μL), lactate standard and lactate reagent (300 μL) were dispensed to microtitre plates. The A540 of a chromophore was measured following a 10 min incubation period at room temperature (~22°C) and concentration was expressed as mol/L. Lactate in muscle samples was determined as described by (Noll 1985). Muscle lactate concentration was expressed as μmol/g tissue (wet weight).
Meat quality measurements
The surface colour was measured after a 30 min ‘bloom’ period at 2°C using a Minolta chromameter (Minolta CR-200, Tokyo, Japan) with a 2° standard observer, D65 lighting and an 8 mm aperture in the measuring head. Colour measurements were reported in the CIE L*, a*, b* system (where L* measures relative lightness, a* relative redness and b* relative yellowness). Three replicate measurements were taken across the same cross section of muscle, avoiding areas of connective tissue or intramuscular fat and the average reading recorded.
Drip loss was measured in duplicate samples (~80 g) attached to plastic netting and suspended in an inflated plastic bag for 48 h at 2°C (Honikel et al. 1986). Drip loss was the difference in weight between pre- and postsuspension and was expressed as a percentage of the initial weight. Meat purge was measured as the percent weight loss in the vacuum packed samples after 6 and 21 days aging at 2°C.
Samples (~65 g) for WBPSF and cooking loss were cooked for 30 min at 70°C, followed by cooling in running water for 30 min and refrigerating for 24 h at 2°C (Moller and Vestergaard 1986). Cooking loss was the difference in weight between the precooked weight and blotted dry postcooked weight and it was expressed as a percentage of the precooked weight. The WBPSF was measured on five 1-cm2 segments of meat cut parallel to the muscle fibres and a minimum of 5 cm long. Shear force was measured by an Instron texture analyser fitted with an inverted V-blade and cross head speed of 300 mm/min perpendicular to fibre orientation (Bouton et al. 1971).
Sarcomere length was determined using a helium–neon laser diffraction unit (custom built by the University of New England, Armidale, Australia), similar to that described by Ruddick and Richards (1975). Frozen muscle samples were cut parallel with the fibres into 4 cm long × 1 cm2 cross sectional area segments, frozen in liquid nitrogen and stored at –20°C. A thin slice of frozen muscle was placed between two microscope slides, thawed and squeezed flat before measuring the distance of light band diffraction. Sarcomere length was calculated as:
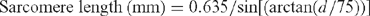
Statistics
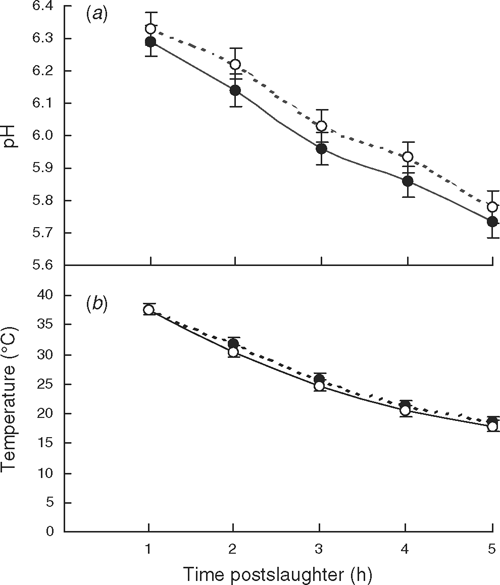
The data was analysed as a paired comparison with a stressed and nonstressed group of 7–10 cattle in each pair. For each measurement, the raw data for analysis was the mean of all cattle in the group. To formally compare the pH and temperature decline with time between treatments, the slope of the linear regression for pH and temperature with time were calculated for all pH and temperature measurements until 5 h postslaughter. Over this time period, the lines were linear (see Fig. 1a, b) and did not show evidence of needing an exponential fit. Temperature at pH 6.0 was calculated for individual carcasses by linearly interpolating the temperature with respect to pH between the first time pH reached a value less than 6.0 and the previous measurement occasion. In cases where the pH was already below pH 6.0 at 1 h postslaughter (only 7 carcasses out of 84), the muscle pH and temperature at time 0 (slaughter) were assumed to be 7.0 and 40°C respectively. The loin pH is likely to be lower than 7.0 at slaughter in carcasses with a low pH at 1 h postslaughter, but this cannot be assumed. Thus, we considered it was safer to use pH 7.0 as the standard.
|
Results
There was no difference (P > 0.05) in flight timespeed between the two treatments for replicate 3 and also no correlations between flight timespeed and any meat quality measures (P > 0.05). Thus, these results are not discussed any further.
Plasma and muscle metabolites
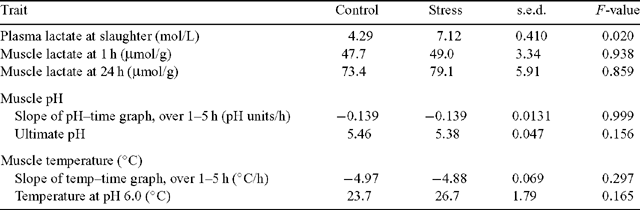
Table 2 shows the effect of the stress treatment on plasma and muscle metabolites. The stress treatment has invoked an acute adrenergic stress response, as indicated by higher (P < 0.05) plasma lactate levels at slaughter. The stress treatment had no effect on LT lactate concentration at either 1 h or 24 h postslaughter (P > 0.05 for both).
|
pH and temperature fall
The postslaughter pH and temperature decline of the LT was not different between treatments (P > 0.05), as indicated by the slopes of the pH–time and temperature–time linear regressions, the temperature at rigor (pH 6) (Table 2) and the pH and temperature of the LT at each time point (Fig. 1a, b). The ultimate pH in the LT was also not different between treatments (P > 0.05). There were seven carcasses where the LT pH was below 6.0 at 1 h postslaughter. Five of these were in the ‘stress’ treatment and two were in the control treatment. Seven carcasses had a pH > 5.7 in the LL at 48 h postslaughter (i.e. dark-cutting) and these were all in the control treatment within one group of cattle.
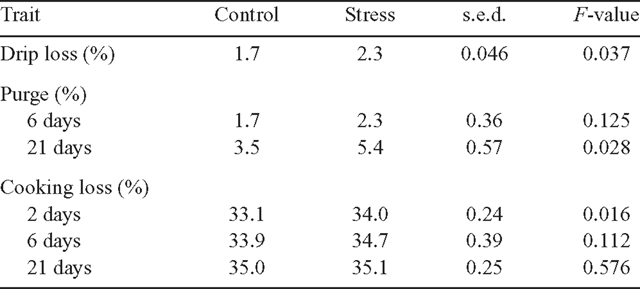
Water-holding capacity traits
Table 3 presents the results of the effect of applying an acute stress preslaughter on the water-holding capacity traits of the meat. Drip loss from the meat after suspension for 2 days at 2°C was higher in the LL from cattle undergoing acute preslaughter stress (P < 0.05). In addition, purge after 21 days of aging and cooking loss at 2 days postslaughter were both higher for ‘stressed’ cattle (P < 0.05 for both).
|
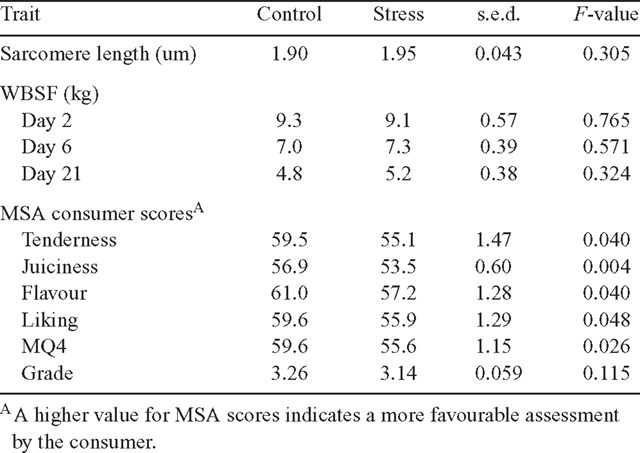
Objective tenderness, sarcomere length and MSA consumer panel evaluation
Sarcomere length and WBPSF were not influenced by stress at any of the time points measured (P > 0.05 for all; Table 4).
|
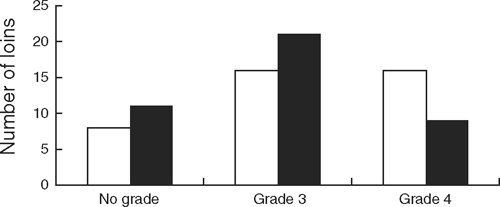
Meat from the carcasses of cattle undergoing acute stress preslaughter was assessed by untrained consumers as being less tender, less juicy with a less acceptable flavour and had lower ‘liking’ and MQ4 scores compared to meat from cattle not subjected to stress (P < 0.05 for all; Table 4). The acute stress treatment shifted the distribution of the consumer assessments of the quality of the striploins towards the lower grade (Grade 3) or to a ‘no grade’ (Fig. 2) relative to the controls.
|
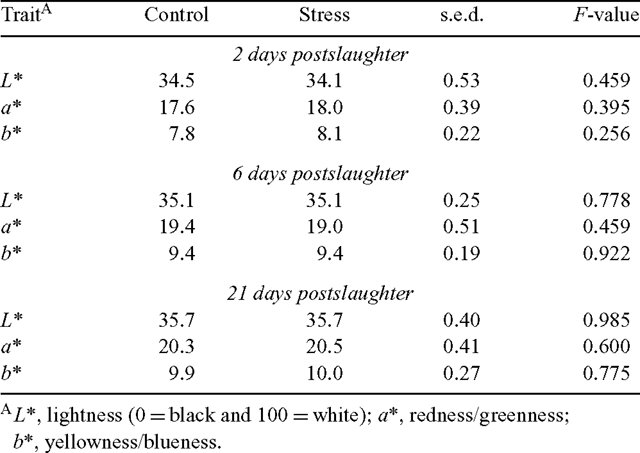
Surface colour
Table 5 presents the effects of the acute preslaughter stress treatment on the surface colour of the LL. The acute stress treatment did not influence the L* (lightness), a* (redness/greenness) or b* (yellowness/blueness) values after 2, 6 or 21 days of aging (P > 0.05 for all). These results, combined with the ultimate pH values presented above, are evidence that the ‘stress’ treatment had no effect on the incidence of dark-cutting in the striploin.
|
Discussion
Subjecting cattle to the acute stress of being electrically prodded just before slaughter resulted in a significant reduction in consumer acceptability (lower tenderness, juiciness, flavour and overall liking scores) of loin steaks. The higher plasma lactate levels at slaughter in the acutely stressed cattle is indicative of sympatho-adrenal stress response, where the stress-mediated increase in the release of catecholamines causes an acceleration in glycogenolysis (Shaw and Tume 1992). The expectation was that the catecholamine released preslaughter during the acute stress would result in an increase in muscle pH fall postslaughter, as well as an increase in muscle temperature. Pigs subjected to acute stress immediately preslaughter, through the use of electric prodders, gave rise to elevated postslaughter muscle temperatures and reduced loin pH (D’Souza et al. 1998a, 1998b; Channon et al. 2000). In stark contrast, similar treatments in cattle in this study elicited no significant change in muscle pH or temperature postslaughter and also no change in ultimate pH.
Plasma lactate appeared to be a better predictor of final consumer eating quality than muscle pH fall postmortem. Catecholamine release, as a result of fear and excitement, can lead to rapid glycolysis and excessive lactate production and hence elevated blood lactate (Shaw and Tume 1992). Generally lactate at slaughter does not predict the dark-cutting meat quality defect, as dark-cutting is a consequence of chronic stress from farm to slaughter (Warriss et al. 1984), whereas plasma lactate indicates ‘acute stress’. In pigs, elevated plasma lactate at slaughter as a result of stress preslaughter has been correlated with a rise in muscle temperature and in ultimate pH in the loin postslaughter (Brown et al. 1998). Negative handling of pigs preslaughter, using prodders, results in higher muscle lactate in loin and leg muscles but not always reduced muscle pH (D’Souza et al. 1998a, 1998b, 1999). Thus, a rise in plasma lactate at slaughter can be used as a guide to stress levels at slaughter due to adrenergic activation and it may be a better predictor of final meat quality than muscle pH.
Cattle subjected to electric prodding every 20 min for 24 h preslaughter have previously been shown to exhibit reduced objective (WBSF) and consumer panel-assessed tenderness in the longissimus, independent of ultimate pH (Lewis et al. 1977). Their prodding regime was excessive and experienced over a sustained period of time, whereas the prodding we imposed was designed to produce an acute stress response. Moreover, the treatment we applied could potentially occur in beef abattoirs, particularly those with poorly designed lairage handling facilities. A toughening effect of stress, independent of ultimate pH, has been observed in lambs (Daly et al. 1995; Bickerstaffe et al. 1996). However, the stress treatment involved extensive ‘swim washing’ over a period and, thus, extensive muscular use.
Meat tenderness is principally determined by the interaction between the degree of myofibrillar shortening and the extent of myofibrillar degradation postmortem. The latter is largely dependent on the activation or suppression of the calpain enzyme system (Dransfeld 1994; Koohmaraie 1994). The calpains (m-calpain and μ-calpain) are inhibited by the endogenous enzyme calpastatin, and Sensky et al. (1996) suggested that tight regulation of calpain is achieved through regulation of calpastatin. Sensky et al. (1996) further demonstrated that epinephrine infusion into pigs for 1 week preslaughter caused an elevation in calpastatin activity in the LT postmortem, with no effect on calpain activity. As calpastatin is the inhibitor of the enzyme calpain, the logical outcome of this effect would be suppression of tenderness and tenderisation postmortem, although they did not measure this. In our case, we did not measure calpain activity but, as there were no effects of the stress treatment on WBPSF, this tends to suggest that the stress effects were not mediated by calpains.
The reduced acceptability of the loins from ‘stress’-treated animals may have been driven by the observed differences in water-holding capacity. Consumer assessments of quality traits are rarely independent. Thus, it is unlikely that the consumer can rate a piece of meat ‘tender’ if it is also ‘dry’. The water-holding capacity of the meat from ‘stress’ treated animals was clearly reduced as the drip loss, purge and cooking loss were increased, even though the rate of pH decline and the ultimate pH were not affected. This would most likely explain the reduced juiciness in the ‘stress’ treated animals. The reduced water-holding capacity could be underpinning the differences in sensory attributes. The question is why the water-holding capacity was reduced by the ‘stress’ treatment?
Reduced water-holding capacity in meat postslaughter is usually attributed to fast pH fall, combined with high muscle temperatures. These conditions are known to result in protein denaturation, particularly of some sarcoplasmic proteins and of the myosin head (Warner et al. 1997), and low ultimate pH (Warner et al. 1997). Together, these cause shrinkage in the myofibrillar lattice and an increase in extracellular water (Offer 1991). Preslaughter stress has been clearly shown to reduce the water-holding capacity of meat in pig carcasses (Tarrant 1989b for review), due to the accelerated rate of pH decline postmortem, and is associated with the occurrence of the quality defect PSE (pale, soft, exudative). Lambs undergoing preslaughter exercise stress during 15 min preslaughter have been reported to have increased drip loss, cooking loss and purge in both loin and leg muscles (Warner et al. 2000; Bond et al. 2004) and this was associated with an accelerated rate of pH decline postmortem. Cattle subjected to electric shocks every 20 min for 24 h preslaughter induced increased cooking loss and total moisture loss in the psoas major and quadriceps femoris, with only a slight elevation in ultimate pH of 0.1 pH units. In our case, the reduction in water-holding capacity due to stress occurred in the total absence of any significant effect on pH or temperature postslaughter. A possible explanation for the loss in water-holding capacity is that the ‘stress’ treatment caused a shift in the distribution of ions in the muscles, with ions passing into the plasma preslaughter. This would decreased the osmolarity of the fluid in the muscle at slaughter, as described by Warner et al. (2001), Bond et al. (2004) and Bond and Warner (2007) in lamb muscle in response to exercise stress. This diffusion of ions out of the muscles with ‘stress’ continues postslaughter, with more ions appearing in the fluid expressed from meat during storage (Bond and Warner 2007). Release and diffusion of free ions across the sarcolemma contribute to the degree of swelling or shrinking of the myofibrillar lattice and hence the space available to hold water.
The reduction in flavour acceptability observed with the ‘stress’ treatment is interesting. Jeremiah et al. (1988) reported that cattle undergoing normal stress preslaughter (mixing, 160 km trucking, 24 h in lairage) exhibited reduced flavour desirability scores relative to cattle undergoing minimal stress (shipped and slaughtered within 4 h of leaving feedlot). They attributed the differences to a higher ultimate pH in the ‘normal stress’ group. When stress causes muscle glycogen loss preslaughter and results in meat with high ultimate pH, it is common to have reports of altered flavour including ‘less desirable flavour’ in beef (Lewis et al. 1977; Wulf et al. 2002), although other studies in lamb have shown no correlation between ultimate pH and flavour (Devine et al. 1993). Jeremiah et al. (1988) reported that the level of preslaughter stress applied to cattle significantly influenced the detection of specific flavour character notes, with minimal preslaughter stress receiving higher flavour desirability scores. Higher levels of preslaughter stress resulted in inappropriate character notes being more intense. Jeremiah et al. (1988) attributed the differences to higher levels of dark-cutting in animals with more stress but, as they did not measure the ultimate pH of the meat samples, this could not be confirmed. We have previously found that exercise stress in the last 15 min preslaughter reduces the concentration of free amino acids in the meat during storage (L. Can and R. Warner, unpubl. data). As flavour is derived partly from amino acids, it is possible that the reduction in flavour acceptability with the ‘stress’ treatment is due to an inhibition of the increase in free amino acids that normally occurs in meat during aging. However, it has to be reiterated that given that an untrained consumer panel was used, the perceived differences in flavour may not be real; rather they may have been underpinned by the sensory differences in juiciness.
Conclusions
Cattle undergoing acute stress immediately preslaughter produced meat of reduced consumer acceptability and lower water-holding capacity. The mechanism for the reduced quality of the loins from cattle undergoing preslaughter stress was not clear, as the factors known to influence tenderness (ultimate pH, rate of pH fall, muscle temperature, sarcomere length) were not affected by the preslaughter stress treatment. The reduced consumer acceptability is most likely associated with the lower juiciness of the acutely stressed cattle, which is driven by the reduced water-holding capacity of the muscle. There would be a reasonable number of cattle that exhibit symptoms of acute stress preslaughter and this may explain observed differences in tenderness between vendor groups, days of slaughter, time of day and abattoir. The stress-induced changes in beef quality identified in this experiment cannot currently be accounted for in a model for predicting eating quality, such as that used by MSA. This is clearly an area for further investigation.
Acknowledgements
This research was funded by the Cooperative Research Centre for Beef, Meat Standards Australia and Meat and Livestock Australia. We thank Matthew Kerr, Peter Walker and Paul Weston for their technical assistance. The biometrical input of Kym Butler is gratefully acknowledged. The study could not have occurred without the cattle provided by Coles Supermarkets through Charlton Feedlot and Kaladbro feedlot and the assistance provided by staff at G and K O’Connor. In particular, the assistance of John Skelton of Coles Supermarkets, Australia, David McKibbin of Coles Supermarkets, Australia and Hamish MacKinnon of Kaladbro is appreciated. We also thank Lotte Nielsen, a student from the Swedish Agricultural University in Uppsala, Sweden, for her input into one of the replicates for the study.
Bond JJ, Warner RD
(2007) Effects of antemortem exercise on water holding capacity of lamb Longissimus thoracis et lumborum: ion distribution and protein proteolysis post mortem. Meat Science 75, 406–414.
| Crossref | GoogleScholarGoogle Scholar |

Bond JJ,
Can LA, Warner RD
(2004) The effect of exercise stress, adrenaline injection and electrical stimulation on changes in quality attributes and proteins in Semimembranosus muscle of lamb. Meat Science 68, 469–477.
| Crossref | GoogleScholarGoogle Scholar |
Bouton PE,
Harris PV, Shorthose WR
(1971) Effect of ultimate pH upon the water-holding capacity and tenderness of mutton. Journal of Food Science 37, 358–360.
Brown SN,
Warriss PD,
Nute GR,
Edwards JE, Knowles TG
(1998) Meat quality in pigs subjected to minimal preslaughter stress. Meat Science 49, 257–265.
| Crossref | GoogleScholarGoogle Scholar |
Burrow HM,
Seifert GW, Corbet NJ
(1988) A new technique for measuring temperament in cattle. Proceedings of Australian Society of Animal Production 17, 154–157.
Channon HA,
Payne A, Warner RD
(2000) Halothane genotype, preslaughter handling and stunning method all influence pork quality. Meat Science 56, 291–299.
| Crossref | GoogleScholarGoogle Scholar |
Devine CE,
Graafhuis AE,
Muir PD, Chrystall BB
(1993) The effect of growth rate and ultimate pH on meat quality of lambs. Meat Science 35, 63–77.
| Crossref | GoogleScholarGoogle Scholar |
Dransfeld E
(1994) Modelling post mortem tenderisation. V. Inactivation of calpains. Meat Science 37, 391–409.
| Crossref | GoogleScholarGoogle Scholar |
D’Souza DN,
Warner RD,
Leury BJ, Dunshea FR
(1998a) The effect of dietary magnesium aspartate supplementation on pork quality. Journal of Animal Science 76, 104–109.
| PubMed |
D’Souza DN,
Dunshea FR,
Warner RD, Leury BJ
(1998b) The effect of handling pre-slaughter and carcass processing rate post-slaughter on pork quality. Meat Science 50, 429–437.
| Crossref | GoogleScholarGoogle Scholar |
D’Souza DN,
Dunshea FR,
Leury BJ, Warner RD
(1999) Effect of mixing boars during lairage and preslaughter handling on pork quality. Australian Journal of Agricultural Research 50, 109–113.
| Crossref | GoogleScholarGoogle Scholar |
Honikel KO,
Kim CJ, Hamm R
(1986) Sarcomere shortening of prerigor muscles and its influence on drip loss. Meat Science 16, 267–282.
| Crossref | GoogleScholarGoogle Scholar |
Jeremiah LE,
Newman JA,
Tong AKW, Gibson LL
(1988) The effects of castration, preslaughter stress and zeranol implants on beef. 2. Cooking properties and flavor of loin steaks from bovine males. Meat Science 22, 103–121.
| Crossref | GoogleScholarGoogle Scholar |
Klont RE, Lambooy E
(1995) Effects of preslaughter muscle exercise on muscle metabolism and meat quality studied in anesthetized pigs of different halothane genotypes. Journal of Animal Science 73, 108–117.
| PubMed |
Koohmaraie M
(1994) Muscle proteinases and meat ageing. Meat Science 36, 93–104.
| Crossref | GoogleScholarGoogle Scholar |
Lewis PK,
Brown CJ, Heck MC
(1977) Effect of pre slaughter treatments on certain chemical and physical characterstics of certain beef muscles. Journal of Animal Science 20, 433–438.
Moller AJ, Vestergaard T
(1986) Effects of altered carcass suspension during rigor mortis on tenderness of pork loin. Meat Science 18, 77–87.
| Crossref | GoogleScholarGoogle Scholar |
Offer G
(1991) Modelling of the formation of pale, soft and exudative meat: Effects of chilling regime and rate and extent of glycolysis. Meat Science 30, 157–184.
| Crossref |
O’Halloran GR,
Troy DJ, Buckley DJ
(1997) The relationship between early postmortem pH and the tenderisation of beef muscles. Meat Science 45, 239–251.
| Crossref | GoogleScholarGoogle Scholar |
Ruddick JE, Richards JF
(1975) Comparison of sarcomere length measurement of cooked chicken pectoralis muscle by laser diffraction and oil immersion microscopy. Journal of Food Science 40, 500–501.
| Crossref | GoogleScholarGoogle Scholar |
Sensky PL,
Parr T,
Bardsley RG, Buttery PJ
(1996) The relationship between plasma epinephrine concentration and the activity of the calpain enzyme system in porcine longissimus muscle. Journal of Animal Science 74, 380–387.
| PubMed |
Shaw FD, Tume RK
(1992) The assessment of preslaughter and slaughter treatments of livestock by measurement of plasma constituents – a review of recent work. Meat Science 32, 311–329.
| Crossref | GoogleScholarGoogle Scholar |
Stoier S,
Aaslyng MD,
Olsen EV, Henckel P
(2001) The effect of stress during lairage and stunning on muscle metabolism and drip loss in Danish pork. Meat Science 59, 127–131.
| Crossref | GoogleScholarGoogle Scholar |
Tarrant PV
(1989a) Animal behaviour and environment in the dark-cutting condition in beef – a review. Irish Journal of Food Science and Technology 13, 1–21.
Tarrant PV
(1989b) The effect of handling, transport, slaughter and chilling on meat quality and yield of pigs. Manipulating Pig Production 3, 1–25.
Tarrant PV, Mothersill C
(1977) Glycolysis and associated changes in beef carcasses. Journal of the Science of Food and Agriculture 28, 739–749.
| Crossref | GoogleScholarGoogle Scholar |
Warner RD,
Kauffman RG, Greaser ML
(1997) Muscle protein changes post mortem in relation to pork quality traits. Meat Science 45(3), 339–352.
| Crossref |
Warner RD,
Channon HA,
Hofmeyr CD,
Can AL,
Cottrell JJ,
Bond JJ,
Greaser ML, Kauffman RG
(2001) Water-holding capacity (WHC) of pork: preslaughter stress and protein denaturation. Proceedings of the Annual Reciprocal Meat Conference 54, 148–154.
Warriss PD
(1990) The handling of cattle preslaughter and its effects on carcass and meat quality. Applied Animal Behaviour Science 28, 171–186.
| Crossref | GoogleScholarGoogle Scholar |
Warriss PD,
Kerstin SC,
Brown SN, Wilkins LJ
(1984) The time required for recovery from mixing stress in young bulls and the prevention of dark cutting meat. Meat Science 10, 53–68.
| Crossref | GoogleScholarGoogle Scholar |
Warriss PD,
Brown SN,
Nute GR,
Knowles TG,
Edwards JE,
Perry AM, Johnson SP
(1995) Potential interactions between the effects of preslaughter stress and postmortem electrical stimulation of the carcasses on meat quality in pigs. Meat Science 41, 55–68.
| Crossref | GoogleScholarGoogle Scholar |
Wulf DM,
Emnett RS,
Leheska JM, Moeller SJ
(2002) Relationships among glycolytic potential, dark cutting (dark, firm, and dry) beef, and cooked beef palatability. Journal of Animal Science 80, 1895–1903.
| PubMed |