

Increasing the proportion of Leucaena leucocephala in hay-fed beef steers reduces methane yield
A. Stifkens A B , E. M. Matthews A , C. S. McSweeney C and E. Charmley A *
A *
A CSIRO Agriculture and Food, Private Mail Bag PO Aitkenvale, Townsville, Qld 4814, Australia.
B University of Liège, Gembloux Agro-Bio Tech, 2 Passage des Déportés, 5030 Gembloux, Belgium.
C CSIRO Agriculture and Food, 306 Carmody Road, St Lucia, Qld 4067, Australia.
Animal Production Science 62(7) 622-632 https://doi.org/10.1071/AN21576
Submitted: 19 November 2021 Accepted: 21 January 2022 Published: 22 February 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Leucaena leucocephala (leucaena) is a leguminous shrub adapted to higher rainfall (>600 mm) in frost-free areas of Australia. It can be a source of high-quality forage for cattle grazing tropical grass-based pastures that are seasonally deficient in the nitrogen content required for adequate levels of performance. Leucaena contains bioactive compounds that may reduce methanogenesis in the rumen, helping to achieve Australia's goal to make red meat production carbon neutral by 2030.
Aim: A study was undertaken to evaluate the response in animal performance and methane production to increasing percentages of leucaena in a hay-based diet.
Methods: Growing steers were fed diets containing 0%, 18%, 36% and 48% leucaena. Intake, liveweight gain, methane production and yield were measured in a cross-over trial with two modern cultivars of leucaena (Redlands and Wondergraze). Methane was measured in open-circuit respiration chambers.
Key results: There were no effects of cultivar on most parameters. Increasing leucaena percentage in the diet increased dry matter intake, animal performance and methane production (g/day) but reduced methane yield (g/kg dry matter intake) according to the equation: methane yield = 19.8 − 0.09 × leucaena percentage in the diet. The inclusion of polyethylene glycol to nullify potential antimethanogenic activity of tannins restored methane yield by 67%, indicating that tannins were responsible for most of the observed reduction in methane yield.
Conclusion: The results demonstrate that leucaena can improve animal performance and reduce methane yield in steers fed low-quality grasses.
Implications: Leucaena can be included in diets of grazing cattle in areas agronomically suited to its production, as a means to reduce enteric methane emissions.
Keywords: Australia, cattle, greenhouse gas, intake, legume, methane, tannin, tropical.
Introduction
In Australia, methane emissions from ruminant livestock are responsible for ∼69% of total agricultural emissions of greenhouse gases (total was 69.7 Mt CO2-e in 2019) and 9.2% of the total national emissions (Australian Greenhouse Emissions Information System 2019). The Australian red meat industry has a goal of carbon neutrality by 2030 (Meat & Livestock Australia, 2021). Leucaena leucocephala (leucaena) is a leguminous shrub adapted to higher rainfall (>600 mm), frost-free areas of Australia. It can provide high-quality forage for cattle grazing tropical grass-based pastures (Tomkins et al. 2019). These C4 grasses are typically deficient in nutrient content for adequate levels of performance, particularly in the dry season (Poppi and McLennan 2010). Shelton and Dalzell (2007) estimate that ∼13.5 Mha of land is suitable for planting leucaena in the state of Queensland, which represents 9.2% of the grazing area in the state. Beutel et al. (2018) estimate that, in 2018, 123 500 ha of pastures contained leucaena. Leucaena has a deep and well-developed taproot, facilitating rapid growth and access to water and nutrient reserves in lower soil horizons than can be reached by grasses. As a result, leucaena is a productive, high-quality forage, typically having crude protein (CP) content of 15–25% (Shelton and Brewbaker 1998; Shelton and Dalzell 2007; Radrizzani et al. 2011). Bowen et al. (2016) compared six forage types in central Queensland and showed annual liveweight (LW) gain to be 198 kg/ha for grass–leucaena pastures, which exceeded LW gain from other forages in the study, including oats (Avena sativa), sorghum (Sorghum spp.), lablab (Lablab purpureus) and mixtures of C4 grass species–butterfly pea (Clitoria ternatea). Harrison et al. (2015) presented data comparing cattle grazing Rhodes grass pastures with and without leucaena and showed a 50% increase in LW gain over 14 months when leucaena was included in the pasture.
Antimethanogenic properties of leucaena have been demonstrated in vivo with cattle and sheep. Kennedy and Charmley (2012) reported an 18% methane abatement when leucaena was fed to cattle at 44% of the diet. Soltan et al. (2013) included leucaena at 35% in sheep diets and achieved a 14% reduction in methane emissions. Archimède et al. (2016) found that leucaena pellets included at 44% of a Dichanthium diet reduced methane production in sheep by 40%.
Tannins have been implicated in reducing methane emissions in ruminants (Jayanegara et al. 2012; Aboagye and Beauchemin 2019 (Aboagye and Beauchemin 2019). However, the relationship between tannins and methane reduction is unclear and results can vary among studies (Aboagye and Beauchemin 2019. The concentration and molecular structure of tannins vary widely, with the major differentiation being between condensed tannins and hydrolysable tannins, the latter being more prevalent in tropical forages (Aboagye and Beauchemin 2019). Condensed tannins are often considered responsible for methane abatement in leucaena. Tannins could lower methane production by directly inhibiting methanogens and H2-producing microorganisms, and/or indirectly by reducing feed degradation in the rumen (Aboagye and Beauchemin 2019). The concentration of condensed tannins in leucaena is variable depending on a range of factors including subspecies and cultivar (Jones and Palmer 2002). Data for L. leucocephala show condensed tannin concentration ranging from 5 to 75 g/kg dry matter (DM) (Jones and Palmer 2002; Tan et al. 2011; Soltan et al. 2013; Archimède et al. 2016; Montoya-Flores et al. 2020).
The objectives of this study were to establish a relationship between the proportion of leucaena in the diet and methane production and feed intake, to determine whether there was a cultivar effect, and to demonstrate that inclusion of polyethylene glycol (PEG) in the diet reversed the suppressive effect of tannins on methane production.
Materials and methods
The experiment was conducted at CSIRO’s Lansdown Research Station (19°39′S, 146°50′E), 45 km south of Townsville, north-eastern Australia, from 18 March until 28 June 2019, with a further study when PEG was included in the diet the following year from 13 June to 19 July 2020. Initially, it was planned that the inclusion of PEG to the diet would follow on from the main dose-response study. However, by mid-June the leucaena was exhibiting leaf loss and there was insufficient biomass available for the PEG study. The experimental protocols complied with the Australian code for the care and use of animals for scientific purposes (8th Edition 2013) and was approved by the CSIRO Queensland Animal Ethics Committee (AEC Numbers: 2019-02 and 2019-32).
Two cultivars of leucaena (Redlands and Wondergraze) were established at Lansdown Research Station in March–April 2017. The tropical legumes were sown into an existing grass pasture of 20 ha of mixed C4 species dominated by Indian couch (Bothriochloa pertusa), Queensland bluegrass (Dichanthium sericeum), black speargrass (Heteropogon contortus) and sabi grass (Urochloa mosambicensis). Leucaena was established in February 2017 in twin rows spaced 12 m apart. Strips to be established with leucaena were sprayed with glyphosate (Roundup; Bayer, Leverkusen, Germany) and disced to provide a tilth suitable for seeding. The paddock was grazed for 3 weeks in April–May 2018 to remove excess biomass, and the leucaena rows were slashed to 30 cm above ground level in July 2018 to promote branching. Then, based on a soil report identifying a phosphorus deficiency (7 μg/g), the paddock was fertilised in September 2018 with 250 kg/ha of single superphosphate and 150 kg/ha of muriate of potash. Following these operations and favoured by an exceptional wet season (∼1500 mm rainfall in January and February 2019), the leucaena plantation reached sufficient biomass to initiate the experiment in March 2019. Grasses in the sward were mowed in April 2019 to make the leucaena rows more accessible for harvesting.
Leucaena was hand harvested as stems cut at 1 m above ground level. Stems were transferred to a feed preparation area where green material (leaves and green stems <10 mm diameter) was removed by hand and shredded with a garden chopper (Greenfield herbage mulcher, Model GT03300; Cox Industries, Acadia Ridge, Qld, Australia). Thicker stems and mature pods were discarded. The leucaena was harvested and prepared 3 days per week (on Mondays, Wednesdays and Fridays) and stored in a cold room at <5°C until feeding.
Experimental design and animals
Sixteen Droughtmaster (50% Bos indicus, 50% Bos taurus) steers with an initial LW of 428 ± 25 kg (mean ± s.e.) were used for the main experiment. After 4 months of grazing, they were housed in individual covered pens (3 m by 4 m) and allocated equally to four groups, based on methane emissions, recorded in a previous experiment.
Prior to introducing treatments, a 4-week period was used to familiarise the cattle with handling and with entry and exit to the methane chambers, and for adaptation to Rhodes grass (Chloris gayana) hay. During this time, individual methane emissions were measured in open-circuit respiration chambers, creating a baseline methane production for each animal fed with 100% Rhodes grass at 90% of ad libitum. There followed a 2-week adaptation period, during which two leucaena cultivars were gradually introduced into the diet (cvv. Redlands and Wondergraze; eight steers per cultivar). On Day 7 of the adaptation period, each animal was dosed with an inoculum (100 mL) of a bacterial suspension of Synergistes jonesii as recommended by Business Queensland (Leucaena inoculum for cattle, Business Queensland: https://www.business.qld.gov.au/industries/farms-fishing-forestry/agriculture/livestock/cattle/leucaena-inoculum-cattle). After 14 days, leucaena inclusion levels of 0%, 18%, 36% and 48% were achieved, with steers remaining on these levels throughout four periods, each of 14 days. Ad libitum intake (10% refusals) was established within the first 7 days of each period; thereafter, intake was reduced to 90% of ad libitum to minimise variation in feed intake during methane measurements. After the first two periods, steers fed Wondergraze were switched to Redlands and vice versa with one extra week included in the design to adapt to the change in cultivars. Individual methane emissions were measured in open-circuit respiration chambers.
Polyethylene glycol experiment
The PEG inclusion study utilised 12 Brangus steers (B. indicus × B. taurus) initially weighing 325 ± 9.8 kg (mean ± s.e.), already adapted to the procedures and a hay–legume diet. Following a 7-day adaption to either cv. Redlands or cv. Wondergraze at 30% of the diet DM and inoculation with S. jonesii mixed culture, PEG4000 was dissolved in water and mixed with feed of eight (four on Redlands, four on Wondergraze) of the animals at 400 g PEG/day. Cattle were dosed once per day for the last 14 days of the 21-day period. Methane was measured in all 12 cattle on the last 2 days of the 14-day period.
Diets and feeding
Steers had ad libitum access to water and diets. The basal diet consisted of Rhodes grass hay chopped (Roto-grind model 760; Burrows Enterprises, Greeley, CO, USA) to particle length of 5–10 cm to facilitate mixing with the fresh leucaena. A mineral–vitamin supplement was provided in the form of a lick block throughout the experiment (Olssons Trace element with copper and cobalt; Olssons, Yennora, NSW, Australia).
Just before feeding, the hay and fresh leucaena were prepared according to the four inclusion levels: 0%, 18%, 36% and 48%, on a DM basis. The diets were distributed between 09:00 and 10:00 hours. Feed intake was measured throughout the experiment by calculating the difference between feed offered and feed removed 23 h later, adjusted for DM. Cattle were weighed weekly before feeding.
Sampling and analysis
Samples of hay and leucaena offered and refusals were collected daily and bulked over 7 days when cattle were not in chambers, and over 3 days when cattle were in chambers, with the 3 days corresponding to the day before and the 2 days during which the steers were in chambers. Samples (10%) of the daily refusals were collected, and the samples pooled by animal.
Feed samples were dried in oven at 65°C, ground to 1 mm and packed in plastic bags before being sent to CSIRO Floreat Laboratory in Perth, Western Australia, for analysis. Ash, acid detergent fibre (ADF), neutral detergent fibre (NDF) and nitrogen (N) contents and DM digestibility (DMD) of feed samples were predicted by near-infrared spectroscopy (NIRS) using a scanning monochromator (Model 6500; NIRSystem, Silver Spring, MD, USA). The calibration database is large, about 1400 samples composed of tropical forages collected in Queensland. All samples (leucaena, hay, refusals) were predicted with the same prediction equations (Coates and Dixon 2011). The chemical composition of feed offered was computed from the proportional composition of the two forages according to their respective inclusion levels in the diet. The DM determinations of the bulked samples collected for the offered hay and leucaena, and refusals, were used to calculate DM intake (DMI).
Validation of NIR predictions was undertaken for N, the fibre fractions (ADF, NDF) and DMD by analysing 10% of samples by wet chemistry. Total N was determined using a CN 628 N analyser (LECO, St. Joseph, MI, USA). ADF and NDF contents were determined by the Ankom method (Ankom, Fairport, NY, USA) and DMD by the in-house pepsin cellulase method corrected for in vivo digestibility for cattle (Klein and Baker 1993). The R2 of correlations between wet chemistry and NIRS were found to be >0.93 for all analytes. Ash content of feed samples was measured by combustion at 550°C. CP content was calculated by multiplying N by 6.25, and hemicellulose was obtained by subtracting the ADF content from the NDF content.
Ruminal samples for pH, ammonia-N and volatile fatty acids (VFAs) were collected 3 h after feeding on Day 14 of each period, following the release of the steers from the methane chambers. The animals were restrained in a commercial cattle crush while the operator inserted an oral stomach tube. The pH was determined immediately, using a digital pH meter. Unstrained rumen fluid (4 mL) was preserved with 1 mL 20% metaphosphoric acid and stored frozen at −80°C prior to analysis for VFAs (Gagen et al. 2014) and ammonia-N (Chaney and Marbach 1962).
Measurement of methane emissions
Four open-circuit respiration chambers were used to determine individual CH4 and H2 emissions. These chambers had an internal volume of 23.04 m3 (4 m by 2.4 m by 2.4 m) and were constructed of a steel frame over which 4.10-mm clear polycarbonate was attached providing full visibility for each animal. A modified squeeze crush defined a confinement area in each chamber, and the floor was made of plastic grid allowing faeces and urine to flow into a container located below. Each chamber was equipped with a nose-activated drinking bowl and wheeled feed bin. Chambers were maintained at 2.0°C below ambient air temperature and under slight negative pressure (−10 Pa). Air was drawn from outside the building at a rate of 3000 L/min. Samples from the airflow coming into and out of the chambers were filtered, dehumidified, dried and refrigerated before analysis. The composition of air samples was determined by infrared for CH4, CO2 and O2 and by gas chromatography for H2. A full description of the components and functioning of these chambers is provided by Martinez-Fernandez et al. (2016).
For the baseline and each of the four 2-week periods, animals were held in individual pens from Day 1 to 11, transferred to open circuit respiration chambers on Day 12, and returned to individual pens on Day 14. Thus, methane emissions were collected over two consecutive days.
Statistical analyses
All statistical analyses were performed using SAS software version 9.4 (SAS Institute, Cary, NC, USA). The individual steer was the experimental unit for DMI, gas (CH4 and H2) emissions, LW gain, ruminal VFAs, ammonia-N and pH. The influence of the fixed factors (level of leucaena in the diet and cultivar) on gas emissions, DMI, and ruminal VFAs, ammonia-N and pH were analysed by using a mixed model procedure. The experiment used a split-plot design (cultivar and level), with polynomial analysis to assess the response (linear or quadratic) to leucaena inclusion level. With the exception of iso-valerate, cultivar had no effect on any variables measured, and excepting H2 production, no interactions were observed between cultivar and level. With the exceptions noted above (Supplementary material Tables S1 and S2), the analysis was restricted to the main effect of level of leucaena inclusion.
Results
Cultivar effect
Both Wondergraze and Redlands are recently developed cultivars of leucaena selected for ease of establishment and yield. Redlands was commercially released in 2019 as a psyllid-resistant cultivar. There were no cultivar effects for any of the parameters measured in this study apart from the exceptions noted above (Tables S1 and S2).
Main experiment: diet quality, intake and performance
The Rhodes grass hay that constituted the basal diet was of poor quality as evidenced by high concentrations of fibre and low CP content. With DMD <50%, the hay quality was considered typical of dry season grasses in northern Australia (Table 1). The chemical composition of the freshly harvested leucaena (leaves plus stems <10 mm diameter) was similar in the two cultivars. As expected, the nutritive value of leucaena was superior to that of Rhodes grass, with CP content and DMD in excess of 14% and 60%, respectively.

|
Over the entire experimental period of 56 days, increasing the percentage of leucaena in the diet from 0% to 48% increased DMI according to a quadratic response (P < 0.001) from ∼13 to 17.5 g/kg bodyweight (BW) at 36% inclusion of leucaena (Table 2). There was no effect of cultivar on DM intake, but intake was apparently 5% lower when cattle were in chambers than pens. During confinement in chambers, treatment response in DMI and chemical components increased in a quadratic fashion (P < 0.001), with greatest intake being achieved when leucaena was included in the diet at 36% (Table 2).

|
Main experiment: rumen fermentation characteristics and gas production
With the exception of iso-valerate where the molar proportion was higher for Redlands than Wondergraze (P = 0.028), cultivar had no effect on any of the rumen fermentation characteristics (Table 3). Total VFA concentration averaged 65 mg/dL across all inclusion levels of leucaena and exhibited a positive linear response to leucaena inclusion (P = 0.024). Proportions of VFAs were not greatly influenced by leucaena inclusion level, although there was a positive linear response for valerate (P = 0.015). Rumen ammonia increased linearly (P < 0.001) as leucaena inclusion level (and CP intake) increased. The pH of rumen liquor exhibited a quadratic response to leucaena inclusion, with higher values at the 18% and 36% inclusion rates (P < 0.001).

|
As a consequence of increasing DMI, methane production increased as leucaena inclusion level increased (quadratic, P < 0.001) (Table 4). However, when methane was expressed per unit DMI, methane yield decreased in a linear fashion (P < 0.001).

|
Relative to results from the same animals during baseline measurements, methane yield was reduced by 15%, 20% and 22% at the 18%, 36% and 48% inclusion rates, respectively (Table 4). Production of H2 exhibited a positive linear response to leucaena inclusion level (P < 0.001). However, there was a significant cultivar × level interaction (P = 0.022; Table S2) such that H2 levels were higher for Wondergraze at intermediate inclusion levels but not at extreme inclusion levels. Yield of H2 also increased with leucaena inclusion level, with the quadratic response being significant (P = 0.011). Relationships are presented between leucaena inclusion level in the diet and DM intake (Fig. 1a), methane production (Fig. 1b) and methane production relative to baseline data (Fig. 1c). The relationship for methane yield was linear and can be defined as:
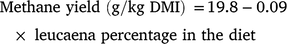

|
Polyethylene glycol experiment
The NIR analysis revealed higher nutritive value of both hay and the two leucaena cultivars in the PEG experiment than in the main trial. However, as with the main experiment, leucaena was of higher quality than Rhodes grass and differences between leucaena cultivars were small (Table 5). Including PEG in the diet did not influence DMI. However, methane production exhibited a numeric increase in response to PEG (P = 0.114), which attained significance when expressed relative to DMI (P = 0.018; Table 6). The pH was unaffected by PEG.

|

|
Discussion
Feed characteristics and animal performance
The composition and nutritive profile of the two leucaena cultivars was very similar but atypical of data recorded in the literature (Shelton and Dalzell 2007; Kennedy and Charmley 2012; Soltan et al. 2013; Archimède et al. 2016). The low CP in the main experiment (∼15%) was surprising and initially attributed to the inclusion of green stems (<10 mm diameter) in the diet. However, the same harvesting method was used the following season, and CP for both cultivars averaged 21%. The months following the 2019 wet season were dry, and the leucaena matured quickly, flowering and setting seed during the main experiment period from March to late June. By contrast, during 2020, out-of-season rainfall in May elicited fresh leucaena growth that coincided with the timing of the PEG experiment. The differences in CP content between years was attributed to the loss of leaf and high proportion of pods in the main study compared with the PEG study.
The Rhodes grass hay used as the basal diet in the main experiment was of poor quality, and this was reflected by the low DMI and LW gain exhibited in steers fed the 0% leucaena diet. Nevertheless, these nutritional characteristics were representative of dry season grass pastures in northern Australia. The response to increasing the proportion of leucaena in intake and growth was predictable considering the increase in N and metabolisable energy (ME) as the proportion of leucaena in the diet was increased. Similar results have been observed elsewhere (Hove et al. 2001; Soltan et al. 2013; Rira et al. 2015; Archimède et al. 2016; Molina et al. 2016). Exceptions to this intake response to leucaena are restricted to studies where the nutrient content of the diet has been held equivalent across treatments (Montoya-Flores et al. 2020).
Effect of leucaena on methane production
Methane yields per kg DMI when feeding 100% grass hay during the baseline measurements (19.6 g/kg DMI) and for the 0% leucaena diet (19.9 g/kg DMI) were close to values described in the literature for non-methanogenic forages, using open-path respiration chambers (Kennedy and Charmley 2012; Hristov et al. 2013; Herd et al. 2014) but lower than those (23–27 g/kg DMI) for a tropical grass hay reported by Martinez-Fernandez et al. (2016, 2017). In a meta-analysis of published Australian data including dairy, temperate beef and tropical beef cattle fed predominantly forage diets, Charmley et al. (2016) obtained a single relationship of 20.7 g methane/kg DMI across an intake range of 3–28 kg/day.
Our results are characteristic of several studies where leucaena and other tropical legumes are added to low-quality grass diets that show methane production per head rising with increasing DMI, but methane yield per unit intake declining (Archimède et al. 2016; Molina et al. 2016; Suybeng et al. 2020). Typically, inclusion of the legume increases DMI as a consequence of increases in the N and ME concentration of the diet. The increase in methane production can be attributed to the increase in rumen available energy and retained N (Hove et al. 2001). When diet quality is equalised by formulating diets to be equivalent in N and ME, the intake response disappears and both methane production and yield decline (Montoya-Flores et al. 2020).
In the present study, the response to leucaena inclusion was measured in two ways: (i) by comparing the dose response relative to the control (0% leucaena in the diet) within each experimental period; and (ii) by measuring the response of individual animals relative to their baseline measurements taken at the start of the experiment. Although the results are broadly similar, the second approach indicated a curvilinear response. Beyond 36% leucaena inclusion, there was no further response in methane yield. These data suggest that an inclusion level of ∼35% leucaena in the diet would provide the maximum benefit in methane avoidance. This is important because, under typical grazing conditions, leucaena may contribute only up to 40% of the dietary DM intake (Dalzell et al. 2012; Cardona et al. 2015; Bowen et al. 2016). Archimède et al. (2016) observed a 40% reduction in methane yield when sheep were fed leucaena leaf pellets at 44% of dietary DM, whereas Molina et al. (2016) observed a 17% reduction in methane yield when leucaena was included at 26% of the dietary DM of growing heifers. Montoya-Flores et al. (2020) observed a 14% reduction in methane yield for a similar level of leucaena inclusion. The results in the present study are consistent with those of Molina et al. (2016) and Montoya-Flores et al. (2020), but the response found by Archimède et al. (2016) was nearly two-fold. This could reveal a difference between cattle and sheep, or could be related to the levels of bioactive compounds in the leucaena. If data from our experiment and from Archimède et al. (2016), Molina et al. (2016) and Montoya-Flores et al. (2020) are standardised to consider methane reduction at a theoretical 35% legume inclusion rate, the reductions in methane yield are 16%, 21%, 28% and 14% for the four datasets, respectively. This range in methane abatement gives some indication of the potential for leucaena to reduce methane emissions across a range of growing conditions and animal species.
Inhibition of methane, particularly when extreme, can lead to increases in hydrogen losses (Martinez-Fernandez et al. 2016). The marked increase in H2 production and yield for the 48% leucaena inclusion diet would suggest that the rumen had reached its potential to redirect hydrogen to alternative sinks such as propionate (Martinez-Fernandez et al. 2017). Excess H2 in the rumen could have been a factor in the quadratic response in performance and DMI because no benefits were observed in either variable for the 48% leucaena inclusion in the diet.
Possible causes for methane reduction
Leucaena contains a number of plant compounds frequently cited as the bioactive agents in reducing methane emissions in ruminants (Soltan et al. 2013; Aboagye et al. 2019). Foremost among these are tannins, a group of polyphenolic compounds broadly classified according to molecular structure and weight as condensed or hydrolysable tannins (Huang et al. 2011; Aboagye and Beauchemin 2019). Tannin content can vary widely in leucaena according to the species (Jones and Palmer 2002), and edaphic and climatic influences, management and growth stage (Jayanegara et al. 2012; Aboagye et al. 2019). The relative concentrations of condensed and hydrolysable tannins may also vary, and because bioactivity is related to the type and molecular weight of tannins, this can influence the relationship between tannin concentration and methane production in the rumen (Aboagye and Beauchemin 2019). For example, Soltan et al. (2013) concluded that the presence of condensed tannins (measured as leucocyanidin equivalents) in leucaena had no effect on methane yield. By contrast, Archimède et al. (2016) demonstrated a relationship between condensed tannin concentration (using quebracho standard) and methane reduction, with higher tannin concentration resulting in a greater reduction in methane yield. Quantification of tannins and their bioactivity is difficult (Makkar 2003) and a biological measure of bioactivity using PEG can be used as a proxy by nullifying tannin bioactivity (Soltan et al. 2013). This approach was chosen for the present study.
Blocking the action of tannins through the addition of PEG to the diet increased methane yield by 8% when leucaena was fed at 30% of dietary DM. This increase was smaller than expected considering the 13% reduction interpolated from the dose-response study and suggests that tannins may have accounted for approximately two-thirds of the methane reduction, but other mechanisms were also at play. However, the possibility that the PEG level was insufficient to nullify the entire tannin effect cannot be ruled out. In that case, the actual tannin effect may have been higher than observed. Tannins are thought to affect methane production through two main processes: a direct antimicrobial effect on methanogens and protozoa (Tavendale et al. 2005; Animut et al. 2008), and an indirect effect on carbohydrate fermentation favouring reduced acetate and increased propionate through an effect on fibrolytic bacteria (Rira et al. 2015). The effect of tannins on protein solubility in the rumen may further influence rumen fermentation by altering the balance between soluble carbohydrate and rumen-degradable protein for microbial protein synthesis (McSweeney et al. 2001).
From the present study, it was concluded that approximately two-thirds of the methane reduction response could be attributed to tannins provided they were completely neutralised by PEG. The remaining one-third of the response could be related to changes in rumen fermentation in response to adding a high-quality legume. It is widely documented that methane yield is influenced by structural carbohydrate concentration in the diet. van Lingen et al. (2019) conducted a meta-analysis of 852 high-forage diets, and although DMI was the major determinant of methane production, NDF was the second most important. If the van Lingen equation (DMI = 23.8 + 13.5 × DMI + 0.855 × NDF%) is applied to the diets in our study, there was a 5% reduction in methane yield attributed to difference in NDF concentration between 0% and 35% inclusion of leucaena. This accounts for the 33% of methane mitigation that could not be attributed to tannins.
The influence of NDF on methane production in the rumen has been attributed to both physical and biochemical effects on methanogenesis. Reducing NDF in the diet increases rate of passage and reduces retention time (Perry et al. 2017) and may reduce methanogenesis. Reducing NDF in the diet also affects the concentration and molar proportions of VFAs in the rumen. These changes lead to a balance away from acetate towards propionate as evidenced by reductions in acetate:propionate ratios resulting in a reduction in H2 production due to utilisation for propionate formation. In this experiment, as with many forage-based studies (Rira et al. 2015; Perry et al. 2017; Suybeng et al. 2020, 2021), the shift in the acetate:propionate ratio was small but consistent with the changes in NDF content of the diet.
Total VFA concentration increased with increasing leucaena inclusion in the diet. This was expected considering the improvement in diet quality resulting from leucaena addition. Others have found decreases in VFA concentration through the addition of tanniniferous compounds in forages (Beauchemin et al. 2007; Rira et al. 2015). However, in the present study, it was apparent that the improvements in diet quality more that countered any potential negative effects of leucaena tannins on rumen fermentation, which have been shown to be minor (McSweeney et al. 1999). Apart from the change in acetate:propionate ratio, there were few effects on the molar proportions of individual VFAs, although a shift in fermentation from acetate to longer chain fatty acids was observed in cattle grazing leucaena (Tomkins et al. 2019). Other studies with leucaena have shown few effects on molar proportions of VFAs other than acetate and propionate (Rira et al. 2015; Montoya-Flores et al. 2020). In our experiment, pH responded quadratically to increasing leucaena in the diet, indicating a reduction in H+ concentration in rumen fluid, particularly at the intermediate inclusion levels of leucaena. This was contrary to expectation based on acetate:propionate ratio and may have been influenced by elevated H2 concentration (Ungerfeld 2020). By contrast, Rira et al. (2015) found that pH was reduced with leucaena feeding, and Montoya-Flores et al. (2020) observed no effects.
Effect of the basal diet on methane response
The methane response to bioactives can also be influenced by the nature of the companion species in the diet. Aboagye et al. (2019) supplemented lucerne silage with different forms of hydrolysable tannin and showed small but variable reductions in methane yield (0–9%). In their study the quality of the companion forage, in this case lucerne silage, was very high (19.8% CP, 42.2% NDF). In the present study the companion forage was poor-quality Rhodes grass hay (5.1% CP, 73.1% NDF), and a much larger response in methane yield was observed. Suybeng et al. (2020) observed a reduction in methane yield when poor-quality Rhodes grass was fed with increasing proportions of the tannin-containing tropical legume Desmanthus. However, in a subsequent study where diets were isonitrogenous (10% CP, 68% NDF), by substituting Desmanthus with lucerne, no effect on methane yield was observed (Suybeng et al. 2021). In both studies, total phenolics (as catechin equivalents) were similar (1.2–2.3% DM).
In the main experiment, the CP content of the basal hay in the ration at only 5.1% was low and potentially insufficient to maintain optimal microbial activity in the rumen. Rumen ammonia concentration on the control diet was 3.7 mg/dL, which is below the generally accepted level for maximum microbial protein synthesis of 5 mL/dL (Satter and Slyter 1974). Increasing the CP of the diet through supplementation with leucaena increased both CP intake and rumen ammonia in a linear fashion. It can be concluded that increasing the availability of ammonia in the rumen would have increased microbial protein synthesis, VFA concentration and DMI. In spite of the predicted increase in microbial protein synthesis, there was no concomitant increase in concentration of branch-chain fatty acids, which are derived from fermentation of protein (Keeney et al. 1962).
Conclusions
The addition of leucaena to poor-quality tropical grass diets increased voluntary intake and reduced methane yield. The relationship between incremental levels of leucaena inclusion was described according to the equation:

Although the relationship was linear, there was evidence that a 36% inclusion rate was optimal in terms of animal performance and methane emissions. The data suggest that the antimethanogenic effect of tannins, if completely neutralised, accounted for two-thirds of the methane reduction, with improved diet quality accounting for the other third. In parts of Queensland where leucaena can be successfully established, it can provide a reliable source of high-quality forage to supplement low-quality grass pastures and can also reduce methane emissions by up to 20%.
Supplementary material
Supplementary material is available online.
Data availability
The data used to generate the results in the paper will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Funding for this research was provide through the Meat & Livestock Australia contract B.GBP.0026. Antoine Stifkens received support through the University of Liège student internship program.
Acknowledgements
The authors thank the staff of the Lansdown Research Station, Wayne Flintham and Steve Austin for management of the leucaena and husbandry of the livestock essential to this paper. Holly Reid, Elizabeth Hulm, Jag Padmanabha, Jane Cheung, Wendy Smith and Pete Giacomantonio are acknowledged for their technical assistance. Special thanks to Greg Bishop-Hurley for statistical analysis and the leucaena harvesting team, especially Jonno Ramsay and Charlie Sinclair.
References
Aboagye IA, Beauchemin KA (2019) Potential of molecular weight and structure of tannins to reduce methane emission from ruminants: a review. Animals 9, 856| Potential of molecular weight and structure of tannins to reduce methane emission from ruminants: a review.Crossref | GoogleScholarGoogle Scholar |
Aboagye IA, Oba M, Koenig KM, Zhao GY, Beauchemin KA (2019) Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. Journal of Animal Science 97, 2230–2244.
| Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage.Crossref | GoogleScholarGoogle Scholar | 30906949PubMed |
Animut G, Puchala R, Goetsch AL, Patra AK, Sahlu T, Varel VH, Wells J (2008) Methane emission by goats consuming different sources of condensed tannins. Animal Feed Science and Technology 144, 228–241.
| Methane emission by goats consuming different sources of condensed tannins.Crossref | GoogleScholarGoogle Scholar |
Archimède H, Rira M, Barde DJ, Labirin F, Marie-Magdeleine C, Calif B, Periacarpin F, Fleury J, Rochette Y, Morgavi DP, Doreau M (2016) Potential of tannin-rich plants, Leucaena leucocephala, Glyricidia sepium and Manihot esculenta, to reduce enteric methane emissions in sheep. Journal of Animal Physiology and Animal Nutrition 100, 1149–1158.
| Potential of tannin-rich plants, Leucaena leucocephala, Glyricidia sepium and Manihot esculenta, to reduce enteric methane emissions in sheep.Crossref | GoogleScholarGoogle Scholar | 27870287PubMed |
Australian Greenhouse Emissions Information System (2019) National greenhouse gas inventory – Paris agreement. Paris Agreement Inventory. Australian Government, Canberra, ACT, Australia. http://climatechange.gov.au. (Accessed 15 October 2021)
Beauchemin KA, McGinn SM, Martinez TF, McAllister TA (2007) Use of condensed tannin extract from quebracho trees to reduce methane missions from cattle. Journal of Animal Science 85, 1990–1996.
| Use of condensed tannin extract from quebracho trees to reduce methane missions from cattle.Crossref | GoogleScholarGoogle Scholar | 17468433PubMed |
Beutel TS, Corbet DH, Hoffmann MB, Buck SR, Kienzle M (2018) Quantifying leucaena cultivation extent on grazing land. The Rangeland Journal 40, 31–38.
| Quantifying leucaena cultivation extent on grazing land.Crossref | GoogleScholarGoogle Scholar |
Bowen MK, Chudleigh F, Buck S, Hopkins K (2016) Productivity and profitability of forage options for beef production in the subtropics of northern Australia. Animal Production Science 58, 332–342.
| Productivity and profitability of forage options for beef production in the subtropics of northern Australia.Crossref | GoogleScholarGoogle Scholar |
Cardona CAC, Naranjo-Ramírez JF, Morales AMT, Correa GA, Rosales RB (2015) Dry matter and nutrient intake and diet composition in Leucaena leucocephala-based intensive silvopastoral systems. Tropical and Subtropical Agroecosystems 18, 303–311.
Chaney AL, Marbach EP (1962) Modified reagents for determination of urea and ammonia. Clinical Chemistry 8, 130–132.
| Modified reagents for determination of urea and ammonia.Crossref | GoogleScholarGoogle Scholar | 13878063PubMed |
Charmley E, Williams SRO, Moate PJ, Hegarty RS, Herd RM, Oddy VH, Reyenga P, Staunton KM, Anderson A, Hannah MC (2016) A universal equation to predict methane production of forage-fed cattle in Australia. Animal Production Science 56, 169–180.
| A universal equation to predict methane production of forage-fed cattle in Australia.Crossref | GoogleScholarGoogle Scholar |
Coates DB, Dixon RM (2011) Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages. Journal of Near Infrared Spectroscopy 19, 507–519.
| Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages.Crossref | GoogleScholarGoogle Scholar |
Dalzell SA, Burnett DJ, Dowsett JE, Forbes VE, Shelton HM (2012) Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala pastures in Queensland, Australia. Animal Production Science 52, 365–372.
| Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala pastures in Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Gagen EJ, Wang J, Padmanabha J, Liu J, de Carvalho IPC, Liu J, Webb RI, Al Jassam R, Morrison M, Denman SE, McSweeney CS (2014) Investigation of a new acetogen isolated from an enrichment of the tamar wallaby forestomach. BMC Microbiology 7, 1122
| Investigation of a new acetogen isolated from an enrichment of the tamar wallaby forestomach.Crossref | GoogleScholarGoogle Scholar |
Harrison MT, McSweeney CS, Tomkins NW, Eckard RJ (2015) Improving greenhouse gas emissions intensities of subtropical and tropical beef farming systems using Leucaena leucocephala. Agricultural Systems 136, 138–146.
| Improving greenhouse gas emissions intensities of subtropical and tropical beef farming systems using Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar |
Herd RM, Arthur PF, Donoghue KA, Bird SH, Bird-Gardiner T, Hegarty RS (2014) Measures of methane production and their phenotypic relationships with growth and body composition traits in beef cattle. Journal of Animal Science 92, 5267–5274.
| Measures of methane production and their phenotypic relationships with growth and body composition traits in beef cattle.Crossref | GoogleScholarGoogle Scholar | 25349368PubMed |
Hove L, Topps JH, Sibanda S, Ndlovu LR (2001) Nutrient intake and utilisation by goats fed dried leaves of the shrubs legumes Acacia angustissima, Calliandra calothyrsus and Leucaena leucocephala as supplements to native pasture hay. Animal Feed Science and Technology 91, 95–106.
| Nutrient intake and utilisation by goats fed dried leaves of the shrubs legumes Acacia angustissima, Calliandra calothyrsus and Leucaena leucocephala as supplements to native pasture hay.Crossref | GoogleScholarGoogle Scholar |
Hristov AN, Oh J, Firkins JL, Dijkstra J, Kebreab E, Waghorn G, Makkar HPS, Adesogan AT, Yang W, Lee C, Gerber PJ, Henderson B, Tricarico JM (2013) Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. Journal of Animal Science 91, 5045–5069.
| Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options.Crossref | GoogleScholarGoogle Scholar | 24045497PubMed |
Huang XD, Liang JB, Tan HY, Yahya R, Ho YW (2011) Effects of Leucaena condensed tannins of differing molecular weights on in vitro CH4 production. Animal Feed Science and Technology 166, 373–376.
| Effects of Leucaena condensed tannins of differing molecular weights on in vitro CH4 production.Crossref | GoogleScholarGoogle Scholar |
Jayanegara A, Leiber F, Kreuzer M (2012) Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments. Journal of Animal Physiology and Animal Nutrition 96, 365–375.
| Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments.Crossref | GoogleScholarGoogle Scholar | 21635574PubMed |
Jones RJ, Palmer B (2002) Assessment of condensed tannin concertation in a collection of Leucaena species using 14C-labelled polyethylene glycol (PEG 4000). Tropical Grasslands 36, 47–53.
Keeney M, Katz I, Allison MJ (1962) On the probable origin of some milk fat acids in rumen microbial lipids. Journal of the American Oil Chemists’ Society 39, 198–201.
| On the probable origin of some milk fat acids in rumen microbial lipids.Crossref | GoogleScholarGoogle Scholar |
Kennedy PM, Charmley E (2012) Methane yields from Brahman cattle fed tropical grasses and legumes. Animal Production Science 52, 225–239.
| Methane yields from Brahman cattle fed tropical grasses and legumes.Crossref | GoogleScholarGoogle Scholar |
Klein L, Baker SK (1993) Composition of the fractions of dry, mature subterranean clover digested in vivo and in vitro. In ‘Proceedings of the 17th International Grassland Congress’. Palmerston North, Hamilton, Lincoln, NZ/Rockhampton, Qld. pp. 593–595. (International Grassland Congress)
Makkar HP (2003) ‘Quantification of tannins in tree and shrub foliage: a laboratory manual.’ (Kluwer Academic Publishers: Dordrecht, The Netherlands)
Martinez-Fernandez G, Denman SE, Yang C, Cheung J, Mitsumori M, McSweeney CS (2016) Methane inhibition alters the microbial community, hydrogen flow, and fermentation response in the rumen of cattle. Frontiers in Microbiology 7, 1122
| Methane inhibition alters the microbial community, hydrogen flow, and fermentation response in the rumen of cattle.Crossref | GoogleScholarGoogle Scholar | 27486452PubMed |
Martinez-Fernandez G, Denman SE, Cheung J, McSweeney CS (2017) Phloroglucinol degradation in the rumen promotes the redirection of hydrogen when methanogenesis is suppressed. Frontiers in Microbiology 8, 1871
| Phloroglucinol degradation in the rumen promotes the redirection of hydrogen when methanogenesis is suppressed.Crossref | GoogleScholarGoogle Scholar | 29051749PubMed |
McSweeney CS, Palmer B, Bunch R, Krause DO (1999) In vitro quality assessment of tannin-containing tropical shrub legumes: protein and fibre digestion. Animal Feed Science and Technology 82, 227–241.
| In vitro quality assessment of tannin-containing tropical shrub legumes: protein and fibre digestion.Crossref | GoogleScholarGoogle Scholar |
McSweeney CS, Palmer B, Bunch R, Krause DO (2001) Effect of the tropical forage calliandra on microbial protein synthesis and ecology in the rumen. Journal of Applied Microbiology 90, 78–88.
| Effect of the tropical forage calliandra on microbial protein synthesis and ecology in the rumen.Crossref | GoogleScholarGoogle Scholar | 11155126PubMed |
Meat & Livestock Australia (2021) CN30 overview CN30: carbon neutral by 2030. Meat & Livestock Australia, Sydney, NSW, Australia. http://mla.com.au (Accessed 15 October 2021)
Molina IC, Angarita EA, Mayorga OL, Chará J, Barahona-Rosales R (2016) Effect of Leucaena leucocephala on methane production of Lucerna heifers fed a diet based on Cynodon plectostachyus. Livestock Science 185, 24–29.
| Effect of Leucaena leucocephala on methane production of Lucerna heifers fed a diet based on Cynodon plectostachyus.Crossref | GoogleScholarGoogle Scholar |
Montoya-Flores MD, Molina-Botero IC, Arango J, Romano-Muñoz JL, Solorio-Sánchez FJ, Aguilar-Pérez CF, Ku-Vera JC (2020) Effect of dried leaves of Leucaena leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers. Animals 10, 300
| Effect of dried leaves of Leucaena leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers.Crossref | GoogleScholarGoogle Scholar |
Perry LA, Al Jassim R, Gaughan JB, Tomkins NW (2017) Effect of feeding forage characteristic of wet- or dry-season tropical C4 grass in northern Australia, on methane production, intake and rumen outflow rates in Bos indicus steers. Animal Production Science 57, 2033–2041.
| Effect of feeding forage characteristic of wet- or dry-season tropical C4 grass in northern Australia, on methane production, intake and rumen outflow rates in Bos indicus steers.Crossref | GoogleScholarGoogle Scholar |
Poppi DP, McLennan SR (2010) Nutritional research to meet future challenges. Animal Production Science 50, 329–338.
| Nutritional research to meet future challenges.Crossref | GoogleScholarGoogle Scholar |
Radrizzani A, Dalzell SA, Shelton HM (2011) Effect of environment and plant phenology on prediction of plant nutrient deficiency using leaf analysis in Leucaena leucocephala. Crop & Pasture Science 62, 248–260.
| Effect of environment and plant phenology on prediction of plant nutrient deficiency using leaf analysis in Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar |
Rira M, Morgavi DP, Archimède H, Marie-Magdeleine C, Popova M, Bousseboua H, Doreau M (2015) Potential of tannin-rich plants for modulating ruminal microbes and ruminal fermentation in sheep. Journal of Animal Science 93, 334–347.
| Potential of tannin-rich plants for modulating ruminal microbes and ruminal fermentation in sheep.Crossref | GoogleScholarGoogle Scholar | 25568379PubMed |
Satter LD, Slyter LL (1974) Effect of ammonia concentration on rumen microbial protein production in vitro. British Journal of Nutrition 32, 199–208.
| Effect of ammonia concentration on rumen microbial protein production in vitro.Crossref | GoogleScholarGoogle Scholar |
Shelton M, Brewbaker JL (1998) Leucaena leucocephala – the most widely used forage tree legume. In ‘Forage tree legumes in tropical agriculture’. (Eds RC Gutteridge, HM Shelton) pp. 15–29. (Tropical Grassland Society of Australia)
Shelton M, Dalzell S (2007) Production, economic and environmental benefits of leucaena pastures. Tropical Grasslands 41, 174–190.
Soltan YA, Morsy AS, Sallam SMA, Lucas RC, Louvandini H, Kreuzer M, Abdalla AL (2013) Contribution of condensed tannins and mimosine to the methane mitigation caused by feeding Leucaena leucocephala. Archives of Animal Nutrition 67, 169–184.
| Contribution of condensed tannins and mimosine to the methane mitigation caused by feeding Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar | 23742642PubMed |
Suybeng B, Charmley E, Gardiner CP, Malu-Aduli BS, Malau-Aduli AEO (2020) Supplementing northern Australian beef cattle with Desmanthus tropical legume reduces in vivo methane emissions. Animals 10, 2097
| Supplementing northern Australian beef cattle with Desmanthus tropical legume reduces in vivo methane emissions.Crossref | GoogleScholarGoogle Scholar |
Suybeng B, Mwangi FW, McSweeney CS, Charmley E, Gardiner CP, Malu-Aduli BS, Malau-Aduli AEO (2021) Response to climate change: evaluation of methane emissions in northern Australian beef cattle on a high quality diet supplemented with Desmanthus using open-circuit respiration chambers and GreenFeed emission monitoring systems. Biology 10, 2097
| Response to climate change: evaluation of methane emissions in northern Australian beef cattle on a high quality diet supplemented with Desmanthus using open-circuit respiration chambers and GreenFeed emission monitoring systems.Crossref | GoogleScholarGoogle Scholar |
Tan HY, Sieo CC, Abdullah N, Liang JB, Huang XD, Ho YW (2011) Effects of condensed tannins from leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro. Animal Feed Science and Technology 169, 185–193.
| Effects of condensed tannins from leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro.Crossref | GoogleScholarGoogle Scholar |
Tavendale MH, Meagher LP, Pacheco D, Walker N, Attwood GT, Sivakumaran S (2005) Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Animal Feed Science and Technology 123, 403–419.
| Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis.Crossref | GoogleScholarGoogle Scholar |
Tomkins N, Harrison M, McSweeney CS, Denman S, Charmley E, Lambrides CJ, Dalal R (2019) Greenhouse gas implications of leucaena-based pastures. Can we develop an emissions reduction methodology for the beef industry? Tropical Grasslands-Forrajes Tropicales 7, 267–272.
| Greenhouse gas implications of leucaena-based pastures. Can we develop an emissions reduction methodology for the beef industry?Crossref | GoogleScholarGoogle Scholar |
Ungerfeld EM (2020) Metabolic hydrogen flows in rumen fermentation: principles and possibilities of interventions. Frontiers in Microbiology 11, 589
| Metabolic hydrogen flows in rumen fermentation: principles and possibilities of interventions.Crossref | GoogleScholarGoogle Scholar | 32351469PubMed |
van Lingen HJ, Niu M, Kebreab E, Valadares Filho SC, Rooke JA, Duthie CA, Schwarm A, Kreuzer M, Hynd PI, Caetano M, Eugène M, Martin C, McGee M, O’Kiely P, Hünerberg M, McAllister TA, Berchielli TT, Messana JD, Peiren N, Chaves AV, Charmley E, Cole NA, Lee S-S, Berndt A, Reynolds CJ, Crompton LA, Bayat A, Yáñez-Ruiz DR, Yu Z, Bannink A, Dijkstra J, Casper DP, Hristov AN (2019) Prediction of beef cattle enteric methane production, yield and intensity using an intercontinental database. Agriculture, Ecosystems & Environment 283, 106575
| Prediction of beef cattle enteric methane production, yield and intensity using an intercontinental database.Crossref | GoogleScholarGoogle Scholar |