

Ruminal microbiota is associated with feed-efficiency phenotype of fattening bulls fed high-concentrate diets
S. Costa-Roura A , D. Villalba A D , M. Blanco B C , I. Casasús B C , J. Balcells A and A. R. Seradj A
A D , M. Blanco B C , I. Casasús B C , J. Balcells A and A. R. Seradj A
A Departament de Ciència Animal, Universitat de Lleida, Avinguda Alcalde Rovira Roure 191, 25198, Lleida, Spain.
B Unidad de Producción y Sanidad Animal, Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA), Avenida Montañana 930, 50059, Zaragoza, Spain.
C Instituto Agroalimentario de Aragón – IA2 (CITA-Universidad de Zaragoza), Zaragoza, Spain.
D Corresponding author. Email: daniel.villalba@udl.cat
Animal Production Science - https://doi.org/10.1071/AN20344
Submitted: 5 June 2020 Accepted: 27 November 2020 Published online: 28 January 2021
Journal Compilation © CSIRO 2021 Open Access CC BY
Abstract
Context: Improving feed efficiency in livestock production is of great importance to reduce feeding costs.
Aims: To examine the relationship between ruminal microbiota and variation in feed efficiency in beef cattle fed concentrate-based diets.
Methods: Residual feed intake of 389 fattening bulls, supplied with corn-based concentrate and forage ad libitum, was used to estimate animals’ feed efficiency. Faeces and ruminal fluid samples, from 48 bulls chosen at random, were collected to estimate their forage intake and to determine their apparent digestibility, ruminal fermentation and microbiota. Those animals with extreme values of feed efficiency (high-efficiency (HE, n = 12) and low-efficiency (LE, n = 13)) were subjected to further comparisons. Alpha biodiversity was calculated on the basis of the normalised sequence data. Beta diversity was approached through performing a canonical correspondence analysis based on log-transformed sequence data. Genera differential abundance was tested with an ANOVA-like differential expression analysis and genera interactions were determined applying the sparse correlations for compositional data technique.
Key results: No differences in dry matter intake were found between the two categories of feed efficiency (P = 0.699); however, HE animals had higher apparent digestibility of dry matter (P = 0.002), organic matter (P = 0.003) and crude protein (P = 0.043). The concentration of volatile fatty acids was unaffected by feed efficiency (P = 0.676) but butyrate proportion increased with time in LE animals (P = 0.047). Ruminal microbiota was different between HE and LE animals (P = 0.022); both α biodiversity and genera network connectance increased with time in LE bulls (P = 0.005 for Shannon index and P = 0.020 for Simpson index), which suggests that LE animals hosted a more robust ruminal microbiota. Certain genera usually related to high energy loss through methane production were found to establish more connections with other genera in LE animals’ rumen than in HE ones. Microbiota function capability suggested that methane metabolism was decreased in HE finishing bulls.
Conclusions: Rumen microbiota was associated with feed efficiency phenotypes in fattening bulls fed concentrate-based diets.
Implications: The possible trade-off between feed efficiency and robustness of ruminal microbiota should be taken into account for the optimisation of cattle production, especially in systems with intrinsic characteristics that may constitute a disturbance to rumen microbial community.
Keywords: apparent digestibility, beef cattle, feed efficiency, rumen microbial community.
Introduction
Improving feed efficiency (FE) in beef cattle production systems provides an opportunity to cut down on the cost of feeding livestock. In that sense, residual feed intake (RFI) can be used as an index of FE that is independent of variation in bodyweight (BW) and average daily gain (ADG; Arthur et al. 2001; Schenkel et al. 2004; Arthur and Herd 2008), being the gold standard index to examine biological mechanisms associated with inter-animal differences in FE. Moreover, some studies have demonstrated the possibility of selection for low RFI as a strategy for greenhouse gas mitigation, as it has been correlated with lower methane emission and greater diet digestibility (Herd and Arthur 2009). Limitations in conducting RFI trials (recording BW and feed intake for a long time) and searching for rumen microbial markers to identify efficient animals with low RFI have become a contemporary challenge.
Research in cattle has focused mostly on the microbial response to dietary changes and management practices, whereas trials for understanding the relationship between host FE phenotype and rumen microbiota are scarce and yet to be undertaken (Myer et al. 2015). Previous studies have shown that rumen microbes are responsible for energy supply through producing organic acids (Huntington 1990), and most taxa associated with variation in FE have been related to cellulolytic, fermentative and metabolic activities (Myer et al. 2015). Therefore, differences in the production rate of organic acids lead to variation in nutrient digestibility and fermentation that ultimately change animals’ phenotypic efficiency (Herd and Arthur 2009).
This experiment aimed to understand the relationship between ruminal microbiota and variation in FE of beef cattle fed concentrate-based diets.
Materials and methods
Animals, diets and housing
Residual feed-intake data from two feeding experiments comprising 389 fattening bulls were used to explore relationships between ruminal microbiota and FE. This dataset included 317 animals raised at the research facilities of Cooperativa d’Ivars d’Urgell, SCCP (Ivars d’Urgell, Spain, 41°41′50″N, 0°58′53″E) and 72 animals from the CITA-La Garcipollera Research Station (Jaca, Spain, 42°37′34″N, 0°30′10″W). All procedures were performed under Project Licence CEEA 01-07/16 and approved by the in-house Ethics Committee for Animal Experiments at the University of Lleida. Care and use of animals were in accordance with the Spanish Policy for Animal Protection RD 53/2013, which meets the European Union Directive 2010/63 on the protection of animals used for experimental and other scientific purposes.
Animals raised at the research facilities of Cooperativa d’Ivars d’Urgell, SCCP were distributed in the following four batches: batches Number 1 to Number 3 included 231 Holstein bulls (63–83 animals per batch) and batch Number 4 included 86 Montbeliard bulls. BW and feed intake data were collected on a daily basis. Animals raised at CITA-La Garcipollera Research Station were distributed in the following three batches: batches Number 5 and Number 6 included 28 and 32 Parda de Montaña bulls respectively, and batch Number 7 included 12 Pirenaica bulls. For these animals, BW was measured weekly and feed intake data were collected on a daily basis.
Bodyweight and feed intake data were recorded throughout the entire fattening phase; the first 150 days were considered as the growing phase (121 days old, s.d. 37 days; and 162 kg BW, s.d. 49 kg), followed by a finishing phase, which lasted until animals reached slaughter weight (336 days old, s.d.: 31; and 501 kg BW, s.d. 56 kg).
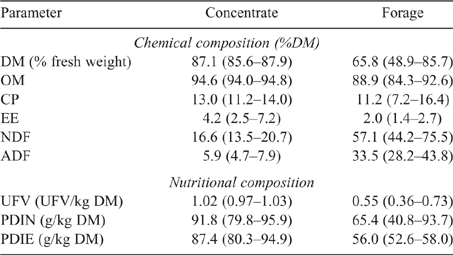
Animals were fed concentrate and forage ad libitum, which were provided separately in two different bunkers, and they had free access to drinking water, following the conventional beef cattle feeding system in Spain. The concentrates used were very similar in composition and their main ingredients were raw corn, corn gluten feed, raw barley, corn dried distillers grains with solubles and raw chickpea; whereas forage used was mainly barley straw (349 animals), oats haylage (20 animals) and vetch haylage (20 animals). Feed chemical and nutritional composition is shown in Table 1.
|
Measurements and sampling
Intake of concentrates was recorded automatically at both research facilities, by using automatic feed stations that were equipped with a feedbunk (provided with a scale) and an individual feeder. When a calf entered the feeder, it was identified and its concentrate intake was obtained by difference between initial and final feedbunk weight. Feed stations available at the research facilities of Cooperativa d’Ivars d’Urgell, SCCP were additionally equipped with a scale under the individual feeder by which the animals were automatically weighed at each visit, whereas at CITA-La Garcipollera Research Station, BW data were recorded manually once a week.
Faeces and ruminal fluid samples from 48 bulls (selected at random within batches) were collected twice, at mid-growing period (GRO, 159 days old and 225 kg BW) and mid-finishing period (FIN, 266 days old and 434 kg BW), for forage intake estimation and digestibility, ruminal fermentation and microbial community characterisation.
Faecal excretion and forage intake were calculated on the basis of concentrate intake and adapting the two indigestible-marker system (Owens and Hanson 1992), by using chromium oxide as an external marker and acid insoluble ash as internal marker. Then, apparent digestibility of dry matter (DM), organic matter (OM) and crude protein (CP) were estimated. Detailed information about marker administration, feed and faeces analytical determinations and apparent digestibility calculations have been described in Costa-Roura et al. (2020).
Ruminal fluid was sampled in the morning by using an oral stomach tube connected to a vacuum pump. Each sample was obtained through two sequential collections. First, ruminal fluid (~200 mL) was collected and discarded to avoid sample contamination with saliva that could get into the tube during its introduction through the animal’s mouth and oesophagus. After that, ruminal fluid (~200 mL) was re-extracted, strained through a cheesecloth and its pH was recorded (Testo 205, Testo AG, Germany). Then, ruminal fluid was sampled for DNA extraction, and determination of ammonia-nitrogen (N) and volatile fatty acid (VFA) concentrations, and immediately frozen on dry ice. Sample preservation conditions and analytical procedures for ammonia-N and VFA determination are detailed in Costa-Roura et al. (2020).
Extraction and sequencing of DNA
Extraction of DNA was performed on freeze-dried ruminal fluid (the initial amount of the sample was 60 mg) through physical disruption (1 min) by using a bead beater (Mini-bead Beater 1, BioSpec Products, USA) and subsequent DNA purification was performed with the QIAamp DNA Stool Mini Kit (ID: 51504; QIAGEN N.V., Germany), with the modifications of greater temperature (95°C) and greater elution time (3 min) to ensure maximum DNA concentration in the final elute. Amplification of DNA was performed by using primers 341F and 805R, which target the V3 and V4 regions of the bacterial and archaeal 16S rRNA. Sequencing was conducted on an Illumina MiSeq 2x300 platform by Era7 Bioinformatics (Spain).
Assembly and filtration of sample reads, as well as operational taxonomic unit (OTU) preparation have been detailed in Costa-Roura et al. (2020).
Estimation of RFI and clustering
Weight data were fitted to a third-degree polynomials model in function of age (Eqn 1) that allows the estimation of the ADG of each animal at any age.
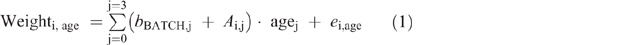
where bBATCH,j is the batch effect (fixed); Ai,j, is the jth random coefficient for the ith animal effect; age is the age of the animal (days) and ei,age the residual term.
Thereafter, ADG_dev was obtained as the first derivative of Eqn 1 for each month by using the monthly average age of each animal (Eqn 2). The individual ADG deviation (ADG_dev) will account for the difference of growth of the animal compared with the average of the batch at each age.

In total, 86 records (3%) out of three standard deviations of the mean were considered as outliers and excluded from the dataset.
Residual feed intake was modelled (Eqn 3) using the random regression coefficient approach proposed by Savietto et al. (2014). The model included batch, age (months), ADGdev, metabolic weight (MW; monthly mean BW0.64) and was defined as follows:
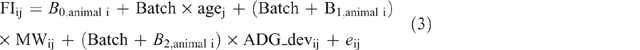
where FIij is DM intake measured for Animal i in Month j and Bk, animal i are the random coefficients for animal effect modelled using an unstructured matrix of variances between them.
The inclusion of batch effect in Eqn 1 and Eqn 3 assured that the FE calculated was not affected by diet differences.
On the basis of the individual coefficients of ADG (B1, animal i) and MW (B2, animal i), animals were segregated into four categories of FE, as follows:
-
Animals with positive coefficients of both ADG and MW belonged to ‘low-efficiency in ADG and low-efficiency in MW’ category.
-
Animals with positive coefficient of ADG but negative coefficient of MW belonged to ‘low-efficiency in ADG and high-efficiency in MW’ category.
-
Animals with negative coefficient of ADG but positive coefficient of MW belonged to ‘high-efficiency in ADG and low-efficiency in MW’ category.
-
Animals with negative coefficients of both ADG and MW belonged to ‘high-efficiency in ADG and high-efficiency in MW’ category.
For the purpose of the present study, the two extreme categories (1 and 4) were considered as high-efficiency (HE, positive RFI) and low-efficiency (LE, negative RFI) animals, respectively. This clustering (HE vs LE) was subjected to bioinformatic analyses of apparent digestibility, ruminal fermentation and microbiota data as explained below.
Bioinformatics
Sequence data were normalised and α biodiversity indices were calculated to measure the variability of OTUs within a sample (R Core Team 2020, Vegan package).
To measure differences in microbiota composition among samples, β diversity was approached through performing a canonical correspondence analysis, based on log-transformed OTU data (zeros were replaced by adding 1 to each value), and including FE (HE vs LE), period (GRO vs FIN) and both ADG and MW coefficients as explanatory variables (R Core Team 2020, Vegan package).
To circumvent the compositional bias problem (Tsilimigras and Fodor 2016; Gloor et al. 2017; Calle 2019), we applied the Aitchison’s centred log ratio (clr) transformation to carry the data to a Euclidean space, after replacing zeros by adding 1 to each value. So as to test the significance of the following effects: FE (HE vs LE), period (GRO vs FIN) and both ADG and MW coefficients on microbiota composition, a permutational multivariate analysis of variance (Adonis) was conducted on the basis of the clr Euclidean distance and calculating statistical significance after 10 000 random permutations (R Core Team 2020, Vegan package). So as to decipher which genera abundance were responsible for the differences among groups, an ANOVA-like differential expression (ALDEx) analysis was conducted over those genera present at least at 50% of the individuals (R Core Team 2020, Aldex2 package; Fernandes et al. 2013). Finally, to describe the interactions within rumen microbial community, we performed a network analysis through sparse correlations for compositional data (SparCC) technique (R Core Team 2020, SpiecEasi package; Friedman and Alm 2012) over those genera present at least at 50% of the individuals. Microbial networks were graphically represented (R Core Team 2020; igraph package) and their complexity was described in terms of number of nodes (genera), number of edges (significant positive or negative correlations), node degree (number of connections that any node establishes with other nodes) and betweenness (measure of centrality in a graph based on shortest paths).
Microbiota functional content was assessed using a topic model approach (R Core Team 2020; themetagenomics package) that consists on (1) capturing groups of co-occurring taxa termed ‘topics’, (2) uncovering within-topic functional potential, and (3) linking these topics and their functional content to specific sample features (e.g. FE phenotypes; Woloszynek et al. 2019).
Statistical analyses
The models of RFI were solved using MIXED procedure of SAS statistical software (SAS v9.4, Cary, NC, USA). Intake, apparent digestibility, ruminal fermentation parameters and microbial α biodiversity data were analysed with a mixed model, including FE (HE vs LE), period (GRO vs FIN) and their interaction as fixed effects and animal as a random effect, to account for repeated measurements (R Core Team 2020, lme4 package). Differences among least square means were assessed using Tukey multiple-comparison test (R Core Team 2020, emmeans package). Individual samples out of three standard deviations of the mean were discarded and not included in the statistical analysis. Results are reported as least square means and standard error of mean. Significant effects were declared at P < 0.05 and tendency to difference at P between 0.05 and 0.10.
Results
The four defined FE categories based on random regression coefficients included between 85 and 108 animals each; the two extreme categories corresponding to HE and LE animals had statistically different means for ADG and MW coefficients (Table S1, available as Supplementary Material to this paper). The set of 48 bulls that were sampled for apparent digestibility, ruminal fermentation and microbiota characterisation were equally distributed within the four categories.
The FE by period interactions were not significant for any of the response variables measured in the present study; thus, only the main effects means are presented and discussed.
Intake, apparent digestibility and ruminal fermentation parameters
Data on DM intake (Table 2) indicated that bulls’ concentrate and forage intakes were similar between FE categories (HE vs LE). However, animals classified as HE had greater apparent digestibility coefficients of DM, OM and CP than did their LE counterparts.

|
Data on ruminal fermentation parameters (Table 2) showed no differences in ruminal pH between HE and LE bulls. Ammonia-N concentration was low and variable among animals; therefore, no statistical differences between FE categories were found. Although the total VFA concentration remained unaffected by FE, numerical differences were found in molar proportions of the main VFA; HE animals had a lower proportion of acetate and a higher proportion of propionate than did LE ones. Butyrate proportion increased with time in the case of LE animals (7.46% vs 9.30% for GRO and FIN periods, respectively; P = 0.047) whereas it remained equal for HE animals (7.95% vs 8.42% for GRO and FIN, respectively; P = 0.908). In contrast, branched-chain VFA proportion (isobutyrate and isovalerate) increased with time only in the case of HE bulls (1.55% vs 2.31% for GRO and FIN, respectively; P = 0.006).
Microbial dataset features
Sequencing procedure yielded an average of (mean ± s.e.m.) 19 862 ± 2215 sequences per sample, resulting in 973 259 sequences in the whole study. In total, 787 OTUs were obtained at the 98% sequence-similarity cut-off levels, with 114 ± 5 as the mean number of OTUs per sample. Good’s coverage value was 99.69 ± 0.03%, suggesting that more than 99% of bacterial and archaeal phylotypes were identified. The unclassified rate of OTUs at genus level was 0.75 ± 0.09%. Shared OTUs by all individuals in each FE category and period were deemed to be core bacterial/archaeal communities. Core community gathered 69.90 ± 2.94% of analysed sequences and was composed of five OTUs, namely, Prevotella ruminicola, unclassified Prevotella (both representing more than 84% of shared sequences), unclassified Roseburia, Sharpea azabuensis and unclassified Methanobrevibacter.
Microbial community biodiversity
Alpha biodiversity (Table 3) was found to be similar among bulls differing in their FE; however, Shannon and Simpson index values increased with time only in LE animals (Shannon index 1.51 vs 2.13, P = 0.005; Simpson index 0.53 vs 0.70, P = 0.020, for GRO and FIN, respectively).

|
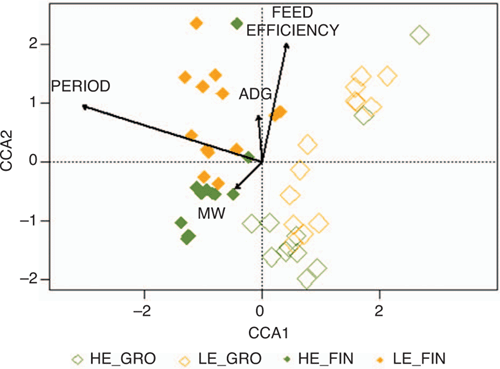
Beta biodiversity is graphically represented in Fig. 1, as well as the effects of explanatory variables included in the model, namely, FE, period, and both MW and ADG coefficients in the RFI model. Samples are clearly clustered by period and FE, with the effects of MW and ADG coefficients being less graphically evident. Adonis test results confirmed the foreseen differences in ruminal microbiota composition when comparing sampling periods (GRO vs FIN, P < 0.001), FE categories (HE vs LE, P = 0.022) and MW coefficient values (P = 0.021), but not in the case of ADG coefficient values (P = 0.276). Statistical differences in abundance of genera between FE categories (HE vs LE) could not be detected by ALDEx analysis, regardless of the sampling period (Fig. S1, available as Supplementary Material to this paper).
|
Microbial network
Microbial networks were built to test interactions among bacterial and archaeal genera (Fig. 2). Degree of interaction was studied through the number of genera (nodes) that established significant interactions (edges) with other genera, as well as the number of interactions established per node (node degree). During the growing period, HE bulls had similar number of nodes taking part in the microbial network as did LE bulls (26 in HE vs 24 in LE), but higher number of edges (57 in HE vs 28 in LE) and a higher average node degree (4.38 in HE vs 2.33 in LE). During the finishing period, microbial network architecture changed; HE bulls continued to have more correlating nodes than did LE bulls (30 in HE vs 21 in LE), but LE animals drastically increased their number of edges (53 in HE vs 59 in LE) and the node degree (3.53 in HE vs 5.62 in LE).

|
Moreover, we investigated microbial genera that act as main information gateways in networks in terms of betweenness centrality, i.e. the extent to which one node lies on paths that connect other nodes. Networks of HE animals presented higher betweenness centrality than did those of LE animals in growing (3.88 in HE vs 1.04 in LE) but not in finishing (2.37 in HE vs 2.52 in LE) period.
Microbial functional capability
After predicting functional content of ruminal microbiota, two pathways were found to be differentially expressed depending on the animal FE phenotype. In the growing period, ABC transporter pathway (ATP-dependent transport of molecules across cell membrane) was more active in HE animals than in LE animals, and, in the finishing period, methane metabolism was downregulated in the rumen of HE individuals when compared with LE animals.
Discussion
RFI and mechanisms underlying the variability of FE
Variations in RFI occur due to potential physiological mechanisms such as digestion, fermentation and metabolism (Herd and Arthur 2009). Our findings showed that HE animals apparently digested more feed, in terms of DM, OM and CP, than did LE ones. These results are in accordance with previous studies in which more efficient animals showed a higher nutrient digestibility and a smaller nutrient loss through waste and methane emission (Richardson et al. 1996; Nkrumah et al. 2006). Negesse et al. (2017) also observed improved apparent digestibility coefficients of DM, OM and CP in HE heifers; these animals excreted a smaller proportion of N through faeces and their N biological value (digestible N ratio) was higher than that of less efficient heifers, suggesting that CP digestion and metabolism may be enhanced in HE animals. In comparison, de Assis Lage et al. (2019) did not find differences in digestibility coefficients of such nutrients but they reported a tendency of HE heifers to better digest ether extract fraction.
Volatile fatty acids are products of rumen microbial fermentation of carbohydrates, constituting the main energy source for ruminants (Bergman 1990). Although differences between HE and LE bulls did not reach significance for any ruminal fermentation parameter, numerical values indicated that LE animals had a fermentation pattern oriented towards the production of higher molar proportion of acetic and less propionic acids than did HE animals, with the consequent effect on acetate-to-propionate ratio. These observed differences in rumen fermentation pattern may be playing a role in the bulls’ FE phenotype, since metabolic hydrogen produced in the first step of acetic acid pathway is later taken up by methanogens, increasing energy loss through gas emissions (Ungerfeld 2020).
During the growing period, molar proportions of the major VFA were similar to those observed by Yuste et al. (2020) in beef heifers fed a similar ad libitum concentrate plus straw diet. However, during the finishing period, the higher amount of total VFA, concomitant with significantly lower ruminal pH, were rooted in the increased DM intake and, consequently, in the higher extent of fermentation process. However, propionic acid showed a different trend and it was higher in younger animals. Hernandez-Urdaneta et al. (1976) reported that forage-to-concentrate ratio affects the molar proportions of VFA, which for high-concentrate diets changes towards decreased acetic and increased propionic acid; therefore, in our experiment, the lower forage-to-concentrate ratio during the growing (11%) than in the finishing period (18%) can explain the observed decreased proportion of propionic acid with time.
RFI and ruminal microbial community
In the present study, Illumina sequencing technology was used to analyse bacterial and archaeal composition, biodiversity, connectance and functional capability within the rumen of intensively reared bulls differing in their FE.
A negative correlation between ruminal microbial α biodiversity and FE has been previously described in milking cows (Shabat et al. 2016), suggesting that efficient microbiotas are less complex but more specialised in providing higher concentrations of relevant output metabolites that can be used to meet host’s energy requirements. In a similar manner, our results showed that microbial α diversity values significantly increased with time only in LE bulls. Microbial diversity is also positively correlated with community stability and robustness, as both differential response to variable conditions and functional redundancy of species are enhanced (McCann 2000; Moya and Ferrer 2016). Thus, it seems reasonable to hypothesise that α biodiversity of the microbiota has positive and negative coexisting effects on ecosystem robustness and feed utilisation efficiency, respectively. The fact that LE bulls considerably increased their genera network connectance with time, while HE bulls kept it constant or even diminished it, supports the hypothesis that LE animals’ ruminal microbiota could be more robust and have an enhanced ability to cope with possible disturbances (Dunne et al. 2002).
Even though β diversity representation showed clear clustering of bulls’ microbial community, no statistical differences in the abundance of main genera could be found between FE categories. Considering that some studies have had success in reporting a relationship between certain microbial taxa and animal’s FE (McCann et al. 2014; Myer et al. 2015; Perea et al. 2017; Delgado et al. 2019), we consider that the following factors could hinder detection of such relationship: (1) there can be substantial animal-to-animal variation in the rumen microbial community, thus requiring a greater number of animals to be able to observe a significant association between microbial taxa and FE (Brulc et al. 2009; Weimer et al. 2010), and (2) the lack of differences observed between FE categories at the main-genera level may indicate that the important variation in microbial communities lies at a finer resolution (e.g. at species level or low-abundance genera).
Kittelmann et al. (2014) described the existence of three ruminal microbial communities linked to different methane yields in sheep; ruminotype H was characterised by the highest methane emissions and harboured the higher abundance of species belonging to Ruminococcus, other Ruminococcaceae, Lachnospiraceae, Catabacteriaceae, Coprococcus, other Clostridiales, Prevotella, other Bacteroidales and Alphaproteobacteria. In a recent study in sheep, Ghanbari Maman et al. (2020) also identified certain genes from Lachnospiraceae, Ruminococcus, Butyrivibrio and Selenomonas taxa that can have significant effects on methane production pathway. In accordance with these studies, our co-abundance analysis showed that certain genera previously related to high methane emission (e.g. Methanobrevibacter, Roseburia, Agathobacter, Butyrivibrio, Pseudobutyrivibrio, Ruminococcus, Selenomonas) either were more central or evolved to be more central in LE animal networks during the transition from growing to finishing periods (Tables S2, Table S3, available as Supplementary Material to this paper), which could at least partially cause their lower FE.
Recent studies have highlighted a possible relationship between microbial metabolic functions and the animal’s FE, but the nature of such relationship is still unclear. Li et al. (2016) observed that HE cattle had a more active metabolism of nucleotides, as well as of various energy-generating molecules (e.g. propanoate, glyoxylate and dicarboxylate, starch and sucrose), hypothesising that such increased metabolic activity could enhance feed digestion and provide the host with more nutrients. Li et al. (2016) and Elolimy et al. (2020) also reported that rumen microbiota of the most efficient cattle was more active in cell proliferation and survivability, inducing cellular growth and increasing tolerance to viral infection; likewise, our results showed enhanced cell membrane transport functions in HE growing animals. Finally, the observed decrease of methane metabolism activity in HE finishing bulls (Shabat et al. 2016) supports the previous idea that high and low methane emitters can have a similar abundance of ruminal methanogens but differential expression and transcription of methanogenesis pathway genes (Shi et al. 2014).
Conclusions
The exploration of the relationship between rumen microbial community and host FE showed increased nutrient digestibility in HE animals. Alpha biodiversity and genera network connectance increased with time in LE bulls, highlighting a possible trade-off between FE and ruminal microbiota robustness. Moreover, certain genera that have previously been related to high methane emission were more central in LE animals’ genera networks. Our results have provided evidence that the rumen microbiota could be one of the biological factors associated with variation in cattle FE.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
Special thanks are extended to J. R. Bertolín Pardos for their laboratory assistance. This study is a part of GenTORE project and received funding from the European Union H2020 program under grant agreement nº 727213 and Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RTA-14-038-C02). S. Costa-Roura is recipient of a research training grant from Spanish Ministry of Science and Innovation (FPU 2016/03761). A. R. Seradj contract is financed by GenTORE project.
References
Arthur JPF, Herd RM (2008) Residual feed intake in beef cattle. Revista Brasileira de Zootecnia 37, 269–279.| Residual feed intake in beef cattle.Crossref | GoogleScholarGoogle Scholar |
Arthur PF, Archer JA, Johnston DJ, Herd RM, Richardson EC, Parnell PF (2001) Genetic and phenotypic variance and covariance components for feed intake, feed efficiency, and other postweaning traits in Angus cattle. Journal of Animal Science 79, 2805–2811.
| Genetic and phenotypic variance and covariance components for feed intake, feed efficiency, and other postweaning traits in Angus cattle.Crossref | GoogleScholarGoogle Scholar | 11768108PubMed |
Bergman EN (1990) Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiological Reviews 70, 567–590.
| Energy contributions of volatile fatty acids from the gastrointestinal tract in various species.Crossref | GoogleScholarGoogle Scholar | 2181501PubMed |
Brulc JM, Antonopoulos DA, Berg Miller ME, Wilson MK, Yannarell AC, Dinsdale EA, Edwards RE, Frank ED, Emerson JB, Wacklin P, Coutinho PM, Henrissat B, Nelson KE, White BA (2009) Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proceedings of the National Academy of Sciences of the United States of America 106, 1948–1953.
| Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases.Crossref | GoogleScholarGoogle Scholar | 19181843PubMed |
Calle ML (2019) Statistical analysis of metagenomics data. Genomics & Informatics 17, e6
| Statistical analysis of metagenomics data.Crossref | GoogleScholarGoogle Scholar |
Costa-Roura S, Balcells J, de la Fuente G, Mora-Gil J, Llanes N, Villalba D (2020) Nutrient utilization efficiency, ruminal fermentation and microbial community in Holstein bulls fed concentrate-based diets with different forage source. Animal Feed Science and Technology 269, 114662
| Nutrient utilization efficiency, ruminal fermentation and microbial community in Holstein bulls fed concentrate-based diets with different forage source.Crossref | GoogleScholarGoogle Scholar |
de Assis Lage CF, Gesteira Coelho S, Diniz Neto H do C, Rocha Malacco VM, Pacheco Rodrigues JP, Sacramento JP, Samarini Machado F, Ribeiro Pereira LG, Ribeiro Tomich T, Magalhães Campos M (2019) Relationship between feed efficiency indexes and performance, body measurements, digestibility, energy partitioning, and nitrogen partitioning in pre-weaning dairy heifers. PLoS One 14, e0223368
| Relationship between feed efficiency indexes and performance, body measurements, digestibility, energy partitioning, and nitrogen partitioning in pre-weaning dairy heifers.Crossref | GoogleScholarGoogle Scholar | 31600254PubMed |
Delgado B, Bach A, Guasch I, González C, Elcoso G, Pryce JE, Gonzalez-Recio O (2019) Whole rumen metagenome sequencing allows classifying and predicting feed efficiency and intake levels in cattle. Scientific Reports 9, 11
| Whole rumen metagenome sequencing allows classifying and predicting feed efficiency and intake levels in cattle.Crossref | GoogleScholarGoogle Scholar | 30626904PubMed |
Dunne JA, Williams RJ, Martinez ND (2002) Network structure and biodiversity loss in food webs: robustness increases with connectance. Ecology Letters 5, 558–567.
| Network structure and biodiversity loss in food webs: robustness increases with connectance.Crossref | GoogleScholarGoogle Scholar |
Elolimy A, Alharthi A, Zeineldin M, Parys C, Loor JJ (2020) Residual feed intake divergence during the preweaning period is associated with unique hindgut microbiome and metabolome profiles in neonatal Holstein heifer calves. Journal of Animal Science and Biotechnology 11, 13
| Residual feed intake divergence during the preweaning period is associated with unique hindgut microbiome and metabolome profiles in neonatal Holstein heifer calves.Crossref | GoogleScholarGoogle Scholar | 31988748PubMed |
Fernandes AD, Macklaim JM, Linn TG, Reid G, Gloor GB (2013) ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-seq. PLoS One 8, e67019
| ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-seq.Crossref | GoogleScholarGoogle Scholar | 23843979PubMed |
Friedman J, Alm EJ (2012) Inferring correlation networks from genomic survey data. PLoS Computational Biology 8, e1002687
| Inferring correlation networks from genomic survey data.Crossref | GoogleScholarGoogle Scholar | 23028285PubMed |
Ghanbari Maman L, Palizban F, Fallah Atanaki F, Elmi Ghiasi N, Ariaeenejad S, Ghaffari MR, Hosseini Salekdeh G, Kavousi K (2020) Co-abundance analysis reveals hidden players associated with high methane yield phenotype in sheep rumen microbiome. Scientific Reports 10, 4995
| Co-abundance analysis reveals hidden players associated with high methane yield phenotype in sheep rumen microbiome.Crossref | GoogleScholarGoogle Scholar | 32193482PubMed |
Gloor GB, Macklaim JM, Pawlowsky-Glahn V, Egozcue JJ (2017) Microbiome datasets are compositional: and this is not optional. Frontiers in Microbiology 8, 2224
| Microbiome datasets are compositional: and this is not optional.Crossref | GoogleScholarGoogle Scholar | 29187837PubMed |
Herd RM, Arthur PF (2009) Physiological basis for residual feed intake. Journal of Animal Science 87, E64–E71.
| Physiological basis for residual feed intake.Crossref | GoogleScholarGoogle Scholar | 19028857PubMed |
Hernandez-Urdaneta A, Coppock CE, McDowell RE, Gianola D, Smith NE (1976) Changes in forage-concentrate ratio of complete feeds for dairy cows. Journal of Dairy Science 59, 695–707.
| Changes in forage-concentrate ratio of complete feeds for dairy cows.Crossref | GoogleScholarGoogle Scholar |
Huntington G (1990) Energy metabolism in the digestive tract and liver of cattle: influence of physiological state and nutrition. Reproduction, Nutrition, Development 30, 35–47.
| Energy metabolism in the digestive tract and liver of cattle: influence of physiological state and nutrition.Crossref | GoogleScholarGoogle Scholar | 2184823PubMed |
Kittelmann S, Pinares-Patiño CS, Seedorf H, Kirk MR, Ganesh S, McEwan JC, Janssen PH (2014) Two different bacterial community types are linked with the low-methane emission trait in sheep. PLoS One 9, e103171
| Two different bacterial community types are linked with the low-methane emission trait in sheep.Crossref | GoogleScholarGoogle Scholar | 25383707PubMed |
Li F, Zhou M, Ominski K, Guan LL (2016) Does the rumen microbiome play a role in feed efficiency of beef cattle? Journal of Animal Science 94, 44–48.
| Does the rumen microbiome play a role in feed efficiency of beef cattle?Crossref | GoogleScholarGoogle Scholar |
McCann KS (2000) The diversity–stability debate. Nature 405, 228–233.
| The diversity–stability debate.Crossref | GoogleScholarGoogle Scholar | 10821283PubMed |
McCann JC, Wiley LM, Forbes TD, Rouquette FM, Tedeschi LO (2014) Relationship between the rumen microbiome and residual feed intake-efficiency of Brahman bulls stocked on bermudagrass pastures. PLoS One 9, e91864
| Relationship between the rumen microbiome and residual feed intake-efficiency of Brahman bulls stocked on bermudagrass pastures.Crossref | GoogleScholarGoogle Scholar | 24642871PubMed |
Moya A, Ferrer M (2016) Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends in Microbiology 24, 402–413.
| Functional redundancy-induced stability of gut microbiota subjected to disturbance.Crossref | GoogleScholarGoogle Scholar | 26996765PubMed |
Myer PR, Smith TPL, Wells JE, Kuehn LA, Freetly HC (2015) Rumen microbiome from steers differing in feed efficiency. PLoS One 10, e0129174
| Rumen microbiome from steers differing in feed efficiency.Crossref | GoogleScholarGoogle Scholar | 26030887PubMed |
Negesse T, Datt C, Kundu SS (2017) Residual feed intake, digestibility of nutrients and efficiency of water utilizations in Murrah buffalo heifers. Journal of Dairy, Veterinary & Animal Research 5, 74–80.
| Residual feed intake, digestibility of nutrients and efficiency of water utilizations in Murrah buffalo heifers.Crossref | GoogleScholarGoogle Scholar |
Nkrumah JD, Okine EK, Mathison GW, Schmid K, Li C, Basarab JA, Price MA, Wang Z, Moore SS (2006) Relationships of feedlot feed efficiency, performance, and feeding behavior with metabolic rate, methane production, and energy partitioning in beef cattle. Journal of Animal Science 84, 145–153.
| Relationships of feedlot feed efficiency, performance, and feeding behavior with metabolic rate, methane production, and energy partitioning in beef cattle.Crossref | GoogleScholarGoogle Scholar | 16361501PubMed |
Owens FN, Hanson CF (1992) External and internal markers for appraising site and extent of digestion in ruminants. Journal of Dairy Science 75, 2605–2617.
| External and internal markers for appraising site and extent of digestion in ruminants.Crossref | GoogleScholarGoogle Scholar | 1452861PubMed |
Perea K, Perz K, Olivo SK, Williams A, Lachman M, Ishaq SL, Thomson J, Yeoman CJ (2017) Feed efficiency phenotypes in lambs involve changes in ruminal, colonic, and small-intestine-located microbiota. Journal of Animal Science 95, 2585–2592.
| Feed efficiency phenotypes in lambs involve changes in ruminal, colonic, and small-intestine-located microbiota.Crossref | GoogleScholarGoogle Scholar | 28727071PubMed |
R Core Team (2020) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria)
Richardson EC, Herd RM, Arthur PF, Wright J, Xu G, Dibley K, Oddy VH (1996) Possible physiological indicators for net feed conversion efficiency in beef cattle. Proceedings of the Australian Society of Animal Production 21, 103–106.
Savietto D, Berry DP, Friggens NC (2014) Towards an improved estimation of the biological components of residual feed intake in growing cattle. Journal of Animal Science 92, 467–476.
| Towards an improved estimation of the biological components of residual feed intake in growing cattle.Crossref | GoogleScholarGoogle Scholar | 24664556PubMed |
Schenkel FS, Miller SP, Wilton JW (2004) Genetic parameters and breed differences for feed efficiency, growth, and body composition traits of young beef bulls. Canadian Journal of Animal Science 84, 177–185.
| Genetic parameters and breed differences for feed efficiency, growth, and body composition traits of young beef bulls.Crossref | GoogleScholarGoogle Scholar |
Shabat SKB, Sasson G, Doron-Faigenboim A, Durman T, Yaacoby S, Berg Miller ME, White BA, Shterzer N, Mizrahi I (2016) Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. The ISME Journal 10, 2958–2972.
| Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants.Crossref | GoogleScholarGoogle Scholar |
Shi W, Moon CD, Leahy SC, Kang D, Froula J, Kittelmann S, Fan C, Deutsch S, Gagic D, Seedorf H, Kelly WJ, Atua R, Sang C, Soni P, Li D, Pinares-Patiño CS, McEwan JC, Janssen PH, Chen F, Visel A, Wang Z, Attwood GT, Rubin EM (2014) Methane yield phenotypes linked to differential gene expression in the sheep rumen microbiome. Genome Research 24, 1517–1525.
| Methane yield phenotypes linked to differential gene expression in the sheep rumen microbiome.Crossref | GoogleScholarGoogle Scholar | 24907284PubMed |
Tsilimigras MCB, Fodor AA (2016) Compositional data analysis of the microbiome: fundamentals, tools, and challenges. Annals of Epidemiology 26, 330–335.
| Compositional data analysis of the microbiome: fundamentals, tools, and challenges.Crossref | GoogleScholarGoogle Scholar |
Ungerfeld EM (2020) Metabolic hydrogen flows in rumen fermentation: principles and possibilities of interventions. Frontiers in Microbiology 11, 589
| Metabolic hydrogen flows in rumen fermentation: principles and possibilities of interventions.Crossref | GoogleScholarGoogle Scholar | 32351469PubMed |
Weimer PJ, Stevenson DM, Mantovani HC, Man SLC (2010) Host specificity of the ruminal bacterial community in the dairy cow following near-total exchange of ruminal contents. Journal of Dairy Science 93, 5902–5912.
| Host specificity of the ruminal bacterial community in the dairy cow following near-total exchange of ruminal contents.Crossref | GoogleScholarGoogle Scholar | 21094763PubMed |
Woloszynek S, Mell JC, Zhao Z, Simpson G, O’Connor MP, Rosen GL (2019) Exploring thematic structure and predicted functionality of 16S rRNA amplicon data. PLoS One 14, e0219235
| Exploring thematic structure and predicted functionality of 16S rRNA amplicon data.Crossref | GoogleScholarGoogle Scholar | 31825995PubMed |
Yuste S, Amanzougarene Z, de Vega A, Fondevila M, Blanco M, Casasús I (2020) Effect of preweaning diet on performance, blood metabolites and rumen fermentation around weaning in calves of two beef breeds. Animal Production Science 60, 1018–1027.
| Effect of preweaning diet on performance, blood metabolites and rumen fermentation around weaning in calves of two beef breeds.Crossref | GoogleScholarGoogle Scholar |