

Effects of silage to concentrate ratio and duration of feeding on the fatty acid composition of ovine muscle and adipose tissue
R. S. Gravador A , A. G. Fahey A , S. M. Harrison A , V. Gkarane A B , A. P. Moloney C , N. P. Brunton A , N. A. Claffey A D , M. G. Diskin D , L. J. Farmer E , P. Allen B and F. J. Monahan A *
A , V. Gkarane A B , A. P. Moloney C , N. P. Brunton A , N. A. Claffey A D , M. G. Diskin D , L. J. Farmer E , P. Allen B and F. J. Monahan A *
A School of Agriculture and Food Science, University College Dublin, Belfield, Dublin 4, Ireland.
B Teagasc Food Research Centre, Ashtown, Dublin 15, Ireland.
C Teagasc, Animal & Grassland Research and Innovation Centre, Grange, Dunsany, Co. Meath, Ireland.
D Teagasc, Animal & Grassland Research and Innovation Centre, Athenry, Co. Galway, Ireland.
E Agri-Food and Biosciences Institute, Newforge Lane, Belfast, BT9 5PX, UK.
Animal Production Science 62(7) 682-690 https://doi.org/10.1071/AN20301
Submitted: 16 May 2020 Accepted: 2 February 2022 Published: 8 March 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Dietary strategies may influence the fatty acid composition of sheep meat and ultimately its nutritional and sensory quality.
Aim: This study investigated the effects of silage to concentrate ratio and feeding duration on the concentration of intramuscular fatty acids and branched chain fatty acid in subcutaneous adipose tissue of lambs.
Methods: Pasture fed lambs were assigned in groups of 11 to one of nine dietary treatments consisting of silage to cereal concentrate at ratios (dry matter basis) of 100:0 (silage), 50:50 (silage:concentrate) or 0:100 (concentrate) for three feeding durations (36, 54 or 72 days) pre-slaughter. A 10th group of 11 lambs remained at pasture until slaughter. Fatty acid methyl esters were prepared using a microwave-assisted method and quantified by gas chromatography with flame ionisation detection.
Results: The concentration of n-3 polyunsaturated fatty acids (PUFA) and proportions of C18:3n-3 and C18:2 cis-9 trans-11 decreased for mixed feed and/or concentrate treatments, but not for silage, from 36 to 72 days. The C18:2n-6 and n-6 PUFA concentrations increased for mixed feed and concentrate, but not for silage, from 36 to 72 days. The animals that remained at pasture had lower C18:2n-6 and n-6 PUFA than those receiving mixed feed for 72 days, concentrate for 54 days and concentrate for 72 days, but higher C18:3n-3, C18:2 cis-9 trans-11 and n-3 PUFA concentrations than the indoor finishing groups. The n-6/n-3 PUFA ratio was lower in pastured lambs compared to the other groups. 4-Methyloctanoic acid and 4-ethyloctanoic acid were higher in concentrate and mixed feed treatments than in silage while 4-methylnonanoic acid increased with increase in concentrate level in the diet.
Conclusion: The ratio of silage to concentrate in the diet and duration of feeding pre-slaughter influence the fatty acid composition of muscle and subcutaneous adipose tissue of lambs.
Implications: The results are important for lamb producers in understanding the nutritional and sensory implications of altering lamb diets.
Keywords: biohydrogenation, branched chain fatty acids, CLA, concentrate-fed, intramuscular fatty acids, lamb meat, PUFA, silage, subcutaneous adipose tissue.
Introduction
The fatty acid composition of ruminant tissues is known to be influenced by the animal’s diet (Aurousseau et al. 2007; Wood et al. 2008) and this can have important implications for the content of nutritionally important fatty acids in tissues, such as muscle, which are consumed by humans as food. For example, grass and grass silage are rich sources of α-linolenic acid (C18:3n-3) and when consumed by ruminants can result in a lower and more nutritionally favourable n-6/n-3 polyunsaturated (PUFA) ratio in muscle (French et al. 2000; Warren et al. 2008; Scerra et al. 2011; Mezgebo et al. 2017). Furthermore, compared to concentrate-fed animals, ruminant muscle from grass-fed animals contains elevated conjugated linoleic acid (CLA) and lower saturated fatty acids (SFA) (Nuernberg et al. 2005), both characteristics favourable from a human health perspective (Daley et al. 2010).
From a production point of view, the use of more energy dense cereal concentrates as feed for lambs can permit animals to grow faster, thus attaining the desired slaughter weight at a younger age in comparison to pasture-fed lambs. Therefore, the utilisation of concentrate-based diets in lamb production is common, especially in countries where fresh grass is not available all year round. Furthermore, a study by Font i Furnols et al. (2009) concluded that European consumers (Spain, Germany, UK and France), preferred lamb from animals fed on cereal concentrate, alone or in combination with pasture, compared to lamb from animals fed pasture only. However, from a nutritional perspective, it may be possible for lambs fed grass at pasture prior to shifting to a concentrate-based diet pre-slaughter, to retain the desirable PUFA n-6/n-3 ratio and fatty acid profile imparted by the grass feeding. It was previously reported that feeding concentrates for a short duration after a period at pasture did not negate the beneficial effects of grass on muscle fatty acid profile in ruminants (Scerra et al. 2014; Mezgebo et al. 2017).
Considering the impact of dietary factors on both the nutritional and sensorial quality of meats, in the current study, lambs fed pasture from birth were finished on three pre-slaughter diets consisting of concentrate and/or silage for three different durations and the effect on muscle and subcutaneous adipose tissue (SAT) fatty acid composition was investigated. It was hypothesised that finishing lambs on a concentrate diet alone for a short duration, or a concentrate diet plus silage or silage alone for a long duration, would yield muscle and SAT fatty acid compositions that are comparable to pasture-fed animals.
Materials and methods
Animal management
One hundred and ten ram lambs (Texel × Scottish Blackface) were sourced from Irish farms in March 2015 (Claffey et al. 2018; Gkarane et al. 2019). Lambs were raised at pasture from birth (March 2015) and were weaned at 130 days of age after which they were transported to the Teagasc Animal & Grassland Research Centre, Athenry, Co. Galway, Ireland. Ninety-nine lambs were maintained at grass pasture (for 66–81 days, average 72 days) until selected for commencement of an intensive indoor finishing period. Lambs were allocated in groups of 11 to one of nine dietary treatments consisting of three grass silage:concentrate ratios (dry matter basis) (100:0 (S), 50:50 (SC) and 0:100 (C)) with each diet being fed for three pre-slaughter feeding durations (36, 54 and 72 days) to give the following dietary treatments: S36, S54, S72, SC36, SC54, SC72, C36, C54, C72. For the indoor finishing period (36, 54 or 72 days) lambs were individually penned in metal floor feeding pens (182 cm × 122 cm). A 10th group of 11 lambs (Pasture treatment) was raised at pasture from birth to slaughter. To maintain commercial relevance, the grazing animals were managed so that they had access to abundant grass, minimising competition for resources and allowing animals to express individual behaviour and maintain independence. The concentrate diet consisted of 30% maize, 30% barley, 16.5% soya hulls, 15.5% soybean meal, 5% molasses and 3% minerals. In line with commercial practice, lambs were selected for treatment based on liveweight at the start of the indoor finishing period and predicted growth rate on the assigned diets to yield lambs with similar weights at slaughter (Claffey et al. 2018). Thus, the lightest lambs were assigned to the C72 treatment and the heaviest to the S36 treatment. At the end of the finishing period, lambs were transported to a commercial abattoir (Gillivan’s, Moate, Co. Westmeath) for slaughter. The mean ages (in days) of the animals at slaughter were 252, 260, 273, 248, 254, 271, 248, 258, 266 for the S36, S54, S72, SC36, SC54, SC72, C36, C54, C72 treatments, respectively, and 273 for the P treatment. The mean slaughter weights (in kg) were 43.9, 45.9, 46.3, 48.3, 48.4, 48.9, 50.8, 48.4, 48.7 for the S36, S54, S72, SC36, SC54, SC72, C36, C54, C72 treatments, respectively, and 53.7 for the P treatment.
After slaughter, carcasses were chilled overnight and transported to Teagasc, Food Research Centre, Ashtown, Dublin 15, Ireland for dissection. Longissimus thoracis et lumborum (LTL) muscle was excised from each carcass, cut into 2.5 cm thick steaks, vacuum packed, aged for 8 days at 4°C and frozen at −20°C until required for analysis.
All animal procedures used in this study were conducted under experimental licence from the Irish Health Products Regulatory Authority (HPRA) in accordance with the European Union Protection of animals used for scientific purposes regulations 2012 (S.I. No. 543 of 2012).
Chemicals
BCFA standards (4-methyloctanoic acid (MOA), 4-ethyloctanoic acid (EOA) and 4-methylnonanoic acid (MNA)), a 37 fatty acid methyl ester (FAME) standard (Supelco) and internal standard C23:0 free fatty acid (IS) (Analytical standard grade), chloroform (high performance liquid chromatography (HPLC) grade) and acetyl chloride (Reagent grade) were purchased from Sigma-Aldrich (Darmstadt, Germany). Potassium hydroxide (KOH; Analytical reagent grade) was purchased from VWR Chemicals (Leuven, Belgium), and methanol (HPLC grade), pentane (Analytical grade), sodium chloride (NaCl; Laboratory reagent grade), and anhydrous sodium sulfate (Na2SO4; Laboratory reagent grade) were purchased from Fisher Scientific (Loughborough, UK). Deionised water was obtained from a Millipore Elix 15 water purification system (Merck Millipore, Darmstadt, Germany).
Feed and muscle fatty acid analysis
The microwave-assisted extraction of total fatty acids and subsequent conversion to FAME was carried out based on the method of Brunton et al. (2015). A 1.5 g portion of feed was used for the feed fatty acid analysis, while a 1 g portion of LTL was used for the muscle fatty acids analysis. The reaction vessel used was a perfluoroalkoxy alkane (PFA) and reactions were carried out in a MARS 6 Express 40 position Microwave Reaction System (CEM Corporation, Matthews, NC, USA). Internal standard (100 μL of 10 mg/mL C23:0) was added to the sample in each reaction vessel. For the saponification, 2.5% KOH in methanol was used, while for the esterification 5% acetyl chloride in methanol was utilised. The FAME were partitioned by pentane and, to aid the separation, saturated NaCl was added. The pentane layer was transferred into a gas chromatograph (GC) vial containing Na2SO4.
The separation of FAME was performed on a PerkinElmer Clarus 580 GC fitted with a flame ionisation detection detector (PerkinElmer, Massachusetts, USA), using a CP-Sil 88 capillary column (100 m length, 0.25 mm internal diameter, 0.2 μm film thickness (Chromapack, The Netherlands). The carrier gas was hydrogen at a constant flow rate of 1.25 mL/min. The injection volume was 0.5 μL at a 10:1 split ratio. The temperatures of the injector and detector were 250°C and 270°C, respectively. The oven temperature was set at 80°C and was increased at 6.2°C/min to 220°C and held for 3.2 min. It was further increased at 6.3°C/min to 240°C and held for 3.2 min. Compounds were identified by comparing their retention times with those of the 37 FAME standard. Results for each FAME were expressed as concentration (mg/100 g muscle) and proportion of the total fatty acids (g fatty acid/100 g total fatty acids).
Subcutaneous adipose tissue branched chain fatty acid analysis
The microwave-assisted extraction described above was used for the preparation BCFA FAME. The detailed procedure is described in Gravador et al. (2019). Results were expressed as the absolute concentration of total BCFA in SAT (μg BCFA/g fat).
Statistical analyses
All data were analysed using SAS v.9.4. Data diagnostics for normality was performed on the fatty acid data (expressed as a concentration in mg/100 g muscle and as a proportion in g/100 g total fatty acids) and the BCFA data (expressed in μg/g SAT) and, for data where the residuals were not normally distributed, a Box–Cox transformation in PROC TRANSREG was used to determine the appropriate lambda value for the transformation (Fahey et al. 2007). Following this, ANOVA, using lamb as the experimental unit, was performed using a mixed model (PROC MIXED) where pre-slaughter diets (S, SC and C), feeding duration (36, 54 and 72 days) and their interactions were considered as fixed effects and individual animal as a random effect. To compare indoor finishing treatments with the P treatment a mixed model (PROC MIXED) was used, where diet was considered as the fixed effect and individual animal as a random effect. A Bonferroni correction was used to adjust the P-values for multiple comparisons. A difference was deemed to be significant when P < 0.05.
Results and discussion
Composition of the diets
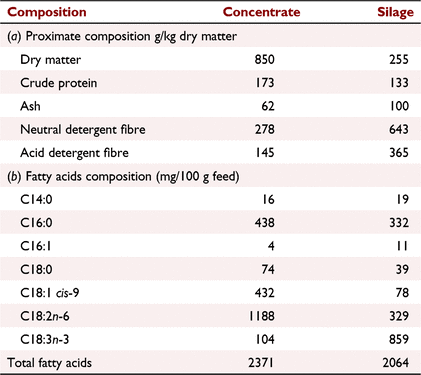
The concentrate had lower C18:3n-3 but higher C18:1 cis-9, C18:2n-6 and total fatty acid concentrations than the silage (Table 1), similar to previous studies (French et al. 2000; Warren et al. 2008; Mezgebo et al. 2017), although the fatty acid concentration of concentrates may vary depending on their formulation. Although grass was not analysed in this study, French et al. (2000) and Warren et al. (2008) reported that grass had a lower proportion of SFA and a higher proportion of unsaturated fatty acids than grass silage, differences that have been attributed to wilting of grass prior to ensiling (Dewhurst and King 1998).
|
Total fatty acids in muscle
The silage to concentrate ratio in the diet affected the total fatty acid concentration (TFA, mg/100 g muscle) in muscle of lambs (P = 0.023), where C (2128) was higher than S (1737) (Table 2). The higher muscle TFA in C was mainly due to higher concentrations of C18:1 cis-9, C18:2n-6, total monosaturated fatty acids (MUFA) and n-6 PUFA, compared to S. Others have also reported higher total lipids and fatty acids in lambs raised at pasture but finished on a concentrate diet compared to lambs finished on pasture alone (Aurousseau et al. 2004, 2007). However, a potential confounding effect of the liveweight of the animals at the start of the indoor finishing period is acknowledged, whereby the lightest animals (with potentially lower intramuscular fat (IMF)) were assigned to the C treatments.

|
Saturated fatty acids in muscle
The proportion of total SFA (g/100 g total fatty acids) in muscle of lambs was influenced by the diets, where S (47.8) and SC (47.5) had higher proportions than C (45.7) (P = 0.004) (Supplementary Table S1). This was a consequence of the higher proportions of C12:0, C14:0, C15:0, and C18:0 in muscle of S and SC compared to C.
An effect of diet on the proportion (g/100 g) of C18:0 was observed, whereby C (16.65) had a lower proportion than SC (18.2) and S (17.5) (P < 0.003; Table S1). This observation could be attributed to the high carbohydrate-containing diet (diet C) having a reduced residence time in the rumen (Demirel et al. 2006), and to the fact that the rumen of these animals is less favourable for the biohydrogenation by cellulolytic microorganisms than the rumen of herbage-fed lambs (S or SC) (Vasta et al. 2009).
There was a significant diet by duration interaction for the proportions of C12:0, C14:0 and C15:0 (P < 0.05; Table S1). The C12:0 decreased with feeding duration especially in SC and C. For C14:0 at 54 days and 72 days, C and SC were lower than S. Moreover, the proportion of C14:0 was lower in C72 and higher in S72 than Pasture (Table S1). Nuernberg et al. (2005) also reported a higher proportion of C14:0 in muscle of lambs fed pasture than of lambs fed concentrate.
Monounsaturated fatty acids in muscle
The diets of lambs had an effect on the concentration and proportion of C18:1 cis-9 (P < 0.05), where values followed the order: C (889 mg/100 g and 41.7 g/100 g) > SC (807 mg/100 g, 39.6 g/100 g) > S (661 mg/100 g, 37.8 g/100 g) (Table 2, Table S1). Similar effects were observed for the concentration and proportion of C16:1 (P < 0.05), where the concentrations (mg/100 g) were: S (26.7) < SC (28.5) < C (33.7), and the proportions (g/100 g) were: SC (1.43) < S (1.55) < C (1.60).
The increase in C18:1 cis-9 and C16:1 could be attributed to enhancement, by concentrate feeding, of the activity of stearoyl-CoA desaturase (SCD or Δ-9 desaturase), which catalyses the conversion of C16:0 and C18:0 to C16:1 and C18:1, respectively. In support of this, Daniel et al. (2004) reported an increase in the gene expression of SCD, while Boughalmi and Araba (2016) observed an increased activity of SCD in response to feeding a concentrate-based diet. In the latter study, semimembranosus muscle of lambs fed concentrate had a higher proportion of C18:1 cis-9 than of lambs fed pasture plus concentrate or pasture alone (Boughalmi and Araba 2016).
The concentrations of C16:1 and C18:1 cis-9, and proportion of C18:1 cis-9 increased with feeding duration (P < 0.05). Between 36 and 72 days, a significant increase in the concentration of C18:1 cis-9 was observed in C (322) and SC (244) but not in S (26) (Table 2). A similar increase between 36 and 72 days was observed for the proportion of C18:1 cis-9 in groups SC and C. Since MUFA was dominated by C18:1 cis-9 (over 37%; Table S1), similar effects of diets and feeding duration on the concentration and proportion of total MUFA were observed (P < 0.05; Table 2 and Table S1). The proportion of total MUFA was higher in C72 than Pasture (P < 0.05; Table S1).
Although the lambs were all raised at pasture before assigning to pre-slaughter groups, the muscle fatty acid profile reflected the fatty acid profile of their respective pre-slaughter diets, especially when fed for the longest duration. Aurousseau et al. (2007) reported a higher concentration of C18:1 cis-9 in muscle of lambs raised at pasture and finished for long periods on a concentrate diet compared to grass-fed lambs. Moreover, Mezgebo et al. (2017) reported a higher muscle concentration of C18:1 cis-9 and total MUFA in bulls fed concentrate in comparison to bulls fed diets containing pasture and/or silage plus concentrate. The concentration and proportion of C18:1 trans-9 were also affected by the dietary treatments (P < 0.001), being lower in C (2.17 mg/100 g, 0.12 g/100 g) than SC (7.05 mg/100 g, 0.36 g/100 g) and S (6.55 mg/100 g, 0.42 g/100 g). This is consistent with Aurousseau et al. (2007) who found that concentrate feeding resulted lower muscle levels of trans-MUFA.
Polyunsaturated fatty acids in muscle
n-6 polyunsaturated fatty acids
There was a significant diet by duration interaction for the concentration of n-6 PUFA and C18:2n-6 (P < 0.01; Table 2). Between 36 and 72 days, the levels increased in SC and C but not in S. Additionally, after 54 days and 72 days, the n-6 PUFA and C18:2n-6 were higher in SC and C than S. When compared with Pasture, the concentration of C18:2n-6 was higher in SC72, C54 and C72 (P < 0.05; Table 2), while the concentration of n-6 PUFA was higher in all diets containing concentrates. There was a significant diet by duration interaction for the concentration of C20:3n-6 (P = 0.039) and C20:4n-6 (P = 0.006) (Table 2). The concentration of C20:3n-6 increased with increase in feeding duration in C but not in S and SC. Similarly, the concentration of C20:4n-6 increased with increase in feeding duration in C and SC but not in S. The proportion of C20:4n-6 also differed between dietary treatments (P = 0.033), where C had a lower proportion than S (1.56 vs 2.03 g/100 g) (Table S1).
The current results suggest that concentrates in the diet generally increase the concentration of n-6 fatty acids in lamb muscle, particularly when feeding takes place for longer durations. For instance, the muscle deposition of C18:2n-6, which was higher in the concentrate than the silage (Table 1), required at least 54 days to reach significance in the case of diet C but 72 days in the case of diet SC. In agreement with this, Aurousseau et al. (2007), Scerra et al. (2011) and Wang et al. (2015) reported an enhancement of C18:2n-6 in muscle of animals with prolonged finishing period on concentrate. In ruminant muscle the PUFA are almost exclusively deposited in the phospholipid fraction (Ponnampalam et al. 2001; Wood et al. 2004), and it was reported that elevated C18:2n-6 from concentrate sources enhances the synthesis of long chain (LC) n-6 PUFA, via desaturation and elongation, and their deposition in the membranal phospholipids (Warren et al. 2008). Therefore, the high content C18:2n-6 in diet C can explain the high muscle content of C20:3n-6, C20:4n-6 as well as of n-6 PUFA in lambs fed concentrate-containing diets. The results also show that the effect of desaturation and elongation was time dependent, i.e. increased with increasing time on the C and SC diets.
n-3 polyunsaturated fatty acids
There was a significant diet by duration interaction for the muscle concentration and proportion of n-3 PUFA and proportion of C18:3n-3 (P < 0.05; Table 2 and Table S1, respectively). Increased feeding duration led to decreases in the muscle levels of n-3 PUFA and C18:3n-3 in C and SC, but not in S. After 54 days and 72 days the levels of n-3 PUFA and C18:3n-3 were higher in S than SC and C. The effect of diets on C18:3n-3 showed a higher concentration (mg/100 g) in S (22.5) than either SC (19.3) or C (16.9) (P < 0.001; Table 2). Although the concentration of n-3 PUFA was higher in Pasture than in all the indoor finishing treatments (P < 0.001; Table 2), overall, S had a higher concentration than SC, which was higher than C (61.1 < 53.7 < 48.8 mg/100 g, respectively). This may be attributed to silage being derived from grass and therefore being more similar in composition to pasture than to concentrates.
It is known that the muscle concentration and proportion of 18:3n-3 is higher in grass-fed lambs in comparison to stall-fed lambs (Aurousseau et al. 2007). The current results showed that the different n-3 PUFA in muscle could be retained when lambs were finished on diets that contain silage regardless of the feeding duration. Conversely, n-3 PUFA in muscle were more likely reduced by finishing lambs on diets that contain concentrate especially at long durations. In agreement with the findings of Scerra et al. (2014), since silage contained higher C18:3n-3 than concentrate (Table 1), finishing the lambs on silage conserved the effect of pasture/grass, on which the animals were raised prior to assignment to the indoor diets, while finishing the lambs on concentrates attenuated the effect. According to Mezgebo et al. (2017), a shorter finishing duration on diets other than grass-based can lead to the retention of residual effects of grazed grass on fatty acid composition of muscle.
Dietary C18:3n-3 undergoes elongation and desaturation resulting in the synthesis of LC n-3 PUFA with deposition of C20:5n-3, C22:5n-3 and C22:6n-3 in the phospholipid fraction of muscle (Nuernberg et al. 2005; Warren et al. 2008). There was a significant diet by feeding duration interaction for the muscle concentration and proportion of C20:5n-3, C22:5n-3, and C22:6n-3 (P < 0.05; Table 2 and Table S1, respectively). Increased feeding duration resulted in decreased levels of C20:5n-3, C22:5n-3 and C22:6n-3 in C and SC, but not in S. At 36 days no effect of diets was observed on the concentration and/or proportion of these LC n-3 PUFA. Wang et al. (2015) also reported a higher level of C20:5n-3 in muscle of lambs that spent longer grazing times and supplemented with less concentrate on a diet, while Warren et al. (2008) reported a higher level of C22:6n-3 in phospholipid of cattle, which could be a result of long feeding periods in grass silage. The concentration and proportion of C18:3n-3 and the concentration of C22:6n-3 in Pasture was higher than in the other groups (P < 0.05; Table 2 and Table S1). The concentration of C20:5n-3 and proportion of C22:6n-3 were also higher in Pasture than in SC54, SC72, C54 and C72, while the concentration of C22:5n-3 and proportion of C20:5n-3 were higher in Pasture than SC72, C54 and C72 (P < 0.05; Table 2). Moreover, the proportion of C22:5n-3 was higher in Pasture than C72 (P < 0.05; Table S1).
Conjugated linoleic acid
The effect of diets on the concentration (mg/100 g) of C18:2 cis-9 trans-11 in muscle (P = 0.018; Table 2) showed that S (14.8) and SC (13.2), which did not differ, were higher than C (9.6), and at 72 days S72 was higher than C72. There was a significant diet by duration interaction observed for the proportion of C18:2 cis-9 trans-11 (P = 0.006; Table S1). There was a reduction in the proportion of C18:2 cis-9 trans-11 in SC and C but not in S with feeding duration, and at 36 days no effect of diet was found. An increase in muscle deposition of C18:2 cis-9 trans-11 was achieved by feeding animals diets such as fresh grass or grass silage (French et al. 2000; Daniel et al. 2004; Noci et al. 2005) or by decreasing the proportion of concentrate in the diet (French et al. 2000), in agreement with the current results. Additionally, muscle from Pasture-fed animals had a higher proportion of C18:2 cis-9 trans-11 than SC54 (Table S1), and concentration than SC72, C36, C54 or C72 (P < 0.05; Table 2).
Ratios of PUFA to SFA and n-6 PUFA to n-3 PUFA
There was a higher proportion of PUFA (g/100 g) in S (10.8) than SC (9.4) and C (8.8) (P < 0.001; Table S1), which could mainly be due to a higher n-3 PUFA in S. The S diets contain a higher concentration of C18:3n-3, which resulted in higher intakes of this fatty acid and n-3 PUFA by animals in this group (French et al. 2000; Realini et al. 2004). When compared with concentrate-fed animals, a higher quantity of n-3 fatty acids is deposited in the phospholipids and triacylglycerols of animals fed grass (Dannenberger et al. 2004).
The ratio of PUFA to SFA (P/S) showed that S (0.23) had a higher ratio than both SC (0.20) and C (0.19) (P = 0.038; Table 2). A significant diet by duration interaction was observed for the ratio of n-6/n-3 PUFA whereby the ratio of n-6/n-3 for groups SC and C increased with increasing feeding duration, while S was not affected. The current observations were in agreement with the study of French et al. (2000), where a decrease in the concentrate intake in grass-based diets resulted in an increased P/S ratio and a decreased n-6/n-3 ratio. Due to the low concentration of n-6 PUFA and high concentration of n-3 PUFA, Pasture had the lowest n-6/n-3 ratio when compared with all indoor finishing treatments (Table 2).
Branched chain fatty acids
The concentration of MOA, EOA and MNA found in SAT of lambs (Table 3) were consistent with other reports. In two lamb genotypes (high and low IMF) finished on one of four diets (perennial ryegrass, lucerne, or two brassica forages), the SAT contained 220–393 μg MOA/g, 89–170 μg EOA/g, and 35.1–63.6 μg MNA/g (Frank et al. 2016). In a study carried out by Schiller et al. (2015), different breeds of lambs fed intensively resulted in SAT with 56.9–103 μg MOA/g, 12.3–19.7 μg EOA/g, and 17.3–46.6 μg MNA/g. The diets of lambs affected all BCFA in SAT (Table 3; P < 0.001). The concentration of MOA was higher in C (335 μg/g) than S (278 μg/g), while SC was intermediate (314 μg/g) (P = 0.005). The concentration of EOA was higher in C (18.4 μg/g) and SC (19.1 μg/g) than S (8.5 μg/g) (P < 0.001). The concentration of MNA increased with increase in concentrate level in the diet (11.9 μg/g for S < 39.8 μg/g for SC < 66.3 μg/g for C) (P < 0.001). Overall, since BCFA are high in SAT, it is likely that when meat is cooked with fat attached the impact of BCFA on cooked meat aroma and flavour would be noticeable by trained or consumer panellists. Others have associated BCFAs with the mutton aroma of cooked sheep meat (Watkins et al. 2013; Frank et al. 2016). Moreover, in an internal preference mapping carried out by Prescott et al. (2001), results demonstrated that high levels of BCFA were associated with typical barnyard/milky/sour/sheep meat flavours. In a study of Young et al. (2003) results showed that, for 132 or 232 day old lambs, BCFA were higher in SAT of lambs fed concentrate than pasture due to increase formation of propionate in the rumen of concentrate-fed lambs (Garton et al. (1972). This supports the lower MOA in Pasture than C36, and lower MNA in Pasture than SC72 and C54 (Table 3; P < 0.05).

|
Conclusions
Silage to concentrate ratio and/or duration of feeding affect the fatty acid profile of the muscle and subcutaneous adipose fat of lambs, which were previously raised at pasture. Finishing lambs on a silage diet, particularly for an extended period, partially conserves the effects of pasture on muscle fatty acids, i.e. muscle contains a higher concentration and proportion of n-3 PUFA than concentrate-fed animals. Conversely, finishing lambs on a cereal concentrate diet reduces the muscle concentrations of n-3 PUFA but increases the n-6 PUFA, relative to pasture feeding, especially when concentrates are fed for long feeding durations. Finishing lambs on concentrate-based diets leads to increased levels of BCFA in the SAT of lambs.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors have no conflicts of interest to declare.
Declaration of funding
This work was funded by the Food Institutional Research Measure of the Irish Department of Agriculture, Food and the Marine (project 11/SF/310). The support of the Teagasc Walsh Fellowship programme (award 2013058) is also gratefully acknowledged.
References
Aurousseau B, Bauchart D, Calichon E, Micol D, Priolo A (2004) Effect of grass or concentrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs. Meat Science 66, 531–541.| Effect of grass or concentrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs.Crossref | GoogleScholarGoogle Scholar | 22060862PubMed |
Aurousseau B, Bauchart D, Faure X, Galot AL, Prache S, Micol D, Priolo A (2007) Indoor fattening of lambs raised on pasture. Part 1: Influence of stall finishing duration on lipid classes and fatty acids in the longissimus thoracis muscle. Meat Science 76, 241–252.
| Indoor fattening of lambs raised on pasture. Part 1: Influence of stall finishing duration on lipid classes and fatty acids in the longissimus thoracis muscle.Crossref | GoogleScholarGoogle Scholar | 22064292PubMed |
Boughalmi A, Araba A (2016) Effect of feeding management from grass to concentrate feed on growth, carcass characteristics, meat quality and fatty acid profile of Timahdite lamb breed. Small Ruminant Research 144, 158–163.
| Effect of feeding management from grass to concentrate feed on growth, carcass characteristics, meat quality and fatty acid profile of Timahdite lamb breed.Crossref | GoogleScholarGoogle Scholar |
Brunton NP, Mason C, Collins MJ (2015) Rapid microwave assisted preparation of fatty acid methyl esters for the analysis of fatty acid profiles in foods. Journal of Analytical Chemistry 70, 1218–1224.
| Rapid microwave assisted preparation of fatty acid methyl esters for the analysis of fatty acid profiles in foods.Crossref | GoogleScholarGoogle Scholar |
Claffey NA, Fahey AG, Gkarane V, Moloney AP, Monahan FJ, Diskin MG (2018) Effect of forage to concentrate ratio and duration of feeding on growth and feed conversion efficiency of male lambs. Translational Animal Science 2, 419–427.
| Effect of forage to concentrate ratio and duration of feeding on growth and feed conversion efficiency of male lambs.Crossref | GoogleScholarGoogle Scholar | 32704724PubMed |
Daley CA, Abbott A, Doyle PS, Nader GA, Larson S (2010) A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutrition Journal 9, 10
| A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef.Crossref | GoogleScholarGoogle Scholar | 20219103PubMed |
Daniel ZCTR, Wynn RJ, Salter AM, Buttery PJ (2004) Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: the role of stearoyl-CoA desaturase. Journal of Animal Science 82, 747–758.
| Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: the role of stearoyl-CoA desaturase.Crossref | GoogleScholarGoogle Scholar |
Dannenberger D, Nuernberg G, Scollan N, Schabbel W, Steinhart H, Ender K, Nuernberg K (2004) Effect of diet on the deposition of n-3 fatty acids, conjugated linoleic and C18:1trans fatty acid isomers in muscle lipids of German Holstein bulls. Journal of Agricultural and Food Chemistry 52, 6607–6615.
| Effect of diet on the deposition of n-3 fatty acids, conjugated linoleic and C18:1trans fatty acid isomers in muscle lipids of German Holstein bulls.Crossref | GoogleScholarGoogle Scholar | 15479030PubMed |
Demirel G, Ozpinar H, Nazli B, Keser O (2006) Fatty acids of lamb meat from two breeds fed different forage: concentrate ratio. Meat Science 72, 229–235.
| Fatty acids of lamb meat from two breeds fed different forage: concentrate ratio.Crossref | GoogleScholarGoogle Scholar | 22061549PubMed |
Dewhurst RJ, King PJ (1998) Effects of extended wilting, shading and chemical additives on the fatty acids in laboratory grass silages. Grass and Forage Science 53, 219–224.
| Effects of extended wilting, shading and chemical additives on the fatty acids in laboratory grass silages.Crossref | GoogleScholarGoogle Scholar |
Fahey AG, Marchant-Forde RM, Cheng HW (2007) Relationship between body weight and beak characteristics in one-day-old White Leghorn chicks: its implications for beak trimming. Poultry Science 86, 1312–1315.
| Relationship between body weight and beak characteristics in one-day-old White Leghorn chicks: its implications for beak trimming.Crossref | GoogleScholarGoogle Scholar | 17575176PubMed |
Font i Furnols M, Realini CE, Guerrero L, Oliver MA, Sañudo C, Campo MM, Nute GR, Cañeque V, Álvarez I, San Julián R, Luzardo S, Brito G, Montossi F (2009) Acceptability of lamb fed on pasture, concentrate or combinations of both systems by European consumers. Meat Science 81, 196–202.
| Acceptability of lamb fed on pasture, concentrate or combinations of both systems by European consumers.Crossref | GoogleScholarGoogle Scholar | 22063982PubMed |
Frank D, Watkins P, Ball A, Krishnamurthy R, Piyasiri U, Sewell J, Ortuño J, Stark J, Warner R (2016) Impact of Brassica and Lucerne finishing feeds and intramuscular fat on lamb eating quality and flavor. a cross-cultural study using Chinese and non-Chinese Australian consumers. Journal of Agricultural and Food Chemistry 64, 6856–6868.
| Impact of Brassica and Lucerne finishing feeds and intramuscular fat on lamb eating quality and flavor. a cross-cultural study using Chinese and non-Chinese Australian consumers.Crossref | GoogleScholarGoogle Scholar | 27523884PubMed |
French P, Stanton C, Lawless F, O’Riordan EG, Monahan FJ, Caffrey PJ, Moloney AP (2000) Fatty acid composition, including conjugated linoleic acid, of intramuscular fat from steers offered grazed grass, grass silage, or concentrate-based diets. Journal of Animal Science 78, 2849–2855.
| Fatty acid composition, including conjugated linoleic acid, of intramuscular fat from steers offered grazed grass, grass silage, or concentrate-based diets.Crossref | GoogleScholarGoogle Scholar | 11063308PubMed |
Garton GA, Hovell FD, Duncan WR (1972) Influence of dietary volatile fatty acids on the fatty-acid composition of lamb triglycerides, with special reference to the effect of propionate on the presence of branched-chain components. The British Journal of Nutrition 28, 409–416.
| Influence of dietary volatile fatty acids on the fatty-acid composition of lamb triglycerides, with special reference to the effect of propionate on the presence of branched-chain components.Crossref | GoogleScholarGoogle Scholar | 5085699PubMed |
Gkarane V, Brunton NP, Allen P, Gravador RS, Claffey NA, Diskin MG, Fahey AG, Farmer LJ, Moloney AP, Alcalde MJ, Murphy P, Monahan FJ (2019) Effect of finishing diet and duration on the sensory quality and volatile profile of lamb meat. Food Research International 115, 54–64.
| Effect of finishing diet and duration on the sensory quality and volatile profile of lamb meat.Crossref | GoogleScholarGoogle Scholar | 30599976PubMed |
Gravador RS, Harrison SM, Monahan FJ, Gkarane V, Farmer LJ, Brunton NP (2019) Validation of a rapid microwave-assisted extraction method and GC-FID quantification of total branched chain fatty acids in lamb subcutaneous adipose tissue. Journal of Food Science 84, 80–85.
| Validation of a rapid microwave-assisted extraction method and GC-FID quantification of total branched chain fatty acids in lamb subcutaneous adipose tissue.Crossref | GoogleScholarGoogle Scholar | 30557909PubMed |
Mezgebo GB, Monahan FJ, McGee M, O’Riordan EG, Richardson IR, Brunton NP, Moloney AP (2017) Fatty acid, volatile and sensory characteristics of beef as affected by grass silage or pasture in the bovine diet. Food Chemistry 235, 86–97.
| Fatty acid, volatile and sensory characteristics of beef as affected by grass silage or pasture in the bovine diet.Crossref | GoogleScholarGoogle Scholar | 28554651PubMed |
Noci F, Monahan FJ, French P, Moloney AP (2005) The fatty acid composition of muscle fat and subcutaneous adipose tissue of pasture-fed beef heifers: influence of the duration of grazing. Journal of Animal Science 83, 1167–1178.
| The fatty acid composition of muscle fat and subcutaneous adipose tissue of pasture-fed beef heifers: influence of the duration of grazing.Crossref | GoogleScholarGoogle Scholar | 15827261PubMed |
Nuernberg K, Nuernberg G, Ender K, Dannenberger D, Schabbel W, Grumbach S, Zupp W, Steinhart H (2005) Effect of grass vs. concentrate feeding on the fatty acid profile of different fat depots in lambs. European Journal of Lipid Science and Technology 107, 737–745.
| Effect of grass vs. concentrate feeding on the fatty acid profile of different fat depots in lambs.Crossref | GoogleScholarGoogle Scholar |
Ponnampalam EN, Sinclairt AJ, Egan AR, Blakeley SJ, Leury BJ (2001) Effect of diets containing n-3 fatty acids on muscle long-chain n-3 fatty acid content in lambs fed low- and medium-quality roughage diets. Journal of Animal Science 79, 698–706.
| Effect of diets containing n-3 fatty acids on muscle long-chain n-3 fatty acid content in lambs fed low- and medium-quality roughage diets.Crossref | GoogleScholarGoogle Scholar | 11263830PubMed |
Prescott J, Young O, O’Neill L (2001) The impact of variations in flavour compounds on meat acceptability: a comparison of Japanese and New Zealand consumers. Food Quality and Preference 12, 257–264.
| The impact of variations in flavour compounds on meat acceptability: a comparison of Japanese and New Zealand consumers.Crossref | GoogleScholarGoogle Scholar |
Realini CE, Duckett SK, Brito GW, Dalla Rizza M, De Mattos D (2004) Effect of pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Science 66, 567–577.
| Effect of pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef.Crossref | GoogleScholarGoogle Scholar | 22060866PubMed |
Scerra M, Luciano G, Caparra P, Foti F, Cilione C, Giorgi A, Scerra V (2011) Influence of stall finishing duration of Italian Merino lambs raised on pasture on intramuscular fatty acid composition. Meat Science 89, 238–242.
| Influence of stall finishing duration of Italian Merino lambs raised on pasture on intramuscular fatty acid composition.Crossref | GoogleScholarGoogle Scholar | 21570777PubMed |
Scerra M, Foti F, Cilione C, Chies L, Scerra V, Caparra P (2014) Influence of stall finishing of podolian young bulls raised on pasture on fatty acid composition and oxidative status of meat. Italian Journal of Animal Science 13, 3432
| Influence of stall finishing of podolian young bulls raised on pasture on fatty acid composition and oxidative status of meat.Crossref | GoogleScholarGoogle Scholar |
Schiller KF, Preuss S, Kaffarnik S, Vetter W, Rodehutscord M, Bennewitz J (2015) Concentration of three branched-chain fatty acids in adipose tissue does not affect meat sensory traits in crossbred and purebred German “Merinolandschaf” lambs. Archives Animal Breeding 58, 159–163.
| Concentration of three branched-chain fatty acids in adipose tissue does not affect meat sensory traits in crossbred and purebred German “Merinolandschaf” lambs.Crossref | GoogleScholarGoogle Scholar |
Vasta V, Mele M, Serra A, Scerra M, Luciano G, Lanza M, Priolo A (2009) Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins. Journal of Animal Science 87, 2674–2684.
| Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins.Crossref | GoogleScholarGoogle Scholar | 19395521PubMed |
Wang Z, Chen Y, Luo H, Liu X, Liu K (2015) Influence of restricted grazing time systems on productive performance and fatty acid composition of Longissimus dorsi in growing lambs. Asian-Australasian Journal of Animal Sciences 28, 1105–1115.
| Influence of restricted grazing time systems on productive performance and fatty acid composition of Longissimus dorsi in growing lambs.Crossref | GoogleScholarGoogle Scholar | 26104518PubMed |
Warren HE, Scollan ND, Enser M, Hughes SI, Richardson RI, Wood JD (2008) Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: Animal performance, carcass quality and muscle fatty acid composition. Meat Science 78, 256–269.
| Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: Animal performance, carcass quality and muscle fatty acid composition.Crossref | GoogleScholarGoogle Scholar | 22062278PubMed |
Watkins PJ, Frank D, Singh TK, Young OA, Warner RD (2013) Sheepmeat flavor and the effect of different feeding systems: a review. Journal of Agricultural and Food Chemistry 61, 3561–3579.
| Sheepmeat flavor and the effect of different feeding systems: a review.Crossref | GoogleScholarGoogle Scholar | 23488874PubMed |
Wood JD, Richardson RI, Nute GR, Fisher AV, Campo MM, Kasapidou E, Sheard PR, Enser M (2004) Effects of fatty acids on meat quality: a review. Meat Science 66, 21–32.
| Effects of fatty acids on meat quality: a review.Crossref | GoogleScholarGoogle Scholar | 22063928PubMed |
Wood JD, Enser M, Fisher AV, Nute GR, Sheard PR, Richardson RI, Hughes SI, Whittington FM (2008) Fat deposition, fatty acid composition and meat quality: a review. Meat Science 78, 343–358.
| Fat deposition, fatty acid composition and meat quality: a review.Crossref | GoogleScholarGoogle Scholar | 22062452PubMed |
Young OA, Lane GA, Priolo A, Fraser K (2003) Pastoral and species flavour in lambs raised on pasture, lucerne or maize. Journal of the Science of Food and Agriculture 83, 93–104.
| Pastoral and species flavour in lambs raised on pasture, lucerne or maize.Crossref | GoogleScholarGoogle Scholar |