

The distribution and significance of stingray feeding pits in Quandamooka (Moreton Bay), Australia
Max L. Giaroli A , Ilha Byrne A , Ben L. Gilby B * , Matthew Taylor A , Craig A. Chargulaf C and Ian R. Tibbetts A
B * , Matthew Taylor A , Craig A. Chargulaf C and Ian R. Tibbetts A
A
B
C
Abstract
Quandamooka (Moreton Bay) is a large, subtropical, urban, Ramsar-listed marine park in which stingray feeding pits (SRFPs) provide nursery habitat for commercially important nekton.
Nearmap aerial images were used to assess the distribution and abundance of SRFPs and their relationship to shore characteristics around Quandamooka’s perimeter.
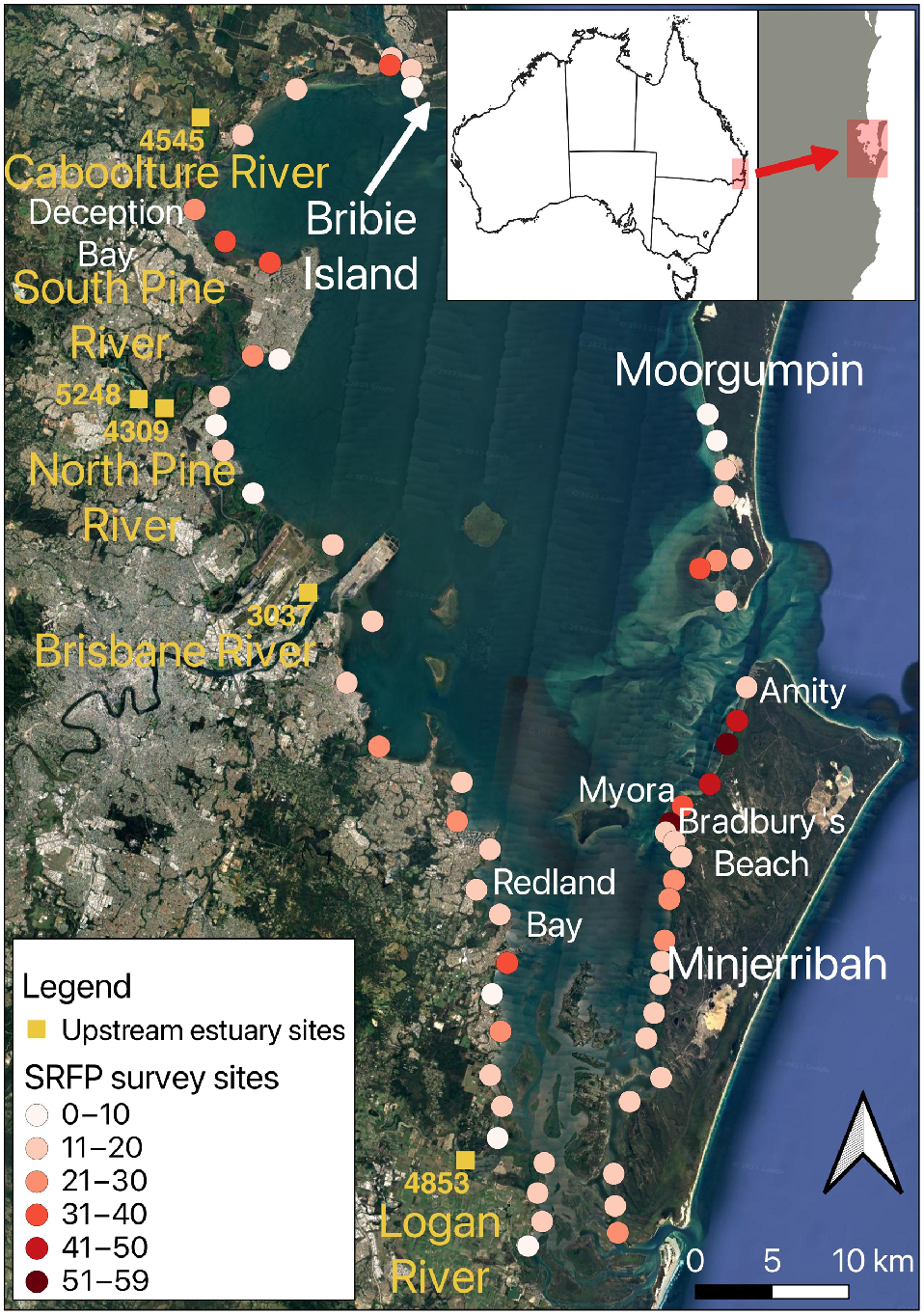
SRFPs were surveyed using 25-m2 quadrats overlain on aerial images at 65 sites on low-energy tidal flats around the perimeter of Quandamooka, and a polygon encompassing the region with the highest SRFP densities was used to make a preliminary assessment of its potential contribution to whiting and prawn fisheries.
SRFPs were present at 63 of the 65 sites surveyed, with habitat type in the high shore (especially mangroves v. seawalls) being the most important spatial driver of SRFP density.
Eastern Quandamooka sites held more SRFPs and may therefore be an important nursery habitat, holding an estimated 600,000 individuals of postlarval whiting (genus Sillago) and 8 million postlarval penaeid prawns.
Given the potentially important role of SRFPs for fisheries, especially those adjacent to mangroves, they should be considered a specific habitat type in future marine park zoning plans and managers should ensure the ongoing protection of stingrays.
Keywords: commercial fisheries, marine park, nursery habitat, penaeidae, recreational fisheries, Sillaginidae, Sillago, smelt-whiting.
Introduction
Stingrays are important ecosystem engineers in subtropical and tropical marine ecosystems globally (McGowan and Kajiura 2009; Flowers et al. 2021; Grew et al. 2024). They influence depositional environments by disturbing sediments while burying for camouflage, ventilating their gills while resting on the seabed, and excavating buried prey (Howard et al. 1977; O’Shea et al. 2012). Such bioturbation can increase the depth of the redox potential discontinuity (i.e. the transition from oxygenated to anoxic sediments), release nutrients into the water column, resuspend fine sediments and detritus, and affect benthic topography (O’Shea et al. 2012; Flowers et al. 2021). When stingrays locate a buried prey organism, they attempt to excavate it by vigorously expelling water from their ventrally located gill slits and driving water currents through postero-lateral undulations of their pectoral fins (Gregory et al. 1979; Pavic 2012). This process, defined as excavation feeding by Crook et al. (2022), results in a pit with two distinctive postero-lateral sediment plumes (Howard et al. 1977) (Fig. 1). In the intertidal zone, stingray feeding pits (SRFPs) located in sediments that are either saturated or have very low permeability (i.e. those that prevent water from draining) form shallow pools when the tide recedes (Krück et al. 2009; Chargulaf et al. 2011). Conversely, SRFPs excavated in well-drained sediments that lie above the beach water table tend to not form low-tide pools (I. R. Tibbetts, pers. obs.).
Images of stingray feeding pits in Quandamooka (Moreton Bay), Australia. (a) A water-filled stingray feeding pit with postero-lateral fans of sediment displaced by a foraging stingray. Bradbury’s Beach, Dunwich (Goompi), Minjerribah (North Stradbroke Island). (b) Stingray feeding pits at Hays Inlet (−27°15′35″, 153°4′19″), western Moreton Bay, showing postero-lateral sediment fans from a Nearmap aerial image. The image date function is visible in the lower right-hand corner and identifies the date the image was taken. Yellow arrows indicate a left sediment plume with respect to the orientation of the stingray.

Evidence from south-eastern Queensland, Australia, indicates that SRFPs are inhabited at low tide by juvenile fishes and crustaceans, including several economically important species, such as sand whiting (Sillago ciliata Cuvier, 1829) (Meager et al. 2005; Chargulaf et al. 2011) and penaeid prawns (Kwik 2003; Tibbetts et al. 2024). These species are associated with significant social and economic benefits for local people. For example, the 2160 tonnes (Mg) of eastern king prawn (Melicertus plebejus) harvested in Queensland in 2019 was valued at A$64.8 million. Similarly, an average of 272 Mg of sand whiting is harvested by commercial and recreational fishers from south-eastern Queensland, corresponding to a retail value of ~A$30 kg−1, and a total fishery value of over A$8.6 million (Wortmann and Hall 2021). Pool-dwelling juvenile nekton is likely to benefit through avoidance of aquatic predators, an abundance of harpacticoid copepod prey, and exposure to higher temperatures that potentially allow for higher growth rates during this critical early developmental period (Krück et al. 2009; Giaroli et al. 2023). Importantly, juvenile whiting (Chargulaf et al. 2011) and juvenile penaeid prawns (Tibbetts et al. 2024) occur year-round in Quandamooka SRFPs. If SRFP pools are important in the development of whiting and penaeid prawns, evaluating pool distribution and abundance could provide insights into the relative contribution of different shores to sustaining these economically important fisheries.
Quantifying the ecological and economic value of these low-energy coastal shores is important because coastal development is increasing rapidly in south-eastern Queensland (Gibbes et al. 2014; Thurstan et al. 2019). For example, although Quandamooka is listed as Ramsar Convention on Wetlands Site 631 because of its importance for migratory wading birds (McPhee 2017; Fuller et al. 2019; Ramsar Sites Information Service 2023), there is continued loss of intertidal wetland habitat because of anthropogenic impacts associated with development (Department of Environment, Science and Innovation, Queensland 2022). Because loss of such intertidal habitats also threatens elasmobranchs that use these habitats, the availability of SRFP pools (Leurs et al. 2023) for whiting and prawn development and recruitment to their respective fisheries may also be affected. This is further compounded by sea-level rise compressing available littoral habitats (Runting et al. 2018; Department of Environment, Science and Innovation, Queensland 2022). Because of the potential risk to both ecosystem and fisheries services provided by stingrays, it is essential that patterns of SRFP abundance are more thoroughly understood.
Quandamooka is a large, subtropical, semi-enclosed estuarine system located in eastern Australia, adjacent to several major population centres (including Brisbane, Redlands, Redcliffe, northern Gold Coast and southern Sunshine Coast). Quandamooka and its adjoining estuaries sustain extensive coastal habitats and wetlands, including mangroves, seagrass meadows, intertidal flats and saltmarsh (Department of Environment, Science and Innovation, Queensland 2022). These are important habitats for several locally occurring and abundant elasmobranchs. Quandamooka’s stingray community is diverse and abundant; however, three of the more commonly observed species on the intertidal sandflats of the bay are the Coral Sea mask ray (Neotrygon trigonoides (Last, White & Séret, 2016)), estuary stingray (Hemitrygon fluviorum (Ogilby, 1908)) and brown whipray (Maculabatis toshi (Whitley, 1939)) (Pierce et al. 2011). Pardo et al. (2015) investigated the diet of these species captured from western Quandamooka, finding that they predominantly ingest polychaete worms, crabs, and shrimp, which are abundant on its intertidal sand and mudflats. Previous studies within the region have also shown significant effects of landscape connectivity on the distribution of fishes across the seascape (Olds et al. 2012), and especially with mangroves for large benthic elasmobranchs (Henderson et al. 2018).
We used high-resolution Nearmap aerial digital images to survey the distribution and density of SRFPs around the perimeter of Quandamooka, and the upstream extent of SRFPs in estuary systems entering the bay. Quandamooka was chosen as the study seascape because of the presence of extensive unvegetated intertidal flats in different contexts across the bay, and previous studies indicating a high abundance of commercially important species within SRFPs of these shores, including sand whiting (Meager et al. 2005; Chargulaf et al. 2011) and penaeid prawns (Kwik 2003; Tibbetts et al. 2024). We hypothesized that (1) mangrove-lined shores would have higher densities of SRFP in comparison to modified shores because of greater habitat connectivity (e.g. Olds et al. 2012; Henderson et al. 2018), and (2) that SRFP density would vary with the shore morphology and sediment type because of the differences in wave energy status and thus SRFP longevity (e.g. O’Shea et al. 2012).
Materials and methods
Study site
Quandamooka is a large, subtropical, semi-enclosed embayment that is wide at the north and narrows to the south where shallow anastomosing channels separate numerous low deltaic islands (Fig. 2). Shores within the bay experience semi-diurnal tides, with a mean spring tidal range of ~1.5 m and a mean neap tidal range of ~0.8 m. Eastern bay shores are sheltered from prevailing south-easterly winds by Moorgumpin (Moreton Island) and Minjerribah (North Stradbroke Island) (McPhee 2017), whereas north-western bay shores typically experience higher-energy wind and waves. Major rivers include the Brisbane, North and South Pine and Caboolture Rivers in the northern bay and the Logan and Albert Rivers in the south (Fig. 2). These rivers deposit significant quantities of fine, muddy sediments on the nearby shores, which may modify the longevity of SRFPs (Beckman 2017). North-western depositional shores feature extensive ridge and runnel systems, whereas eastern shores are protected from wave action and strong currents by sinuous low-tide sand bars. Southern bay shores are protected by the high dunes of Minjerribah and small fetches between adjacent islands and sediment bars (Gibbes et al. 2014). As the energy status of a shore largely determines its sediment composition, detritus load and mobility (Martin et al. 2005), the longevity of features such as SRFPs depends on the prevailing energy regime of different shores (O’Shea et al. 2012). Furthermore, the western shores and catchments abut several large and heavily urbanised population centres, whereas the sand islands that enclose the eastern bay, by comparison, are largely undeveloped and protected by several national parks and land managed by Traditional Owners under Native Title awards.
Nearmap aerial images
Spatio-temporal data on SRFP density and abundance in Quandamooka were extracted from digital aerial images in Nearmap (Nearmap Australia), which was established in 2007 and covers 90% of Australia. Nearmap has archived aerial and satellite images of Quandamooka, with some locations being photographed up to six times per year, as per user requests and contracts. The frequency of images available for each site varies on the basis of user interest, resulting in the western more urbanised shores typically being photographed more regularly (several times per year) than the eastern shores (once every few years). The irregularity of images captured meant we were unable assess images of Quandamooka shores from the same period. Instead, the clearest available image for each location taken between 2010 and 2023, determined by good lighting and taken during low tide, was used. Most sampled images were taken during winter (n = 39, equating to 60% of images) and between May 2020 and June 2023 (n = 51, equating to 78% of images) (Supplementary Fig. S1 and Table S1). Nearmap images are taken normal to the earth surface, to which had been applied a panoramic correlation of all four cardinal directions, and a measurable oblique method that allows users to accurately calculate the height and dimensions of ground features (Heimgartner 2017). We used the vertical imagery approach, with the aerial and satellite images being presented in a cloud-based interface that allowed for their manipulation and analysis (Carroll 2023). All SRFP counts were made using aerial images with a resolution of 5.5–7.5-cm ground sampling distance (GSD) per pixel.
Pit identity and presence of pools
Recently excavated SRFPs were readily identifiable on the ground by the presence of postero-lateral plumes of sediment (Fig. 1a). At low virtual altitudes in aerial images, it was sometimes possible to observe a pair of postero-lateral fans adjacent to a pit (Fig. 1b), indicating its creation by a feeding stingray; however, typically all that could be discerned was the presence of SRFP of a size consistent with genesis by stingrays. Moreover, it was challenging to differentiate between SRFPs from which water had drained and those that formed pools. We chose to count all presumptive SRFPs, recognising that only those that remain pools while emersed are likely to serve any nursery-habitat function for juvenile nekton. Additionally, although we were able to identify recently dug SRFPs, which had sediment plumes, we were unable to assess SRFP longevity, which is likely to vary among shores, owing to the irregularity with which aerial images were taken.
SRFP surveys
The perimeter shores of Quandamooka were surveyed using Nearmap aerial images displayed on a 60.5-cm Dell SE2422 high resolution (1920 × 1080p) monitor set at Nearmap screen scale bar of 10 m giving a field of view ~200 × 120 m, at which any SRFPs were visible. The survey of eastern bay shores extended from the north-western tip of Moorgumpin (−27°3′36″, 153°21′42″) to the Jumpinpin Passage (−27°44′37″, 153°25′2″), which lies between North and South Minjerribah. The survey of western bay shores extended from the Bribie Island Bridge in the north (−27°4′15″, 153°8′43″) to Steiglitz (−27°44′24″, 153°21′26″) in the south. The image-date function was used to locate an image taken at low tide (e.g. bottom of Fig. 1b), which differed between some locations because of differences in coverage. For example, the urbanised western shores of Quandamooka are heavily sampled because of Nearmap client interests (Carroll 2023), whereas the eastern bay shores receive far less attention, with only three images in total available for our northern-most site on Moorgumpin. Despite this, sites were spread evenly to encompass the range of shore forms characteristic of the bay (Fig. 2).
When SRFPs were observed, a digital marker was placed to record latitude and longitude of the site and a single 5- × 5-m quadrat was placed at 0° orientation near the marker. The quadrat was moved over the highest SRFP density, and the pits counted at Nearmap screen scale bar set at 1 m, giving a field of view ~25 × 15 m. Highest density was used for this first quantification because of the uneven distribution of pits (highly clumped), rendering other uniform approaches untenable. This method was not designed to estimate the realised number of SRFP within the Bay, but to compare among sites. Pits on the perimeter of the quadrat were counted only if more than 50% of the pit lay within the quadrat boundary.
The position of the most upstream extent of SRFPs was determined for the Caboolture, North and South Pine, Brisbane, and Logan Rivers, by moving upstream progressively by using the clearest available image for each location taken between 2010 and 2023, determined by good lighting and taken during low tide. The stream midline distance from the river mouth was estimated using the line function in Nearmap and a mean and variance were derived.
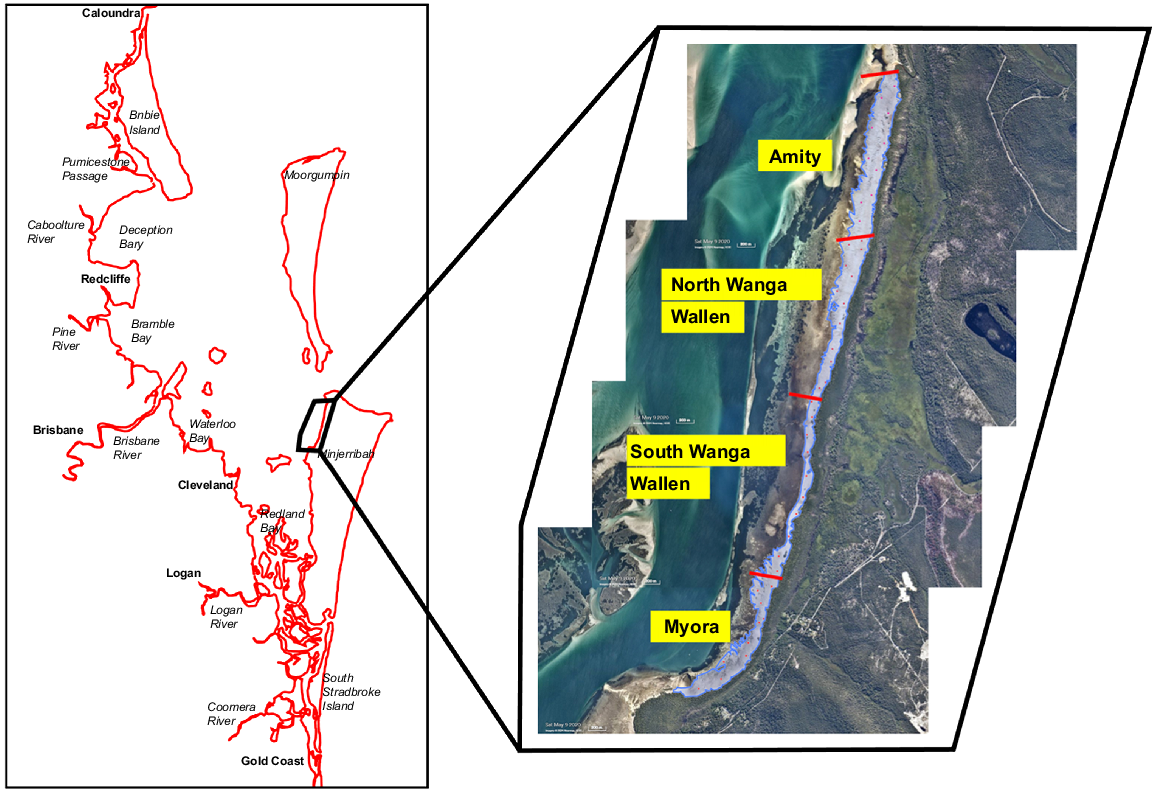
There appeared to be a very high density of SRFPs along the shore between Amity and Myora, North Minjerribah (Fig. 2). Consequently, the drawing overlay feature of Nearmap was used to superimpose four adjacent polygons over the area of an image taken at low tide on 9 May 2020, from Amity to Myora, North Minjerribah (Fig. 3). We identified that this area warranted closer scrutiny to obtain more accurate counts of SRFPs to help estimate the potential value of the area to economically important nekton (e.g. Sillago spp.), given the higher abundance of SRFPs found, rather than as a comparison between other sites in Quandamooka. It was not possible to overlay a single polygon because of the limit on the number of points available in the Nearmap polygon function and the complexity of the perimeter of unvegetated sediment because of seagrass and mangrove patches. In each ~2000-m-long polygon, 10 Nearmap balloon markers were dropped haphazardly at Nearmap scale 200 m; a scale at which SRFPs were not discernible. The top of a single, spatially locked, virtual 5- × 5-m quadrat was centred on the basal apex of the balloon marker and SRFPs were counted at a 1-m scale. To approximate the potential contribution of the nekton nursery service provided by this area, mean SRFP densities per 25-m2 quadrat were divided into the area of each polygon and then multiplied by modal densities of whiting and prawns per SRFP pool observed at Bradbury’s Beach and One Mile, Minjerribah, in surveys between September 2022 and March 2023 (Tibbetts et al. 2024).
The position of four adjacent polygons over unvegetated intertidal habitat along north-western Minjerribah (eastern Quandamooka (Moreton Bay), Australia) used to investigate sites that had very high abundances of stingray feeding pits (SRFPs). This area was subsequently termed an SRFP hotspot, and the four polygons were used to calculate surface area and sampled to derive mean SRFP densities rather than maximum abundance. Image is from Nearmap and taken on 9 May 2020. Red lines indicate the three boundaries between adjacent polygons.
Data analysis
Patterns in SRFP density from the bay perimeter surveys were identified using a generalised linear mixed model (GLMM) in the mgcv package (ver. 1.8-23, see https://cran.r-project.org/package=mgcv/; Wood 2017) of R (ver. 4.4.1, R Foundation for Statistical Computing, Vienna, Austria, see https://www.r-project.org/). The distribution of the data was visually checked to confirm that a Poisson distribution would be most appropriate. Five predictor variables were tested as fixed effects; upper shore habitat (three levels; mangroves, sand beach, seawall), sediment type (three levels; sand, muddy sand, sandy mud) (Fig. 4) and shore morphology (four levels; ridge and runnel, low-tide bar, channel, flat), which were each determined by visual inspection of Nearmap images (Fig. 5), and season (two levels; winter from March to August and summer all remaining months), and bay side (two levels; east and west) (Table S2). The variables year and site were included as random effects because images for each site could not be obtained from the same year. We minimised GLMM overfitting by running all combinations of fixed variables from the full GLMM by using the dredge function of the MuMIn package (ver. 1.48.4, K. Bartoń, see https://CRAN.R-project.org/package=MuMIn). Both random variables (year and site) were included in all dredged models. The model with the lowest Akaike’s information criterion (AIC) value was considered the best-fit model. The relative importance of each fixed variable was calculated by taking the sum of the weighted AIC values for each model in which the variable was included, resulting in an importance value between 0 and 1 for each variable. Here, importance values closer to 1 indicate a very strong contribution of the variable to explaining overall patterns, whereas values of 0 indicate no effect.
The different sediment types observed through analysis of Nearmap images of shores in Quandamooka (Moreton Bay), Australia. (a) A sandy shore of south-western Moorgumpin. (b) A muddy sand shore of Bradbury’s Beach. (c) A sandy mud shore on southern Minjerribah. These classifications were determined on the basis of the colouration of sediments and field trips to Quandamooka shores.

The different shore morphologies observed through analysis of Nearmap images of shores in Quandamooka (Moreton Bay), Australia. (a) A low-tide bar at Myora. (b) A channel on central south-western Minjerribah. (c) A flat. (d) A ridge and runnel system in western Quandamooka. Low-tide bars were easily identifiable due to the presence of a bar adjacent to the flat and ridge and runnel systems by the ‘ribbed’ pattern. Flats and channels were separated on the basis of the relative steepness of the drop-off, whereby flats had a gradual increase in depth, whereas channels had a steep depth increase.

One-way ANOVA and post hoc Tukey’s multiple-comparison tests (α = 0.05) were used to compare SRFP densities among the four contiguous polygons between Amity and Myora using GraphPad Prism (ver. 9.5.1, see https://www.graphpad.com/features).
Results
Bay-perimeter SRFP survey
Few shores around Quandamooka lacked visible SRFPs (Fig. 2), including north-western Moorgumpin from −27°15′10″, 153°23′4″, eastern shores of Redcliffe Peninsula from −27°15′49″, 153°6′13″to −27°11′22″, 153°6′46″, the north-western shores of Minjerribah from −27°24′28″, 153°26′15″ and two Redland Bay sites, located at 1–27°36′32″, 153°18′10″and at 6–27°41′31″, 153°19′15″), which lacked SRFPs. By contrast, the western shores of North Minjerribah (between −27°24′28″, 153°26′15″and 27°29′37″, 153°24′5″) had the highest concentrations of SRFP (maximum of 59 SRFPs per 25 m2). Other areas of notable SRFP density included Deception Bay and southern Moorgumpin (Fig. 2).
River SRFP maximum penetrations
Mean upstream penetration of SRFPs observable by using Nearmap images was 4326 m (±838 m s.d.). Highest penetration occurred in the South Pine River (5284 m) and the lowest in the highly urbanised Brisbane River (3037 m) (Table 1, Fig. 2). Muddy shores dominate these higher reaches, apart from the Logan River, where the most upstream SRFP seen was on a sand bar.
| River | Latitude | Longitude | Habitat type | Image date | Distance from estuary mouth (m) | |
|---|---|---|---|---|---|---|
| Caboolture | 27°7′33″ | 153°1′31″ | Muddy shore near mangroves | 31 May 2022 | 4545 | |
| North Pine | 27°14′7″, | 153°3′22″ | Muddy shore near mangrove island tail | 15 Nov. 2021 | 4309 | |
| South Pine | 27°17′50″ | 153°2′7″ | Muddy shore near mangroves | 15 Nov. 2021 | 5248 | |
| Brisbane | 27°23′32″ | 153°8′34″ | Muddy shore near mangroves | 10 July 2022 | 3037 | |
| Logan | 27°42′28″ | 153°18′8″ | Sand bar | 4 Nov. 2020 | 4853 |
Spatial distribution of SRFPs
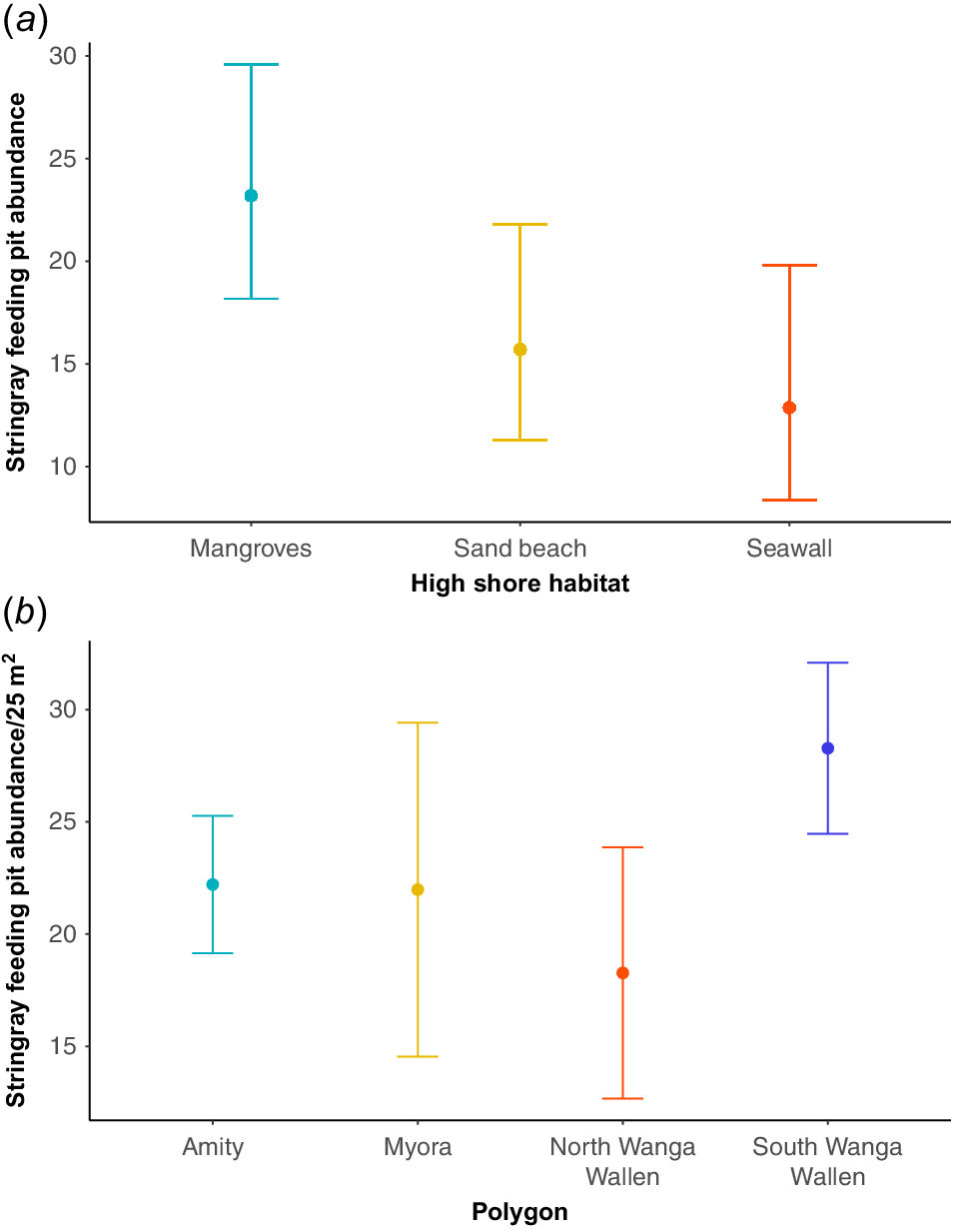
Variability in SRFP abundance was best explained by the high shore habitat at each site (F(2) = 6.11, P < 0.001), with mangrove-lined shores containing nearly double the average abundance of SRFPs (mean = 29.58 SRFPs) compared with seawall-lined shores (mean = 12.86), and 13.89 more STFPs on average than sand beach-lined shores (mean = 15.69) (Fig. 6a). All other fixed variables were removed during the model simplification and dredge process. The best-fit model explained 91.5% of deviance and had an adjusted R2 value of 0.79 (AIC = 577.9). Importantly, no other models were within five AIC of this best-fit model (next-best model 583.6). The large effect of high shore habitat for SRFP abundance was further supported by the variable importance values, with high shore habitats (importance = 1) being 20 times more important in explaining patterns than the next-most important fixed variable (season importance = 0.05).
(a) Generalised linear mixed-model (GLMM) output (with 95% confidence intervals), illustrating the effects of high shore habitat on stingray feeding pit abundance in Quandamooka (Moreton Bay), Australia. (b) Mean (±s.d.) stingray feeding pit density at four focal sites in eastern Quandamooka.
Eastern Bay SRFP hotspot and fisheries productivity
Mean SRFP densities differed significantly among the four hotspot polygons between Amity and Myora (F(3, 36) = 4.997, P < 0.01). Tukey’s post hoc analysis indicated that differences were due to higher densities at South Wanga Wallen area than in Northern Wanga Wallen (Fig. 6b), with no other pairwise differences. Multiplying the approximate annual daily modal abundance of juvenile nekton found in shore surveys in SRFPs at One Mile and Bradbury’s Beach on Minjerribah (Tibbetts et al. 2024) by the number of SRFPs on north-eastern Minjerribah, showed that this area could support ~0.57 million juvenile whiting and 8 million juvenile penaeids (Table 2).
| Polygon | Area (m2) | Mean SRFP (s.d.) | Number of 25-m2 quadrats | Total number of SRFPs | Whiting (0.5 per SRFP) | Prawns (7 per SRFP) | |
|---|---|---|---|---|---|---|---|
| Amity | 330,715 | 22.21 (3.06) | 13,229 | 293,777 | 0.15 × 106 | 2.06 × 106 | |
| North Wanga Wallen | 442,381 | 18.27 (5.60) | 17,695 | 323,313 | 0.16 × 106 | 2.26 × 106 | |
| South Wanga Wallen | 198,094 | 28.28 (3.81) | 7924 | 224,056 | 0.11 × 106 | 1.57 × 106 | |
| Myora | 346,031 | 21.98 (7.44) | 13,841 | 304,242 | 0.15 × 106 | 2.13 × 106 | |
| Total | 1,317,221 | 52,689 | 1,145,389 | 0.57 × 106 | 8.02 × 106 |
Four contiguous polygons were drawn around unvegetated intertidal habitats and their areas calculated by Nearmap. Total areas of each polygon were divided by 25-m2 quadrats where SRFPs were counted to determine the total number of SRFPs in each polygon.
Discussion
The presence of juvenile fish and prawns in intertidal SRFPs has been widely reported (e.g. Krück et al. 2009; Chargulaf et al. 2011). However, the potential for these formations to contribute to overall fisheries nursery value across entire seascapes has not been quantified, and nor have the environmental conditions that most contribute towards these effects. Mangrove-lined shores of Quandamooka had higher densities of SRFPs than did modified shores, as was hypothesised, with sandy shores having an intermediate value. These findings support our first hypothesis, and the notion that hardening and urbanisation of shorelines in marine environments has significantly detrimental effects for fish. However, in this study we have shown that these effects extend beyond localised effects in individual species, to likely effects on habitat-forming species in soft-shore environments, and potential flow-on effects for nursery habitat and fisheries values. Crucially, we found no evidence to suggest that SRFP density varied with bay side, sediment type, season or shore morphology. Indeed, relative importance values indicated that highshore habitat type was at least 20 times more important in explaining SRFP distribution than was each of these variables. Therefore, our second hypothesis was not supported. Given these findings, mangrove-lined shores between Amity and Myora in eastern Quandamooka supported a higher density of SRFPs than did other locations, resulting in a very high abundance of both larval prawns and whiting at these locations. Maintaining connectivity between high shore mangrove forests and soft-sediment shores should therefore be a priority for conservation, restoration and management. More broadly, however, these findings add significant credence to the importance of soft-shore habitats in supporting SRFPs, irrespective of their position across embayments or different sediment types. Therefore, these habitats should be prioritised in future conservation initiatives to assist in maintaining these likely crucial nursery sites for commercially important invertebrates and fish.
Stingrays are sensitive to changes in water quality, including salinity, temperature, dissolved oxygen and pH (Chin et al. 2010; Schlaff et al. 2014). Three of the four most abundant stingrays in Quandamooka have limited freshwater penetration capacity (Neotrygon trigonoides, Maculabatis toshi, Pateobatis fai) (Pierce et al. 2011). This limited tolerance is due to the decreasing salinity overcoming their ability to maintain ionic homeostasis and thus losing ions to the surrounding environment, forcing non-euryhaline species to stay in water with higher salinity (Tam et al. 2003). This is likely to be one of the most important influences on the relative low penetration of SRFP into estuaries that are subjected to higher freshwater input within the region (Gibbes et al. 2014). Anthropogenic influences may also contribute to this trend, with increased runoff and degraded water quality from catchments with extensive urbanisation, agriculture, and industry occurring within these estuaries (Pierce et al. 2011; Gibbes et al. 2014; Maxwell et al. 2014). In contrast, the central bay typically experiences lower turbidity and has relatively stable salinity, owing to a strong oceanic influence from warm, oligotrophic South Pacific water (Dennison and Abal 1999). Indeed, this upstream estuarine limitation was the only spatial restriction identified in this study, beyond the major effects of high shore habitat.
We were surprised by the lack of detected influence of bay side and sediment type on SRFP abundance in this study, given potential effects of wind and wave exposure and SRFP longevity. Western bay shores (especially north of the Brisbane River) experience a higher-energy environment because they are subjected to larger waves from dominant south-easterly trade winds and larger wave fetch (Dennison and Abal 1999). We expected that this would result in higher sediment-transport rates, more rapid SRFP extinction and, therefore, lower SRFP abundance in imagery. For example, along more protected western shores, such as southern Deception Bay, it is possible that the deposition of muddier sediments from nearby rivers might increase SRFP pool longevity (Beckman 2017). Eastern Bay shores have a low profile and are protected from wave action by low-tide sand bars that exceed the elevation of the middle shore where stingray pits are predominantly located (H. Bostock, pers. comm.). The low shore bars are likely to shield eastern Quandamooka shores (especially between central-west Minjerribah and Amity) from significant wave and wind action, decreasing the rate of pit erasure by wave and tidal action. Decreased wave action may also help support the shoreline mangrove communities that our data suggest correlate with higher SRFP density. Although the longevity of SRFPs was not able to be assessed in this study because of the sporadic nature of Nearmap imagery, future studies could further tease apart the effects of sediment and shoreline exposure on SRFP persistence and longevity, and linked effects on nursery-habitat values.
Our a priori prediction that mangrove-backed shores would have a higher mean SRFP density was supported. This supports the results of previous research, suggesting that mangrove-lined intertidal shores are important habitat features for stingrays and other elasmobranchs (Schlaff et al. 2014; Henderson et al. 2018; Kanno et al. 2023). For example, it has been established that juvenile mangrove whiprays (Urolophus granulatus) use both mangrove habitats and sandy flats in Western Australia (Kanno et al. 2019) and mangroves are important for giant shovel-nosed rays (Glaucostegus typus) in Quandamooka (Henderson et al. 2018). Martins et al. (2020, 2021) visually tracked juvenile cowtail rays (Pastinachus ater) and juvenile mangrove whiprays (Urolophus granulatus) on a reef-flat embayment with a mangrove-lined upper shore in the central Great Barrier Reef. They found that both species moved in with the rising tide to sand flats and mangrove edges, surmising that this was due to food availability and protection from predators. We were unable to assess signs of stingray activity in the mangroves from aerial images because the sediment surface was obscured by the mangrove canopy and pneumatophores. Ideally, telemetry or direct observations of mangrove habitat use by stingrays, such as those by Martins et al. (2020, 2021), should be used to determine the role of mangrove habitats for Quandamooka stingrays. Studies quantifying the attributes of mangrove forests that mostly drive these effects (e.g. tree density, food availability) could also further assist in prioritising conservation and restoration actions within Quandamooka.
It is likely that the patterns found in this study for mangrove-backed shorelines are due to the combined effects of higher food availability, shelter and wave abatement near mangroves resulting in stingrays prioritising feeding in these areas. It is well established that mangroves support a higher abundance of invertebrate prey (Kabir et al. 2014), and that these effects are likely to expand into the surrounding soft-shore environments (Sheaves et al. 2016; Giaroli et al. 2023). Indeed, prey abundance and diversity have been previously suggested as a significant determinant of elasmobranch distribution and habitat use across the Quandamooka seascape (Henderson et al. 2018). This is likely to be a crucial effect in this study because the presence of SRFPs is a direct indicator of feeding and not simply of stingray presence in these environments. Sandy shores also support an abundance of invertebrates, but likely not to the same degree in this system as do mangroves, typically owing to lower subsidies on sand-backed shorelines (Hyndes et al. 2022). The effects of hardened shorelines with extensive, low-complexity seawalls are increasingly recognised globally (e.g. Heery et al. 2017), with key effects being found for fish (e.g. Brook et al. 2018), invertebrates (e.g. Vozzo et al. 2021), and primary producers (e.g. Lai et al. 2018). In contrast to high-value mangrove-lined shores, it is likely, therefore, that hardened shorelines support both a lower stingray abundance as well as less food. Shores with seawalls are also more likely to be frequented by people for recreation (e.g. shoreline walking, fishing), which may also lead to reduced prevalence of timid stingray species at these shores. It is also more likely that shorelines lined by seawalls have higher local sediment mobility and wave movement because of the reflection of wave energy and boat wake off vertical surfaces (Miles et al. 2001), leading to less persistent SRFPs. The increased sediment mobility on such shores could also lead to burial and decreased survival of invertebrates such as polychaetes (Hinchey et al. 2006), which feature heavily in the diet of stingrays (Jacobsen and Bennett 2012), leading to lower stingray feeding activity at these locations.
Ground observations confirmed that the features identified as SRFPs in aerial images were stingray pits; however, those on elevated features had drained and only those in either muddy sediment or interacting with the beach water table supported pools. More extensive ground surveys or higher-resolution imagery from drone-mounted cameras (e.g. Grew et al. 2024) could better characterise the location and size of SRFPs useful as nurseries for prawns and whiting. Future work should focus on quantifying the abundance of juvenile whiting and prawns in different areas of the bay and at different times throughout the year with spawning and recruitment pulses. Crucially, this must be undertaken independently from simply counting SRFPs, because results of this study demonstrated no variation in SRFP abundance between seasons. Similarly, surveys undertaken in this study were limited by the availability and timing of clear aerial imagery on Nearmap. This limited potential seasonal and annual analyses of SRFP prevalence, meaning that future studies could more thoroughly test these effects.
This study used aerial images to quantify the distribution of SRFPs around Quandamooka, finding them to occur on most soft-sediment shores throughout the bay, but detectable only a few kilometres up nearby major rivers. The north-western shores of Minjerribah had consistently high abundances of SRFPs over an extensive area, indicating that they might be particularly important to protect. Conserving connectivity between intertidal flats and adjacent mangrove habitats appears crucial in maximising fisheries outcomes, a finding supported by previous research both within Quandamooka and beyond. Given the wide tropical and subtropical distribution of stingrays, whiting and prawns throughout the Indo-Pacific, our approach and findings will be of relevance to habitat and fisheries managers for use in fisheries habitat protection and marine-park zoning. Stingrays are important habitat-forming species on sandy depositional shores, resulting in shallow pits where the juveniles of commercially important nekton are common (Krück et al. 2009; Chargulaf et al. 2011). However, stingrays are increasingly threatened by extinction (Pierce et al. 2011; Dulvy et al. 2021; Dulvy and Simpfendorfer 2022), which might threaten these key habitat features on sedimentary shores of the tropics and subtropics.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Declaration of funding
This study was supported by The University of Queensland through a licencing agreement with Nearmap Australia.
Acknowledgements
The authors acknowledge the many thousands of years of custodianship of Quandamooka Sea Country by Traditional Owners including Nunukul, Ngugi, and Goenpul First Nations People of Minjerribah and Moorgumpin, and the Turrbul and Kabi Kabi Traditional Custodians of the shores examined in this study. We pay our respects to their Elders past, present and emerging. The authors thank two anonymous reviewers for comments on this paper.
References
Brook TW, Gilby BL, Olds AD, Connolly RM, Henderson CJ, Schlacher TA (2018) The effects of shoreline armouring on estuarine fish are contingent upon the broader urbanisation context. Marine Ecology Progress Series 605, 195-206.
| Crossref | Google Scholar |
Carroll S (2023) A different point-of-view – using aerial imagery to build stronger cities. Photogrammetric Engineering & Remote Sensing 89(1), 5-8.
| Crossref | Google Scholar |
Chargulaf CA, Townsend KA, Tibbetts IR (2011) Community structure of soft sediment pool fishes in Moreton Bay, Australia. Journal of Fish Biology 78(2), 479-494.
| Crossref | Google Scholar | PubMed |
Chin A, Kyne PM, Walker TI, McAuley RB (2010) An integrated risk assessment for climate change: analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef. Global Change Biology 16(7), 1936-1953.
| Crossref | Google Scholar |
Crook KA, Sheaves M, Barnett A (2022) Species-specific foraging behaviors define the functional roles of sympatric stingrays. Limnology and Oceanography 67(1), 219-230.
| Crossref | Google Scholar |
Department of Environment, Science and Innovation, Queensland (2022) Mangroves and associated communities of Moreton Bay. WetlandInfo website. Available at https://wetlandinfo.des.qld.gov.au/wetlands/ecology/components/biota/flora/flora-structural/mangroves/mangrove-moreton.html [Verified 26 April 2024]
Dulvy NK, Pacoureau N, Rigby CL, Pollom RA, Jabado RW, Ebert DA, Finucci B, Pollock CM, Cheok J, Derrick DH, Herman KB, Sherman CS, VanderWright WJ, Lawson JM, Walls RHL, Carlson JK, Charvet P, Bineesh KK, Fernando D, Ralph GM, Matsushiba JH, Hilton-Taylor C, Fordham SV, Simpfendorfer CA (2021) Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Current Biology 31(21), 4773-4787.e8.
| Crossref | Google Scholar | PubMed |
Flowers KI, Heithaus MR, Papastamatiou YP (2021) Buried in the sand: uncovering the ecological roles and importance of rays. Fish and Fisheries 22(1), 105-127.
| Crossref | Google Scholar |
Giaroli ML, Chargulaf CA, Gilby BL, Tibbetts IR (2023) Tidal migrations of juvenile Sillago spp. in a subtropical intertidal nursery seascape. Marine and Freshwater Research 74(14), 1193-1210.
| Crossref | Google Scholar |
Gibbes B, Grinham A, Neil D, Olds A, Maxwell P, Connolly R, Weber T, Udy N, Udy J (2014) Moreton Bay and its estuaries: a sub-tropical system under pressure from rapid population growth. In ‘Estuaries of Australia in 2050 and beyond’. Estuaries of the World. (Ed. E Wolanski) pp. 203–222. (Springer: Dordrecht, Netherlands) 10.1007/978-94-007-7019-5_12
Gregory MR, Balance PF, Gibson GW, Ayling AM (1979) On how some rays (Elasmobranchia) excavate feeding depressions by jetting water. SEPM Journal of Sedimentary Research 49, 1125-1129.
| Crossref | Google Scholar |
Grew M, Gaston TF, Griffin AS, Duce SJ, Raoult V (2024) Ray bioturbation rates suggest they shape estuary processes. Remote Sensing in Ecology and Conservation
| Crossref | Google Scholar |
Heery EC, Bishop MJ, Critchley LP, Bugnot AB, Airoldi L, Mayer-Pinto M, Sheehan EV, Coleman RA, Loke LHL, Johnston EL, Komyakova V, Morris RL, Strain EMA, Naylor LA, Dafforn KA (2017) Identifying the consequences of ocean sprawl for sedimentary habitats. Journal of Experimental Marine Biology and Ecology 492, 31-48.
| Crossref | Google Scholar |
Heimgartner J (2017) Service provides clear, orthorectified and current images of construction site. (engineering.com) Available at https://mobile.engineering.com/amp/16087.html [Verified November 2023]
Henderson CJ, Stevens T, Gilby BL, Lee SY (2018) Spatial conservation of large mobile elasmobranchs requires an understanding of spatio-temporal seascape utilization. ICES Journal of Marine Science 75(2), 553-561.
| Crossref | Google Scholar |
Hinchey EK, Schaffner LC, Hoar CC, Vogt BW, Batte LP (2006) Responses of estuarine benthic invertebrates to sediment burial: the importance of mobility and adaptation. Hydrobiologia 556(1), 85-98.
| Crossref | Google Scholar |
Howard JD, Mayou TV, Heard RW (1977) Biogenic sedimentary structures formed by rays. SEPM Journal of Sedimentary Research 47(1), 339-346.
| Crossref | Google Scholar |
Hyndes GA, Berdan EL, Duarte C, Dugan JE, Emery KA, Hambäck PA, Henderson CJ, Hubbard DM, Lastra M, Mateo MA, Olds A, Schlacher TA (2022) The role of inputs of marine wrack and carrion in sandy-beach ecosystems: a global review. Biological Reviews 97(6), 2127-2161.
| Crossref | Google Scholar | PubMed |
Jacobsen IP, Bennett MB (2012) Feeding ecology and dietary comparisons among three sympatric Neotrygon (Myliobatoidei: Dasyatidae) species. Journal of Fish Biology 80(5), 1580-1594.
| Crossref | Google Scholar | PubMed |
Kabir M, Abolfathi M, Hajimoradloo A, Zahedi S, Kathiresan K, Goli S (2014) Effect of mangroves on distribution, diversity and abundance of molluscs in mangrove ecosystem: a review. Aquaculture, Aquarium, Conservation & Legislation 7(4), 286-300.
| Google Scholar |
Kanno S, Schlaff AM, Heupel MR, Simpfendorfer CA (2019) Stationary video monitoring reveals habitat use of stingrays in mangroves. Marine Ecology Progress Series 621, 155-168.
| Crossref | Google Scholar |
Kanno S, Heupel MR, Sheaves MJ, Simpfendorfer CA (2023) Mangrove use by sharks and rays: a review. Marine Ecology Progress Series 724, 167-183.
| Crossref | Google Scholar |
Krück NC, Chargulaf CA, Saint-Paul U, Tibbetts IR (2009) Early post-settlement habitat and diet shifts and the nursery function of tidepools during Sillago spp. recruitment in Moreton Bay, Australia. Marine Ecology Progress Series 384, 207-219.
| Crossref | Google Scholar |
Lai S, Loke LHL, Bouma TJ, Todd PA (2018) Biodiversity surveys and stable isotope analyses reveal key differences in intertidal assemblages between tropical seawalls and rocky shores. Marine Ecology Progress Series 587, 41-53.
| Crossref | Google Scholar |
Leurs G, Nieuwenhuis BO, Zuidewind TJ, Hijner N, Olff H, Govers LL (2023) Where land meets sea: intertidal areas as key-habitats for sharks and rays. Fish and Fisheries 24(3), 407-426.
| Crossref | Google Scholar |
Martin D, Bertasi F, Colangelo MA, de Vries M, Frost M, Hawkins SJ, Macpherson E, Moschella PS, Satta MP, Thompson RC, Ceccherelli VU (2005) Ecological impact of coastal defence structures on sediment and mobile fauna: evaluating and forecasting consequences of unavoidable modifications of native habitats. Coastal Engineering 52(10–11), 1027-1051.
| Crossref | Google Scholar |
Martins APB, Heupel MR, Bierwagen SL, Chin A, Simpfendorfer C (2020) Diurnal activity patterns and habitat use of juvenile Pastinachus ater in a coral reef flat environment. PLoS ONE 15(2), e0228280.
| Crossref | Google Scholar |
Martins APB, Heupel MR, Bierwagen SL, Chin A, Simpfendorfer CA (2021) Tidal–diel patterns of movement, activity and habitat use by juvenile mangrove whiprays using towed-float GPS telemetry. Marine and Freshwater Research 72(4), 534-541.
| Crossref | Google Scholar |
Maxwell PS, Pitt KA, Burfeind DD, Olds AD, Babcock RC, Connolly RM (2014) Phenotypic plasticity promotes persistence following severe events: physiological and morphological responses of seagrass to flooding. Journal of Ecology 102(1), 54-64.
| Crossref | Google Scholar |
McGowan DW, Kajiura SM (2009) Electroreception in the euryhaline stingray, Dasyatis sabina. Journal of Experimental Biology 212(10), 1544-1552.
| Crossref | Google Scholar | PubMed |
Meager JJ, Williamson I, King CR (2005) Factors affecting the distribution, abundance and diversity of fishes of small, soft-substrata tidal pools within Moreton Bay, Australia. Hydrobiologia 537, 71-80.
| Crossref | Google Scholar |
Miles JR, Russell PE, Huntley DA (2001) Field measurements of sediment dynamics in front of a seawall. Journal of Coastal Research 17(1), 195-206.
| Google Scholar |
Olds AD, Connolly RM, Pitt KA, Maxwell PS (2012) Habitat connectivity improves reserve performance. Conservation Letters 5(1), 56-63.
| Crossref | Google Scholar |
O’Shea OR, Thums M, van Keulen M, Meekan M (2012) Bioturbation by stingrays at Ningaloo Reef, Western Australia. Marine and Freshwater Research 63(3), 189-197.
| Crossref | Google Scholar |
Pardo SA, Burgess KB, Teixeira D, Bennett MB (2015) Local-scale resource partitioning by stingrays on an intertidal flat. Marine Ecology Progress Series 533, 205-218.
| Crossref | Google Scholar |
Pavic I (2012) Dasyatis americana Kit (also: Southern Stingray; Stingaree; Whip stingray). In ‘Animal Diversity Web’. (University of Michigan) Available at https://animaldiversity.org/accounts/Dasyatis_americana/#35669C7A-B34D-44D7-AA3F-58702170870B [Verified May 2022]
Pierce SJ, Scott-Holland TB, Bennett MB (2011) Community composition of elasmobranch fishes utilizing intertidal sand flats in Moreton Bay, Queensland, Australia. Pacific Science 65(2), 235-247.
| Crossref | Google Scholar |
Ramsar Sites Information Service (2023) Ramsar information sheet for Site number 631, Moreton Bay, Australia. Ramsar. Available at https://rsis.ramsar.org/RISapp/files/RISrep/AU631RIS_2312_en.pdf [Verified 05 July 2024]
Runting RK, Beyer HL, Dujardin Y, Lovelock CE, Bryan BA, Rhodes JR (2018) Reducing risk in reserve selection using Modern Portfolio Theory: coastal planning under sea-level rise. Journal of Applied Ecology 55(5), 2193-2203.
| Crossref | Google Scholar |
Schlaff AM, Heupel MR, Simpfendorfer CA (2014) Influence of environmental factors on shark and ray movement, behaviour and habitat use: a review. Reviews in Fish Biology and Fisheries 24, 1089-1103.
| Crossref | Google Scholar |
Sheaves M, Dingle L, Mattone C (2016) Biotic hotspots in mangrove-dominated estuaries: macro-invertebrate aggregation in unvegetated lower intertidal flats. Marine Ecology Progress Series 556, 31-43.
| Crossref | Google Scholar |
Tam WL, Wong WP, Loong AM, Hiong KC, Chew SF, Ballantyne JS, Ip YK (2003) The osmotic response of the Asian freshwater stingray (Himantura signifer) to increased salinity: a comparison with marine (Taeniura lymma) and Amazonian freshwater (Potamotrygon motoro) stingrays. Journal of Experimental Biology 206(17), 2931-2940.
| Crossref | Google Scholar | PubMed |
Thurstan R, Fraser K, Brewer D, Buckley S, Dinesen Z, Skewes T, Courtney T, Pollock B (2019) Fishers and fisheries of Moreton Bay. In ‘Moreton Bay (Quandamooka) & catchment: past, present, and future’. (Eds IR Tibbetts, PC Rothlisberg, DT Neil, TA Homburg, DT Brewer, AH Arthington) pp. 521–536. (The Moreton Bay Foundation: Brisbane, Qld, Australia)
Vozzo ML, Cumbo VR, Crosswell JR, Bishop MJ (2021) Wave energy alters biodiversity by shaping intraspecific traits of a habitat-forming species. Oikos 130(1), 52-65.
| Crossref | Google Scholar |