

Growth, metal partitioning and antioxidant enzyme activities of mung beans as influenced by zinc oxide nanoparticles under cadmium stress
Md Harunur Rashid A B C , Mohammad Mahmudur Rahman A B * , Md Abdul Halim A B and Ravi Naidu A B *
A B C , Mohammad Mahmudur Rahman A B * , Md Abdul Halim A B and Ravi Naidu A B *
A Global Centre for Environmental Remediation (GCER), College of Engineering, Science and Environment, The University of Newcastle, Callaghan, Newcastle, NSW 2308, Australia.
B Cooperative Research Centre for Contamination Assessment and Remediation of the Environment (CRC CARE), ATC Building, The University of Newcastle, Callaghan, Newcastle, NSW 2308, Australia.
C Bangladesh Agricultural Research Institute (BARI), Gazipur 1701, Bangladesh.
Handling Editor: Shahid Hussain
Crop & Pasture Science - https://doi.org/10.1071/CP21598
Submitted: 4 May 2021 Accepted: 8 December 2021 Published online: 11 May 2022
© 2021 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Cadmium (Cd) toxicity and zinc (Zn) deficiency are of major concerns for crop growth and quality. Moreover, their interactive effects exert some controversial reports.
Aims: The effects of zinc oxide nanoparticles (ZnO NPs) and Cd on growth, physiology, and metal distribution in mung beans (Vigna radiata L.) was investigated.
Methods: Seven-day-old seedlings were treated with Zn (0, 1, 2, 4, 8, 16 and 32 μM) and Cd (0, 0.5, 1 μM) for 14 days.
Key results: Photosynthetic pigments, antioxidant enzyme activities, dry matter yield and metal concentration in tissues were significantly influenced by ZnO NPs and Cd. Considered on its own as a main effect, Zn application (16 μM) enhanced its accumulation in roots, stem and leaf by about 33-fold (314 mg kg−1), 10-fold (60.6 mg kg−1) and 17-fold (110.8 mg kg−1), respectively, compared to control. However, accumulation was slower for interactions with Cd. While leaf Zn increased approximately 27 times (180 mg kg−1) at 32 μM Zn, its interactions with lower and higher Cd increased only 6-fold (41.2 mg kg−1) and 3-fold (21.4 mg kg−1), respectively. Added ZnO NPs up to 4 μM under Cd contamination elevated the leaf Cd, which was restricted by higher supply. However, Cd accumulation in stem and root consistently rose, indicating a synergistic effect. ZnO NPs induced an upregulation of antioxidant enzymes to avert oxidative stress and maintain growth performance.
Implications: These findings may be suitable for formulating nanomaterials of desired particle sizes and testing on other crop to remediate Cd.
Keywords: antioxidant enzymes, biofortification, cadmium, chlorophyll, metal partitioning, mung beans, photosynthetic pigments, reactive oxygen species (ROS), transfer/translocation factor, zinc oxide nanoparticles.
Introduction
Cadmium (Cd) is a toxic element, causing kidney damage and other diseases in humans (IARC 2018). Exposure to Cd primarily occurs through food crops grown in Cd-contaminated soils, so diet is the crucial source of Cd entry into the human body (Martos et al. 2016; Zare et al. 2018). It has a direct impact on plant growth including reduction of the root and shoot length, biomass, photosynthetic properties and enzyme activities (Rizwan et al. 2019b). Cd toxicity in crops is more severe under Zn-deficient conditions (Köleli et al. 2004). Plants can tolerate a certain level of Cd through binding with cell walls, compartmentalisation of it in inactive portions, chelation and enhancement in certain enzymes (Rizwan et al. 2016), and gene activation to block metal transport (Martos et al. 2016). However, presence of Cd in higher concentrations than optimal interferes with normal metabolic functions of plant cell, induces the accumulation of reactive oxygen species (ROS) including H2O2, malondialdehyde (MDA), and O2− in plants (Rizwan et al. 2016) which lead to inactivation of enzymes and dysfunction of various plant physiological and biochemical processes (Hayat et al. 2012). Plants have their own mechanism to prevent the negative effects of ROS through activation of antioxidant enzyme systems, e.g. ascorbate peroxidase (APX), catalase (CAT), peroxidase (POD), superoxide dismutase (SOD) (Mittler 2002).
Zinc (Zn) is essential for plants and can greatly minimise the uptake of Cd by roots and other parts of crops. Due to their chemical similarity, they compete with each other for binding sites in soils and on the root surface (Zare et al. 2018) demonstrating both antagonistic (Murtaza et al. 2017) and synergistic outcomes (Cojocaru et al. 2016). In one study, it was reported that diminishing the Cd:Zn ratio by increasing Zn concentration in the nutrient solution caused a considerable reduction of Cd transport and accumulation in lettuce leaves (Zare et al. 2018). Soil application of Zn at 15 and 30 mg kg−1 and foliar application at 0.3% and 0.6% were found to be effective in alleviating Cd concentration in wheat grains grown in soil spiked with 30 mg Cd kg−1 (Saifullah et al. 2014). Conversely, in bread and durum wheat, Köleli et al. (2004) observed that application of 10 and 25 mg Zn kg−1 on 10 mg Cd kg−1 spiked soil increased shoot Cd in 35-day-old plants and decreased in 65-day-old plants. Application of 5 mg Zn kg−1 increased Cd concentration in roots, shoots and leaves of tomato and this fell with higher doses (Mohammad and Moheman 2010). Murtaza et al. (2017) discovered that Zn application in soils suppressed uptake of Cd and improved the performance of cereals (wheat and maize), pulses (mung bean) and legumes (chickpea). It was also reported that applying 1.1 kg of Zn as Zn-EDTA per hectare as a foliar supplement in combination with nitrogen significantly reduced grain Cd and increased grain Zn (Forster et al. 2018).
Nanotechnology, a recently emerging scientific technology, is considered to be an ideal solution to combat environmental pollution. The use of ZnO NPs has recently drawn much attention for their potential to enhance the accumulation of nutrients by plants and to ensure quality outcomes are produced (Rizwan et al. 2019a). ZnO NPs are now extensively used in agriculture as well as biomedical platforms (Santhoshkumar et al. 2017). The salient features (optical, physical, and antimicrobial activities) of ZnO NPs are beneficial to plants (Tripathi et al. 2017) and can tackle environmental contaminants, for example, Cd (Rizwan et al. 2019a). Due to their tiny size and wider surface area, it is easy for ZnO NPs to penetrate contaminated plant sites and they show a high tendency to combat Cd (Khan et al. 2017). ZnO NPs reduced Cd accumulation in Sorghum bicolor (Wang et al. 2018) and in Leucaena leucocephala (Venkatachalam et al. 2017a). The effect of ZnO NPs application on the accumulation of Cd by pulse crops is crucial as this will provide useful information on the uptake in different parts of plants as well as their translocation from root to shoots. The use of ZnO NPs has not been properly examined but they have great potential as a viable option for agronomic biofortification of crops (Ghasemi et al. 2013).
The effect of Zn fertiliser applications on Cd accumulation in crops is well investigated particularly for cereals (Huang et al. 2019) but very limited studies have been done on pulse crops. Mung bean is an important pulse crop, and is beneficial for soil fertility. In addition to this, it is a good source of Zn, iron and vitamins and is very sensitive to Cd stress. How mung beans accumulate metals in their tissues and activate antioxidant enzymes (e.g. APX, CAT, POD, SOD) via interactions between ZnO NPs and Cd is not yet understood. The present study sets out to determine the Cd absorption, translocation, and growth performance in mung beans under various Cd and Zn interactions. To achieve these objectives, various plant parts, such as root, stem and leaves were evaluated for both Cd and Zn to determine the accumulation and translocation of these metals as well as activities of antioxidant enzymes in leaves.
Materials and methods
Plant growth conditions
One specific variety, Jade-AU – which is promoted by the Australian Mung Bean Association – was used in this study. It was characterised as fairly weather resistant with good lodgement resistance, tall, moderately susceptible to powdery mildew, tan spot and halo blight, with 100 seed weight 6–7.3 g. The seeds were surface sterilised with hydrogen peroxide 3% (vol/vol), and rinsed with Milli-Q water (18.2 MΩ cm). Seeds were placed in the 0.5 cm holes of the plastic lids and wrapped with cotton for germination. To facilitate germination and root growth to a certain length the lower end of the cotton was suspended in Milli-Q water in the pail. After 7 days of germination the pails were filled with a half-strength Hoagland’s nutrient solution [pH 5.6; composition (mg kg−1): N 210, K 235, Ca 200, P 31, S 64, Cl 0.65, Na 1.2, Mg 48.6, B 0.5, Fe 2.9, Mn 0.5, Zn 0.05, Cu 0.02, Mo 0.05] (Spomer et al. 1997) to facilitate growth and acclimatise the seedlings. Black polyethylene was used to cover the openings to stop light from reaching the nutrient solution and roots. Each pot contained three seedlings in the three holes. The solution was changed every 2 days. ZnO NPs and cadmium nitrate [Cd(NO3)2.4H2O] were added to the solution after the 7-day acclimation phase at the following concentrations: 0 (control), 1, 2, 4, 8, 16 and 32 μM Zn and 0 (control), 0.5 and 1 μM Cd, respectively. The experiment was set up using a complete randomised design with four replications. Mung beans were grown hydroponically in a greenhouse under the following conditions: 25/20°C day/night temperature, 75% relative humidity, and 16/8 h photoperiod.
Plant sampling and analysis
After 21 days, plants were removed from the solution, roots and shoots were separated using a stainless-steel cutter, washed with Milli-Q water and then placed in an oven at 72°C for 48 h. The dry biomass was recorded and the same samples were used for measurement of Zn and Cd. Plant samples were ground with a stainless-steel grinder to digest in a microwave digester (CEM, MARS 6). Approximately 0.25 g of ground samples was weighed directly into a Teflon vessel and 3 mL concentrated HNO3 and 2 mL H2O2 were added. Samples were pre-digested overnight (16 h) in a fume hood. Syringe filters (PTFE 0.45 μm) were employed to filter the samples and these were then stored at 4°C unless analysed on the day of preparation. The concentrations of Zn and Cd in the digested plant samples were measured by ICP-MS. Standard reference material (SRM) was used in each batch throughout the elemental analysis.
Determination of photosynthetic pigments
Chlorophyll a, b and total were measured utilising the Arnon (1949) method, which comprises a mixture of acetone and water at a ratio of 80%:20% (v/v). 2 g of mung bean leaves was homogenised with 25 mL of acetone solution (80%) and filtered. Absorption was measured at 663 and 645 nm using a UV spectrophotometer (Thermo Scientific-Biomate 3S). Using the following formulae the chlorophyll a, b and total were measured, where fw is fresh weight, A is absorption at a given wavelength, X is total volume of filtrate and n represents tissue weight:
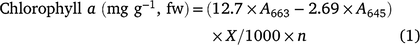
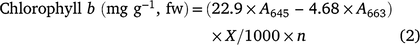
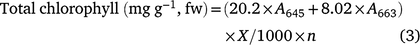
Antioxidant enzyme assay
Enzyme extracts were prepared according to Kamal and Komatsu (2015). Briefly, fresh leaves (approximately 250 mg) were homogenised with a pestle in an ice-cold mortar containing 1 mL of extraction buffer (25 mM potassium phosphate buffer [PPB, pH 7.8], 0.4 mM EDTA, 1 mM ascorbic acid, and 2% polyvinylpyrrolidone-40). The homogenates were centrifuged twice at 15 000g for 20 min at 4°C. The supernatants were collected and used for assays of enzymatic activity. Protein concentration was quantified according to the method of Bradford with bovine serum albumin as the standard. The enzyme activities were calculated according to the following equation (Borker et al. 2020):
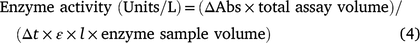
Where, Δt is the time of incubation (min), ΔAbs is the change in absorbance, ε is the extinction coefficient of substrates in units of M−1 cm−1), and l is the cuvette diameter (1 cm). Enzyme activity (Unit) was defined as the amount of enzyme that oxidised 1 μmol of substrate min−1. Finally we have converted the unit to unit (U) g−1 fw by calculation.
APX activity was determined in a 1 mL reaction mixture containing 25 mM PPB (pH 7.0), 0.25 mM ascorbic acid, 0.1 mM EDTA, 10 mM H2O2, and 0.1 mL of enzyme extract. The decline in ascorbic acid was monitored at 290 nm, and APX activity was calculated by using the extinction coefficient (EC290) of ascorbic acid (2.8 mM−1 cm−1).
CAT activity was determined in a reaction mixture containing 50 mM PPB (pH 7.0), 15 mM H2O2, and enzyme extract. The decomposition of H2O2 after 10 min of reaction time was measured at 240 nm in both the blanks and the samples by using a DU 730 UV-Vis spectrophotometer (Beckman Coulter, CA, USA), and CAT activity was calculated by using the EC240 of H2O2 (40 mM−1 cm−1).
POD activity was measured in a reaction mixture containing 100 mM PPB (pH 7.0), 2% H2O2, 4% p-phenylenediamine, 5 N H2SO4, and enzyme extract; the reaction was started by addition of the enzyme extract and stopped 10 min later by addition of 5 N H2SO4. In corresponding blanks, H2SO4 was added to the reaction mixture prior to the addition of enzyme extract. The tubes were kept in the refrigerator for 30 min at 4°C, and then the colour intensity was measured at 485 nm with the use of a UV-Vis spectrophotometer. POD activity was calculated by using the EC485 of p-phenylenediamine (2.8 mM−1 cm−1).
SOD activity was measured in a reaction mixture containing 200 mM PPB (pH 7.8), 250 μM nitro blue tetrazolium (NBT), 10 μM riboflavin, 0.1% tetramethylethylenediamine, and enzyme extract. With the use of a UV-Vis spectrophotometer, activity was calculated as the degree of inhibition of the photochemical reduction of NBT at 560 nm (EC560 = 2.1 mM−1 cm−1). The enzyme extract that corresponded to a 50% inhibition of the reaction was considered to be one enzyme unit.
Standard reference materials (SRMs)
To verify the results, spinach leaves SRM (1573a) from the National Institute of Standards and Technology (NIST) were used as standard reference materials (SRMs). Spinach leaves were digested following the same protocol as employed for the plant samples. The recovery percentages of the samples were 92% and 91% for Zn and Cd, respectively.
Statistical analysis
Data were analysed with the statistical package JMP Pro 14.2.0 software. Means were compared through the two-way ANOVA using Student’s t-test. Correlations between different traits were done using JMP Pro 14.2.0 software, and two significant levels of P < 0.05 and 0.01 were used in presenting the results. Figures were generated using GraphPad Prism Software (version 9.0.0). Computation of standard errors (s.e.) were done (n = 4) using Microsoft Excel.
Zn/Cd transfer/translocation factors (TFs) were calculated by the following formulae:

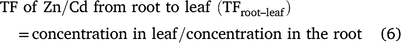

Results
Shoot and root dry matter yield
Shoot dry matter was significantly varied by Cd (P < 0.01) but non-significantly by ZnO NPs and their interactions (Fig. 1a). For instance, the reduction in shoot dry matter was about 20% (1.05 g) at lower Cd (0.5 μM) and 44% (0.73 g) at higher Cd (1.0 μM) levels as compared to the control (1.31 g). Although not significant, individually Zn increased shoot dry matter and reached the peak (1.68 g) at 16 μM ZnO NPs treatment, about 1.3 times greater than control (1.31 g). While Cd at 0.5 μM and 1.0 μM in solution exerted their detrimental effects through producing only 1.05 g and 0.73 g shoot dry matter, their interactions with ZnO NPs prevented the yield loss and yield in turn was boosted. Interaction of 8 μM ZnO NPs with lower and higher Cd levels increased dry matter to 1.53 g and 1.10 g, respectively, which was about 1.5 times larger than that of their sole Cd levels. However, the effect of elevated levels of ZnO NPs was less prominent at higher Cd (1.0 μM) than lower Cd (0.5 μM) concentration in solution.

|
Similar to shoot dry matter, root dry matter was also significantly (P < 0.01) influenced by Cd but not with ZnO NPs or their interactions. Root dry matter diminished to about 83% (0.235 g) and 77% (0.218 g) from the control (0.283 g) due to the presence of 0.5 μM and 1.0 μM Cd in solution, respectively (Fig. 1b). Sole application of ZnO NPs from 1.0 μM to 16 μM did not significantly increase root biomass, and this was followed by a slight decline at the highest level (32 μM). Although not significant, adding ZnO NPs to Cd-treated solution gradually enhanced root growth of mung beans and Zn at 8.0 μM yielded the highest amount.
Zn concentration in tissues
The concentration of Zn in leaf was significantly (P < 0.01) influenced due to ZnO NPs, Cd and their interactions (Fig. 2a–c). Without Cd added to the solution, leaf Zn concentration linearly increased as a result of elevated ZnO NPs levels. The highest Zn concentration in leaf (180 mg kg−1) was obtained at the highest Zn addition (32 μM) which was about 27 times more than the control (6.61 mg kg−1). However, the rate of increase in leaf Zn concentration was drastically reduced because of the interactions between ZnO NPs and Cd. When plants were grown in 0.5 μM Cd concentration, the addition of ZnO NPs in solution gradually increased leaf Zn concentration where the lower doses of ZnO NPs were not significantly affected. Interaction between 32 μM Zn and 0.5 μM Cd accumulated approximately five times the leaf Zn concentration (41.2 mg kg−1) than the sole Cd in solution (6.39 mg kg−1). Application of elevated ZnO NPs to a higher level of Cd (1.0 μM) also gradually increased leaf Zn concentration although this was non-significant.

|
Accumulation of Zn in the stem was significantly (P < 0.01) influenced by ZnO NPs, Cd and their interactions. The addition of individual ZnO NPs gradually enhanced stem Zn concentration and reached 60 mg kg−1 at 16 μM application which was approximately 10 times higher than the control. In comparison to the respective sole Zn and Cd treatments, their interactions exerted more influence on the accumulation of Zn in the stem. Concentration of Zn in the stem increased more than ten times with Zn at 32 μM coupled with either of the Cd levels (Fig. 2b).
Concentration of Zn in roots was significantly (P < 0.01) influenced by ZnO NPs, Cd and their interactions as well. Elevated levels of ZnO NPs linearly increased root Zn concentration and skyrocketed to 314 mg kg−1 at 16 μM Zn level which was approximately 33 times higher than the control (9.55 mg kg−1) (Fig. 2c). Slightly less accumulation of Zn in roots was observed in the case of ZnO NPs×Cd interaction as compared to sole ZnO NPs treatments. Interaction between the elevated Zn levels with either concentration of Cd contamination increased Zn accumulation in roots, where the highest figure was obtained with top levels of ZnO NPs and Cd.
Cd concentration in tissues
Cd accumulation in leaf was significantly (P < 0.01) affected by individual ZnO NPs and Cd but not by their interactions. Cadmium concentration at 0.5 and 1.0 μM increased in the leaf by more than six times (0.296 mg kg−1) and seven times (0.334 mg kg−1) compared to the control (0.047 mg kg−1) (Fig. 3a). However, no significant variation was observed in leaf Cd concentration between the two Cd levels. Application of ZnO NPs in solution containing 0.5 and 1.0 μM Cd did not restrict translocation of Cd to leaf. With the addition of up to 4 μM ZnO NPs, leaf Cd concentration linearly and significantly increased, and this was followed by a decline at further doses (Fig. 3a).

|
When compared to the control (0.123 mg kg−1) stem Cd concentration rose to 6.92 mg kg−1 (56 times) and 7.75 mg kg−1 (63 times) as a result of 0.5 μM Cd and 1.0 μM Cd, respectively, in solution (Fig. 3b). Interaction with Cd, ZnO NPs increased the stem Cd concentration. As compared to the accumulation due to sole Cd (0.5 μM and 1.0 μM), the presence of ZnO NPs with Cd displayed a 2–3 times higher accumulation in the stem. The highest level of ZnO NPs (32 μM) with either level of Cd accounted for higher concentrations of stem Cd (21 mg kg−1).
ZnO NPs and Cd individually as well as in combination significantly (P < 0.01) influenced Cd accumulation in the mung beans’ roots. Plants grown in non-spiked media (control) demonstrated only 0.957 mg Cd kg−1 in roots which sharply increased to 28 mg kg−1 and 83 mg kg−1 due to the addition of 0.5 and 1.0 μM Cd in solution, respectively (Fig. 3c). Like other tissues, Cd accumulation in roots was also enhanced by the addition of ZnO NPs to the solution containing Cd. This increase in Cd concentration was as high as 40.7–70 mg kg−1 and 90.4–136 mg kg−1 as a result of this addition, leading to increasing levels of Zn in lower (0.5 μM) and higher Cd-contaminated (1.0 μM) solutions.
Chlorophyll pigments in leaves
Both ZnO NPs and Cd displayed their significant (P < 0.01) effect on chlorophyll a contents in mung bean leaf but their interaction did not. Chlorophyll a concentration reduced to about 88% (0.718 mg kg−1, fw) and 79% (0.650 mg kg−1, fw) from the control (0.817 mg kg−1, fw.) due to the addition of 0.5 and 1.0 μM Cd in solution (Fig. 4a). On the other hand, individual application of Zn gradually boosted chlorophyll a contents and at 16 μM for the ZnO NPs level this content doubled. Chlorophyll a pigment was less prominent in the interactions between ZnO NPs and Cd in comparison to the main effect of ZnO NPs. When the solution contained either of Cd levels, chlorophyll a accumulation revealed an upward trend for 4 μM of ZnO NPs application.

|
Chlorophyll b was also significantly (P < 0.01) influenced by ZnO NPs and Cd, although their interaction effect was non-significant. Concentration of chlorophyll b fell by 9% (0.787 mg kg−1, fw) and 18% (0.713 mg kg−1, fw) from the control (0.865 mg kg−1, fw) as a result of adding Cd at 0.5 and 1.0 μM, respectively (Fig. 4b). Presumably, ZnO NPs enhanced chlorophyll b accumulation steadily at approximately an increase of 1.6 times (1.40 mg kg−1, fw) was observed at the treatment of 16 μM Zn than the control. However, the influence of elevated levels of ZnO NPs on chlorophyll b accumulation was restricted in the presence of Cd in solution. Adding more than 4 μM of ZnO NPs to either of the solutions containing Cd caused a gradual decline in chlorophyll b content.
A similar trend that was evident in chlorophyll a and b was observed for the total chlorophyll contents in mung bean leaves. Up to 15% reduction and a 41% increase in total chlorophyll concentration were observed due to the main effect of Cd and ZnO NPs, respectively (Fig. 4c). Combined presence of ZnO NPs and Cd in solution displayed larger and smaller accumulation compared to sole Cd and ZnO NPs, respectively. However, total chlorophyll contents were identical with the larger and smaller Cd levels when they reacted in combination with Zn.
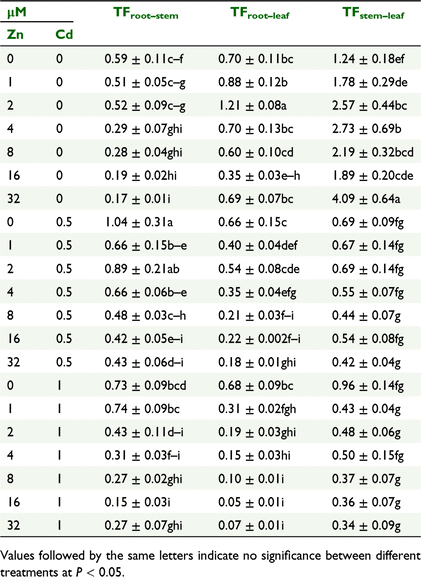
TFs of Zn within mung beans
The TFs of Zn within tissues, e.g. from root-to-stem (TFZnroot–stem), root-to-leaf (TFZnroot–leaf), and from stem-to-leaf (TFZnstem–leaf) are presented in Table 1. Added ZnO NPs decreased Zn transfer from root-to-stem (TFZnroot–stem) where no Cd was added, implying that Zn was mostly retained in roots and difficult to translocate to stem. With the exception of ZnO NPs application at 2 μM, translocation of Zn was almost steady from root-to-leaf (TFZnroot–leaf), which means Zn absorption by leaf mostly occurred from roots. In the Cd control translocation of Zn from stem-to-leaf (TFZnstem–leaf) was the highest, showing that Zn was not sequestrated to stem. The trend of Zn transfer from root-to-stem (TFZnroot–stem) was downward for both the main effect of ZnO NPs and its interaction with Cd. Due to the interaction between Zn and Cd in solution, a similar downward trend was observed for Zn translocation from root-to-leaf which indicates Zn was sequestrated to roots.
|
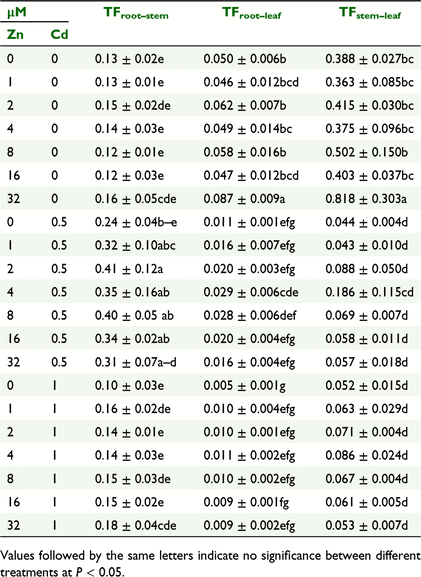
TFs of Cd within mung beans
The TFs of Cd from root-to-stem (TFCdroot–stem), root-to-leaf (TFCdroot–leaf), and from stem-to-leaf (TFCdstem–leaf) are presented in Table 2. The TFs of Cd within different parts of mung bean differed greatly. At Cd control the TFCdstem–leaf were about 3–5 times and 8–9 times higher than those for TFCdroot–leaf and TFCdroot–stem, respectively, suggesting a higher translocation of Cd from root-to-leaf. On the other hand, in presence of Cd, the highest translocation of Cd was observed from root-to-stem (TFCdroot–stem) which indicates the higher absorption capacity of Cd by stem than those for leaf. While added ZnO NPs gradually enhanced the absorption of Cd by leaf to 0.818 (at 32 μM Zn) from 0.388 (at control), its transfer was drastically restricted by added Cd to as low as 0.053 at same levels of added ZnO NPs, suggesting that the the additional Cd was sequestrated to stem. However, the translocation of Cd from root-to-leaf was always the lowest irrespective of ZnO NPs and Cd added in solution as compared to the TFCdroot–stem and TFCdstem–leaf, showing that it was very difficult for Cd to be transported from root-to-leaf and very easy to transport from stem (Table 2).
|
Correlation matrix
Shoot and root dry matter had a strong positive correlation with leaf Zn concentration and all the photosynthetic pigments (chlorophyll a, b and total chlorophyll) and strong negative correlations with Cd concentration of root, stem and leaf (Table 3). Similarly, root Zn concentration had strong positive correlations with Zn concentration of stem (r = 0.758**) and leaf (r = 0.595**) and positive correlations with all other parameters except total chlorophyll contents. Root Cd concentration displayed strong positive correlations with stem Cd (r = 0.701**) and leaf Cd (r = 0.667**) and strong negative correlations with dry matter yield (r = −0.512**, −0.529**), leaf Zn (r = −0.419**) and all the photosynthetic pigments. A strong positive correlation was observed for stem Zn concentration with root Zn, leaf Zn, stem and leaf Cd concentration and negative correlation with root dry matter and chlorophyll a. Cd concentration in stem showed strong positive correlation with root Cd, stem Zn and leaf Cd and strong negative correlation with root and shoot biomass production, leaf Zn and all the chlorophyll contents. Leaf Zn had strong positive correlation with root (r = 0.436**) and shoot dry matter (r = 0.251*), root Zn, stem Zn and all the photosynthetic pigments and strong negative correlations were observed with Cd concentration of root, stem and leaf. Cd concentration in leaf demonstrated strong positive correlations with Cd concentration of root and stem and strong negative correlations with root and shoot dry matter, leaf Zn and all the chlorophyll types. Chlorophyll a, chlorophyll b and total chlorophyll had strong positive correlations with root and shoot dry matter and strong negative correlations with Cd concentrations of root, stem and leaf (Table 3). Transfer of Zn from root-to-leaf (TFZnroot–leaf) was significantly negatively correlated with Cd concentration of root (r = −0.682**), stem (r = −0.667**) and leaf (r = −0.668**), suggesting that increase in the concentration of Cd in root, stem and leaf caused additional Zn to be sequestrated in the root, for that reason Zn treatment increased the concentration in the roots (Fig. 3c). An insignificant positive relationship (r = 0.118) was observed between root Zn concentration and TFCdroot–leaf, which indicates that the more accumulation of Zn in root, the less Cd was transferred to the leaf and more Cd was sequestrated to roots.

|
Effects of Cd and Zn on antioxidant enzyme activities
APX activity
Results revealed that APX activity was significantly (P < 0.01) increased by Zn supply (up to 4 μM Zn) and Cd addition (up to 1.0 μM Cd) in mung bean leaves (Fig. 5a). APX activity boosted up to 400.4 U g−1 fw from 353.2 U g−1 fw due to the presence of 1 μM Cd in solution which was about 13% higher than control. Similar increased APX activity (about 13% higher than control) was observed for the added Zn (4 μM) in solution. As compared to the individual effect of Zn and Cd, their interactions were responsible for higher increased APX activity (26% higher than control).

|
CAT activity
CAT activity was enhanced (P < 0.01) by increasing Zn (up to 16 μM Zn) and Cd concentration (up to 0.5 μM Cd). The least CAT activity was observed at control (41.75 U g−1 fw). At 0.5 μM Cd addition, CAT activity boosted up by about 19% (49.75 U g−1 fw) as compared to control (Fig. 5b). Increasing amount of Zn up to 16 μM was responsible for a gradual enhancement of CAT activity up to 60.00 U g−1 fw which was about 44% higher than control. Interaction between Zn and Cd further increased CAT activity by about 55% than control.
POD activity
Zn (up to 16 μM Zn) and Cd (up to 0.5 μM Zn) treatments were responsible (P < 0.01) for increasing POD activity (20.1 U g−1 fw) in a concentration-dependent manner in mung bean leaves. About 15% increased POD activity was observed due to 1 μM Cd addition as compared to control (17.5 U g−1 fw) (Fig. 5c). With the increasing of Zn supply, POD activity was also enhanced which reached to the peak (25.4 U g−1 fw) at 16 μM Zn application. As compared to the mean effect of Zn and Cd, their interactions induced more profound influence to enhance the POD activity. As compared to control, the highest increment (26.7 U g−1 fw) was observed at the interaction between 8 μM Zn and 1 μM Cd.
SOD activity
The induced activity of the SOD enzyme was noted (P < 0.01) in Zn (up to 16 μM Zn) and Cd-treated (up to 1.0 μM Zn) mung beans in a concentration-dependent manner. Addition of 0.5 μM and 1 μM Cd were responsible for the increment of about 16% (19.9 U g−1 fw) and 37% (23.6 U g−1 fw) increased SOD activity as compared to control (17.2 U g−1 fw) (Fig. 5d). Also the mean effect of Zn up to 16 μM increased SOD activity to 23.9 U g−1 fw from 17.2 U g−1 fw at control. The results revealed that Zn supply together with Cd accelerated SOD activity tremendously where 16 μM Zn × 1 μM Cd caused about 86% higher activity as compared to control.
Discussion
The objective of this trial was to investigate whether ZnO NPs can remediate the adverse effect of Cd in mung beans. Due to presence of Cd in solution, plants demonstrated a retarded growth (Fig. 1a and b). Studies have revealed that, as a toxic metal, Cd caused reduced growth and development (Bayçu et al. 2017). Our findings confirmed that ZnO NPs enhanced the growth and biomass yield of mung beans individually as well as by interactions with Cd (Fig. 1a and b). ZnO NPs at certain concentrations enhanced growth performance of crops (Faizan et al. 2018). ZnO NPs supply Zn2+ which enhance plant growth (Liu et al. 2015). The toxic effect of Cd might be neutralised by accelerated uptake of Zn and decreasing Cd uptake in plants treated with ZnO NPs (Garg and Kaur 2013). In their study, Venkatachalam et al. (2017a) observed superior growth of Leucaena leucocephala seedlings under Cd and lead (Pb) contamination as compared to the control as an effect of bioengineered ZnO NPs application. In another study with nanoscale zero-valent iron, an increase in dry matter yield of Boehimeria nivea was observed under Cd stress (Gong et al. 2017). Similarly, Singh and Lee (2016) found enhanced soybean growth utilising nano-TiO2 particles under Cd stress. Moreover, enhanced growth was observed by ZnO NPs in different crops (Rizwan et al. 2019a).
The present study revealed that concentration of chlorophylls linearly increased up to 4 μM of ZnO NPs application followed by a decline (Fig. 4a–c) and that improved plant growth was the result of enhanced accumulation of chlorophyll (Fig. 2a–c). Photosynthesis is one of many metabolic processes of plants which is very sensitive to environmental stresses (Khan et al. 2017). The prevalence of heavy metals, e.g. Cd is responsible for reduced photosynthetic activities and breakdown of chlorophyll pigments, for which plants show toxicity symptoms (Malar et al. 2016). Exogenous supply of ZnO NPs improved chlorophyll concentration in different plant species under metal stress condition (Venkatachalam et al. 2017a; Rizwan et al. 2019a). Enhanced photosynthetic activity caused by NPs is due to the acceleration of photolysis of water and the electron transport chain (Pradhan et al. 2013). Our findings are in agreement with those of previous studies (Venkatachalam et al. 2017a).
In our study the combined effects of ZnO NPs and Cd significantly increased Cd accumulation in tissues, although a decline in leaves was observed at certain levels (Fig. 3a–c). It is interesting that the root-to-stem, root-to-leaf, and stem-to-leaf TFs of mung bean varied greatly with Zn and Cd concentration in solution (Tables 1, 2). In this study TFs of Cd from root-to-stem and from root-to-leaf decreased and that of stem-to-leaf increased with added Cd (Table 2) which is in conformity with Khaliq et al. (2019) on rice. In this study added ZnO NPs favoured the upward movement of Cd (Table 2). However, Khaliq et al. (2019) found the opposite result in rice with soil application of zinc sulfate where root-to-shoot TFs of Cd was decreased by 28–41%. Root Zn concentration positively but non-significantly correlated with root Cd (r = 0.159), stem Cd (r = 0.148) and leaf Cd (r = 0.105) in our study and positively and significantly correlated with stem Zn (r = 0.758), and leaf Zn (r = 0.595). In a study with upland rice, Khaliq et al. (2019) obtained negative correlations between Zn concentrations and Cd concentrations in root, stem, leaf and brown rice. Zn may either positively or negatively correlate with Cd concentrations in plants (Green and Olsen 2017). Lasat et al. (2000) stated that ZNT1 is a Zn transporter and enhanced Cd in plants and Cohen et al. (1998) found that IRT1 might facilitate the transport of divalent cations such as Zn2+, Fe2+ and Cd2+.
It appears that Cd concentration was larger in roots than any other parts (Fig. 3c), and this may be one of the plant’s protective mechanisms and also a consequence of direct contact of root with Cd in the solution (Rizwan et al. 2018). The influence of NPs on metal uptake by plants is not consistent in the literature (Singh and Lee 2016). Some literature supports our results, for example: in L. leucocephala plant by ZnO NPs (Venkatachalam et al. 2017b); in B. nivea by nano zero-valent Fe (Gong et al. 2017); and in soybean using TiO2 NPs (Singh and Lee 2016). However, other research came to very different conclusions (Qaswar et al. 2017). Consequently, it can be stated that the interaction of metals with NPs is influenced by the type of NPs, plants, conditions of growth, etc. A sharp increase in the accumulation of Zn in tissues was observed due to the application of ZnO NPs (Fig. 2a–c), and this has been confirmed in one analysis on green pea and beet (García-Gómez et al. 2018). Plants experience a shortfall in essential nutrients and particularly Zn due to competition with Cd under Cd stress (Rizwan et al. 2016). According to our results, the influence of ZnO NPs on Cd uptake depends on concentration levels and this corroborates other studies confirming that higher Zn partially inhibits Cd toxicity for some mechanisms. These include, for example, co-precipitation of Cd and Zn in metabolically inactive parts (Rizwan et al. 2016). Expectedly, ZnO NPs may penetrate from roots or leaf surfaces and could transfer to other parts of plants, but more detailed studies are necessary to measure the Zn speciation and biotransformation of ZnO NPs in plants as well as underlying mechanisms. Generally in an oxidising environment plants continuously produce ROS [super oxide (O2−), singlet oxygen (1O2), hydroxyl radical (HO−), and H2O2] in cells as by products. An extensive production of ROS causes oxidative stress which leads to damage to DNA, lipid and proteins and causes cell death (Tripathy and Oelmüller 2012). As a defence mechanism plants activate enzymatic [APX, CAT, POD, SOD, and glutathione reductase (GR) etc.] and non-enzymatic (proline) antioxidants (Tripathy and Oelmüller 2012). In this study, application of ZnO NPs to solution enhanced antioxidant enzyme activity under Cd stress (Fig. 5a–d). The APX activity in our study ranged from 323.0 to 456.0 U g−1 fw where the presence of 0.5 μM and 1.0 μM Cd were responsible for about 6 and 13% enhancement over control. Similar results (about 250–400 U g−1 fw) were found in mung bean leaves with an exception of a negligible decline due to Cd toxicity (Leng et al. 2021). However, as compared to control, an enhanced activity of APX enzyme due to Cd toxicity was observed in mung bean seedlings which also supports our results (Hassan et al. 2021). About 19% increased (41.0–67.2 U g−1 fw) CAT activity due to Cd stress was observed in this study which was about 64% higher in mung bean seedlings (Leng et al. 2021) as compared to control. Likewise, CAT activity in mung beans ranged from 17.5 to 18.2 μmoles min−1 mg−1 protein (Farahani and Taghavi 2016). With increasing ZnO NPs and Cd up to a certain level, POD and SOD activities increased in our study which were from 16.6 to 29.7 U g−1 fw and 16.0 to 37.0 U g−1 fw, respectively. In literature, the POD activity in mung beans ranged from 30.4 to 32.6 μmol min−1 mg−1 protein (Farahani and Taghavi 2016). As compared to the uncontaminated control about 138–139% increased SOD activity was observed in mung beans due to Cd contamination (Ramzani et al. 2017). However, under Cd treatments, Hassan et al. (2021) found an enhanced SOD activity while a slight decreased activity was observed by Leng et al. (2021). In this study, activities of all the antioxidant enzymes were increased with ZnO NPs application under Cd stress. Improved antioxidant enzyme activities by ZnO NPs under Cd stress were documented in maize (Rizwan et al. 2019a) and in tomato (Faizan et al. 2021). As a source of Zn, application of ZnCl2 to solution also improved SOD, CAT, APX, and GR activity in tomato under Cd stress, indicating the efficacy of Zn supply to boost these antioxidant enzyme activities as compared to no Zn application (Cherif et al. 2011). An improvement in the antioxidant defence system due to ZnO NPs application was also investigated on different crops (Rizwan et al. 2019a). Such development of antioxidant defence system within plants is the outcome of the expression of genes (Nair and Chung 2014) or its association in different oxidative processes (Hossain et al. 2015). Overall, under Cd contamination, the application of ZnO NPs could be an effective tool to strengthen plants’ defence mechanisms, maintain yield performance and Cd tolerance.
Conclusion
ZnO NPs up to 8 μM level promoted growth performance in both control and Cd-stressed conditions, and thus prevented the adverse effect of Cd to some extent. Added ZnO NPs to Cd-treated solution favoured Cd accumulation consistently in all tissues except leaf. Up to 4 μM ZnO NPs application, Cd in leaf increased followed by a decrease at higher levels. Cd contamination limits accumulation of Zn in mung bean leaves to a great extent as compared to the accumulation in roots and stems, indicating that Zn transfer to leaves from roots and stems was impaired by prevalence of Cd in the media. ZnO NPs demonstrate a synergistic effect on Cd accumulation in mung bean tissues especially in the roots and stems, which is contrast to the antagonistic effects documented in some literature. Despite the higher accumulation of Cd in tissues, plant growth displayed an upward trend and this supports the hypothesis that Zn plays a key role in protecting plants from Cd stress. It does this through activation of robust antioxidant enzyme systems against Cd-induced oxidative stress. Further research on this topic at the field levels may help to better understand the influence of ZnO NPs along with their application protocols in diverse soil conditions. The emphasis must be on physiological mechanisms, metal accumulation and translocation to the edible portions of plants.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare that they have no known conflicts of interest.
Declaration of funding
This research was partly funded by the CRC CARE and the IZA (G2000677).
Acknowledgements
The authors are grateful to the University of Newcastle (UoN) for providing the necessary facilities. Also, we acknowledge the contribution of CRC CARE and the International Zinc Association (IZA) for their financial support.
References
Arnon DI (1949) Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiology 24, 1| Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris.Crossref | GoogleScholarGoogle Scholar | 16654194PubMed |
Bayçu G, Gevrek-Kürüm N, Moustaka J, Csatári I, Rognes SE, Moustakas M (2017) Cadmium–zinc accumulation and photosystem II responses of Noccaea caerulescens to Cd and Zn exposure. Environmental Science and Pollution Research 24, 2840–2850.
| Cadmium–zinc accumulation and photosystem II responses of Noccaea caerulescens to Cd and Zn exposure.Crossref | GoogleScholarGoogle Scholar | 27838905PubMed |
Borker AR, David K, Singhal N (2020) Analysis of time varying response on uptake patterns of Cu and Zn ions under application of ethylene diamine disuccinic acid and gibberellic acid in Lolium perenne. Chemosphere 260, 127541
| Analysis of time varying response on uptake patterns of Cu and Zn ions under application of ethylene diamine disuccinic acid and gibberellic acid in Lolium perenne.Crossref | GoogleScholarGoogle Scholar | 32688311PubMed |
Cherif J, Mediouni C, Ammar WB, Jemal F (2011) Interactions of zinc and cadmium toxicity in their effects on growth and in antioxidative systems in tomato plants (Solarium lycopersicum). Journal of Environmental Sciences 23, 837–844.
| Interactions of zinc and cadmium toxicity in their effects on growth and in antioxidative systems in tomato plants (Solarium lycopersicum).Crossref | GoogleScholarGoogle Scholar |
Cohen CK, Fox TC, Garvin DF, Kochian LV (1998) The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants. Plant Physiology 116, 1063–1072.
| The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants.Crossref | GoogleScholarGoogle Scholar | 9501139PubMed |
Cojocaru P, Gusiatin ZM, Cretescu I (2016) Phytoextraction of Cd and Zn as single or mixed pollutants from soil by rape (Brassica napus). Environmental Science and Pollution Research 23, 10693–10701.
| Phytoextraction of Cd and Zn as single or mixed pollutants from soil by rape (Brassica napus).Crossref | GoogleScholarGoogle Scholar | 26884243PubMed |
Faizan M, Faraz A, Yusuf M, Khan ST, Hayat S (2018) Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 56, 678–686.
| Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants.Crossref | GoogleScholarGoogle Scholar |
Faizan M, Faraz A, Mir AR, Hayat S (2021) Role of zinc oxide nanoparticles in countering negative effects generated by cadmium in Lycopersicon esculentum. Journal of Plant Growth Regulation 40, 101–115.
| Role of zinc oxide nanoparticles in countering negative effects generated by cadmium in Lycopersicon esculentum.Crossref | GoogleScholarGoogle Scholar |
Farahani AS, Taghavi M (2016) Changes of antioxidant enzymes of mung bean [Vigna radiata (L.) R. Wilczek] in response to host and non-host bacterial pathogens. Journal of Plant Protection Research 56, 95–99.
| Changes of antioxidant enzymes of mung bean [Vigna radiata (L.) R. Wilczek] in response to host and non-host bacterial pathogens.Crossref | GoogleScholarGoogle Scholar |
Forster SM, Rickertsen JR, Mehring GH, Ransom JK (2018) Type and placement of zinc fertilizer impacts cadmium content of harvested durum wheat grain. Journal of Plant Nutrition 41, 1471–1481.
| Type and placement of zinc fertilizer impacts cadmium content of harvested durum wheat grain.Crossref | GoogleScholarGoogle Scholar |
García-Gómez C, García S, Obrador AF, González D, Babín M, Fernández MD (2018) Effects of aged ZnO NPs and soil type on Zn availability, accumulation and toxicity to pea and beet in a greenhouse experiment. Ecotoxicology and Environmental Safety 160, 222–230.
| Effects of aged ZnO NPs and soil type on Zn availability, accumulation and toxicity to pea and beet in a greenhouse experiment.Crossref | GoogleScholarGoogle Scholar | 29807295PubMed |
Garg N, Kaur H (2013) Response of antioxidant enzymes, phytochelatins and glutathione production towards Cd and Zn stresses in Cajanus cajan (L.) Millsp. genotypes colonized by arbuscular mycorrhizal fungi. Journal of Agronomy and Crop Science 199, 118–133.
| Response of antioxidant enzymes, phytochelatins and glutathione production towards Cd and Zn stresses in Cajanus cajan (L.) Millsp. genotypes colonized by arbuscular mycorrhizal fungi.Crossref | GoogleScholarGoogle Scholar |
Ghasemi S, Khoshgoftarmanesh AH, Afyuni M, Hadadzadeh H (2013) The effectiveness of foliar applications of synthesized zinc-amino acid chelates in comparison with zinc sulfate to increase yield and grain nutritional quality of wheat. European Journal of Agronomy 45, 68–74.
| The effectiveness of foliar applications of synthesized zinc-amino acid chelates in comparison with zinc sulfate to increase yield and grain nutritional quality of wheat.Crossref | GoogleScholarGoogle Scholar |
Gong X, Huang D, Liu Y, Zeng G, Wang R, Wan J, Zhang C, Cheng M, Qin X, Xue W (2017) Stabilized nanoscale zerovalent iron mediated cadmium accumulation and oxidative damage of Boehmeria nivea (L.) Gaudich cultivated in cadmium contaminated sediments. Environmental Science & Technology 51, 11308–11316.
| Stabilized nanoscale zerovalent iron mediated cadmium accumulation and oxidative damage of Boehmeria nivea (L.) Gaudich cultivated in cadmium contaminated sediments.Crossref | GoogleScholarGoogle Scholar |
Green AC, Olsen CM (2017) Cutaneous squamous cell carcinoma: an epidemiological review. British Journal of Dermatology 177, 373–381.
| Cutaneous squamous cell carcinoma: an epidemiological review.Crossref | GoogleScholarGoogle Scholar |
Hassan M, Israr M, Mansoor S, Hussain SA, Basheer F, Azizullah A, Ur Rehman S (2021) Acclimation of cadmium-induced genotoxicity and oxidative stress in mung bean seedlings by priming effect of phytohormones and proline. PLoS ONE 16, e0257924
| Acclimation of cadmium-induced genotoxicity and oxidative stress in mung bean seedlings by priming effect of phytohormones and proline.Crossref | GoogleScholarGoogle Scholar | 34587203PubMed |
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments: a review. Plant Signaling & Behavior 7, 1456–1466.
| Role of proline under changing environments: a review.Crossref | GoogleScholarGoogle Scholar |
Hossain MA, Bhattacharjee S, Armin S-M, Qian P, Xin W, Li H-Y, Burritt DJ, Fujita M, Tran L-SP (2015) Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: insights from ROS detoxification and scavenging. Frontiers in Plant Science 6, 420
| Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: insights from ROS detoxification and scavenging.Crossref | GoogleScholarGoogle Scholar | 26136756PubMed |
Huang G, Ding C, Zhou Z, Zhang T, Wang X (2019) A tillering application of zinc fertilizer based on basal stabilization reduces Cd accumulation in rice (Oryza sativa L.). Ecotoxicology and Environmental Safety 167, 338–344.
| A tillering application of zinc fertilizer based on basal stabilization reduces Cd accumulation in rice (Oryza sativa L.).Crossref | GoogleScholarGoogle Scholar | 30359900PubMed |
IARC (2018) List of classifications by cancer sites with sufficient or limited evidence in humans, volumes 1 to 121 [WWW document]. International Agency for Research on Cancer. Available at http://monographs.iarc.fr/ENG/Classification/Table4.pdf. [Accessed 27 June 2018]
Kamal AHM, Komatsu S (2015) Involvement of reactive oxygen species and mitochondrial proteins in biophoton emission in roots of soybean plants under flooding stress. Journal of Proteome Research 14, 2219–2236.
| Involvement of reactive oxygen species and mitochondrial proteins in biophoton emission in roots of soybean plants under flooding stress.Crossref | GoogleScholarGoogle Scholar |
Khaliq MA, James B, Chen YH, Saqib HSA, Li HH, Jayasuriya P, Guo W (2019) Uptake, translocation, and accumulation of Cd and its interaction with mineral nutrients (Fe, Zn, Ni, Ca, Mg) in upland rice. Chemosphere 215, 916–924.
| Uptake, translocation, and accumulation of Cd and its interaction with mineral nutrients (Fe, Zn, Ni, Ca, Mg) in upland rice.Crossref | GoogleScholarGoogle Scholar | 30408887PubMed |
Khan MN, Mobin M, Abbas ZK, AlMutairi KA, Siddiqui ZH (2017) Role of nanomaterials in plants under challenging environments. Plant Physiology and Biochemistry 110, 194–209.
| Role of nanomaterials in plants under challenging environments.Crossref | GoogleScholarGoogle Scholar | 27269705PubMed |
Köleli N, Eker S, Cakmak I (2004) Effect of zinc fertilization on cadmium toxicity in durum and bread wheat grown in zinc-deficient soil. Environmental Pollution 131, 453–459.
| Effect of zinc fertilization on cadmium toxicity in durum and bread wheat grown in zinc-deficient soil.Crossref | GoogleScholarGoogle Scholar | 15261409PubMed |
Lasat MM, Pence NS, Garvin DF, Ebbs SD, Kochian LV (2000) Molecular physiology of zinc transport in the Zn hyperaccumulator Thlaspi caerulescens. Journal of Experimental Botany 51, 71–79.
| Molecular physiology of zinc transport in the Zn hyperaccumulator Thlaspi caerulescens.Crossref | GoogleScholarGoogle Scholar | 10938797PubMed |
Leng Y, Li Y, Ma Y-H, He L-F, Li S-W (2021) Abscisic acid modulates differential physiological and biochemical responses of roots, stems, and leaves in mung bean seedlings to cadmium stress. Environmental Science and Pollution Research 28, 6030–6043.
| Abscisic acid modulates differential physiological and biochemical responses of roots, stems, and leaves in mung bean seedlings to cadmium stress.Crossref | GoogleScholarGoogle Scholar | 32986195PubMed |
Liu X, Wang F, Shi Z, Tong R, Shi X (2015) Bioavailability of Zn in ZnO nanoparticle-spiked soil and the implications to maize plants. Journal of Nanoparticle Research 17, 175
| Bioavailability of Zn in ZnO nanoparticle-spiked soil and the implications to maize plants.Crossref | GoogleScholarGoogle Scholar |
Malar S, Vikram SS, Favas PJC, Perumal V (2016) Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Botanical Studies 55, 54
| Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)].Crossref | GoogleScholarGoogle Scholar | 28597420PubMed |
Martos S, Gallego B, Sáez L, López-Alvarado J, Cabot C, Poschenrieder C (2016) Characterization of zinc and cadmium hyperaccumulation in three Noccaea (Brassicaceae) populations from non-metalliferous sites in the eastern Pyrenees. Frontiers in Plant Science 7, 128
| Characterization of zinc and cadmium hyperaccumulation in three Noccaea (Brassicaceae) populations from non-metalliferous sites in the eastern Pyrenees.Crossref | GoogleScholarGoogle Scholar | 26904085PubMed |
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science 7, 405–410.
| Oxidative stress, antioxidants and stress tolerance.Crossref | GoogleScholarGoogle Scholar | 12234732PubMed |
Mohammad A, Moheman A (2010) The effects of cadmium and zinc interactions on the accumulation and tissue distribution of cadmium and zinc in tomato (Lycopersicon esculentum Mill.). Archives of Agronomy and Soil Science 56, 551–561.
| The effects of cadmium and zinc interactions on the accumulation and tissue distribution of cadmium and zinc in tomato (Lycopersicon esculentum Mill.).Crossref | GoogleScholarGoogle Scholar |
Murtaza G, Javed W, Hussain A, Qadir M, Aslam M (2017) Soil-applied zinc and copper suppress cadmium uptake and improve the performance of cereals and legumes. International Journal of Phytoremediation 19, 199–206.
| Soil-applied zinc and copper suppress cadmium uptake and improve the performance of cereals and legumes.Crossref | GoogleScholarGoogle Scholar | 27419530PubMed |
Nair PMG, Chung IM (2014) Assessment of silver nanoparticle-induced physiological and molecular changes in Arabidopsisthaliana. Environmental Science and Pollution Research 21, 8858–8869.
| Assessment of silver nanoparticle-induced physiological and molecular changes in Arabidopsisthaliana.Crossref | GoogleScholarGoogle Scholar |
Pradhan S, Patra P, Das S, Chandra S, Mitra S, Dey KK, Akbar S, Palit P, Goswami A (2013) Photochemical modulation of biosafe manganese nanoparticles on Vigna radiata: a detailed molecular, biochemical, and biophysical study. Environmental Science & Technology 47, 13122–13131.
| Photochemical modulation of biosafe manganese nanoparticles on Vigna radiata: a detailed molecular, biochemical, and biophysical study.Crossref | GoogleScholarGoogle Scholar |
Qaswar M, Hussain S, Rengel Z (2017) Zinc fertilisation increases grain zinc and reduces grain lead and cadmium concentrations more in zinc-biofortified than standard wheat cultivar. Science of The Total Environment 605, 454–460.
| Zinc fertilisation increases grain zinc and reduces grain lead and cadmium concentrations more in zinc-biofortified than standard wheat cultivar.Crossref | GoogleScholarGoogle Scholar |
Ramzani PMA, Coyne MS, Anjum S, Khan W-U-D, Iqbal M (2017) In situ immobilization of Cd by organic amendments and their effect on antioxidant enzyme defense mechanism in mung bean (Vigna radiata L.) seedlings. Plant Physiology and Biochemistry 118, 561–570.
| In situ immobilization of Cd by organic amendments and their effect on antioxidant enzyme defense mechanism in mung bean (Vigna radiata L.) seedlings.Crossref | GoogleScholarGoogle Scholar | 28783510PubMed |
Rizwan M, Ali S, Abbas T, Zia-ur-Rehman M, Hannan F, Keller C, Al-Wabel MI, Ok YS (2016) Cadmium minimization in wheat: a critical review. Ecotoxicology and Environmental Safety 130, 43–53.
| Cadmium minimization in wheat: a critical review.Crossref | GoogleScholarGoogle Scholar | 27062345PubMed |
Rizwan M, Ali S, Zia-ur-Rehman MZ, Rinklebe J, Tsang DC, Bashir A, Maqbool A, Tack F, Ok YS (2018) Cadmium phytoremediation potential of Brassica crop species: a review. Science of The Total Environment 631, 1175–1191.
| Cadmium phytoremediation potential of Brassica crop species: a review.Crossref | GoogleScholarGoogle Scholar |
Rizwan M, Ali S, Zia-ur-Rehman MZ, Adrees M, Arshad M, Qayyum MF, Ali L, Hussain A, Chatha SAS, Imran M (2019a) Alleviation of cadmium accumulation in maize (Zea mays L.) by foliar spray of zinc oxide nanoparticles and biochar to contaminated soil. Environmental Pollution 248, 358–367.
| Alleviation of cadmium accumulation in maize (Zea mays L.) by foliar spray of zinc oxide nanoparticles and biochar to contaminated soil.Crossref | GoogleScholarGoogle Scholar | 30818115PubMed |
Rizwan M, Ali S, Zia-ur-Rehman MZ, Maqbool A (2019b) A critical review on the effects of zinc at toxic levels of cadmium in plants. Environmental Science and Pollution Research 26, 6279–6289.
| A critical review on the effects of zinc at toxic levels of cadmium in plants.Crossref | GoogleScholarGoogle Scholar | 30635881PubMed |
Saifullah Saifullah (2014) Effectiveness of zinc application to minimize cadmium toxicity and accumulation in wheat (Triticum aestivum L.). Environmental Earth Sciences 71, 1663–1672.
| Effectiveness of zinc application to minimize cadmium toxicity and accumulation in wheat (Triticum aestivum L.).Crossref | GoogleScholarGoogle Scholar |
Santhoshkumar J, Kumar SV, Rajeshkumar S (2017) Synthesis of zinc oxide nanoparticles using plant leaf extract against urinary tract infection pathogen. Resource-Efficient Technologies 3, 459–465.
| Synthesis of zinc oxide nanoparticles using plant leaf extract against urinary tract infection pathogen.Crossref | GoogleScholarGoogle Scholar |
Singh J, Lee B-K (2016) Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max): a possible mechanism for the removal of Cd from the contaminated soil. Journal of Environmental Management 170, 88–96.
| Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max): a possible mechanism for the removal of Cd from the contaminated soil.Crossref | GoogleScholarGoogle Scholar | 26803259PubMed |
Spomer LA, Berry WL, Tibbitts TW (1997) Plant culture in solid media. In ‘Plant growth chamber handbook’. (Eds RW Langhans, TW Tibbitts) pp. 105–118. (Iowa Agricultural and Home Economics Experiment Station: Iowa, IA, USA)
Tripathi DK, Singh S, Singh S, Pandey R, Singh VP, Sharma NC, Prasad SM, Dubey NK, Chauhan DK (2017) An overview on manufactured nanoparticles in plants: uptake, translocation, accumulation and phytotoxicity. Plant Physiology and Biochemistry 110, 2–12.
| An overview on manufactured nanoparticles in plants: uptake, translocation, accumulation and phytotoxicity.Crossref | GoogleScholarGoogle Scholar | 27601425PubMed |
Tripathy BC, Oelmüller R (2012) Reactive oxygen species generation and signaling in plants. Plant Signaling & Behavior 7, 1621–1633.
| Reactive oxygen species generation and signaling in plants.Crossref | GoogleScholarGoogle Scholar |
Venkatachalam P, Jayaraj M, Manikandan R, Geetha N, Rene ER, Sharma NC, Sahi SV (2017a) Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: a physiochemical analysis. Plant Physiology and Biochemistry 110, 59–69.
| Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: a physiochemical analysis.Crossref | GoogleScholarGoogle Scholar | 27622846PubMed |
Venkatachalam P, Priyanka N, Manikandan K, Ganeshbabu I, Indiraarulselvi P, Geetha N, Muralikrishna K, Bhattacharya RC, Tiwari M, Sharma N, Sahi SV (2017b) Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.). Plant Physiology and Biochemistry 110, 118–127.
| Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.).Crossref | GoogleScholarGoogle Scholar | 27622847PubMed |
Wang F, Jing X, Adams CA, Shi Z, Sun Y (2018) Decreased ZnO nanoparticle phytotoxicity to maize by arbuscular mycorrhizal fungus and organic phosphorus. Environmental Science and Pollution Research 25, 23736–23747.
| Decreased ZnO nanoparticle phytotoxicity to maize by arbuscular mycorrhizal fungus and organic phosphorus.Crossref | GoogleScholarGoogle Scholar | 29876848PubMed |
Zare AA, Khoshgoftarmanesh AH, Malakouti MJ, Bahrami HA, Chaney RL (2018) Root uptake and shoot accumulation of cadmium by lettuce at various Cd:Zn ratios in nutrient solution. Ecotoxicology and Environmental Safety 148, 441–446.
| Root uptake and shoot accumulation of cadmium by lettuce at various Cd:Zn ratios in nutrient solution.Crossref | GoogleScholarGoogle Scholar | 29102904PubMed |